
Collaborative Impact of Compost and Beneficial Rhizobacteria on Soil Properties, Physiological Attributes, and Productivity of Wheat Subjected to Deficit Irrigation in Salt Affected Soil

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
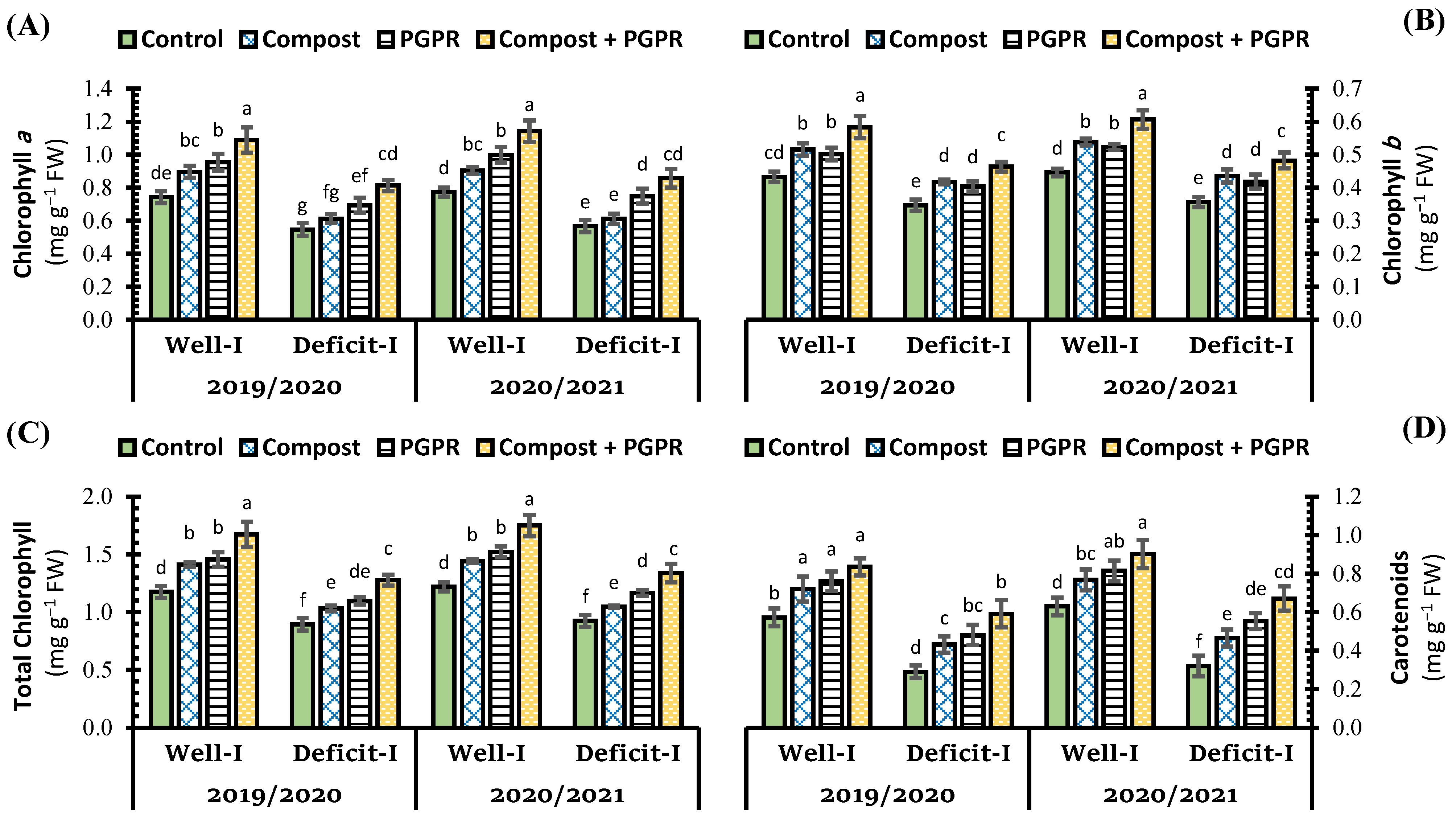
2.1. Effect of Soil Treatments on Soil Exchangeable Na Percentage, Soil Enzymes, Ion Selectivity and Photosynthetic Pigment Contents under Abiotic Stress
2.2. Effect of Soil Treatments on Physiological Attributes under Abiotic Stress
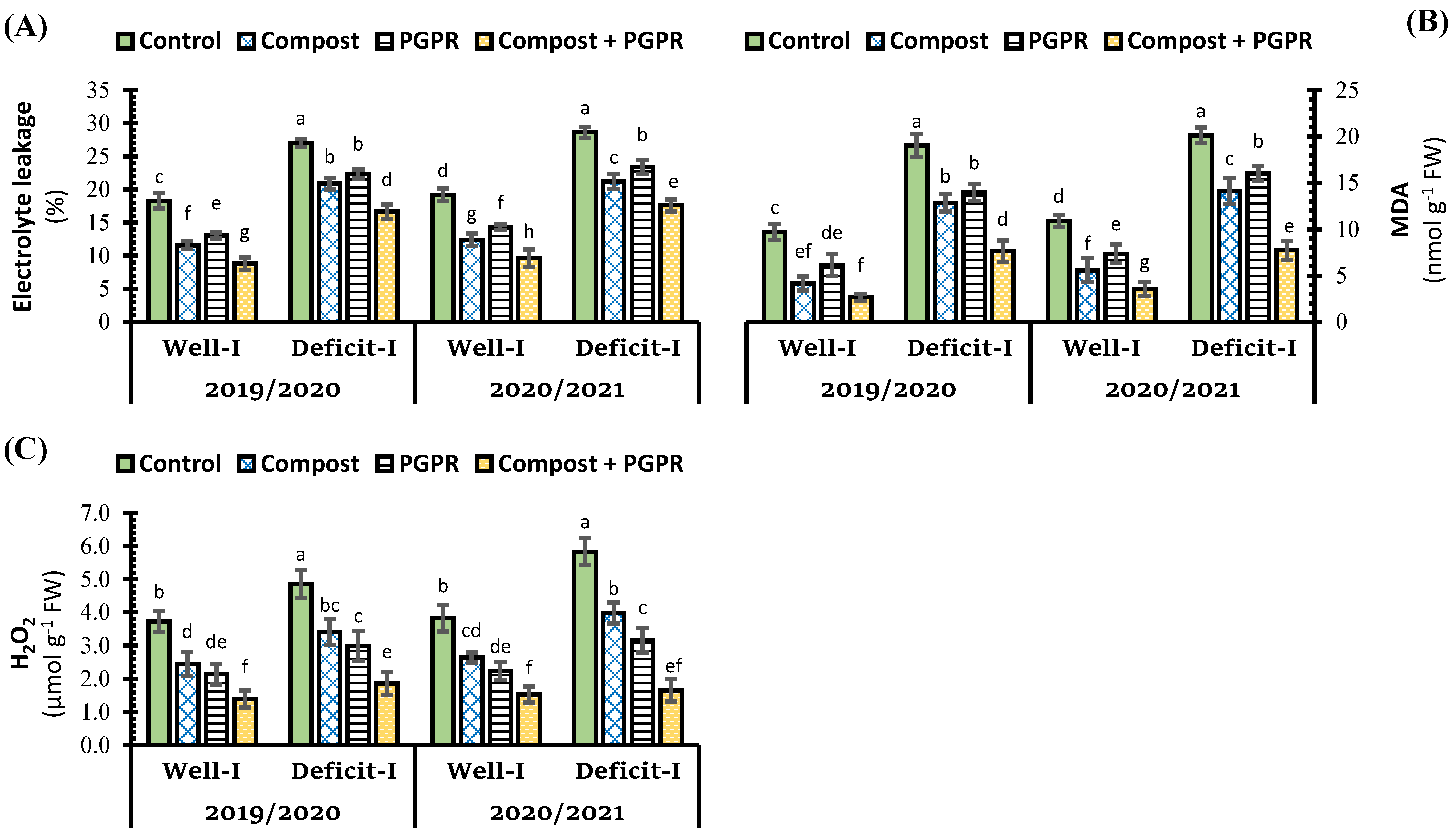
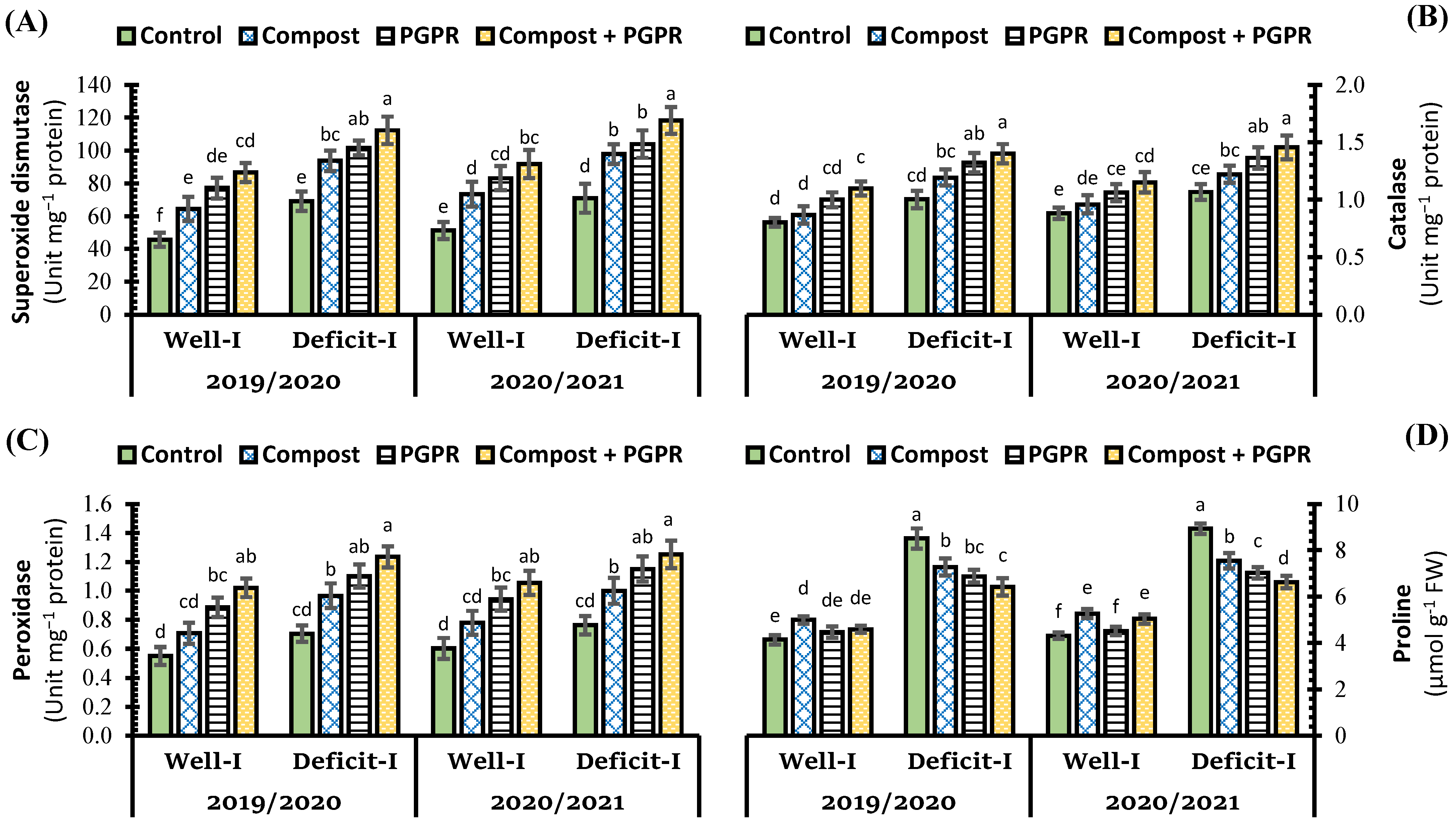
2.3. Effect of Soil Treatments on Oxidative Stress Indicators and Antioxidant System in Wheat Leaves under Abiotic Stress
2.4. Effect of Soil Treatments on Yield Parameters and Related Traits under Abiotic Stress
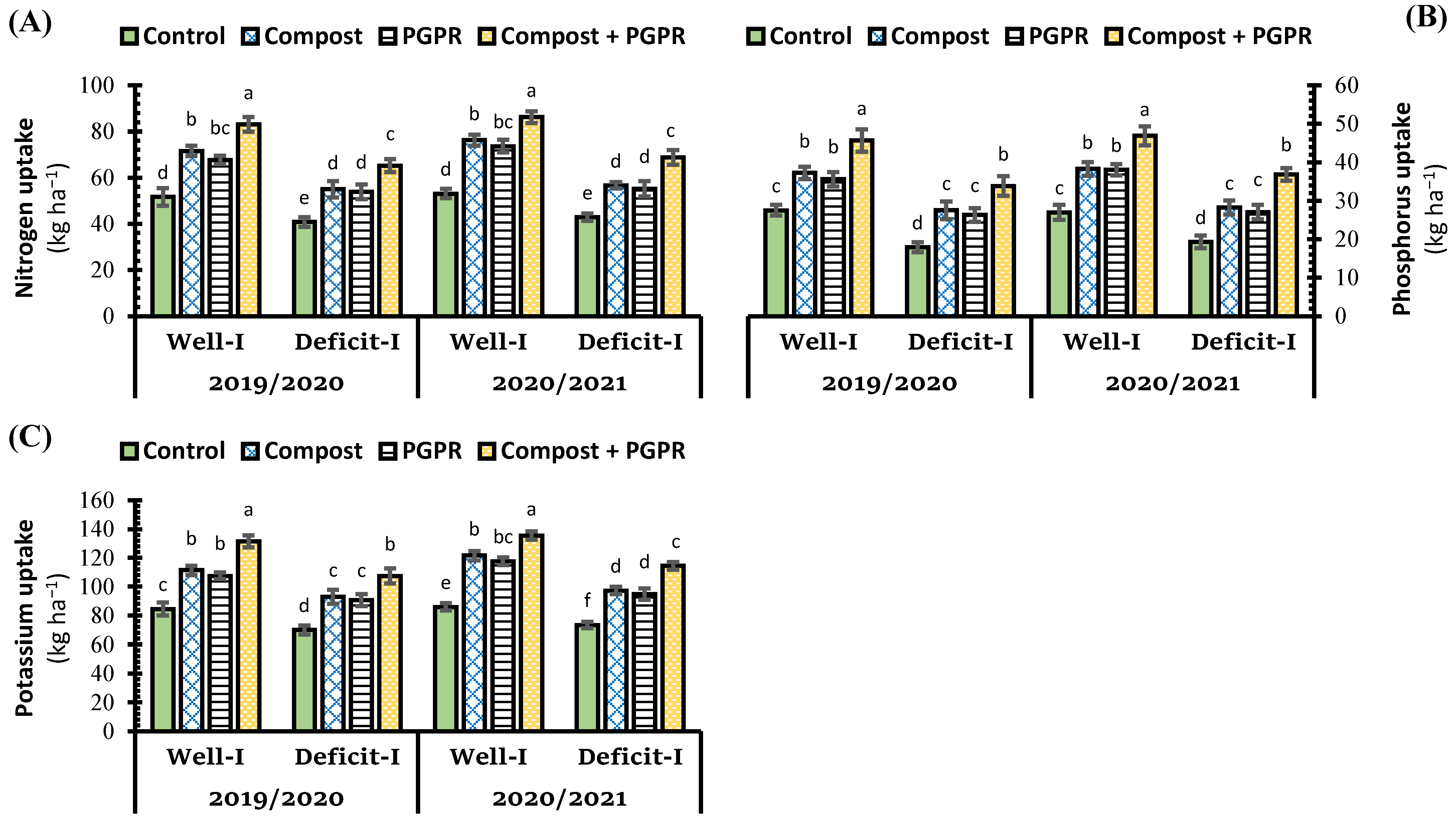
2.5. Effect of Soil Treatments on Nutrient Uptake under Abiotic Stress
3. Discussion
3.1. Effect of Soil Treatments on Soil Exchangeable Na Percentage, Soil Enzymes, Ion Selectivity and Photosynthetic Pigment Contents under Abiotic Stress
3.2. Effect of Soil Treatments on Physiological Attributes under Abiotic Stress
3.3. Effect of Soil Treatments on Antioxidant Enzyme Activity and Oxidative Stress Indicators in Wheat Leaves under Abiotic Stress
3.4. Effect of Soil Treatments on Yield Parameters and Related Traits under Abiotic Stress
3.5. Effect of Soil Treatments on Nutrient Uptake under Abiotic Stress
4. Materials and Methods
4.1. Study Site Description and Plant Material
4.2. Experimental Design and Treatments
4.2.1. Compost Characterization
4.2.2. Microorganisms and Culture Conditions
4.3. Soil Measurements
4.3.1. Soil Exchangeable Percentage of Na+
4.3.2. Soil Enzymes (Dehydrogenase and Urease) Activity
4.4. Plant Measurements
4.4.1. Measurements of Na+ and K+ Ions in Leaves
4.4.2. Physiological Attributes
Photosynthetic Pigment Contents
Stomatal Conductance
Leaf Water Relations
4.4.3. Oxidative Stress Indicators and Antioxidant System in Wheat Leaves
Oxidative Stress Indicators
Antioxidant System
4.5. Yield Parameters and Related Traits
Nutrient Uptake
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Food Outlook—Biannual Report on Global Food Markets; FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Tahir, S.; Ahmad, A.; Khaliq, T.; Cheema, M.J. Evaluating the impact of seed rate and sowing dates on wheat productivity in semi-arid environment. Int. J. Agric. Biol. 2019, 22, 57–64. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munns, R.; Day, D.A.; Fricke, W.; Watt, M.; Arsova, B.; Barkla, B.J.; Bose, J.; Byrt, C.S.; Chen, Z.-H.; Foster, K.J.; et al. Energy costs of salt tolerance in crop plants. New Phytol. 2020, 225, 1072–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royo, A.; Abió, D. Salt tolerance in durum wheat cultivars. Span. J. Agric. Res. 2003, 1, 27–35. [Google Scholar] [CrossRef]
- Olawumi, T.O.; Chan, D.W.M. A scientometric review of global research on sustainability and sustainable development. J. Clean. Prod. 2018, 183, 231–250. [Google Scholar] [CrossRef]
- Hafez, E.; Omara, A.E.D.; Ahmed, A. The coupling effects of plant growth promoting rhizobacteria and salicylic acid on physiological modifications, yield traits, and productivity of wheat under water deficient conditions. Agronomy 2019, 9, 524. [Google Scholar] [CrossRef] [Green Version]
- Hafez, E.M.; Omara, A.E.D.; Alhumaydhi, F.A.; El-Esawi, M.A. Minimizing hazard impacts of soil salinity and water stress on wheat plants by soil application of vermicompost and biochar. Physiol. Plant. 2020, 172, 587–602. [Google Scholar] [CrossRef]
- FAO. The future of food and agriculture–Trends and challenges. Annu. Rep. 2017, 296, 1–180. [Google Scholar]
- Allison, L.; Richards, L. Diagnosis and Improvement of Saline and Alkali Soils; No. 60. Soil and Water Conservative Research Branch, Agricultural Research Service, US Department of Agriculture: Washington, DC, USA, 1954.
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Flowers, T.J.; Galal, H.K.; Bromham, L. Evolution of halophytes: Multiple origins of salt tolerance in land plants. Funct. Plant Biol. 2010, 37, 604–612. [Google Scholar] [CrossRef]
- Talebnejad, R.; Sepaskhah, A.R. Physiological characteristics, gas exchange, and plant ion relations of quinoa to different saline groundwater depths and water salinity. Arch. Agron. Soil Sci. 2016, 62, 1347–1367. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killi, D.; Haworth, M. Diffusive and metabolic constraints to photosynthesis in quinoa during drought and salt stress. Plants 2017, 6, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, A.; Aslam, Z.; Javed, T.; Hussain, S.; Raza, A.; Shabbir, R.; Mora-Poblete, F.; Saeed, T.; Zulfiqar, F.; Ali, M.M.; et al. Screening of wheat (Triticum aestivum L.) genotypes for drought tolerance through agronomic and physiological response. Agronomy 2022, 12, 287. [Google Scholar] [CrossRef]
- Valdivia-Cea, W.; Bustamante, L.; Jara, J.; Fischer, S.; Holzapfel, E.; Wilckens, R. Effect of soil water availability on physiological parameters, yield, and seed quality in four quinoa genotypes (Chenopodium quinoa Willd.). Agronomy 2021, 11, 1012. [Google Scholar] [CrossRef]
- Hafez, E.M.; Osman, H.S.; El-Razek, U.A.A.; Elbagory, M.; Omara, A.E.-D.; Eid, M.A.; Gowayed, S.M. Foliar-applied potassium silicate coupled with plant growth-promoting rhizobacteria improves growth, physiology, nutrient uptake and productivity of faba bean (Vicia faba L.) irrigated with saline water in salt-affected soil. Plants 2021, 10, 894. [Google Scholar] [CrossRef]
- Osman, H.S.; Gowayed, S.M.; Elbagory, M.; Omara, A.E.; El-Monem, A.M.A.; Abd El-Razek, U.A.; Hafez, E.M. Interactive impacts of beneficial microbes and Si-Zn nanocomposite on growth and productivity of soybean subjected to water deficit under salt-affected soil conditions. Plants 2021, 10, 1396. [Google Scholar] [CrossRef]
- Zhu, J.-K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Abdelrasheed, K.G.; Mazrou, Y.; Omara, A.E.-D.; Osman, H.S.; Nehela, Y.; Hafez, E.M.; Rady, A.M.S.; El-Moneim, D.A.; Alowaiesh, B.F.; Gowayed, S.M. Soil amendment using biochar and application of K-humate enhance the growth, productivity, and nutritional value of onion (Allium cepa L.) under deficit irrigation conditions. Plants 2021, 10, 2598. [Google Scholar] [CrossRef]
- Leal, L.d.S.G.; Pessoa, L.G.M.; de Oliveira, J.P.; Santos, N.A.; Silva, L.F.d.S.; Júnior, G.B.; Freire, M.B.G.d.S.; de Souza, E.S. Do applications of soil conditioner mixtures improve the salt extraction ability of Atriplex nummularia at early growth stage? Int. J. Phytoremediation 2020, 22, 482–489. [Google Scholar] [CrossRef]
- Osman, H.S.; Rady, A.M.S.; Awadalla, A.; Omara, A.E.-D.; Hafez, E.M. Improving the antioxidants system, growth, and sugar beet quality subjected to long-term osmotic stress by phosphate solubilizing bacteria and compost tea. Inter. J. Plant Prod. 2021, 16, 119–135. [Google Scholar] [CrossRef]
- Hafez, E.M.; Osman, H.S.; Gowayed, S.M.; Okasha, S.A.; Omara, A.E.-D.; Sami, R.; Abd El-Monem, A.M.; Abd El-Razek, U.A. Minimizing the adversely impacts of water deficit and soil salinity on maize growth and productivity in response to the application of plant growth-promoting rhizobacteria and silica nanoparticles. Agronomy 2021, 11, 676. [Google Scholar] [CrossRef]
- Sadegh-Zadeh, F.; Tolekolai, S.F.; Bahmanyar, M.A.; Emadi, M. Application of biochar and compost for enhancement of rice (Oryza sativa L.) grain yield in calcareous sandy soil. Comm. Soil Sci. Plant Anal. 2018, 49, 552–566. [Google Scholar] [CrossRef]
- Youssif, N.E.E.; Osman, H.S.M.; Salama, Y.A.M.; Zaghlool, S.A.M. Effect of rice straw and applications of potassium silicate, potassium humate and seaweed extract on growth and some macronutrients of sweet pepper plants under irrigation deficit. Arab Univ. J. Agric. Sci. 2018, 26, 755–773. [Google Scholar] [CrossRef]
- Mahmoud, E.; Ibrahim, M.; Ali, N.; Ali, H. Effect of biochar and compost amendments on soil biochemical properties and dry weight of canola plant grown in soil contaminated with heavy metals. Comm. Soil Sci. Plant Anal. 2020, 51, 1561–1571. [Google Scholar] [CrossRef]
- Diacono, M.; Montemurro, F. Long-Term Effects of Organic Amendments on Soil Fertility. In Sustainable Agriculture Volume 2; Lichtfouse, E., Hamelin, M., Navarrete, M., Debaeke, P., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 761–786. [Google Scholar]
- Wu, Y.; Li, Y.; Zhang, Y.; Bi, Y.; Sun, Z. Responses of saline soil properties and cotton growth to different organic amendments. Pedosphere 2018, 28, 521–529. [Google Scholar] [CrossRef]
- Wu, Y.; Li, Y.; Zheng, C.; Zhang, Y.; Sun, Z. Organic amendment application influence soil organism abundance in saline alkali soil. Eur. J. Soil Biol. 2013, 54, 32–40. [Google Scholar] [CrossRef]
- Walker, D.J.; Bernal, M.P. The effects of olive mill waste compost and poultry manure on the availability and plant uptake of nutrients in a highly saline soil. Bioresour. Technol. 2008, 99, 396–403. [Google Scholar] [CrossRef]
- Eboibi, O.; Akpokodje, O.I.; Nyorere, O.; Oghenerukevwe, P.; Uguru, H. Effect of pre-harvest applications of organic manure and calcium chloride on the storability of tomato fruits. Ann. Agric. Sci. 2021, 66, 142–151. [Google Scholar] [CrossRef]
- Swarnalakshmi, K.; Yadav, V.; Tyagi, D.; Dhar, D.W.; Kannepalli, A.; Kumar, S. Significance of plant growth promoting rhizobacteria in grain legumes: Growth promotion and crop production. Plants 2020, 9, 1596. [Google Scholar] [CrossRef]
- Paul, D.; Lade, H. Plant-growth-promoting rhizobacteria to improve crop growth in saline soils: A review. Agron. Sustain. Develop. 2014, 34, 737–752. [Google Scholar] [CrossRef]
- Fghire, R.; Anaya, F.; Ali, O.I.; Benlhabib, O.; Ragab, R.; Wahbi, S. Physiological and photosynthetic response of quinoa to drought stress. Chil. J. Agric. Res. 2015, 75, 174–183. [Google Scholar] [CrossRef] [Green Version]
- Ilangumaran, G.; Smith, D.L. Plant growth promoting rhizobacteria in amelioration of salinity stress: A systems biology perspective. Front. Plant Sci. 2017, 8, 1768. [Google Scholar] [CrossRef] [PubMed]
- Ayuso-Calles, M.; Flores-Félix, J.D.; Rivas, R. Overview of the role of rhizobacteria in plant salt stress tolerance. Agronomy 2021, 11, 1759. [Google Scholar] [CrossRef]
- Hafez, E.M.; Abou El Hassan, W.H.; Gaafar, I.A.; Seleiman, M.F. Effect of gypsum application and irrigation intervals on clay saline-sodic soil characterization, rice water use efficiency, growth, and yield. J. Agric. Sci. 2015, 7, 208. [Google Scholar] [CrossRef] [Green Version]
- Nehela, Y.; Mazrou, Y.S.A.; Alshaal, T.; Rady, A.M.S.; El-Sherif, A.M.A.; Omara, A.E.-D.; Abd El-Monem, A.M.; Hafez, E.M. The integrated amendment of sodic-saline soils using biochar and plant growth-promoting rhizobacteria enhances maize (Zea mays L.) resilience to water salinity. Plants 2021, 10, 1960. [Google Scholar] [CrossRef]
- Hafez, E.M.; Kheir, A.M.S.; Badawy, S.A.; Rashwan, E.; Farig, M.; Osman, H.S. Differences in physiological and biochemical attributes of wheat in response to single and combined salicylic acid and biochar subjected to limited water irrigation in saline sodic soil. Plants 2020, 9, 1346. [Google Scholar] [CrossRef]
- Ismail, F.; Badawi, F.S.F.; Ahmed, F.H. Effect of different irrigation intervals, compost and bacterial inoculation on sesame productity in sandy soils. J. Agric. Chem. Biotechnol. 2013, 4, 181–195. [Google Scholar] [CrossRef]
- Kanwal, S.; Ilyas, N.; Batool, N.; Arshad, M. Amelioration of drought stress in wheat by combined application of PGPR, compost, and mineral fertilizer. J. Plant Nutr. 2017, 40, 1250–1260. [Google Scholar] [CrossRef]
- Ramzani, P.M.A.; Shan, L.; Anjum, S.; Khan, W.-u.-D.; Ronggui, H.; Iqbal, M.; Virk, Z.A.; Kausar, S. Improved quinoa growth, physiological response, and seed nutritional quality in three soils having different stresses by the application of acidified biochar and compost. Plant Physiol. Biochem. 2017, 116, 127–138. [Google Scholar] [CrossRef]
- Singh, R.; Singh, P.; Singh, H.; Raghubanshi, A.S. Impact of sole and combined application of biochar, organic and chemical fertilizers on wheat crop yield and water productivity in a dry tropical agro-ecosystem. Biochar 2019, 1, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Hafez, E.M.; Alsohim, A.S.; Farig, M.; Omara, A.E.-D.; Rashwan, E.; Kamara, M.M. Synergistic effect of biochar and plant growth promoting rhizobacteria on alleviation of water deficit in rice plants under salt-affected soil. Agronomy 2019, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Bashan, Y.; Salazar, B.G.; Moreno, M.; Lopez, B.R.; Linderman, R.G. Restoration of eroded soil in the Sonoran Desert with native leguminous trees using plant growth-promoting microorganisms and limited amounts of compost and water. J. Environ. Manag. 2012, 102, 26–36. [Google Scholar] [CrossRef]
- Duo, L.A.; Liu, C.X.; Zhao, S.L. Alleviation of drought stress in turfgrass by the combined application of nano-compost and microbes from compost. Russ. J. Plant Physiol. 2018, 65, 419–426. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Imran, M.; Naveed, M.; Khan, M.Y.; Ahmad, M.; Zahir, Z.A.; Crowley, D.E. Synergistic use of biochar, compost and plant growth-promoting rhizobacteria for enhancing cucumber growth under water deficit conditions. J. Sci. Food Agric. 2017, 97, 5139–5145. [Google Scholar] [CrossRef]
- Uzinger, N.; Takács, T.; Szili-Kovács, T.; Radimszky, L.; Füzy, A.; Draskovits, E.; Szűcs-Vásárhelyi, N.; Molnár, M.; Farkas, É.; Kutasi, J.; et al. Fertility impact of separate and combined treatments with biochar, sewage sludge compost and bacterial inocula on acidic sandy soil. Agronomy 2020, 10, 1612. [Google Scholar] [CrossRef]
- Yaseen, R.; Aziz, O.; Saleem, M.H.; Riaz, M.; Zafar-ul-Hye, M.; Rehman, M.; Ali, S.; Rizwan, M.; Nasser Alyemeni, M.; El-Serehy, H.A.; et al. Ameliorating the drought stress for wheat growth through application of ACC-deaminase containing rhizobacteria along with biogas slurry. Sustainability 2020, 12, 6022. [Google Scholar] [CrossRef]
- Kamara, M.M.; Rehan, M.; Ibrahim, K.M.; Alsohim, A.S.; Elsharkawy, M.M.; Kheir, A.M.S.; Hafez, E.M.; El-Esawi, M.A. Genetic diversity and combining ability of white maize inbred lines under different plant densities. Plants 2020, 9, 1140. [Google Scholar] [CrossRef]
- Salim, B.B.M.; Hikal, M.S.; Osman, H.S. Ameliorating the deleterious effects of saline water on the antioxidants defense system and yield of eggplant using foliar application of zinc sulphate. Ann. Agric. Sci. 2019, 64, 244–251. [Google Scholar] [CrossRef]
- Salim, B.B.M.; Abou El-Yazied, A.; Salama, Y.A.M.; Raza, A.; Osman, H.S. Impact of silicon foliar application in enhancing antioxidants, growth, flowering and yield of squash plants under deficit irrigation condition. Ann. Agric. Sci. 2021, 66, 176–183. [Google Scholar] [CrossRef]
- Osman, H.S.; Salim, B.B. Improving yield and quality of kohlrabi stems growing under NaCl salinity using foliar application of urea and seaweed extract. J. Hort. Sci. Ornament. Plant 2016, 8, 149–160. [Google Scholar]
- Kheir, A.M.S.; Abouelsoud, H.M.; Hafez, E.M.; Ali, O.A.M. Integrated effect of nano-Zn, nano-Si, and drainage using crop straw–filled ditches on saline sodic soil properties and rice productivity. Arab. J. Geosci. 2019, 12, 471. [Google Scholar] [CrossRef]
- Ding, Z.; Kheir, A.M.S.; Ali, O.A.M.; Hafez, E.M.; ElShamey, E.A.; Zhou, Z.; Wang, B.; Lin, X.e.; Ge, Y.; Fahmy, A.E.; et al. A vermicompost and deep tillage system to improve saline-sodic soil quality and wheat productivity. J. Environ. Manag. 2021, 277, 111388. [Google Scholar] [CrossRef] [PubMed]
- Hafez, E.; Farig, M. Efficacy of salicylic acid as a cofactor for ameliorating effects of water stress and enhancing wheat yield and water use efficiency in saline soil. Inter. J. Plant Prod. 2019, 13, 163–176. [Google Scholar] [CrossRef]
- Hafez, E.M. Influence of salicylic acid on ion distribution, enzymatic activity and some agromorphological characteristics of wheat under salt-affected soil. Egypt. J. Agron. 2016, 38, 455–469. [Google Scholar] [CrossRef] [Green Version]
- Osman, H.S.; Salim, B.B.M. Enhancing antioxidants defense system of snap bean under NaCl salinity using foliar application of salicylic acid, spermidine and glycine betaine. Am.-Eurasian J. Agric. Environ. Sci. 2016, 16, 1200–1210. [Google Scholar]
- Seleiman, M.F.; Refay, Y.; Al-Suhaibani, N.; Al-Ashkar, I.; El-Hendawy, S.; Hafez, E.M. Integrative effects of rice-straw biochar and silicon on oil and seed quality, yield and physiological traits of Helianthus annuus L. grown under water deficit stress. Agronomy 2019, 9, 637. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, S. ROS and Oxidative Stress: Origin and Implication. In Reactive Oxygen Species in Plant Biology; Springer: New Delhi, India, 2019; pp. 1–31. [Google Scholar]
- Nafees, M.; Fahad, S.; Shah, A.N.; Bukhari, M.A.; Maryam; Ahmed, I.; Ahmad, S.; Hussain, S. Reactive Oxygen Species Signaling in Plants. In Plant Abiotic Stress Tolerance: Agronomic, Molecular and Biotechnological Approaches; Hasanuzzaman, M., Hakeem, K.R., Nahar, K., Alharby, H.F., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 259–272. [Google Scholar]
- Chung, W.-H. Unraveling new functions of superoxide dismutase using yeast model system: Beyond its conventional role in superoxide radical scavenging. J. Microbiol. 2017, 55, 409–416. [Google Scholar] [CrossRef]
- Ahmad, R.; Hussain, S.; Anjum, M.A.; Khalid, M.F.; Saqib, M.; Zakir, I.; Hassan, A.; Fahad, S.; Ahmad, S. Oxidative stress and antioxidant defense mechanisms in plants under salt stress. In Plant Abiotic Stress Tolerance: Agronomic, Molecular and Biotechnological Approaches; Hasanuzzaman, M., Hakeem, K.R., Nahar, K., Alharby, H.F., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 191–205. [Google Scholar]
- Alam, M.Z.; Carpenter-Boggs, L.; Hoque, M.A.; Ahammed, G.J. Effect of soil amendments on antioxidant activity and photosynthetic pigments in pea crops grown in arsenic contaminated soil. Heliyon 2020, 6, e05475. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the regulation of reactive oxygen species metabolism in plants under abiotic stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef]
- Hafez, E.M.; Gowayed, S.M.; Nehela, Y.; Sakran, R.M.; Rady, A.M.S.; Awadalla, A.; Omara, A.E.-D.; Alowaiesh, B.F. Incorporated biochar-based soil amendment and exogenous glycine betaine foliar application ameliorate rice (Oryza sativa L.) tolerance and resilience to osmotic stress. Plants 2021, 10, 1930. [Google Scholar] [CrossRef] [PubMed]
- Atlas, R.M. Handbook of Microbiological Media, 4th ed.; CRC Press: Boca Raton, FL, USA, 2010; p. 2040. [Google Scholar]
- Arshad, M.A.C.; Lowery, B.; Grossman, B. Physical Tests for Monitoring Soil Quality. In SSSA Special Publications; Soil Science Society of America: Madison, WI, USA, 2015; pp. 123–141. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Mersi, W.V. Dehydrogenase activity with the substrate INT. In Methods in Soil Biology; Schinner, F., Öhlinger, R., Kandeler, E., Margesin, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 243–245. [Google Scholar]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Johnston, C.T.; Sumner, M.E. Methods of Soil Analysis: Part 3 Chemical Methods; Soil Science Society of America, Inc.; American Society of Agronomy, Inc.: Madison, WI, USA, 1996. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Izanloo, A.; Condon, A.G.; Langridge, P.; Tester, M.; Schnurbusch, T. Different mechanisms of adaptation to cyclic water stress in two South Australian bread wheat cultivars. J. Exp. Bot. 2008, 59, 3327–3346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weatherley, P.E. Studies in the water relations of the cotton plant. I. The field measurement of water deficits in leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Du, Z.; Bramlage, W.J. Modified thiobarbituric acid assay for measuring lipid oxidation in sugar-rich plant tissue extracts. J. Agric. Food Chemist. 1992, 40, 1566–1570. [Google Scholar] [CrossRef]
- Bajji, M.; Kinet, J.-M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Analyt. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; pp. 121–126. [Google Scholar]
- Vetter, J.L.; Steinberg, M.P.; Nelson, A.I. Enzyme assay, quantitative determination of peroxidase in sweet corn. J. Agric. Food Chemist. 1958, 6, 39–41. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; Association of Official Analytical Chemists: Washengton, DC, USA, 2005. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Irrigation | Soil Amendments | ESP (%) | Urease (mg N-NH4+ g−1 soil h−1) | Dehydrogenase (mg TPF g−1 DM soil d−1) | |||
|---|---|---|---|---|---|---|---|
| 2019/2020 | 2020/2021 | 2019/2020 | 2020/2021 | 2019/2020 | 2020/2021 | ||
| Well Irrigation | Control | 18.0 ± 0.4 c | 16.5 ± 1.2 b | 115.9 ± 4.1 e | 124.0 ± 5.5 d | 59.3 ± 4.7 e | 62.9 ± 3.1 f |
| Compost | 11.9 ± 0.6 f | 11.4 ± 0.4 d | 149.3 ± 6.2 d | 158.8 ± 5.0 c | 90.8 ± 3.7 cd | 96.0 ± 4.0 d | |
| PGPR | 14.8 ± 0.6 e | 13.1 ± 1.0 cd | 167.1 ± 4.6 b | 181.9 ± 5.1 b | 110.9 ± 5.3 b | 116.7 ± 4.7 b | |
| Compost + PGPR | 9.6 ± 1.0 g | 9.1 ± 1.0 e | 214.4 ± 6.0 a | 223.4 ± 7.2 a | 130.4 ± 5.5 a | 134.6 ± 5.3 a | |
| Deficit Irrigation | Control | 23.8 ± 0.7 a | 22.4 ± 0.6 a | 85.8 ± 3.2 g | 94.7 ± 3.4 e | 36.6 ± 3.1 f | 48.5 ± 4.0 g |
| Compost | 18.2 ± 1.2 c | 17.4 ± 0.4 b | 109.0 ± 4.0 f | 116.6 ± 5.2 d | 69.3 ± 2.6 e | 79.8 ± 5.1 e | |
| PGPR | 21.2 ± 1.3 b | 20.7 ± 1.0 a | 118.8 ± 5.1 e | 122.8 ± 4.7 d | 83.6 ± 3.1 d | 92.6 ± 4.3 d | |
| Compost + PGPR | 16.6 ± 0.9 d | 14.3 ± 0.5 c | 153.0 ± 4.9 c | 160.3 ± 5.6 c | 101.4 ± 4.1 bc | 103.7 ± 4.2 c | |
| Irrigation | ** | *** | *** | ** | ** | ** | |
| Soil amendments treatments | *** | *** | *** | *** | *** | *** | |
| Irrigation × treatments | * | * | *** | ** | * | * | |
| Irrigation | Soil Amendments | Na (mg g−1 DW) | K (mg g−1 DW) | K/Na (Ratio) | |||
|---|---|---|---|---|---|---|---|
| 2019/2020 | 2020/2021 | 2019/2020 | 2020/2021 | 2019/2020 | 2020/2021 | ||
| Well Irrigation | Control | 4.24 ± 0.19 d | 4.30 ± 0.33 d | 3.04 ± 0.08 de | 3.10 ± 0.09 e | 0.72 ± 0.05 de | 0.72 ± 0.06 d |
| Compost | 3.37 ± 0.14 f | 3.45 ± 0.08 f | 3.87 ± 0.10 b | 3.88 ± 0.13 b | 1.15 ± 0.03 b | 1.13 ± 0.06 b | |
| PGPR | 3.68 ± 0.21 e | 3.75 ± 0.07 e | 3.56 ± 0.11 c | 3.65 ± 0.12 c | 0.97 ± 0.06 c | 0.98 ± 0.05 c | |
| Compost + PGPR | 3.02 ± 0.19 g | 3.02 ± 0.14 g | 4.28 ± 0.18 a | 4.40 ± 0.10 a | 1.42 ± 0.15 a | 1.46 ± 0.09 a | |
| Deficit Irrigation | Control | 6.16 ± 0.30 a | 6.01 ± 0.21 a | 2.28 ± 0.11 g | 2.38 ± 0.13 g | 0.37 ± 0.01 g | 0.04 ± 0.03 f |
| Compost | 4.73 ± 0.15 c | 4.74 ± 0.20 c | 2.9 ± 0.09 ef | 2.98 ± 0.08 ef | 0.61 ± 0.03 ef | 0.63 ± 0.02 e | |
| PGPR | 5.11 ± 0.15 b | 5.19 ± 0.14 b | 2.67 ± 0.10 f | 2.83 ± 0.10 f | 0.52 ± 0.04 f | 0.54 ± 0.01 e | |
| Compost + PGPR | 4.17 ± 0.16 d | 4.25 ± 0.18 d | 3.31 ± 0.13 cd | 3.43 ± 0.11 d | 0.79 ± 0.03 d | 0.81 ± 0.05 d | |
| Irrigation | ** | ** | ** | ** | ** | ** | |
| Soil amendments treatments | *** | *** | *** | *** | *** | *** | |
| Irrigation × Treatments | * | * | ns | ns | ** | ** | |
| Year Month | 2019/2020 | 2020/2021 | ||||||
|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | Rainfall | Relative Humidity (%) | Temperature (°C) | Rainfall | Relative Humidity (%) | |||
| Max | Min | (mm) | Max | Min | (mm) | |||
| Nov | 26.3 | 17.2 | 0.98 | 32.6 | 25.3 | 16.2 | 0.94 | 31.6 |
| Dec | 25.9 | 15.3 | 0.85 | 34.2 | 24.9 | 14.3 | 0.82 | 33.2 |
| Jan | 24.5 | 13.2 | 1.1 | 35.1 | 23.2 | 12.4 | 0.54 | 32.7 |
| Feb | 22.3 | 10.3 | 3.1 | 46.2 | 20.3 | 11.1 | 3.32 | 42.4 |
| Mar | 21.4 | 9.7 | 6.4 | 44.3 | 20.6 | 10.7 | 6.85 | 43.1 |
| April | 23.7 | 13.8 | 0.5 | 43.8 | 22.5 | 12.5 | 0.63 | 44.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omara, A.E.-D.; Hafez, E.M.; Osman, H.S.; Rashwan, E.; El-Said, M.A.A.; Alharbi, K.; Abd El-Moneim, D.; Gowayed, S.M. Collaborative Impact of Compost and Beneficial Rhizobacteria on Soil Properties, Physiological Attributes, and Productivity of Wheat Subjected to Deficit Irrigation in Salt Affected Soil. Plants 2022, 11, 877. https://doi.org/10.3390/plants11070877
Omara AE-D, Hafez EM, Osman HS, Rashwan E, El-Said MAA, Alharbi K, Abd El-Moneim D, Gowayed SM. Collaborative Impact of Compost and Beneficial Rhizobacteria on Soil Properties, Physiological Attributes, and Productivity of Wheat Subjected to Deficit Irrigation in Salt Affected Soil. Plants. 2022; 11(7):877. https://doi.org/10.3390/plants11070877
Chicago/Turabian StyleOmara, Alaa El-Dein, Emad M. Hafez, Hany S. Osman, Emadeldeen Rashwan, Mohamed A. A. El-Said, Khadiga Alharbi, Diaa Abd El-Moneim, and Salah M. Gowayed. 2022. "Collaborative Impact of Compost and Beneficial Rhizobacteria on Soil Properties, Physiological Attributes, and Productivity of Wheat Subjected to Deficit Irrigation in Salt Affected Soil" Plants 11, no. 7: 877. https://doi.org/10.3390/plants11070877
APA StyleOmara, A. E.-D., Hafez, E. M., Osman, H. S., Rashwan, E., El-Said, M. A. A., Alharbi, K., Abd El-Moneim, D., & Gowayed, S. M. (2022). Collaborative Impact of Compost and Beneficial Rhizobacteria on Soil Properties, Physiological Attributes, and Productivity of Wheat Subjected to Deficit Irrigation in Salt Affected Soil. Plants, 11(7), 877. https://doi.org/10.3390/plants11070877