Phenology Response to Climatic Dynamic across China’s Grasslands from 1985 to 2010


Abstract
1. Introduction
2. Materials and Methods
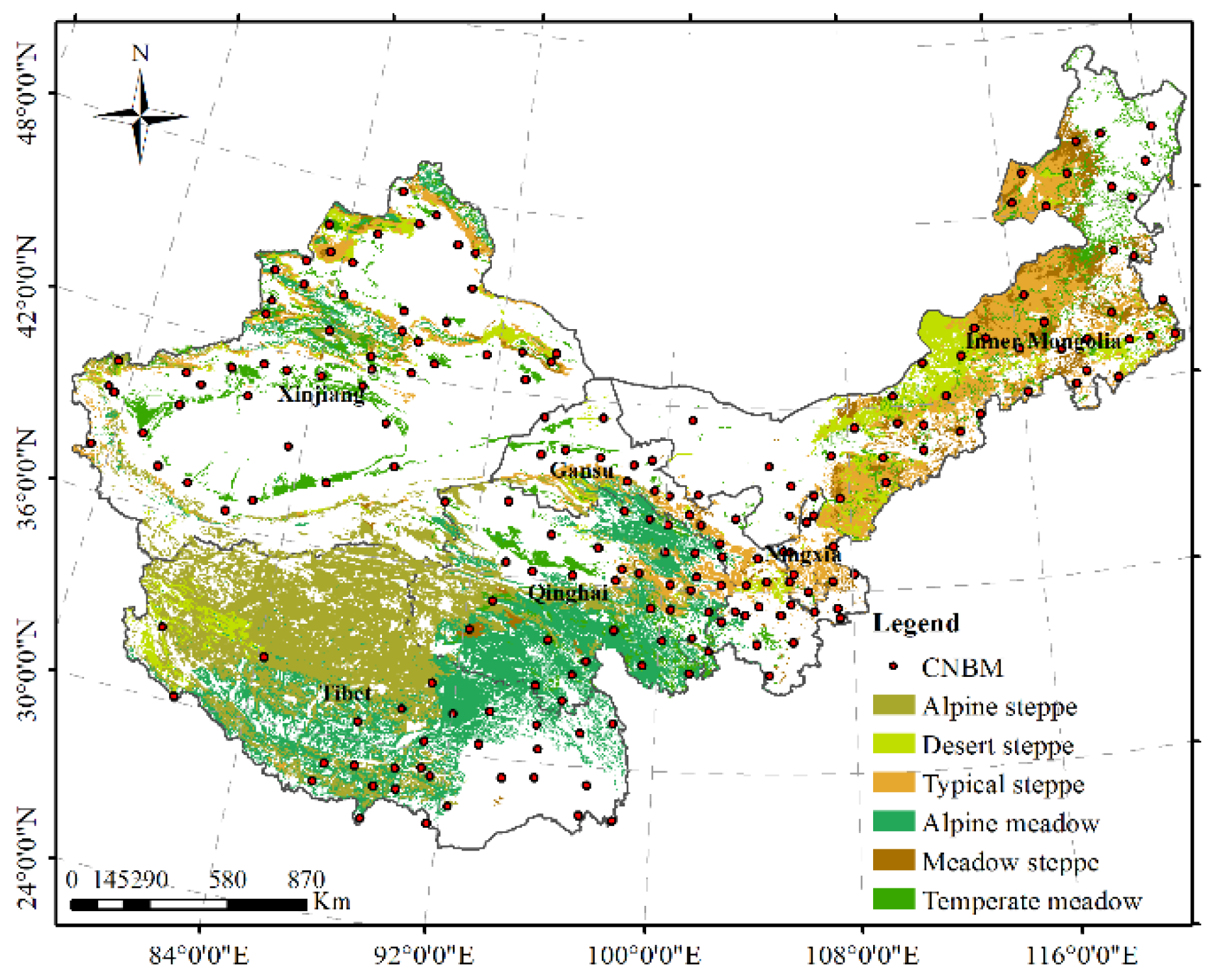
2.1. Study Area
2.2. NDVI Database
2.3. Climate Database
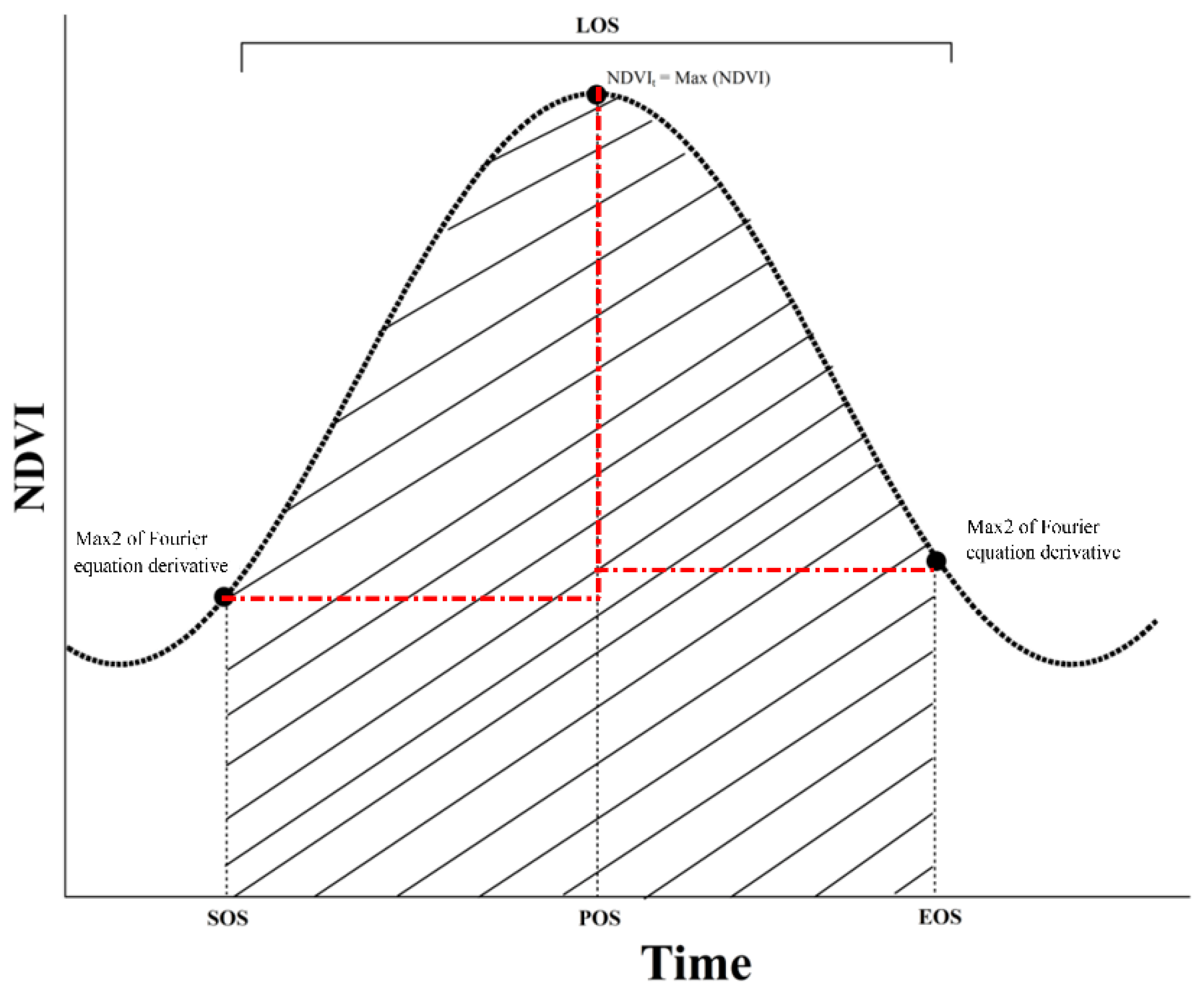
2.4. SOS, EOS, POS, and LOS Were Obtained with the Retrieval Method
2.5. Data Analyses
3. Results
3.1. Dynamics and Relationships of Plant Phenology, AMT, and AMP across China’s Grasslands
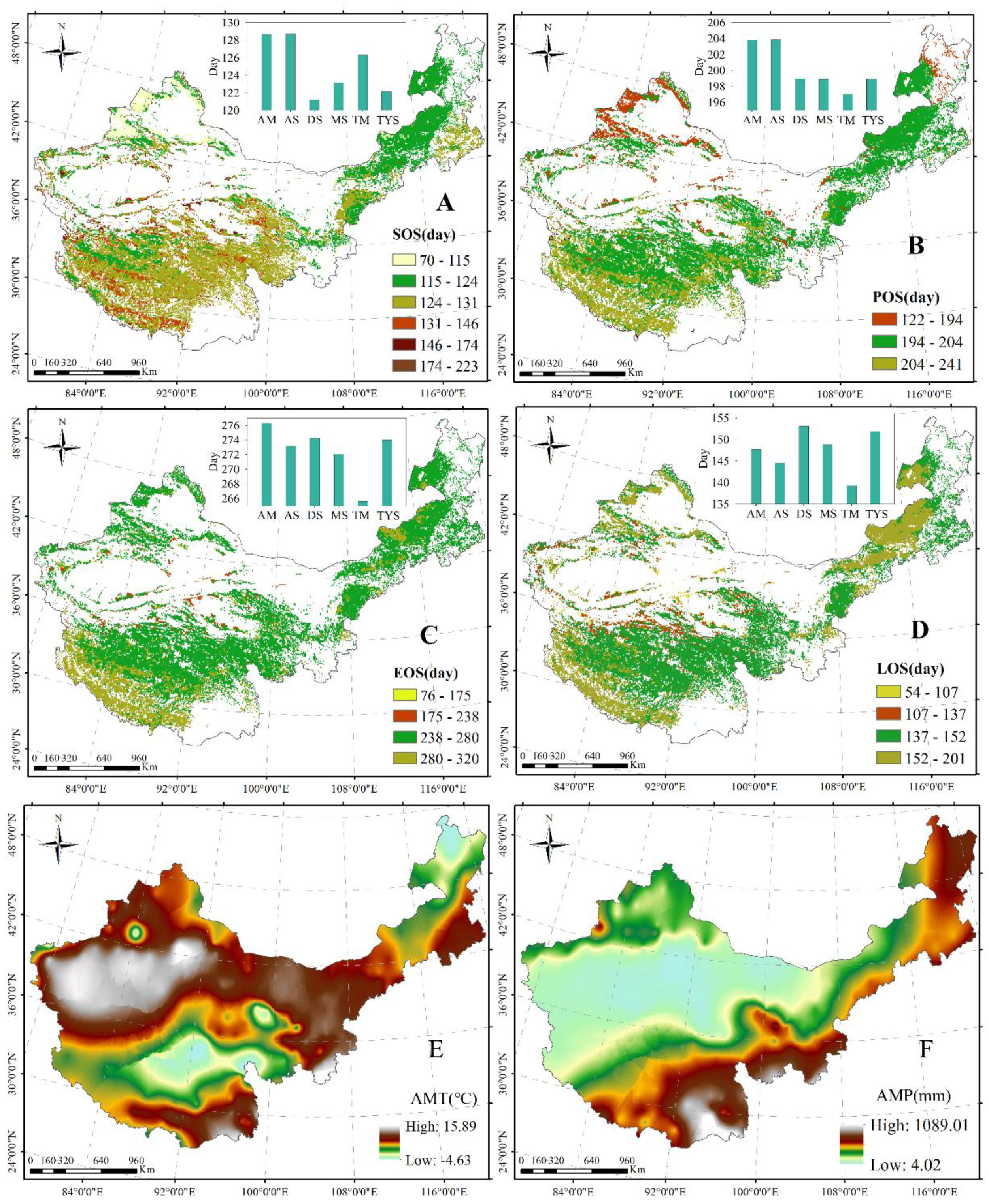
3.1.1. Spatial Distribution of Plant Phenology, AMT, and AMP in China’s Grasslands
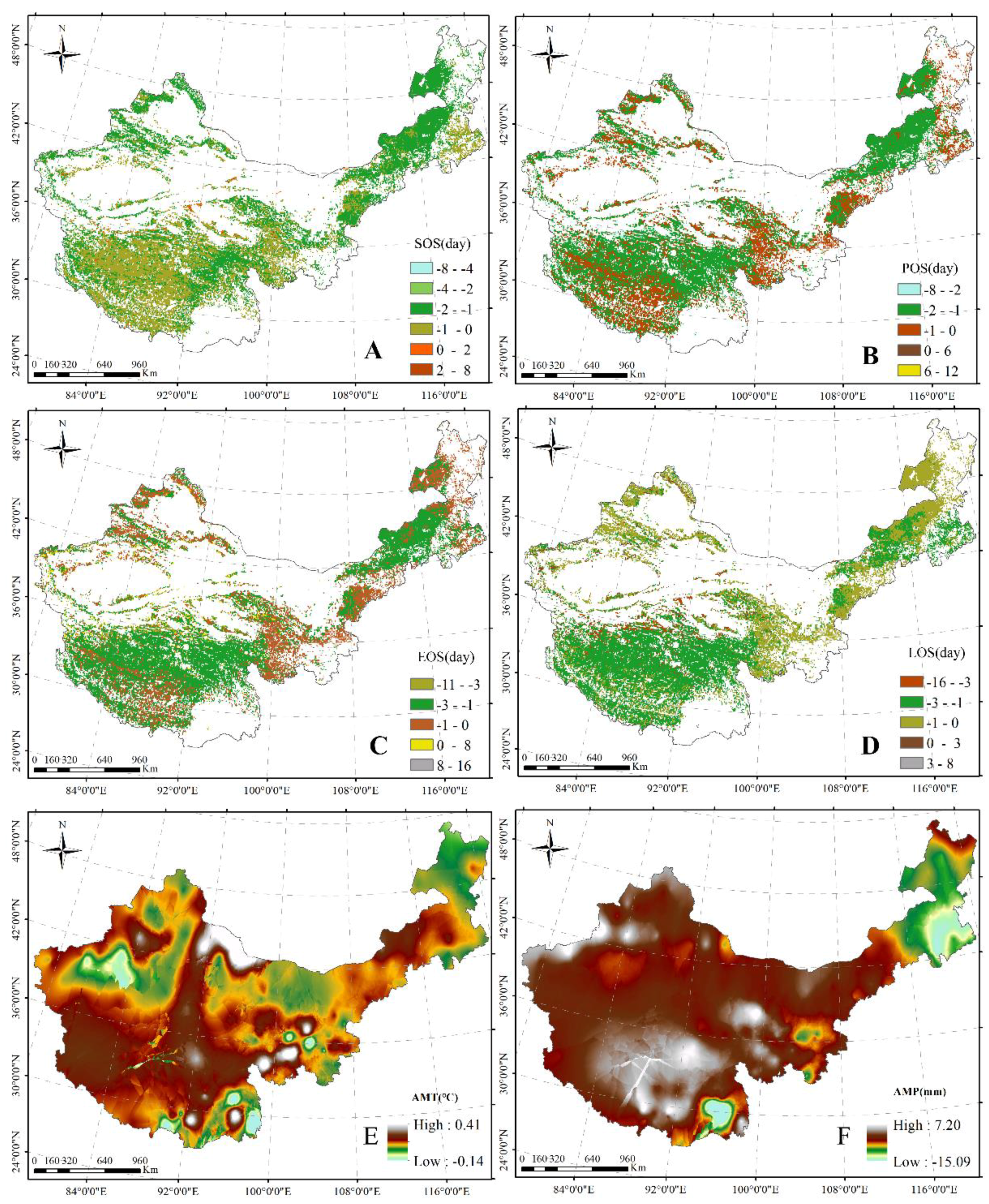
3.1.2. Dynamics of Plant Phenology, AMT, and AMP in China’s Grasslands
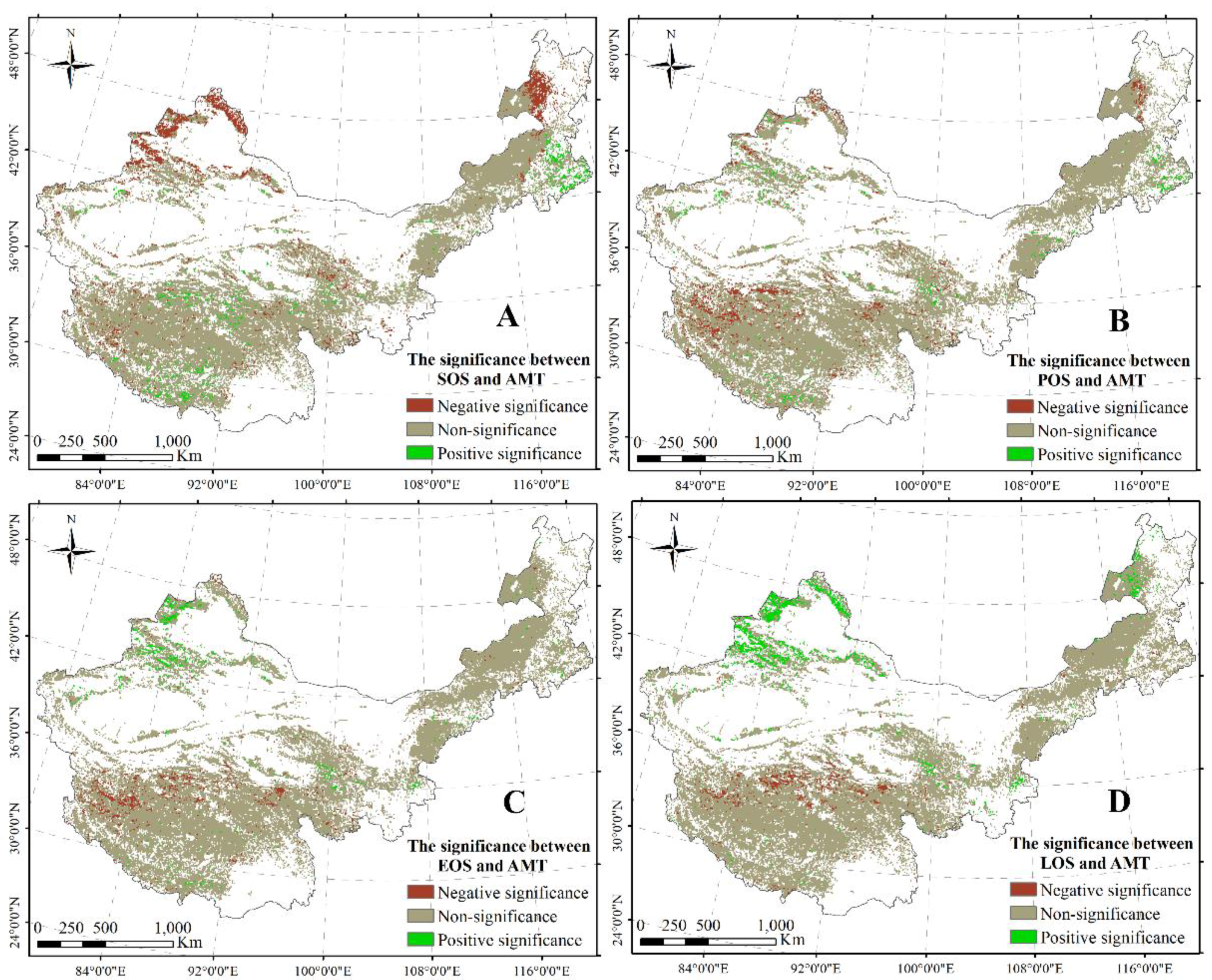
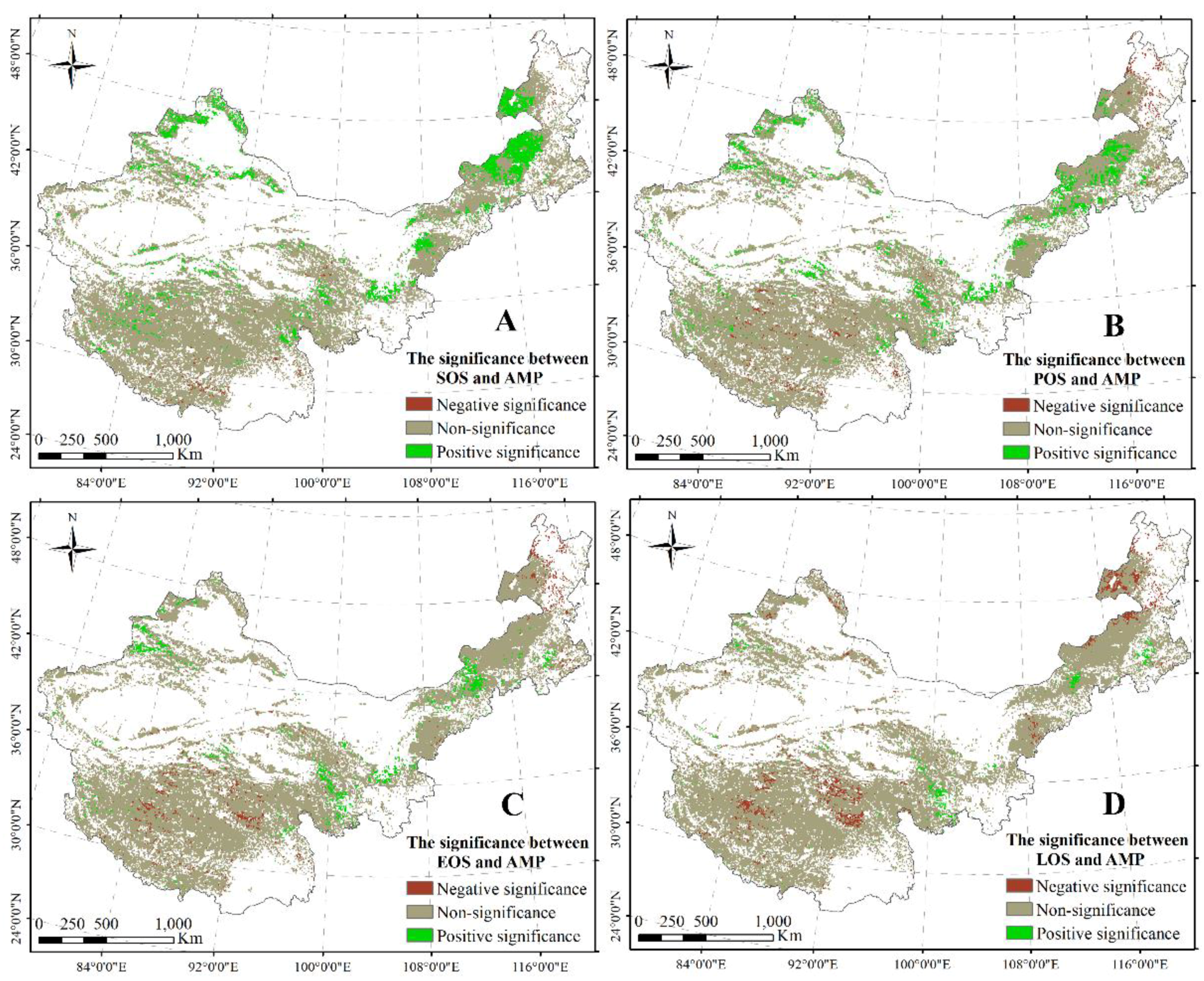
3.1.3. Responses of Plant Phenology to AMT and AMP across China’s Grasslands
3.2. Dynamics and Relationships in Plant Phenology, AMT, and AMP in Different Grassland Types
3.2.1. Dynamics of Plant Phenology in Different Grassland Types
3.2.2. Dynamics of AMT and AMP in Different Grassland Types
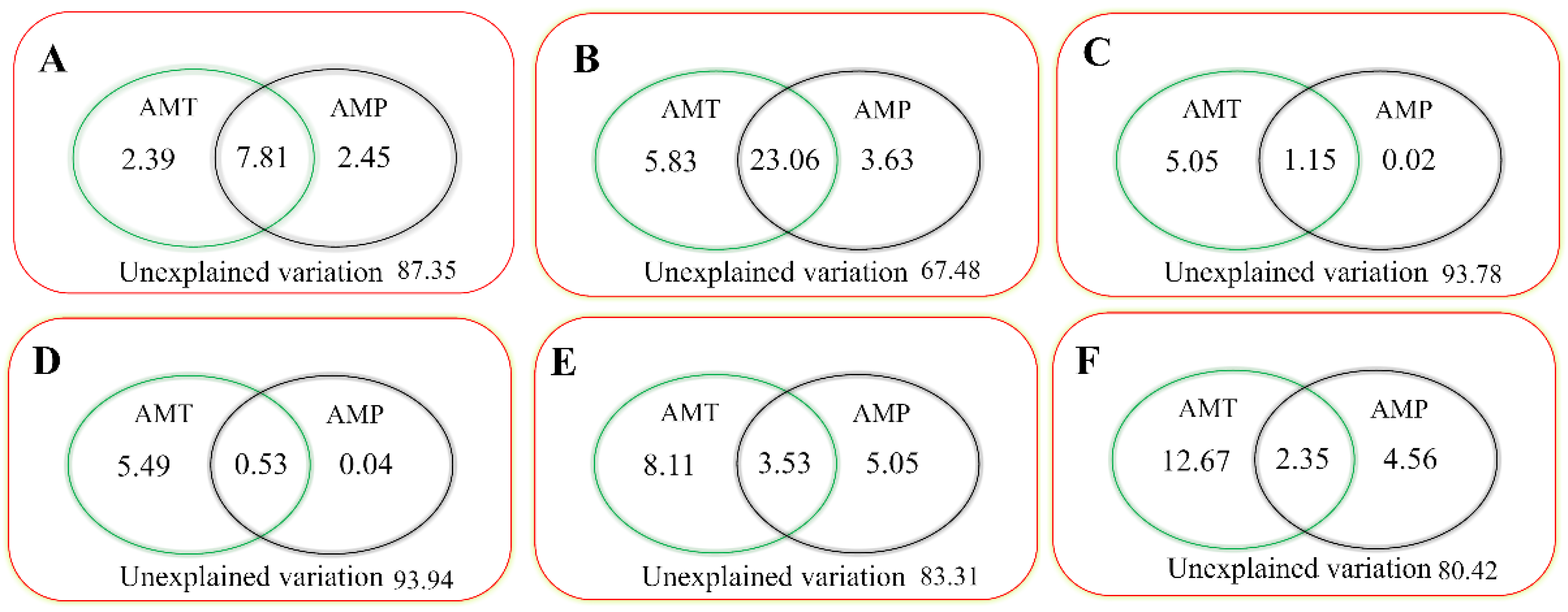
3.2.3. Responses of Plant Phenology to AMT and AMP in Different Grassland Types
4. Discussion
4.1. Spatial Distribution and Changes in Plant Phenology across China’s Grasslands
4.2. Control Mechanisms of Plant Phenology and Climate in Different Grasslands
4.3. Limitations of the Current Study
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- You, X.Z.; Meng, J.H.; Zhang, M.; Dong, T.F. Remote Sensing Based Detection of Crop Phenology for Agricultural Zones in China Using a New Threshold Method. Remote Sens. 2013, 5, 3190–3211. [Google Scholar] [CrossRef]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.L.; Cui, M.D.; Chen, A.P.; Wang, X.H.; Ciais, P.; Liu, J.; Tang, Y. Altitude and temperature dependence of change in the spring vegetation green-up date from 1982 to 2006 in the Qinghai-Xizang Plateau. Agric. For. Meteorol. 2011, 151, 1599–1608. [Google Scholar] [CrossRef]
- Ding, M.J.; Li, L.H.; Zhang, Y.L.; Sun, X.M.; Liu, L.S.; Gao, J.G.; Wang, Z.; Li, Y. Start of vegetation growing season on the Tibetan Plateau inferred from multiple methods based on GIMMS and SPOT NDVI data. J. Geogr. Sci. 2015, 25, 131–148. [Google Scholar] [CrossRef]
- Prevey, J.S.; Seastedt, T.R. Seasonality of precipitation interacts with exotic species to alter composition and phenology of a semi-arid grassland. J. Ecol. 2014, 102, 1549–1561. [Google Scholar] [CrossRef]
- Lesica, P.; Kittelson, P.M. Precipitation and temperature are associated with advanced flowering phenology in a semi-arid grassland. J. Arid Environ. 2010, 74, 1013–1017. [Google Scholar] [CrossRef]
- Bradley, A.V.; Gerard, F.F.; Barbier, N.; Weedon, G.P.; Anderson, L.O.; Huntingford, C.; Aragão, L.E.; Zelazowski, P.; Arai, E. Relationships between phenology, radiation and precipitation in the Amazon region. Glob. Change Biol. 2011, 17, 2245–2260. [Google Scholar] [CrossRef]
- Hmimina, G.; Dufrene, E.; Pontailler, J.Y.; Delpierre, N.; Aubinet, M.; Caquet, B.; De Grandcourt, A.; Burban, B.; Flechard, C.; Granier, A.; et al. Evaluation of the potential of MODIS satellite data to predict vegetation phenology in different biomes: An investigation using ground-based NDVI measurements. Remote Sens. Environ. 2013, 132, 145–158. [Google Scholar] [CrossRef]
- Leeuwen, W.J.D.V.; Davison, J.E.; Casady, G.M.; Marsh, S.E. Phenological Characterization of Desert Sky Island Vegetation Communities with Remotely Sensed and Climate Time Series Data. Remote Sens. 2010, 2, 388–415. [Google Scholar] [CrossRef]
- Shen, M.G.; Piao, S.L.; Cong, N.; Zhang, G.X.; Janssens, I.A. Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Glob. Change Biol. 2015, 21, 3647–3656. [Google Scholar] [CrossRef] [PubMed]
- Han, G.F.; Xu, J.H. Land Surface Phenology and Land Surface Temperature Changes Along an Urban-Rural Gradient in Yangtze River Delta, China. Environ. Manag. 2013, 52, 234–249. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Cao, R.Y.; Chen, J.; Rao, Y.H.; Tang, Y.H. Temperature sensitivity of spring vegetation phenology correlates to within-spring warming speed over the Northern Hemisphere. Ecol. Indic. 2015, 50, 62–68. [Google Scholar] [CrossRef]
- He, Z.B.; Du, J.; Zhao, W.Z.; Yang, J.J.; Chen, L.F.; Zhu, X.; Chang, X.; Liu, H. Assessing temperature sensitivity of subalpine shrub phenology in semi-arid mountain regions of China. Agric. For. Meteorol. 2015, 213, 42–52. [Google Scholar] [CrossRef]
- Brown, M.E.; Beurs, K.M.D.; Marshall, M. Global phenological response to climate change in crop areas using satellite remote sensing of vegetation, humidity and temperature over 26 years. Remote Sens. Environ. 2012, 126, 174–183. [Google Scholar] [CrossRef]
- Shen, M.G.; Zhang, G.X.; Cong, N.; Wang, S.P.; Kong, W.D.; Piao, S.L. Increasing altitudinal gradient of spring vegetation phenology during the last decade on the Qinghai-Tibetan Plateau. Agric. For. Meteorol. 2014, 189, 71–80. [Google Scholar] [CrossRef]
- Qiu, B.W.; Zhong, M.; Zeng, C.Y.; Tang, Z.H.; Chen, C.C. Effect of topography and accessibility on vegetation dynamic pattern in Mountain-hill Region. J. Mt. Sci. 2012, 9, 879–890. [Google Scholar] [CrossRef]
- Hwang, T.; Song, C.H.; Vose, J.M.; Band, L.E. Topography-mediated controls on local vegetation phenology estimated from MODIS vegetation index. Landsc. Ecol. 2011, 26, 541–556. [Google Scholar] [CrossRef]
- Brown, L.A.; Dash, J.; Ogutu, B.O.; Richardson, A.D. On the relationship between continuous measures of canopy greenness derived using near-surface remote sensing and satellite-derived vegetation products. Agric. For. Meteorol. 2017, 247, 280–292. [Google Scholar] [CrossRef]
- Hufkens, K.; Friedl, M.; Sonnentag, O.; Braswell, B.H.; Milliman, T.; Richardson, A.D. Linking near-surface and satellite remote sensing measurements of deciduous broadleaf forest phenology. Remote Sens. Environ. 2012, 117, 307–321. [Google Scholar] [CrossRef]
- Klosterman, S.T.; Hufkens, K.; Gray, J.M.; Melaas, E.; Sonnentag, O.; Lavine, I.; Mitchell, L.; Norman, R.; Friedl, M.A.; Richardson, A.D. Evaluating remote sensing of deciduous forest phenology at multiple spatial scales using PhenoCam imagery. Biogeosciences 2014, 11, 4305–4320. [Google Scholar] [CrossRef]
- Liu, Y.; Hill, M.J.; Zhang, X.; Wang, Z.; Richardson, A.D.; Hufkens, K.; Filippa, G.; Baldocchi, D.D.; Ma, S.; Verfaillie, J. Using data from Landsat, MODIS, VIIRS and PhenoCams to monitor the phenology of California oak/grass savanna and open grassland across spatial scales. Agric. For. Meteorol. 2017, 237, 311–325. [Google Scholar] [CrossRef]
- Melaas, E.K.; Sulla-Menashe, D.; Gray, J.M.; Black, T.A.; Morin, T.H.; Richardson, A.D.; Friedl, M.A. Multisite analysis of land surface phenology in North American temperate and boreal deciduous forests from Landsat. Remote Sens. Environ. 2016, 186, 452–464. [Google Scholar] [CrossRef]
- Robinson, N.P.; Allred, B.W.; Jones, M.O.; Moreno, A.; Kimball, J.S.; Naugle, D.E.; Erickson, T.A.; Richardson, A.D.; Thenkabail, P. A Dynamic Landsat Derived Normalized Difference Vegetation Index (NDVI) Product for the Conterminous United States. Remote Sens. 2017, 9, 863. [Google Scholar] [CrossRef]
- Benhadj, I.; Duchemin, B.; Maisongrande, P.; Simonneaux, V.; Khabba, S.; Chehbouni, A. Automatic unmixing of MODIS multi-temporal data for inter-annual monitoring of land use at a regional scale (Tensift, Morocco). Int. J. Remote Sens. 2012, 33, 1325–1348. [Google Scholar] [CrossRef]
- Cook, B.I.; Smith, T.M.; Mann, M.E. The North Atlantic Oscillation and regional phenology prediction over europe. Glob. Change Biol. 2005, 11, 919–926. [Google Scholar] [CrossRef]
- Arrogante-Funes, P.; Novillo, C.; Romero-Calcerrada, R.; Vázquez-Jiménez, R.; Ramos-Bernal, R. Relationship between MRPV Model Parameters from MISRL2 Land Surface Product and Land Covers: A Case Study within Mainland Spain. ISPRS Int. J. Geo-Inf. 2017, 6, 353. [Google Scholar] [CrossRef]
- Gressler, E.; Jochner, S.; Capdevielle-Vargas, R.M.; Morellato, L.P.C.; Menzel, A. Vertical variation in autumn leaf phenology of Fagus sylvatica L. in southern Germany. Agric. For. Meteorol. 2015, 201, 176–186. [Google Scholar] [CrossRef]
- Sharma, R.; Tateishi, R.; Hara, K. A Biophysical Image Compositing Technique for the Global-Scale Extraction and Mapping of Barren Lands. ISPRS Int. J. Geo-Inf. 2016, 5, 225. [Google Scholar] [CrossRef]
- Dahlke, C.; Loew, A.; Reick, C. Robust Identification of Global Greening Phase Patterns from Remote Sensing Vegetation Products. J. Clim. 2012, 25, 8289–8307. [Google Scholar] [CrossRef]
- Verhegghen, A.; Bontemps, S.; Defourny, P. A global NDVI and EVI reference data set for land-surface phenology using 13 years of daily SPOT-VEGETATION observations. Int. J. Remote Sens. 2014, 35, 2440–2471. [Google Scholar] [CrossRef]
- Ding, M.J.; Zhang, Y.L.; Sun, X.M.; Liu, L.S.; Wang, Z.F.; Bai, W.Q. Spatiotemporal variation in alpine grassland phenology in the Qinghai-Tibetan Plateau from 1999 to 2009. Chin. Sci. Bull. 2013, 58, 396–405. [Google Scholar] [CrossRef]
- Yu, H.Y.; Luedeling, E.; Xu, J.C. Winter and spring warming result in delayed spring phenology on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2010, 107, 22151–22156. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Tang, Y.; Chen, J.; Zhu, X.; Zheng, Y. Influences of temperature and precipitation before the growing season on spring phenology in grasslands of the central and eastern Qinghai-Tibetan Plateau. Agric. For. Meteorol. 2011, 151, 1711–1722. [Google Scholar] [CrossRef]
- Zhang, G.L; Zhang, Y.J; Dong, J.W; Xiao, X.M. Green-up dates in the Tibetan Plateau have continuously advanced from 1982 to 2011. Proc. Natl. Acad. Sci. USA 2013, 110, 4309–4314. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Cheng, G.W.; Li, W.P. Meta-analysis of relationships between environmental factors and aboveground biomass in the alpine grassland on the Tibetan Plateau. Biogeosciences 2013, 10, 1707–1715. [Google Scholar] [CrossRef]
- Sun, J.; Wang, H.M. Soil nitrogen and carbon determine the trade-off of the above- and below-ground biomass across alpine grasslands, Tibetan Plateau. Ecol. Indic. 2016, 60, 1070–1076. [Google Scholar] [CrossRef]
- Wang, C.Z.; Guo, H.D.; Zhang, L.; Liu, S.Y.; Qiu, Y.B.; Sun, Z.C. Assessing phenological change and climatic control of alpine grasslands in the Tibetan Plateau with MODIS time series. Int. J. Biometeorol. 2015, 59, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.L.; Li, Q.T.; Wang, C.Z.; Guo, H.D.; Zhang, L. Spatiotemporal Variations of Satellite-Derived Phenology in the Tibetan Plateau. In Proceedings of the 2012 IEEE International Geoscience and Remote Sensing Symposium (Igarss), Munich, Germany, 22–27 July 2012; pp. 6451–6454. [Google Scholar]
- Yu, H.Y.; Xu, J.C.; Okuto, E.; Luedeling, E. Seasonal Response of Grasslands to Climate Change on the Tibetan Plateau. PLoS ONE 2012, 7, e49230. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.H.; Zhou, Z.Y. Increasing contamination might have delayed spring phenology on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2011, 108, E94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.L.; Dong, J.W.; Zhang, Y.J.; Xiao, X.M. Reply to Shen et al.: No evidence to show nongrowing season NDVI affects spring phenology trend in the Tibetan Plateau over the last decade. Proc. Natl. Acad. Sci. USA 2013, 110, E2330–E2331. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.G.; Sun, Z.Z.; Wang, S.P.; Zhang, G.X.; Kong, W.D.; Chen, A.P.; Piao, S. No evidence of continuously advanced green-up dates in the Tibetan Plateau over the last decade. Proc. Natl. Acad. Sci. USA 2013, 110, E2329. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.W.; Zhang, G.L.; Zhang, Y.J.; Xiao, X.M. Reply to Wang et al.: Snow cover and air temperature affect the rate of changes in spring phenology in the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2013, 110, E2856–E2857. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Peng, S.S.; Lin, X.; Chang, J.F. Declining snow cover may affect spring phenological trend on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2013, 110, E2854–E2855. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fang, J.; Ji, C.; Datta, A.; Li, P.; Ma, W.; Mohammat, A.; Shen, H.; Hu, H.; Knapp, B.O.; et al. Stoichiometric shifts in surface soils over broad geographical scales: Evidence from China’s grasslands. Glob. Ecol. Biogeogr. 2014, 23, 947–955. [Google Scholar] [CrossRef]
- Sun, J.; Cheng, G.W.; Li, W.P.; Sha, Y.K.; Yang, Y.C. On the Variation of NDVI with the Principal Climatic Elements in the Tibetan Plateau. Remote Sens. 2013, 5, 1894–1911. [Google Scholar] [CrossRef]
- Almadani, S.; Ibrahim, E.; Abdelrahman, K.; Al-Bassam, A.; Al-Shmrani, A. Magnetic and seismic refraction survey for site investigation of an urban expansion site in Abha District, Southwest Saudi Arabia. Arab. J. Geosci. 2015, 8, 2299–2312. [Google Scholar] [CrossRef]
- Chen, Y.L.; Ding, J.Z.; Peng, Y.F.; Li, F.; Yang, G.B.; Liu, L.; Qin, S.Q.; Fang, K.; Yang, Y.H. Patterns and drivers of soil microbial communities in Tibetan alpine and global terrestrial ecosystems. J. Biogeogr. 2016, 43, 2027–2039. [Google Scholar] [CrossRef]
- Tang, W.; Zhou, T.; Sun, J.; Li, Y.; Li, W. Accelerated Urban Expansion in Lhasa City and the Implications for Sustainable Development in a Plateau City. Sustainability 2017, 9, 1499. [Google Scholar] [CrossRef]
- Myneni, R.B.; Keeling, C.D.; Tucker, C.J.; Asrar, G.; Nemani, R.R. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Sherry, R.A.; Zhou, X.; Gu, S.; Rd, A.J.; Schimel, D.S.; Verburg, P.S.; Wallace, L.L.; Luo, Y. Divergence of reproductive phenology under climate warming. Proc. Natl. Acad. Sci. USA 2007, 104, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Slayback, D.A.; Pinzon, J.E.; Los, S.O.; Tucker, C.J. Northern hemisphere photosynthetic trends 1982–99. Glob. Change Biol. 2010, 9, 1–15. [Google Scholar] [CrossRef]
- Shilong, P.; Fang, J.; Zhou, L.; Philippe, C.; Zhu, B. Variations in satellite-derived phenology in China’s temperate vegetation. Glob. Change Biol. 2010, 12, 672–685. [Google Scholar]
- Zhou, L.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res. Atmos. 2001, 106, 20069–20083. [Google Scholar] [CrossRef]
- Jeong, S.J.; Chang-Hoi, H.O.; Gim, H.J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Change Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Julien, Y.; Sobrino, J.A. Global land surface phenology trends from GIMMS database. Int. J. Remote Sens. 2009, 30, 3495–3513. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Dunne, J.A.; Harte, J.; Taylor, K.J. Subalpine meadow flowering phenology responses to climate change: integrating experimental and gradient methods. Ecol. Monogr. 2003, 73, 69–86. [Google Scholar] [CrossRef]
- Bradley, N.L.; Leopold, A.C.; Ross, J.; Huffaker, W. Phenological changes reflect climate change in Wisconsin. Proc. Natl. Acad. Sci. USA 1999, 96, 9701–9704. [Google Scholar] [CrossRef] [PubMed]
- Chmielewski, F.M.; Rötzer, T. Response of tree phenology to climate change across Europe. Agric. For. Meteorol. 2001, 108, 101–112. [Google Scholar] [CrossRef]
- CAMILLE. Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Glob. Change Biol. 2007, 13, 1860–1872. [Google Scholar]
- Menzel, A.; Fabian, P. Growing season extended in Europe. Nature 1999, 397, 659. [Google Scholar] [CrossRef]
- Wang, X.; Piao, S.; Ciais, P.; Li, J.; Friedlingstein, P.; Koven, C.; Chen, A. Spring temperature change and its implication in the change of vegetation growth in North America from 1982 to 2006. Proc. Natl. Acad. Sci. USA 2011, 108, 1240–1245. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Sun, J.; Wang, X. Plant coverage is more sensitive than species diversity in indicating the dynamics of the above-ground biomass along a precipitation gradient on the Tibetan Plateau. Ecol. Indic. 2018, 84, 507–514. [Google Scholar] [CrossRef]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Lucht, W.; Prentice, I.C.; Myneni, R.B.; Sitch, S.; Friedlingstein, P.; Cramer, W.; Bousquet, P.; Buermann, W.; Smith, B. Climatic Control of the High-Latitude Vegetation Greening Trend and Pinatubo Effect. Science 2002, 296, 1687–1689. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Zhou, L.; Chen, A. Effect of climate and CO2 changes on the greening of the Northern Hemisphere over the past two decades. Geophys. Res. Lett. 2006, 33, 265–288. [Google Scholar] [CrossRef]
- Braswell, B.H.; Schimel, D.S.; Linder, E.; Moore, B. The Response of Global Terrestrial Ecosystems to Interannual Temperature Variability. Science 1997, 278, 870–872. [Google Scholar] [CrossRef]
- Austin, A.T.; Vitousek, P.M. Nutrient dynamics on a precipitation gradient in Hawai’i. Oecologia 1998, 113, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Paruelo, J.M.; Lauenroth, W.K.; Burke, I.C.; Sala, O.E. Grassland Precipitation-Use Efficiency Varies Across a Resource Gradient. Ecosystems 1999, 2, 64–68. [Google Scholar] [CrossRef]
- Halse, S.A.; Scanlon, M.D.; Cocking, J.S.; Smith, M.J.; Kay, W.R. Factors affecting river health and its assessment over broad geographic ranges: The Western Australian experience. Environ. Monit. Assess. 2007, 134, 161–175. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator | Grasslands | y = ax + y0 | R2 | p | Mean (day) |
|---|---|---|---|---|---|
| SOS (day) | Alpine meadow | y = 0.0066x + 115.5370 | 0.0009 | 0.8856 | 128.6547 |
| Alpine steppe | y = 0.0805x − 32.0302 | 0.0488 | 0.2784 | 128.7045 | |
| Desert steppe | y = −0.0453x + 211.6438 | 0.0145 | 0.5586 | 121.2055 | |
| Meadow steppe | y = −0.1264x + 375.7233 | 0.2648 | 0.0072 ** | 123.1431 | |
| Temperate meadow | y = −0.0061x + 138.4402 | 0.0006 | 0.9081 | 126.3115 | |
| Typical steppe | y = −0.1559x + 433.6264 | 0.281 | 0.0053 ** | 122.1544 | |
| POS (day) | Alpine meadow | y = −0.0353x + 274.3249 | 0.0125 | 0.5866 | 203.8044 |
| Alpine steppe | y = −0.1289x + 461.3085 | 0.0604 | 0.2263 | 203.8654 | |
| Desert steppe | y = −0.0910x + 380.6512 | 0.0606 | 0.2255 | 198.9293 | |
| Meadow steppe | y = −0.0520x + 302.7789 | 0.0561 | 0.2442 | 198.9030 | |
| Temperate meadow | y = −0.0029x + 202.8453 | 0.0002 | 0.9437 | 197.0213 | |
| Typical steppe | y = −0.0564x + 311.6396 | 0.0524 | 0.2605 | 198.9553 | |
| EOS (day) | Alpine meadow | y = −0.0981x + 472.2792 | 0.0411 | 0.3207 | 276.2977 |
| Alpine steppe | y = −0.3772x + 1026.7458 | 0.1614 | 0.0419 ** | 273.2050 | |
| Desert steppe | y = −0.0896x + 453.2871 | 0.0372 | 0.3450 | 274.3001 | |
| Meadow steppe | y = 0.0737x + 124.8830 | 0.043 | 0.3095 | 272.0691 | |
| Temperate meadow | y = 0.0490x + 167.7751 | 0.0216 | 0.4741 | 265.6026 | |
| Typical steppe | y = 0.0648x + 144.5951 | 0.029 | 0.4053 | 273.9888 | |
| LOS (day) | Alpine meadow | y = −0.1047x + 356.7421 | 0.0546 | 0.2505 | 147.6430 |
| Alpine steppe | y = −0.4577x + 1058.7759 | 0.202 | 0.0213 ** | 144.5005 | |
| Desert steppe | y = −0.0443x + 241.6433 | 0.0061 | 0.7051 | 153.0946 | |
| Meadow steppe | y = 0.1866x − 223.9604 | 0.1807 | 0.0304 ** | 148.7567 | |
| Temperate meadow | y = 0.0550x + 29.3350 | 0.0128 | 0.5816 | 139.2911 | |
| Typical steppe | y = 0.2207x − 289.0314 | 0.1761 | 0.0329 ** | 151.8344 |
| Factor | Grasslands | Mean | Min | Max | Range | Std |
|---|---|---|---|---|---|---|
| AMT (°C/year) | Alpine steppe | 0.05 | 0.00 | 0.23 | 0.23 | 0.02 |
| Meadow steppe | 0.06 | −0.02 | 0.17 | 0.19 | 0.02 | |
| Typical steppe | 0.06 | −0.01 | 0.18 | 0.19 | 0.02 | |
| AMP (mm/year) | Alpine steppe | −1.75 | −7.76 | 4.29 | 12.05 | 2.39 |
| Meadow steppe | −0.60 | −7.93 | 3.98 | 11.91 | 1.83 | |
| Typical steppe | −0.72 | −7.50 | 3.98 | 11.48 | 2.53 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhou, T.; Peng, P. Phenology Response to Climatic Dynamic across China’s Grasslands from 1985 to 2010. ISPRS Int. J. Geo-Inf. 2018, 7, 290. https://doi.org/10.3390/ijgi7080290
Wang J, Zhou T, Peng P. Phenology Response to Climatic Dynamic across China’s Grasslands from 1985 to 2010. ISPRS International Journal of Geo-Information. 2018; 7(8):290. https://doi.org/10.3390/ijgi7080290
Chicago/Turabian StyleWang, Jun, Tiancai Zhou, and Peihao Peng. 2018. "Phenology Response to Climatic Dynamic across China’s Grasslands from 1985 to 2010" ISPRS International Journal of Geo-Information 7, no. 8: 290. https://doi.org/10.3390/ijgi7080290
APA StyleWang, J., Zhou, T., & Peng, P. (2018). Phenology Response to Climatic Dynamic across China’s Grasslands from 1985 to 2010. ISPRS International Journal of Geo-Information, 7(8), 290. https://doi.org/10.3390/ijgi7080290