
Glucosinolates and Isothiocyantes in Processed Rapeseed Determined by HPLC-DAD-qTOF

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Glucosinolates in Raw Material
2.2. Glucosinolates during the Processing of the Pressed Rapeseed Cake
2.3. Glucosinolates in Different Extracted Rapeseed Protein Sample
2.4. Isothiocyanates in Raw Material and during Processing of Rapeseed Pressed Cake
3. Materials and Methods
3.1. Sample Description
Pilot-Scale Rapeseed Protein Extraction
3.2. Determination of Glucosinolates
3.3. Determination of Isothiocyanates
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority (EFSA). Glucosinolates as undesirable substances in animal feed-Scientific Opinion of the Panel on Contaminants in the Food Chain. EFSA J. 2008, 6, 1–76. [Google Scholar]
- Wallsgrove, R.; Benett, R.; Kiddle, G.; Bartlet, E.; Ludwig-Mueller, J. Glucosinolate biosynthesis and pest/disease interactions. Young 1999, 1, 49. Available online: http://www.regional.org.au/au/gcirc/3/393.htm/ (accessed on 12 June 2021).
- Tolrà, R.P.; Alonso, R.; Poschenrieder, C.; Barceló, D.; Barceló, J. Determination of glucosinolates in rapeseed and Thlaspi caerulescens plants by liquid chromatography–atmospheric pressure chemical ionization mass spectrometry. J. Chromatogr. A 2000, 889, 75–81. [Google Scholar] [CrossRef]
- Song, L.; Morrison, J.J.; Botting, N.P.; Thornalley, P.J. Analysis of glucosinolates, isothiocyanates, and amine degradation products in vegetable extracts and blood plasma by LC–MS/MS. Anal. Biochem. 2005, 347, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.; Bones, J.; Rossiter, T. Sub-cellular immunolocalization of the glucosinolate sinigrin in seedlings of Brassica juncea. Planta 1998, 206, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, T.A.; Fahey, J.W.; Wade, K.L.; Stephenson, K.K.; Talalay, P. Chemoprotective glucosinolates and isothiocyanates of broccoli sprouts: Metabolism and excretion in humans. Cancer Epidemiol. Prev. Biomark. 2001, 10, 501–508. Available online: https://cebp.aacrjournals.org/content/10/5/501/ (accessed on 18 March 2021).
- Rodrigues, E.A.S.; Rosa, E. Effect of post-harvest treatments on the level of glucosinolates in broccoli. J. Sci. Food Agric. 1999, 79, 1028–1032. [Google Scholar] [CrossRef]
- Verkerk, R.; Dekker, M.; Jongen, W.M.F. Post-harvest increase of indolyl glucosinolates in response to chopping and storage of Brassica vegetables. J. Sci. Food Agric. 2001, 81, 953–958. [Google Scholar] [CrossRef]
- Quinsac, A.; Charrier, D.; Ribaillier, D.; Péron, J.Y. Glucosinolates in etiolated sprouts of sea-kale (Crambe maritima L.). J. Sci. Food Agric. 1994, 65, 201–207. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Geng, F.; Tang, L.; Li, Y.; Yang, L.; Choi, K.-S.; Kazim, A.L.; Zhang, Y. Allyl isothiocyanate arrests cancer cells in mitosis, and mitotic arrest in turn leads to apoptosis via Bcl-2 protein phosphorylation. J. Biol. Chem. 2011, 286, 32259–32267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, T.K.; Lund, E.K.; Parker, M.L.; Clarke, R.G.; Johnson, I.T. Allyl-isothiocyanate causes mitotic block, loss of cell adhesion and disrupted cytoskeletal structure in HT29 cells. Carcinogenesis 2004, 25, 1409–1415. [Google Scholar] [CrossRef] [Green Version]
- Pham, N.-A.; Jacobberger, J.W.; Schimmer, A.D.; Cao, P.; Gronda, M.; Hedley, D.W. The dietary isothiocyanate sulforaphane targets pathways of apoptosis, cell cycle arrest, and oxidative stress in human pancreatic cancer cells and inhibits tumor growth in severe combined immunodeficient mice. Mol. Cancer Ther. 2004, 3, 1239–1248. [Google Scholar] [PubMed]
- Mellon, F.A.; Bennett, R.N.; Holst, B.; Williamson, G. Intact glucosinolate analysis in plant extracts by programmed cone voltage electrospray LC/MS: Performance and comparison with LC/MS/MS methods. Anal. Biochem. 2002, 306, 83–91. [Google Scholar] [CrossRef]
- Singh, H.; Mehta, P.; Vig, A.P. Removal of Glucosinolates from Rapeseed Meal Using Bowl-Shaped Tetrameric Molecules in Apolar Solvent. J. Agric. Food Chem. 1997, 45, 4522–4524. [Google Scholar] [CrossRef]
- Serraino, M.R.; Thompson, L.U. Removal of phytic acid and protein-phytic acid interactions in rapeseed. J. Agric. Food Chem. 1984, 32, 38–40. [Google Scholar] [CrossRef]
- Heaney, R.K.; Fenwick, G.R. The analysis of glucosinolates in Brassica species using gas chromatography. Direct determination of the thiocyanate ion precursors, glucobrassicin and neoglucobrassicin. J. Sci. Food Agric. 1980, 31, 593–599. [Google Scholar] [CrossRef]
- Velasco, P.; Soengas, P.; Vilar, M.; Cartea, M.E.; Del Rio, M. Comparison of glucosinolate profiles in leaf and seed tissues of different Brassica napus crops. J. Am. Soc. Hortic. Sci. 2008, 133, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Sosulski, F.W.; Dabrowski, K.J. Determination of glucosinolates in canola meal and protein products by desulfation and capillary gas-liquid chromatography. J. Agric. Food Chem. 1984, 32, 1172–1175. [Google Scholar] [CrossRef]
- Millán, S.; Sampedro, M.C.; Gallejones, P.; Castellón, A.; Ibargoitia, M.L.; Goicolea, M.A.; Barrio, R.J. Identification and quantification of glucosinolates in rapeseed using liquid chromatography–ion trap mass spectrometry. Anal. Bioanal. Chem. 2009, 394, 1661–1669. [Google Scholar] [CrossRef] [PubMed]
- van Poppel, G.; Verhoeven, D.T.; Verhagen, H.; Goldbohm, R.A. Brassica vegetables and cancer prevention. Epidemiology and mechanisms. Adv. Exp. Med. Biol. 1999, 472, 159–313. [Google Scholar] [CrossRef] [PubMed]
- Seow, A.; Yuan, J.M.; Sun, C.L.; van den Berg, D.; Lee, H.; Yu, M.C. Dietary isothiocyanates, glutathione S-transferase polymorphisms and colorectal cancer risk in the Singapore Chinese health study. Carcinogenesis 2002, 23, 2055–2061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, A.; Tang, L.; Li, Y.; Geng, F.; Paonessa, J.D.; Chen, S.C.; Wong, M.K.; Zhang, Y. Inhibition of bladder cancer development by allyl isothiocyanate. Carcinogenesis 2010, 31, 281–286. [Google Scholar] [CrossRef]
- London, S.J.; Yuan, J.-M.; Chung, F.L.; Gao, Y.-T.; Coetzee, G.A.; Ross, R.K.; Yu, M.C. Isothiocyanates, glutathione S-transferase M1 and T1 polymorphisms, and lung-cancer risk: A prospective study of men in Shanghai, China. Lancet 2000, 356, 724–729. [Google Scholar] [CrossRef]
- Fowke, J.H.; Chung, F.L.; Jin, F.; Qi, D.; Cai, Q.Y.; Conaway, C.; Cheng, J.R.; Shu, X.O.; Gao, Y.T.; Zheng, W. Urinary isothiocyanate levels, brassica, and human breast cancer. Cancer Res. 2003, 63, 3980–3986. [Google Scholar] [PubMed]
- Kirsh, V.A.; Peters, U.; Mayne, S.T.; Subar, A.F.; Chatterjee, N.; Johnson, C.C.; Hayes, R.B. Prospective study of fruit and vegetable intake and risk of prostate cancer. J. Natl. Cancer Inst. 2007, 99, 1200–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vig, A.P.; Rampal, G.; Singh, T.S.; Arora, S. Bioprotective effects of glucosinolates—A review. LWT Food Sci. Technol. 2009, 42, 1561–1572. [Google Scholar] [CrossRef]
- Holst, B.; Williamson, G. A critical review of the bioavailability of glucosinolates and related compounds. Nat. Prod. Rep. 2004, 21, 425–447. [Google Scholar] [CrossRef]
- Andersen, K.E.; Frandsen, H.B.; Jensen, S.K.; Bellostas, N.M.; Sørensen, A.D.; Sørensen, J.C.; Sørensen, H. Glucosinolates in Brassica–Health risks, but also benefits. Nor. Acad. Sci. Lett. 2010, 1, 104–124. [Google Scholar]
- Mawson, R.; Heaney, R.K.; Zdunczyk, Z.; Kozłowska, H. Rapeseed meal-glucosinolates and their antinutritional effects, Part 3: Animal growth and performance. Nahrung 1994, 38, 167–177. [Google Scholar] [CrossRef]
- Mawson, R.; Heaney, R.K.; Piskuła, M.; Kozłowska, H. Rapeseed meal-glucosinolates and their antinutritional effects. Part 1. Rapeseed production and chemistry of glucosinolates. Nahrung 1993, 37, 131–140. [Google Scholar] [CrossRef]
- Burel, C.; Boujard, T.; Kaushik, S.J.; Boeuf, G.; Mol, K.A.; Van der, G.S.; Darras, V.M.; Kuhn, E.R.; Prade Balade, B.; Querat, B.; et al. Effects of rapeseed meal-glucosinolates on thyroid metabolism and feed utilization in rainbow trout. GenComp. Endocrinol. 2001, 124, 343–358. [Google Scholar] [CrossRef] [Green Version]
- Conaway, C.C.; Yang, Y.M.; Chung, F.L. Isothiocyanates as cancer chemopreventive agents: Their biological activities and metabolism in rodents and humans. Curr. Drug Metab. 2002, 3, 233–255. [Google Scholar] [CrossRef] [PubMed]
- Szydłowska-Czerniak, A.; Bartkowiak-Broda, I.; Karlović, I.; Karlovits, G.; Szłyk, E. Antioxidant capacity, total phenolics, glucosinolates and colour parameters of rapeseed cultivars. Food Chem. 2011, 127, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Rosa, E.A.S.; Heaney, R.K.; Fenwick, G.R.; Portaz, C.A.M. Glucosinolates in crop plants. Hort. Rev. 1997, 19, 99–215. [Google Scholar] [CrossRef]
- El-Beltagi, H.E.D.S.; Mohamed, A.A. Variations in fatty acid composition, glucosinolate profile and some phytochemical contents in selected oil seed rape (Brassica napus L.) cultivars. Grasas Aceites 2010, 61, 143–150. [Google Scholar] [CrossRef]
- Travers-Martin, N.; Kuhlmann, F.; Müller, C. Revised determination of free and complexed myrosinase activities in plant extracts. Plant Physiol. Biochem. 2008, 46, 506–516. [Google Scholar] [CrossRef]
- Verkerk, R.; Schreiner, M.; Krumbein, A.; Ciska, E.; Holst, B.; Rowland, I.; De Schrjiver, R.; Hansen, M.; Gerhauser, C.; Mithen, R.; et al. Glucosinolates in Brassica vegetables: The influence of the food supply chain on intake, bioavailability and human health. Mol. Nutr. Food Res. 2008, 53, 219–256. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kushad, M.M. Purification and characterization of myrosinase from horseradish (Armoracia rusticana) roots. Plant Physiol. Biochem. 2005, 43, 503–511. [Google Scholar] [CrossRef]
- Jing, P.; Zhao, S.J.; Ruan, S.Y.; Xie, Z.H.; Dong, Y.; Yu, L.L. Anthocyanin and glucosinolate occurrences in the roots of Chinese red radish (Raphanus sativus L.), and their stability to heat and pH. Food Chem. 2012, 133, 1569–1576. [Google Scholar] [CrossRef]
- Gevers, E.; Andrade, I.; Hallikainen, A. Information on inherent food plant toxicants: A resource generated by the EU-AIR NETTOX project (1995–1997). In Nettox Compilation of Consumption Data; Report No. 4; Danish Veterinary and Food Administration: Søborg, Denmark, 1998; p. 144. [Google Scholar]
- Deng, Q.; Zinoviadou, K.G.; Galanakis, C.M.; Orlien, V.; Grimi, N.; Vorobiev, E.; Lobovka, N.; Barba, F.J. The effects of conventional and non-conventional processing on glucosinolates and its derived forms, isothiocyanates: Extraction, degradation, and applications. Food Eng. Rev. 2015, 7, 357–381. [Google Scholar] [CrossRef]
- Kozlowska, H.J.; Nowak, H.; Nowak, J. Characterisation of myrosinase in polish varieties of rapeseed. J. Sci. Food Agric. 1983, 34, 1171–1178. [Google Scholar] [CrossRef]
- Morra, M.J.; Kirkegaard, J.A. Isothiocyanate release from soil-incorporated Brassica tissues. Soil Biol. Biochem. 2002, 34, 1683–1690. [Google Scholar] [CrossRef]
- Mithen, R. Glucosinolates and their degradation products. Adv. Bot. Res. 2001, 35, 213–262. [Google Scholar] [CrossRef]
- Lüthy, B.; Matile, P. The mustard oil bomb: Rectified analysis of the subcellular organisation of the myrosinase system. Biochem. Physiol. Pflanzen 1984, 179, 5–12. [Google Scholar] [CrossRef]
- Langer, P.; Michajlovskij, N.; Sedlak, J.; Kutka, M. Studies on the antithyroid activity of naturally occurring L-5-vinyl-2-thiooxazolidone in man. Endokrinologie 1971, 57, 225–229. [Google Scholar] [PubMed]
- Jie, C.; Luo, Z.; Chen, H.; Wang, M.; Yan, C.; Mao, Z.F.; Xiao, G.K.; Kurihara, H.; Li, Y.F.; He, R.R. Indirubin, a bisindole alkaloid from Isatis indigotica, reduces H1N1 susceptibility in stressed mice by regulating MAVS signaling. Oncotarget 2017, 8, 105615–105629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Li, J.; Shi, Y.; Yang, L.; Wang, Z.; Han, H.; Wang, R. Stereoselective pharmacokinetic study of epiprogoitrin and progoitrin in rats with UHPLC–MS/MS method. J. Pharm. Biomed. Anal. 2020, 187, 113356. [Google Scholar] [CrossRef] [PubMed]
- Cakaloglu, B.; Vasfiye, H.O.; Semih, O. Cold press in oil extraction. A review. Ukr. Food J. 2018, 7, 640–654. [Google Scholar] [CrossRef]
- EEC. Rapessed—Determination of glucosinolates content—Part 1: Method using highperformance liquid chromatography. ISO 9167–1, 1–9. Available online: https://www.iso.org/standard/16763.html/ (accessed on 1 July 2019).
- Szűcs, Z.; Plaszkó, T.; Cziáky, Z.; Kiss-Szikszai, A.; Emri, T.; Bertóti, R.; Sinka, L.T.; Vasas, G.; Gonda, S. Endophytic fungi from the roots of horseradish (Armoracia rusticana) and their interactions with the defensive metabolites of the glucosinolate-myrosinase-isothiocyanate system. BMC Plant Biol. 2018, 18, 85. [Google Scholar] [CrossRef] [Green Version]
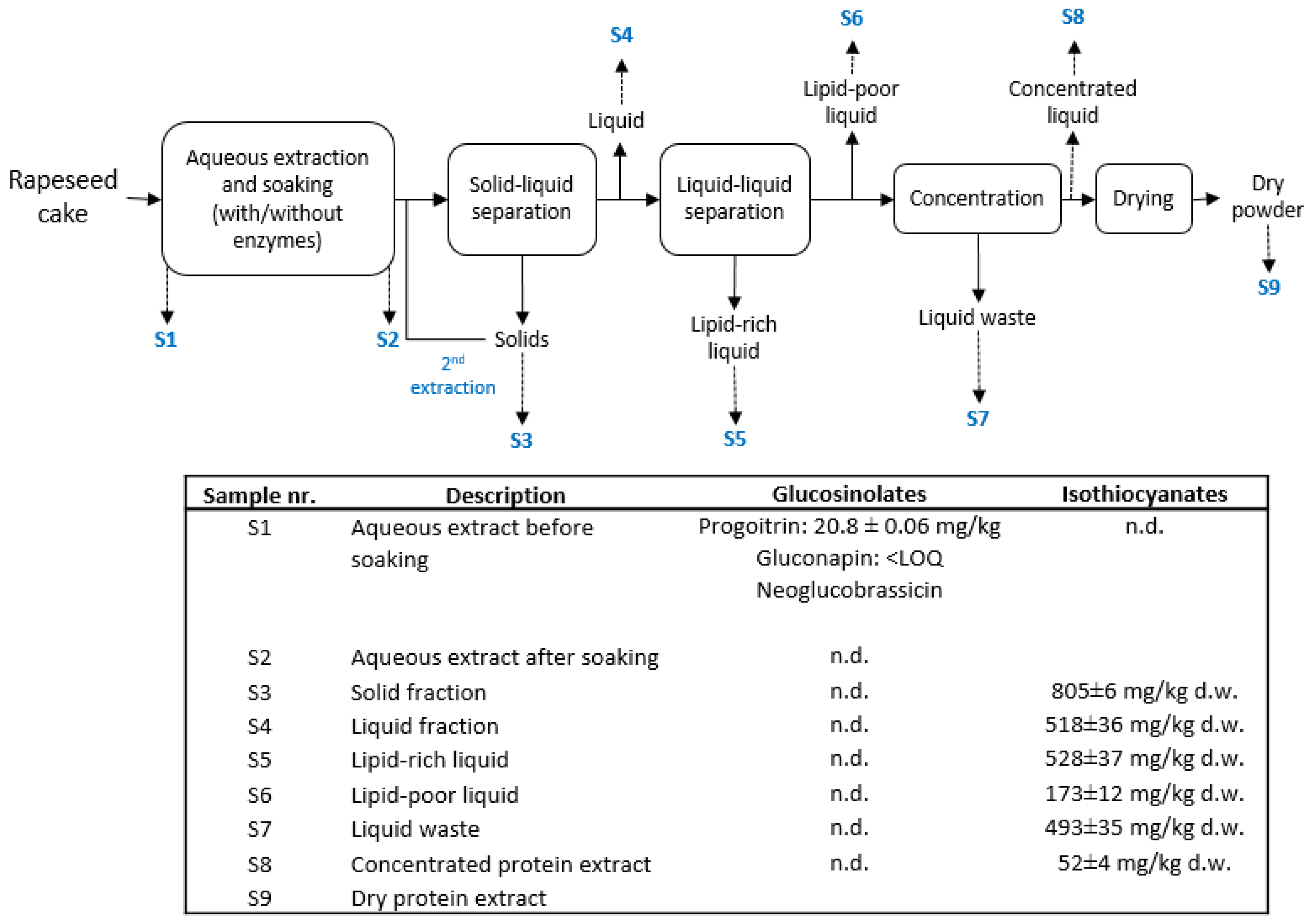

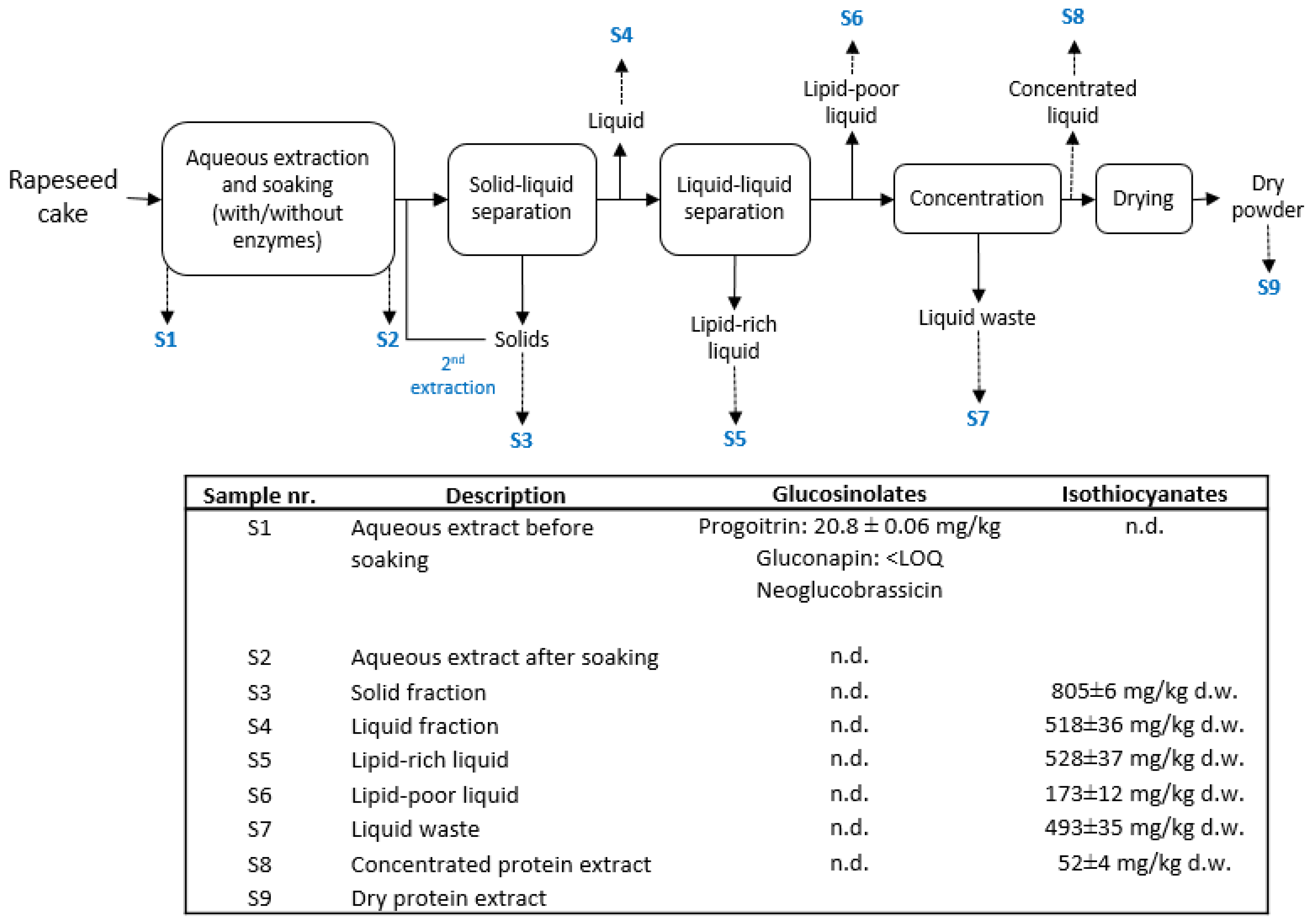{kind=link}
| Name of the Compound | Formula | M | [M − H]− | Characteristic Fragments | Fragment 96/97 Ratio | RT | Conc. |
|---|---|---|---|---|---|---|---|
| (min) | |||||||
| 2-Hydroxy-3-butenyl glucosinolate (progoitrin) | C11H19NO10S2 | 389.0460 | 388.0389 | 96.9595 | 1: 2 | 2.26 | 1014 ± 56 mg/kg |
| 259.0183 | (2.6 mmol/kg) | ||||||
| 2-Propenyl glucosinolate (sinigrin) | C10H17NO9S2 | 359.0287 | 358.0287 | 96.9586 | 1: 2 | 2.92 | 3.6 ± 0.09 |
| mg/kg | |||||||
| 259.0012 | (0.01 mmol/kg) | ||||||
| p-Hydroxybenzyl glucosinolate (sinalbin) | C14H19NO10S2 | 425.0460 | 424.0387 | 96.9595 | 1: 4 | 4.39 | 21.7 ± 2.2 |
| 259.0106 | mg/kg | ||||||
| (0.05 mmol/kg) | |||||||
| 2-Hydroxy-4-pentenyl glucosinolate (gluconapoliferin) | C12H21NO10S2 | 403.0620 | 402.0549 | 96.9602 | 1: 3 | 4.98 | 114.8 * ± 8.0 |
| 259.0130 | mg/kg | ||||||
| (0.28 mmol/kg) | |||||||
| Glucoalyssin | C13H25NO10S3 | 451.0660 | 450.0586 | 96.9602 | 1: 2 | 5.51 | 52.1 * ± 0.04 |
| 259.0079 | mg/kg | ||||||
| (0.12 mmol/kg) | |||||||
| 3-Butenyl glucosinolate (gluconapin) | C11H19NO9S2 | 373.0510 | 372.0439 | 96.9601 | 1: 3 | 5.93 | 2413 ± 27 |
| 259.0146 | mg/kg | ||||||
| (6.5 mmol/kg) | |||||||
| 2-Methylpropyl glucosinolate/butyl glucosinolate | C11H21NO9S2 | 375.0670 | 374.0598 | 96.9523 | 1: 3 | 10.2 | 16.3 * ± 1.1 |
| 259.0247 | mg/kg | ||||||
| (0.04 mmol/kg) | |||||||
| 4-Pentenyl glucosinolate | C12H21NO9S2 | 387.0670 | 386.0598 | 96.9601 | 1: 3 | 12.3 | 23.7 * ± 1.7 |
| (glucobrassicanapin) | 259.0124 | mg/kg | |||||
| (0.06 mmol/kg) | |||||||
| 4-Methyltiobutyl glucosinolate | C12H23NO9S3 | 421.0540 | 420.0472 | 96.9602 | 1: 3 | 15.0 | <LOQ |
| 259.0137 | |||||||
| 2-Phenylethyl glucosinolate (gluconasturtiin) | C15H21NO9S2 | 423.0670 | 422.0597 | 96.9608 | 1: 3 | 20.2 | 22.0 ± 0.8 |
| 259.0075 | mg/kg | ||||||
| (0.05 mmol/kg) | |||||||
| Methylpentyl glucosinolate | C13H25NO9S2 | 403.0990 | 402.0915 | 96.9604 | 1: 2 | 24.2 | 31.7 * ± 2.2 |
| 259.0098 | mg/kg | ||||||
| (0.08 mmol/kg) | |||||||
| Methylpentyl glucosinolate | C13H25NO9S2 | 403,098 | 402.0911 | 96.9591 | 1: 2 | 24.7 | 19.5 * ± 1.4 |
| 259.0118 | mg/kg | ||||||
| (0.05 mmol/kg) | |||||||
| Methylpentyl glucosinolate | C13H25NO9S2 | 403,099 | 402.0913 | 96.9627 | 1: 2 | 25.7 | 11.5 *± 0.8 |
| mg/kg | |||||||
| (0.03 mmol/kg) | |||||||
| Neoglucobrassicin | C17H22N2O10S2 | 478.073 | 477.0657 | 96.9605 | 1: 3 | 26.6 | <LOQ |
| 259.0131 | |||||||
| 446.0503 | |||||||
| Total determined glucosinolates | 3740 ± 261 mg/kg | ||||||
| (9.9 ± 0.7 mmol/kg) |
| HPR-1 | HPR-2 | HPR-3 | |
|---|---|---|---|
| Progoitrin | 1600 ± 160 mg/kg | 1720 ± 10 mg/kg | 2565 ± 3.0 mg/kg |
| Gluconapoliferin | 28.3 * ± 0.1 mg/kg | 54 * ± 5 mg/kg | 3.0 * ± 0.3 mg/kg |
| Glucoalyssin | <LOD | 35.4*± 0.3 mg/kg | 52 * ± 0.3 mg/kg |
| Gluconapin | 1479 ± 5.8 mg/kg | 2024 ± 2.7 mg/kg | 2209 ± 29 mg/kg |
| Butyl glucosinolate | 5.3 * ± 0.05 mg/kg | 4.8 * ± 0.03 mg/kg | 5.3 * ± 0.1 mg/kg |
| 4-Pentenyl glucosinolate | 16.5 * ± 0.08 mg/kg | 64.0 *± 1.0 mg/kg | 60.9 * ± 0.2 mg/kg |
| Gluconasturtiin | <LOD | 11.3 ± 0.3 mg/kg | 28.9 ± 0.4 mg/kg |
| Neoglucobrassicin | <LOD | 1.9 * ± 0.1 mg/kg | 10.3 * ± 0.2 mg/kg |
| 3-Methylpentyl glucosinolate | <LOD | 1.5 *± 0.05 mg/kg | 4.1 *± 0.3 mg/kg |
| 4-Methyltiobutyl glucosinolate | <LOD | <LOD | 4.7 * ± 0.3 mg/kg |
| Name of the Isothiocyanate Identified | Spray-Dried Protein Extract | Oven-Dried Protein Extract (Trial CPR-3) | Spray-Dried Product Extract (Trial CPR-4) |
|---|---|---|---|
| (Trial CPR-2) | |||
| Goitrin | 95 ± 7 mg/kg | 7 ± 0.5 mg/kg | 22 ± 1.5 mg/kg |
| 2-Phenylethyl isothiocyanate | <LOD | <LOQ | <LOD |
| (from Gluconasturtiin) | |||
| 4-Methylthiobutyl isothiocyanate | <LOQ | <LOD | <LOD |
| 3-Butenyl isothiocyanate | <LOD | <LOD | <LOQ |
| (from Gluconapin) |
| Raw Material | Trial No. | Brief Process Description |
|---|---|---|
| CPR | 1 | Soaking overnight, manual liquid–liquid separation and 5 kDa ultrafiltration for concentration, spray drying. 5-day process |
| CPR | 2 | Soaking overnight, 0.2 µ microfiltration for liquid–liquid separation and 5 kDa ultrafiltration for concentration, spray drying. 4-day process |
| CPR | 3 | Soaking overnight, 0.2 µ microfiltration for liquid–liquid separation and 10 kDa ultrafiltration for concentration, oven drying. 5-day process |
| CPR | 4 | Soaking overnight, 0.2 µ microfiltration for liquid–liquid separation and 10 kDa ultrafiltration for concentration, final product in liquid form. 5-day process |
| HPR | 1 | Soaking/hydrolysis overnight at 55 °C, enzymatic extraction at pH 8, 0.2 µ microfiltration for liquid–liquid separation and 10 kDa ultrafiltration for concentration, spray drying. |
| HPR | 2 | Soaking/hydrolysis overnight at 58 °C, enzymatic extraction at pH 6.7, 0.2 µ microfiltration for liquid–liquid separation and 300 Da nanofiltration for concentration, spray drying. |
| HPR | 3 | Soaking/hydrolysis for ca. 4 h at 50 °C, enzymatic extraction at pH 5.0 for 1 h followed by pH adjustment to pH 7.0 a subsequent 30 min reaction time, 0.2 µ microfiltration for liquid–liquid separation and 300 Da nanofiltration for concentration, final product in liquid form. |
| Raw Material | Trial No. | Dry Product Yield |
|---|---|---|
| (g/100 g Dry Raw Material) | ||
| CPR | 1 | 4.3 |
| CPR | 2 | 3.6 |
| CPR | 3 | 12.3 |
| CPR | 4 | 6.1 |
| HPR | 1 | 18.1 |
| HPR | 2 | 23.1 |
| HPR | 3 | 14.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miklavčič Višnjevec, A.; Tamayo Tenorio, A.; Steenkjær Hastrup, A.C.; Hansen, N.M.L.; Peeters, K.; Schwarzkopf, M. Glucosinolates and Isothiocyantes in Processed Rapeseed Determined by HPLC-DAD-qTOF. Plants 2021, 10, 2548. https://doi.org/10.3390/plants10112548
Miklavčič Višnjevec A, Tamayo Tenorio A, Steenkjær Hastrup AC, Hansen NML, Peeters K, Schwarzkopf M. Glucosinolates and Isothiocyantes in Processed Rapeseed Determined by HPLC-DAD-qTOF. Plants. 2021; 10(11):2548. https://doi.org/10.3390/plants10112548
Chicago/Turabian StyleMiklavčič Višnjevec, Ana, Angelica Tamayo Tenorio, Anne Christine Steenkjær Hastrup, Natanya Majbritt Louie Hansen, Kelly Peeters, and Matthew Schwarzkopf. 2021. "Glucosinolates and Isothiocyantes in Processed Rapeseed Determined by HPLC-DAD-qTOF" Plants 10, no. 11: 2548. https://doi.org/10.3390/plants10112548
APA StyleMiklavčič Višnjevec, A., Tamayo Tenorio, A., Steenkjær Hastrup, A. C., Hansen, N. M. L., Peeters, K., & Schwarzkopf, M. (2021). Glucosinolates and Isothiocyantes in Processed Rapeseed Determined by HPLC-DAD-qTOF. Plants, 10(11), 2548. https://doi.org/10.3390/plants10112548