Identification and Bioinformatic Analysis of the GmDOG1-Like Family in Soybean and Investigation of Their Expression in Response to Gibberellic Acid and Abscisic Acid

 ,
, 

Abstract
1. Introduction
2. Results
2.1. Identification of GmDOG1Ls Genes
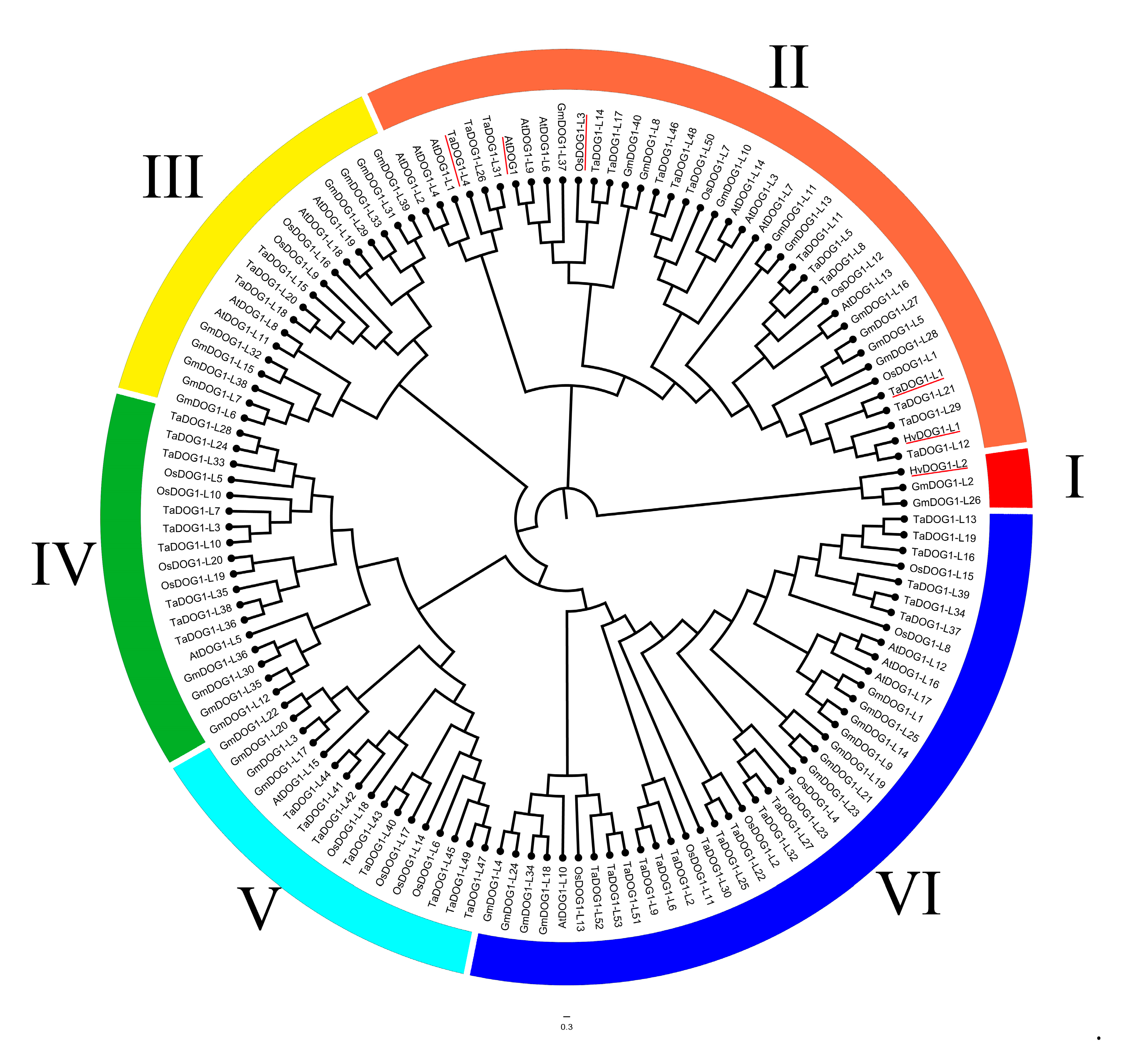
2.2. GmDOG1L Genes Structure and Phylogenetic Analysis
2.3. Chromosomal Location and Gene Duplication
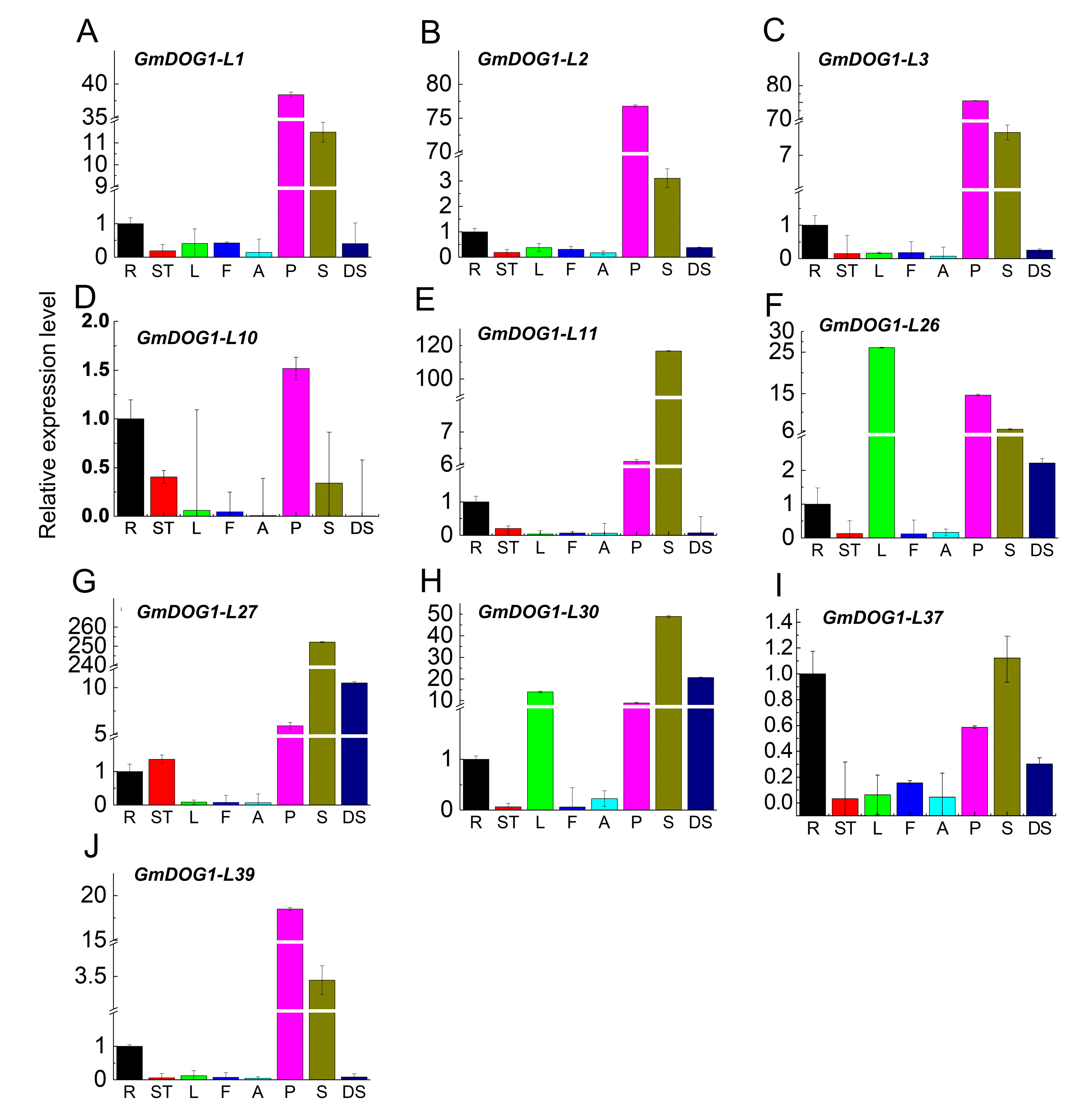
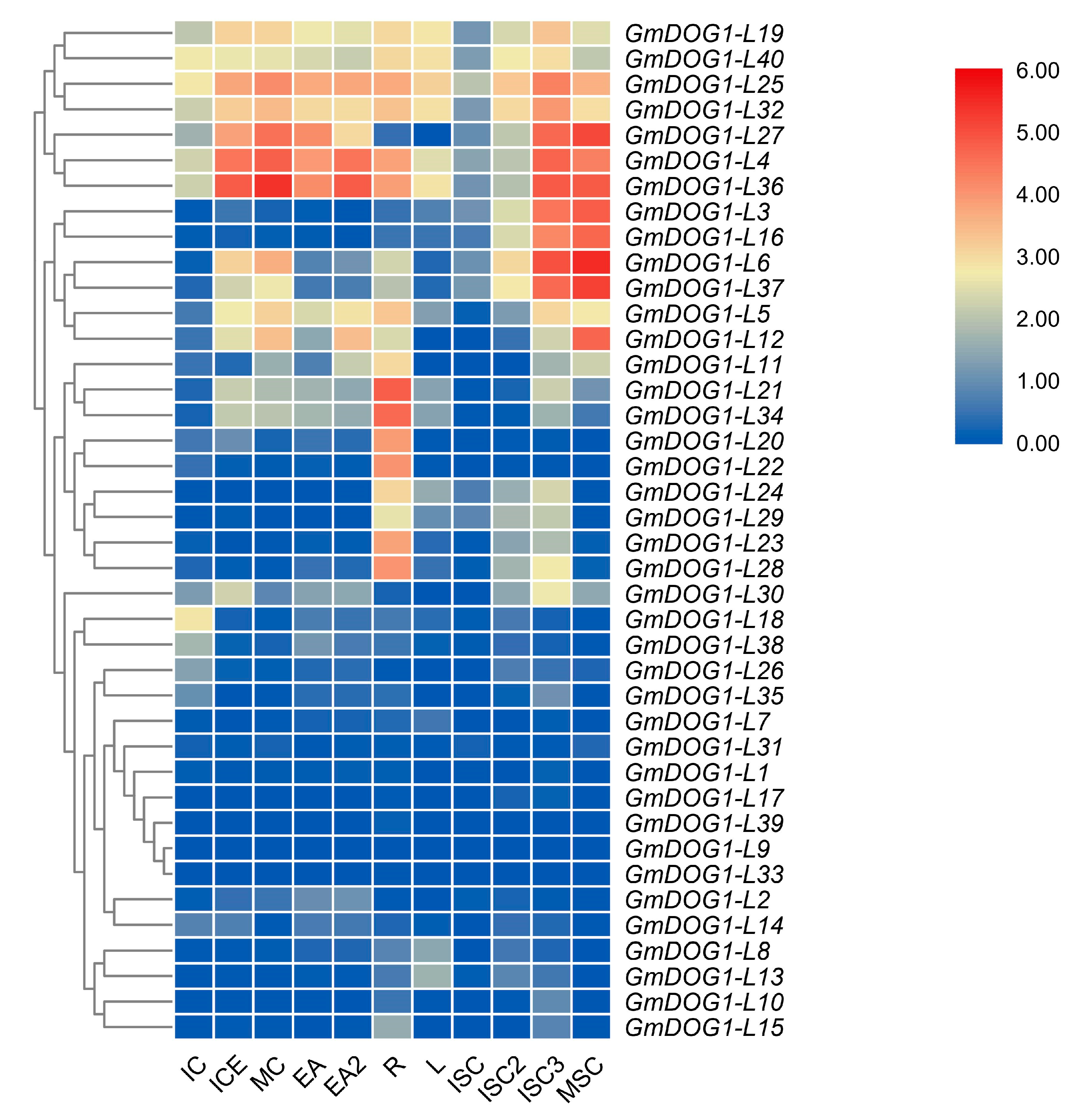
2.4. Expression Profiles of GmDOG1L Genes in Various Tissues of Soybean
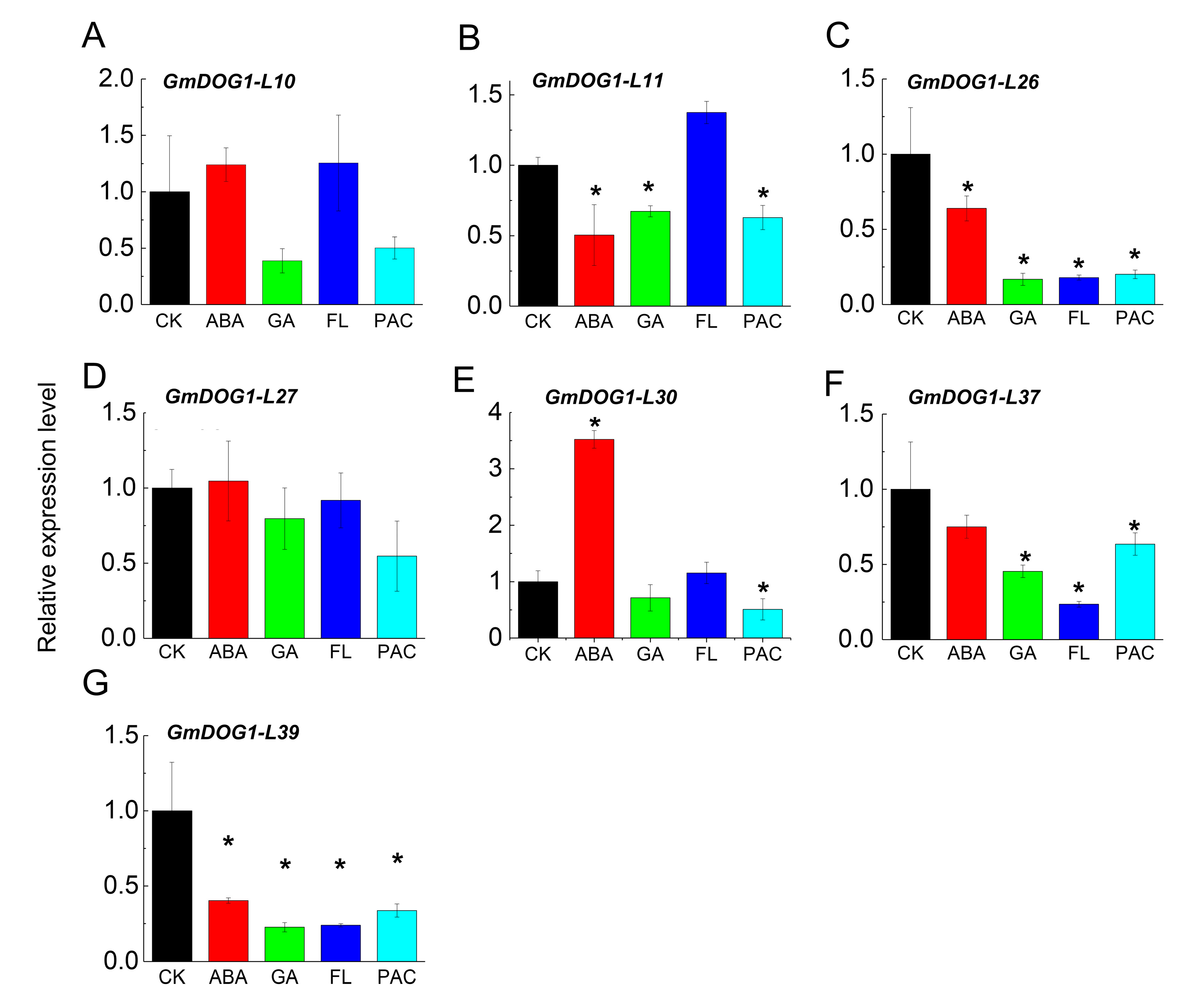
2.5. Expression Analysis of GmDOG1L under Phytohormones Treatment
3. Discussions
4. Conclusions
5. Materials and Methods
5.1. Identification of GmDOG1Ls
5.2. Chromosomal Distribution, Gene Structure and Phylogenetic Analysis
5.3. Synteny Analysis of GmDOG1Ls
5.4. Gene Expression Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lee, G.-A.; Crawford, G.W.; Liu, L.; Sasaki, Y.; Chen, X. Archaeological Soybean (Glycine max) in East Asia: Does Size Matter? PLoS ONE 2011, 6, e26720. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Chen, F.; Shuai, H.; Luo, X.; Ding, J.; Tang, S.; Xu, S.; Liu, J.; Liu, W.; Du, J.; et al. Karrikins delay soybean seed germination by mediating abscisic acid and gibberellin biogenesis under shaded conditions. Sci. Rep. 2016, 6, 22073. [Google Scholar] [CrossRef]
- Zhou, W.; Chen, F.; Zhao, S.; Yang, C.; Meng, Y.; Shuai, H.; Luo, X.; Dai, Y.; Yin, H.; Du, J.; et al. DA-6 promotes germination and seedling establishment from aged soybean seeds by mediating fatty acid metabolism and glycometabolism. J. Exp. Bot. 2018, 70, 101–114. [Google Scholar] [CrossRef]
- Meng, Y.; Shuai, H.; Luo, X.; Chen, F.; Zhou, W.; Yang, W.; Shu, K. Karrikins: Regulators Involved in Phytohormone Signaling Networks during Seed Germination and Seedling Development. Front. Plant Sci. 2017, 7, e0157. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Chen, F.; Luo, X.; Dai, Y.; Yang, Y.; Zheng, C.; Yang, W.; Shu, K. A matter of life and death: Molecular, physiological, and environmental regulation of seed longevity. Plant Cell Environ. 2019, 43, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Zhou, W.; Chen, F.; Luo, X.; Yang, W. Abscisic Acid and Gibberellins Antagonistically Mediate Plant Development and Abiotic Stress Responses. Front. Plant Sci. 2018, 9, 416. [Google Scholar] [CrossRef] [PubMed]
- Bentsink, L.; Jowett, J.; Hanhart, C.J.; Koornneef, M. Cloning of DOG1, a quantitative trait locus controlling seed dormancy in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 17042–17047. [Google Scholar] [CrossRef]
- Nakabayashi, K.; Bartsch, M.; Xiang, Y.; Miatton, E.; Pellengahr, S.; Yano, R.; Seo, M.; Soppe, W.J.J. The Time Required for Dormancy Release in Arabidopsis Is Determined by Delay of Germination1 Protein Levels in Freshly Harvested Seeds. Plant Cell 2012, 24, 2826–2838. [Google Scholar] [CrossRef]
- Footitt, S.; Huang, Z.; Clay, H.A.; Mead, A.; Finch-Savage, W.E. Temperature, light and nitrate sensing coordinate Arabidopsis seed dormancy cycling, resulting in winter and summer annual phenotypes. Plant J. 2013, 74, 1003–1015. [Google Scholar] [CrossRef]
- Huo, H.; Wei, S.; Bradford, K.J. Delay of Germination1(DOG1) regulates both seed dormancy and flowering time through microRNA pathways. Proc. Natl. Acad. Sci. USA 2016, 113, e2199–e2206. [Google Scholar] [CrossRef]
- Graeber, K.; Linkies, A.; Müller, K.; Wunchova, A.; Rott, A.; Leubner-Metzger, G. Cross-Species approaches to seed dormancy and germination: Conservation and biodiversity of ABA-regulated mechanisms and the Brassicaceae DOG1 genes. Plant Mol. Biol. 2009, 73, 67–87. [Google Scholar] [CrossRef]
- Sugimoto, K.; Takeuchi, Y.; Ebana, K.; Miyao, A.; Hirochika, H.; Hara, N.; Ishiyama, K.; Kobayashi, M.; Ban, Y.; Hattori, T.; et al. Molecular cloning of Sdr4, a regulator involved in seed dormancy and domestication of rice. Proc. Natl. Acad. Sci. USA 2010, 107, 5792–5797. [Google Scholar] [CrossRef] [PubMed]
- Ashikawa, I.; Abe, F.; Nakamura, S. DOG1-like genes in cereals: Investigation of their function by means of ectopic expression in Arabidopsis. Plant Sci. 2013, 208, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Née, G.; Kramer, K.; Nakabayashi, K.; Yuan, B.; Xiang, Y.; Miatton, E.; Finkemeier, I.; Soppe, W.J. Delay of Germination1 requires PP2C phosphatases of the ABA signalling pathway to control seed dormancy. Nat. Commun. 2017, 8, 72. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Qi, Y.; Chen, F.; Meng, Y.; Luo, X.; Shuai, H.; Zhou, W.; Ding, J.; Du, J.; Liu, J.; et al. Salt Stress Represses Soybean Seed Germination by Negatively Regulating GA Biosynthesis While Positively Mediating ABA Biosynthesis. Front. Plant Sci. 2017, 8, 1372. [Google Scholar] [CrossRef]
- Chen, F.; Zhou, W.; Yin, H.; Luo, X.; Chen, W.; Liu, X.; Wang, X.; Meng, Y.; Feng, L.; Qin, Y.; et al. Shading of the mother plant during seed development promotes subsequent seed germination in soybean. J. Exp. Bot. 2020, 71, 2072–2084. [Google Scholar] [CrossRef]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.A.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef]
- Buggs, R.J.A.; Elliott, N.M.; Zhang, L.; Koh, J.; Viccini, L.F.; Soltis, U.E.; Soltis, P.S. Tissue-Specific silencing of homoeologs in natural populations of the recent allopolyploid Tragopogon mirus. New Phytol. 2010, 186, 175–183. [Google Scholar] [CrossRef]
- Roulin, A.; Auer, P.L.; Libault, M.; Schlueter, J.; Farmer, A.; May, G.; Stacey, G.; Doerge, R.W.; Jackson, S.A. The fate of duplicated genes in a polyploid plant genome. Plant J. 2012, 73, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Woodhouse, M.R.; Pedersen, B.; Freeling, M. Transposed Genes in Arabidopsis Are Often Associated with Flanking Repeats. PLoS Genet. 2010, 6, e1000949. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Tang, H.; Tan, X.; Ficklin, S.P.; Feltus, F.A.; Paterson, A.H. Modes of Gene Duplication Contribute Differently to Genetic Novelty and Redundancy, but Show Parallels across Divergent Angiosperms. PLoS ONE 2011, 6, e28150. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Freeling, M. Bias in Plant Gene Content Following Different Sorts of Duplication: Tandem, Whole-Genome, Segmental, or by Transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef] [PubMed]
- Juretic, N.; Hoen, D.R.; Huynh, M.L.; Harrison, P.M.; Bureau, T.E. The evolutionary fate of MULE-mediated duplications of host gene fragments in rice. Genome Res. 2005, 15, 1292–1297. [Google Scholar] [CrossRef]
- Singh, V.K.; Jain, M. Genome-Wide survey and comprehensive expression profiling of Aux/IAA gene family in chickpea and soybean. Front. Plant Sci. 2015, 6, 6. [Google Scholar] [CrossRef]
- Ghosh, A.; Islam, T. Genome-Wide analysis and expression profiling of glyoxalase gene families in soybean (Glycine max) indicate their development and abiotic stress specific response. BMC Plant Biol. 2016, 16, 87. [Google Scholar] [CrossRef]
- Kim, S.; Park, M.; Yeom, S.-I.; Kim, Y.-M.; Lee, J.M.; Lee, H.-A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.-T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef]
- Chen, F.; Yang, Y.; Luo, X.; Zhou, W.; Dai, Y.; Zheng, C.; Weiguo, L.; Yang, W.; Shu, K. Genome-Wide identification of GRF transcription factors in soybean and expression analysis of GmGRF family under shade stress. BMC Plant Biol. 2019, 19, 269. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.-D.; Xie, Q.; He, Z. Two Faces of One Seed: Hormonal Regulation of Dormancy and Germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef]
- Shu, K.; Zhou, W.; Yang, W. Apetala 2-domain-containing transcription factors: focusing on abscisic acid and gibberellins antagonism. New Phytol. 2017, 217, 977–983. [Google Scholar] [CrossRef]
- Sarris, P.F.; Cevik, V.; Dagdas, G.; Jones, J.D.G.; Krasileva, K.V. Comparative analysis of plant immune receptor architectures uncovers host proteins likely targeted by pathogens. BMC Biol. 2016, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Peng, Y.; Rao, Y.; Li, S.; Zeng, L. Genome-Wide Identification and Expression Analysis of Sugar Transporter (ST) Gene Family in Longan (Dimocarpus longan L.). Plants 2020, 9, 342. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2013, 42, d222–d230. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39 (Suppl. 2), w29–w37. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2016, 45, d200–d203. [Google Scholar] [CrossRef]
- Jiangtao, C.; Yingzhen, K.; Qian, W.; Yuhe, S.; Daping, G.; Jing, L.; Guanshan, L. MapGene2Chrom, a tool to draw gene physical map based on Perl and SVG languages. Hered. Beijing 2015, 37, 91. [Google Scholar]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296–1297. [Google Scholar] [CrossRef]
- Bailey, T.L.; Bodén, M.; Buske, F.A.; Frith, M.C.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37 (Suppl. 2), w202–w208. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Murshudov, G.N.; Vagin, A.A.; Dodson, E.J. Refinement of Macromolecular Structures by the Maximum-Likelihood Method. Acta Crystallogr. Sect. D Biol. Crystallogr. 1997, 53, 240–255. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Li, L.; Zheng, T.; Zhuo, X.; Li, S.; Qiu, L.; Wang, J.; Cheng, T.; Zhang, Q. Genome-Wide identification, characterization and expression analysis of the HD-Zip gene family in the stem development of the woody plant Prunus mume. PeerJ 2019, 7, e7499. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, J.; Zhao, X.-Q.; Wang, J.; Wong, G.K.-S.; Yu, J. KaKs_Calculator: Calculating Ka and Ks Through Model Selection and Model Averaging. Genom. Proteom. Bioinform. 2006, 4, 259–263. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.M.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, S.; Zhou, Y.; Zhou, Y.; Yang, J.; Tang, X.-Q. Genome-Wide identification and characterization of GRAS transcription factors in sacred lotus (Nelumbo nucifera). PeerJ 2016, 4, e2388. [Google Scholar] [CrossRef]
- Shu, K.; Chen, Q.; Wu, Y.; Liu, R.; Zhang, H.; Wang, P.; Li, Y.; Wang, S.; Tang, S.; Liu, C.; et al. ABI4 mediates antagonistic effects of abscisic acid and gibberellins at transcript and protein levels. Plant J. 2016, 85, 348–361. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Locus ID | CDS (bp) | Chromosome Number | Number of Exons | Length (aa) | pI | MW (Da) |
|---|---|---|---|---|---|---|---|
| GmDOG1-L1 | Glyma.01G084200 | 1461 | 1 | 12 | 486 | 7.32 | 54309.28 |
| GmDOG1-L2 | Glyma.02G097900 | 1404 | 2 | 11 | 467 | 7.32 | 51853.46 |
| GmDOG1-L3 | Glyma.02G176800 | 1455 | 2 | 11 | 484 | 7.77 | 53927.08 |
| GmDOG1-L4 | Glyma.03G127600 | 1383 | 3 | 11 | 460 | 7 | 50928.00 |
| GmDOG1-L5 | Glyma.03G128200 | 873 | 3 | 7 | 290 | 8.82 | 32365.88 |
| GmDOG1-L6 | Glyma.03G142400 | 1476 | 3 | 11 | 491 | 8.22 | 55200.29 |
| GmDOG1-L7 | Glyma.04G151800 | 285 | 4 | 2 | 94 | 5.04 | 10511.23 |
| GmDOG1-L8 | Glyma.04G254800 | 1089 | 4 | 8 | 362 | 8.69 | 40929.00 |
| GmDOG1-L9 | Glyma.05G113200 | 579 | 5 | 2 | 192 | 4.97 | 22591.38 |
| GmDOG1-L10 | Glyma.05G182500 | 1113 | 5 | 8 | 370 | 7.11 | 42096.79 |
| GmDOG1-L11 | Glyma.05G195200 | 858 | 5 | 1 | 285 | 5.28 | 32592.96 |
| GmDOG1-L12 | Glyma.06G090900 | 495 | 6 | 2 | 164 | 9.97 | 18889.84 |
| GmDOG1-L13 | Glyma.06G107300 | 1068 | 6 | 8 | 355 | 6.21 | 39984.68 |
| GmDOG1-L14 | Glyma.07G151600 | 702 | 7 | 1 | 233 | 5.32 | 26409.83 |
| GmDOG1-L15 | Glyma.08G140100 | 1140 | 8 | 8 | 379 | 7.78 | 43105.04 |
| GmDOG1-L16 | Glyma.10G092100 | 1554 | 10 | 10 | 517 | 6.61 | 57921.5 |
| GmDOG1-L17 | Glyma.10G194800 | 696 | 10 | 4 | 231 | 7 | 26599.48 |
| GmDOG1-L18 | Glyma.10G276100 | 1371 | 10 | 11 | 456 | 6.08 | 50993.85 |
| GmDOG1-L19 | Glyma.10G296200 | 999 | 10 | 8 | 332 | 8.94 | 37141.79 |
| GmDOG1-L20 | Glyma.11G183700 | 1482 | 11 | 12 | 493 | 5.87 | 54385.80 |
| GmDOG1-L21 | Glyma.11G236300 | 1095 | 11 | 8 | 364 | 6.28 | 41178.84 |
| GmDOG1-L22 | Glyma.12G088700 | 1506 | 12 | 12 | 501 | 6.61 | 55253.16 |
| GmDOG1-L23 | Glyma.12G184500 | 1467 | 12 | 12 | 488 | 6.75 | 54118.97 |
| GmDOG1-L24 | Glyma.13G085100 | 1113 | 13 | 8 | 370 | 7.13 | 41915.73 |
| GmDOG1-L25 | Glyma.13G193700 | 1410 | 13 | 11 | 469 | 5.98 | 51989.98 |
| GmDOG1-L26 | Glyma.13G228600 | 765 | 13 | 1 | 254 | 8.70 | 28971.62 |
| GmDOG1-L27 | Glyma.13G269500 | 807 | 13 | 2 | 268 | 5.86 | 30487.66 |
| GmDOG1-L28 | Glyma.13G316900 | 1473 | 13 | 12 | 490 | 6.96 | 54630.43 |
| GmDOG1-L29 | Glyma.14G167000 | 1113 | 14 | 8 | 370 | 8.26 | 41888.77 |
| GmDOG1-L30 | Glyma.15G083900 | 873 | 15 | 1 | 290 | 9.37 | 32967.42 |
| GmDOG1-L31 | Glyma.15G182700 | 888 | 15 | 2 | 295 | 6.71 | 33829.21 |
| GmDOG1-L32 | Glyma.15G232000 | 1494 | 15 | 11 | 497 | 6.1 | 55125.35 |
| GmDOG1-L33 | Glyma.17G154000 | 636 | 17 | 3 | 211 | 5.95 | 24440.01 |
| GmDOG1-L34 | Glyma.18G020900 | 1089 | 18 | 8 | 362 | 7.78 | 41052.71 |
| GmDOG1-L35 | Glyma.18G202900 | 699 | 18 | 1 | 232 | 5.72 | 26078.56 |
| GmDOG1-L36 | Glyma.19G130200 | 1380 | 19 | 11 | 459 | 8.53 | 50717.82 |
| GmDOG1-L37 | Glyma.19G145300 | 1476 | 19 | 11 | 491 | 8.71 | 55358.4 |
| GmDOG1-L38 | Glyma.20G113600 | 1368 | 20 | 11 | 455 | 5.98 | 50671.73 |
| GmDOG1-L39 | Glyma.20G195100 | 708 | 20 | 4 | 235 | 8.2 | 26896.70 |
| GmDOG1-L40 | Glyma.20G246400 | 1335 | 20 | 11 | 444 | 5.91 | 49505.41 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Zheng, C.; Chandrasekaran, U.; Yu, L.; Liu, C.; Pu, T.; Wang, X.; Du, J.; Liu, J.; Yang, F.; et al. Identification and Bioinformatic Analysis of the GmDOG1-Like Family in Soybean and Investigation of Their Expression in Response to Gibberellic Acid and Abscisic Acid. Plants 2020, 9, 937. https://doi.org/10.3390/plants9080937
Yang Y, Zheng C, Chandrasekaran U, Yu L, Liu C, Pu T, Wang X, Du J, Liu J, Yang F, et al. Identification and Bioinformatic Analysis of the GmDOG1-Like Family in Soybean and Investigation of Their Expression in Response to Gibberellic Acid and Abscisic Acid. Plants. 2020; 9(8):937. https://doi.org/10.3390/plants9080937
Chicago/Turabian StyleYang, Yingzeng, Chuan Zheng, Umashankar Chandrasekaran, Liang Yu, Chunyan Liu, Tian Pu, Xiaochun Wang, Junbo Du, Jiang Liu, Feng Yang, and et al. 2020. "Identification and Bioinformatic Analysis of the GmDOG1-Like Family in Soybean and Investigation of Their Expression in Response to Gibberellic Acid and Abscisic Acid" Plants 9, no. 8: 937. https://doi.org/10.3390/plants9080937
APA StyleYang, Y., Zheng, C., Chandrasekaran, U., Yu, L., Liu, C., Pu, T., Wang, X., Du, J., Liu, J., Yang, F., Yong, T., Yang, W., Liu, W., & Shu, K. (2020). Identification and Bioinformatic Analysis of the GmDOG1-Like Family in Soybean and Investigation of Their Expression in Response to Gibberellic Acid and Abscisic Acid. Plants, 9(8), 937. https://doi.org/10.3390/plants9080937