Phenolic Content of Brown Algae (Pheophyceae) Species: Extraction, Identification, and Quantification

 ,
, 
 ,
,  , and
, and
Abstract
:1. Introduction
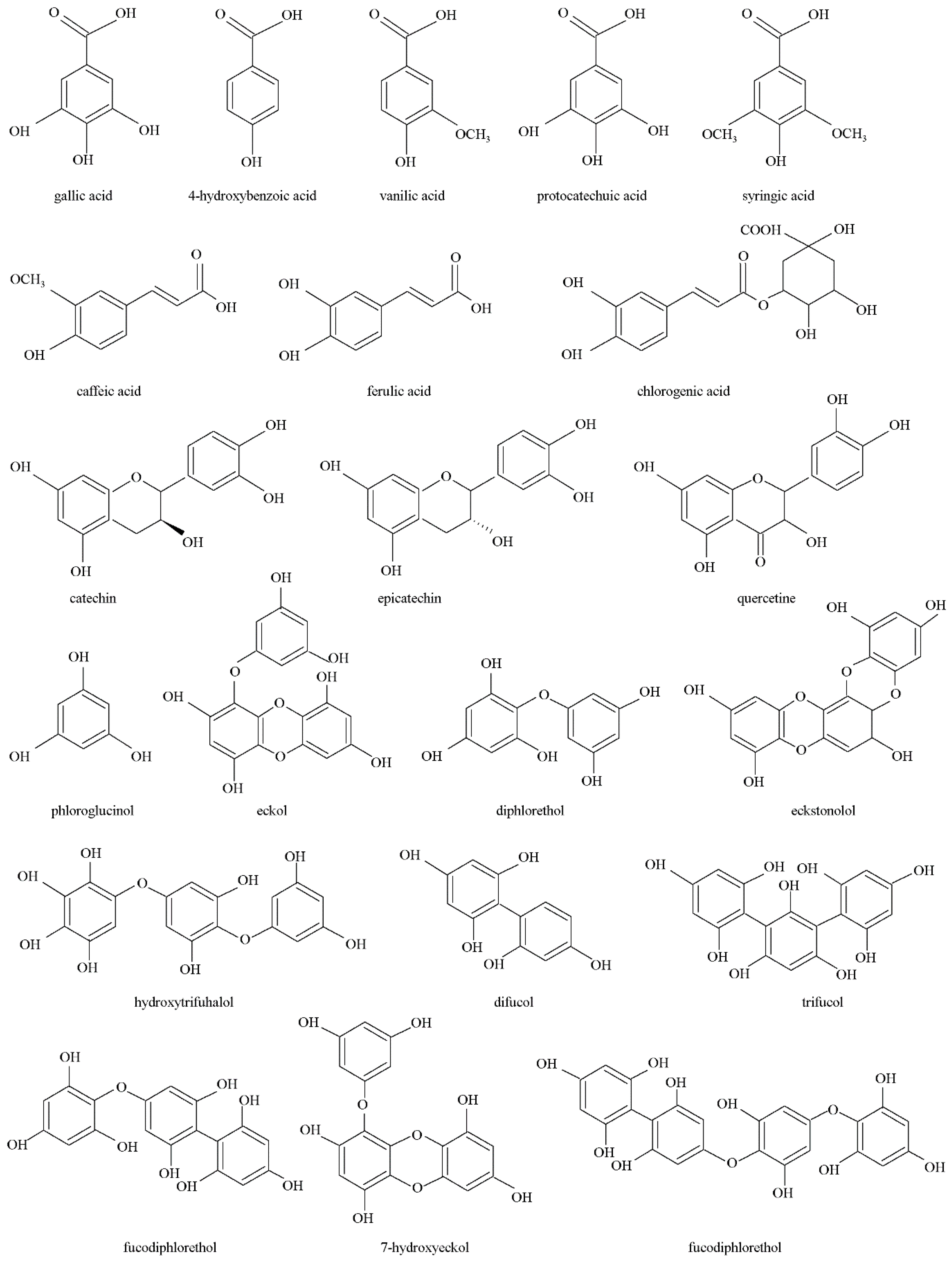
2. Brown Algae Phenolics
2.1. Extraction
2.2. Quantification and Identification
3. Conclusions
Funding
Conflicts of Interest
References
- Brodie, J.; Lewis, J. Unravelling the Algae, the Past, Present and the Future of Algal Systematics; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2007. [Google Scholar]
- Gupta, S.; Abu-Ghannam, N. Bioactive potential and possible health effects of edible brown seaweeds. Trends Food Sci. Tech. 2011, 22, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.X.; Wijesekara, I.; Li, Y.; Kim, S.K. Phlorotannins as bioactive agents from brown algae. Process Biochem. 2011, 46, 2219–2224. [Google Scholar] [CrossRef]
- Abdelhamid, A.; Jouini, M.; Amor, H.B.H.; Mzoughi, Z.; Dridi, M.; Said, R.B.; Bouraoui, A. Phytochemical analysis and evaluation of the antioxidant, anti-inflammatory, and antinociceptive potential of phlorotannin-rich fractions from three Mediterranean brown seaweeds. Mar. Biotechnol. 2018, 20, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef]
- Lezcano, V.; Fernández, C.; Parodi, E.R.; Morelli, S. Antitumor and antioxidant activity of the freshwater macroalga Cladophora surera. J. Appl. Phycol. 2018, 1–9. [Google Scholar] [CrossRef]
- Chakraborty, K.; Joseph, D. Antioxidant Potential and Phenolic Compounds of Brown Seaweeds Turbinaria conoides and Turbinaria ornata (Class: Phaeophyceae). J. Aquat. Food Prod. T. 2016, 25, 1249–1265. [Google Scholar] [CrossRef]
- FAO. The state of world fisheries and aquaculture. 2018. Available online: http://www.fao.org/3/i9540en/i9540en.pdf (accessed on 21 June 2019).
- Yuan, Y.V.; Bone, D.E.; Carrington, M.F. Antioxidant activity of dulse (Palmaria palmata) extract evaluated in vitro. Food Chem. 2005, 91, 485–494. [Google Scholar] [CrossRef]
- Cox, S.; Gupta, S.; Abu-Ghannam, N. Effect of different rehydration temperatures on the moisture, content of phenolic compounds, antioxidant capacity and textural properties of edible Irish brown seaweed. LWT-Food Sci. Technol. 2012, 47, 300–307. [Google Scholar] [CrossRef]
- Chakaborty, K.; Joseph, D.; Praveen, N.K. Antioxidant activities and phenolic contents of three red seaweeds (Division: Rhodophyta) harvested from the Gulf of Mannar of Peninsular India. J. Food Sci. Technol. 2015, 52, 1924–1935. [Google Scholar] [CrossRef] [PubMed]
- Dixit, D.C.; Reddy, C.R.K.; Balar, N.; Suthar, P.; Gajaria, T.; Gadhavi, D.K. Assessment of the Nutritive, Biochemical, Antioxidant and Antibacterial Potential of Eight Tropical Macro algae Along Kachchh Coast, India as Human Food Supplements. J. Aquat. Food Prod. T. 2018, 27, 61–79. [Google Scholar] [CrossRef]
- Liu, L.; Heinrich, M.; Myers, S.; Dworjanyn, S.A. Towards a better understanding of medicinal uses of the brown seaweed Sargassum in Traditional Chinese Medicine: A phytochemical and pharmacological review. J. Ethnopharmacol. 2012, 142, 591–619. [Google Scholar] [CrossRef]
- Herrero, M.; Mendiola, J.A.; Plaza, M.; Ibanez, E. Screening for Bioactive Compounds from Algae. In Advanced Biofuels and Bioproducts; Lee, J.W., Ed.; Springer: New York, NY, USA, 2013; pp. 833–872. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. Algaebase. World-Wide Electronic Publication. National University of Ireland, Galway. 2019. Available online: http://www.algaebase.org (accessed on 9 June 2019).
- Dawes, C. Seaweed in Health and Disease Prevention; Chapter 4 - Macroalgae Systematics; Fleurence, J., Levine, I., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 107–148. [Google Scholar] [CrossRef]
- Rindi, F.; Soler-Vila, A.; Guiry, M.D. Taxonomy of Marine Macroalgae Used as Sources of Bioactive Compounds. In Marine Bioactive Compounds; Hayes, M., Ed.; Springer: Boston, MA, USA, 2012. [Google Scholar]
- Alves, A.; Sousa, R.A.; Reis, R.L. A practical perspective on ulvan extracted from green algae. J. Appl. Phycol. 2013, 25, 407–424. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Domínguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef] [PubMed]
- Van den Hoek, C.; Mann, D.; Jahns, H.M. Algae, an Introduction to Phycology; Press Syndicate of the University of Cambridge: Cambridge, UK, 1995. [Google Scholar]
- Montero, L.; Herrero, M.; Ibanez, E.; Cifuentes, A. Separation and characterization of phlorotannins from brown algae Cystoseira abies-marina by comprehensive two-dimensional liquid chromatography. Electrophoresis 2014, 35, 1644–1651. [Google Scholar] [CrossRef] [PubMed]
- Bravo, L. Polyphenols: Chemistry, Dietary Sources, Metabolism, and Nutritional Significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 3rd ed.; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Shahidi, F.; Naczk, M. Phenolics in Food and Nutraceuticals; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, H. Algae as a source of biologically active ingredients for the formulation of functional foods and nutraceuticals. In Woodhead Publishing Series in Food Science, Technology and Nutrition, Functional Ingredients from Algae for Foods and Nutraceuticals; Dominguez, H., Ed.; Woodhead Publishing: Cambridge, MA, USA, 2013. [Google Scholar]
- Singh, I.P.; Sidana, J. Phlorotannins. In Woodhead Publishing Series in Food Science, Technology and Nutrition, Functional Ingredients from Algae for Foods and Nutraceuticals; Dominguez, H., Ed.; Woodhead Publishing: Cambridge, MA, USA, 2013. [Google Scholar]
- Dang, T.T.; Bowyer, M.C.; Van Altena, I.A.; Scarlett, C.J. Comparison of chemical profile and antioxidant properties of the brown algae. Int. J. Food Sci. Tech. 2018, 53, 174–181. [Google Scholar] [CrossRef]
- Koivikko, R.; Loponen, J.; Honkanen, T.; Jormalainen, V. Contents of soluble, cell-wall-bound and exuded phlorotannins in the brown alga Fucus vesiculosus, with implications on their ecological functions. J. Chem.Ecol. 2005, 31, 195–212. [Google Scholar] [CrossRef]
- Schoenwaelder, M.E.A. The occurrence and cellular significance of physodes in brown algae. Phycologia 2002, 41, 125–139. [Google Scholar] [CrossRef]
- Ank, G.; Da Costa Paradas, W.; Amado-Filho, G.M.; Perez Da Gama, B.A.; Pereira, R.C. Within-thallus variation on phlorotannin contents and physodes amount in Stypopodium zonale (Phaeophyceae). Pan-Am. J. Aquat. Sci. 2014, 9, 1–7. [Google Scholar]
- Li, Y.; Qian, Z.J.; Ryu, B.; Lee, S.H.; Kim, M.M.; Kim, S.K. Chemical components and its antioxidant properties in vitro: An edible marine brown alga Ecklonia cava. Bioorg. Med. Chem. 2009, 17, 1963–1973. [Google Scholar] [CrossRef] [PubMed]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Koivikko, R. Brown Algal Phlorotannins: Improving and Applying Chemical Methods. PhD Thesis, University of Turku, Turku, Finland, 22 February 2008. [Google Scholar]
- Li, Y.; Fu, X.; Duan, D.; Liu, X.; Xu, J.; Gao, X. Extraction and Identification of Phlorotannins from the Brown Alga, Sargassum fusiforme (Harvey) Setchell. Mar. Drugs 2017, 15, 49. [Google Scholar] [CrossRef] [PubMed]
- Thomas, N.V.; Kim, S.K. Review: Potential pharmacological applications of polyphenolic derivatives from marine brown algae. Environ. Toxicol. Pharmacol. 2011, 32, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Le Lann, K.; Surget, G.; Couteau, C.; Coiffard, L.; Cerantola, S.; Gaillard, F.; Larnicol, M.; Zubia, M.; Guerard, F.; Poupart, N.; et al. Sunscreen, antioxidant, and bactericide capacities of phlorotannins from the brown macroalga Halidrys siliquosa. J. Appl. Phycol. 2016, 28, 3547–3559. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, K.; Maneesh, A.; Makkar, F. Antioxidant Activity of Brown Seaweeds. J. Aquat. Food Prod. Technol. 2017, 26, 406–419. [Google Scholar] [CrossRef]
- Fellah, F.; Louaileche, H.; Dehbi-Zebboudj, A.; Touati, N. Seasonal variations in the phenolic compound content and antioxidant activities of three selected species of seaweeds from Tiskerth islet, Bejaia, Algeria. J. Mater. Environ. Sci. 2017, 8, 4451–4456. [Google Scholar] [CrossRef]
- Dang, T.T.; Bowyer, M.C.; Van Altena, I.A.; Scarlett, C.J. Optimum conditions of microwave-assisted extraction for phenolic compounds and antioxidant capacity of the brown alga Sargassum vestitum. Sep. Sci. Technol. 2017, 53, 1711–1723. [Google Scholar] [CrossRef]
- Dang, T.T.; Van Vuong, Q.; Schreider, M.J.; Bowyer, M.C.; Van Altena, I.A.; Scarlett, C.J. Optimisation of ultrasound-assisted extraction conditions for phenolic content and antioxidant activities of the alga Hormosira banksii using response surface methodology. J. Appl. Phycol. 2017, 29, 3161–3173. [Google Scholar] [CrossRef]
- Magnusson, M.; Yuen, A.K.; Zhang, R.; Wright, J.T.; Taylor, R.B.; Maschmeyer, T.; de Nys, R. A comparative assessment of microwave assisted (MAE) and conventional solid-liquid (SLE) techniques for the extraction of phloroglucinol from brown seaweed. Algal Res. 2017, 23, 28–36. [Google Scholar] [CrossRef]
- Júnior, S.Q.; Carneiro, V.H.A.; Fontenelle, T.P.C.; de Sousa Chaves, L.; Mesquita, J.X.; de Brito, T.V.; Prudêncio, R.S.; de Oliveira, J.S.; Rolim Medeiros, J.V.; Saboia Aragão, K.; et al. Antioxidant and anti-inflammatory activities of methanol extract and its fractions from the brown seaweed Spatoglossum schroederi. J. Appl. Phycol. 2015, 27, 2367–2376. [Google Scholar] [CrossRef]
- Liu, N.; Fu, X.; Duan, D.; Xu, J.; Gao, X.; Zhao, L. Evaluation of bioactivity of phenolic compounds from the brown seaweed of Sargassum fusiforme and development of their stable emulsion. J. Appl. Phycol. 2018, 30, 1955–1970. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave assisted extraction of phenolic compounds from four economic brown macroalgae species and evaluation of their antioxidant activities and inhibitory effects on α-amylase, α-glucosidase, pancreatic lipase and tyrosinase. Food Res. Int. 2018, 113, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Hermund, D.B.; Heung, S.Y.; Raagaard Thomsen, B.; Akoh, C.C.; Jacobsen, C. Improving oxidative stability of skin-care emulsions with antioxidant extracts from brown alga Fucus vesiculosus. J. Am. Oil Chem.’ Soc. 2018, 95, 1509–1520. [Google Scholar] [CrossRef]
- Zaragozá, M.C.; López, D.; Sáiz, M.P.; Poquet, M.; Pérez, J.; Puig-Parellada, P.; Màrmol, F.; Simonetti, P.; Gardana, C.; Lerat, Y.; et al. Toxicity and antioxidant activity in vitro and in vivo of two Fucus vesiculosus extracts. J. Agr. Food Chem. 2008, 56, 7773–7780. [Google Scholar] [CrossRef] [PubMed]
- Zubia, M.; Fabre, M.S.; Kerjean, V.; Le Lann, K.; Stiger-Pouvreau, V.; Fauchon, M.; Deslandes, E. Antioxidant and antitumoural activities of some Phaeophyta from Brittany coasts. Food Chem. 2009, 116, 693–701. [Google Scholar] [CrossRef]
- Del Pilar Sánchez-Camargo, A.; Montero, L.; Stiger-Pouvreau, V.; Tanniou, A.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Considerations on the use of enzyme-assisted extraction in combination with pressurized liquids to recover bioactive compounds from algae. Food Chem. 2016, 192, 67–74. [Google Scholar] [CrossRef]
- Neto, R.T.; Marçal, C.; Queir, A.S.; Abreu, H.; Silva, A.M.S.; Cardoso, S.M. Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissimi as functional ingredient. Int. J. Mol. Sci. 2018, 19, 2987. [Google Scholar] [CrossRef]
- Wang, T.; Jonsdottir, R.; Ólafsdóttir, G. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Zahra, R.; Mehrnaz, M.; Farzaneh, V.; Kohzad, S. Antioxidant activity of extract from a brown alga Sargassum boveanum. Afr. J. Biotechnol. 2007, 6, 2740–2745. [Google Scholar] [CrossRef]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish seaweeds. Int. Food Res. J. 2010, 17, 205–220. [Google Scholar] [CrossRef]
- Rajauria, G.; Jaiswal, A.K.; Abu Ghannam, N.; Gupta, S. Effect of hydrothermal processing on colour, antioxidant and free radical scavenging capacities of edible Irish brown seaweeds. Int. J. Food Sci. Tech. 2010, 45, 2485–2493. [Google Scholar] [CrossRef]
- O’Sullivan, A.M.; O’Callaghan, Y.C.; O’Grady, M.N.; Queguineur, B.; Hanniffy, D.; Troy, D.J.; Kerry, J.P.; O’Brien, N.M. In vitro and cellular antioxidant activities of seaweed extracts prepared from five brown seaweeds harvested in spring from the west coast of Ireland. Food Chem. 2011, 126, 1064–1070. [Google Scholar] [CrossRef]
- Tierney, M.S.; Smyth, T.J.; Hayes, M.; Soler-Vila, A.; Croft, A.K.; Brunton, N. Influence of pressurised liquid extraction and solid–liquid extraction methods on the phenolic content and antioxidant activities of Irish macroalgae. Int. J. Food Sci. Tech. 2013, 48, 860–869. [Google Scholar] [CrossRef]
- Heffernan, N.; Smyth, T.J.; Soler-Villa, A.; Fitzgerald, R.J.; Brunton, N.P. Phenolic content and antioxidant activity of fractions obtained from selected Irish macroalgae species (Laminaria digitata, Fucus serratus, Gracilaria gracilis and Codium fragile). J. Appl. Phycol. 2015, 27, 519–530. [Google Scholar] [CrossRef]
- Kadam, S.U.; O’Donnell, C.P.; Rai, D.K.; Hossain, M.B.; Burgess, C.M.; Walsh, D.; Tiwari, B.K. Laminarin from Irish brown seaweeds Ascophyllum nodosum and Laminaria hyperborea: Ultrasound assisted extraction, characterization and bioactivity. Mar. Drugs 2015, 13, 4270–4280. [Google Scholar] [CrossRef] [PubMed]
- Kuda, T.; Kunii, T.; Goto, H.; Suzuki, T.; Yano, T. Varieties of antioxidant and antibacterial properties of Ecklonia stolonifera and Ecklonia kurome products harvested and processed in the Noto peninsula, Japan. Food Chem. 2007, 103, 900–905. [Google Scholar] [CrossRef]
- Iwai, K. Antidiabetic and antioxidant effects of polyphenols in brown alga Ecklonia stolonifera in genetically diabetic KK-A y mice. Plant. Food Hum. Nutr. 2008, 63, 163. [Google Scholar] [CrossRef] [PubMed]
- Airanthi, M.K.; Hosokawa, M.; Miyashita, K. Comparative antioxidant activity of edible Japanese brown seaweeds. J. Food Sci. 2011, 76, C104–C111. [Google Scholar] [CrossRef]
- Machu, L.; Misurcova, L.; Vavra Ambrozova, J.; Orsavova, J.; Mlcek, J.; Sochor, J.; Jurikova, T. Phenolic content and antioxidant capacity in algal food products. Molecules 2015, 20, 1118–1133. [Google Scholar] [CrossRef]
- Shin, H.C.; Hwang, H.J.; Kang, K.J.; Lee, B.H. An antioxidative and antiinflammatory agent for potential treatment of osteoarthritis from Ecklonia cava. Arch. Pharm. Res. 2006, 29, 165. [Google Scholar] [CrossRef] [PubMed]
- Khaled, N.; Hiba, M.; Asma, C. Antioxidant and antifungal activities of Padina pavonica and Sargassum vulgare from the Lebanese Mediterranean coast. Adv. Environ. Biol. 2012, 6, 42–48. [Google Scholar]
- Chew, Y.L.; Lim, Y.Y.; Omar, M.; Khoo, K.S. Antioxidant activity of three edible seaweeds from two areas in South East Asia. LWT-Food Sci. Technol. 2008, 41, 1067–1072. [Google Scholar] [CrossRef]
- Matanjun, P.; Mohamed, S.; Mustapha, N.M.; Muhammad, K.; Ming, C.H. Antioxidant activities and phenolics content of eight species of seaweeds from north Borneo. J. Appl. Phycol. 2008, 20, 367. [Google Scholar] [CrossRef]
- Fu, C.W.F.; Ho, C.W.; Yong, W.T.L.; Abas, F.; Tan, C.P. Effects of phenolic antioxidants extraction from four selected seaweeds obtained from Sabah. PeerJ PrePrints 2015, 3, e1249v1. [Google Scholar] [CrossRef]
- Senevirathne, M.; Kim, S.H.; Siriwardhana, N.; Ha, J.H.; Lee, K.W.; Jeon, Y.J. Antioxidant potential of ecklonia cava on reactive oxygen species scavenging, metal chelating, reducing power and lipid peroxidation inhibition. Food Sci. Technol. Int. 2006, 12, 27–38. [Google Scholar] [CrossRef]
- Chkhikvishvili, I.D.; Ramazanov, Z.M. Phenolic substances of brown algae and their antioxidant activity. Appl. Biochem. Micro. 2000, 36, 289–291. [Google Scholar] [CrossRef]
- Plaza, M.; Amigo-Benavent, M.; Del Castillo, M.D.; Ibáñez, E.; Herrero, M. Facts about the formation of new antioxidants in natural samples after subcritical water extraction. Food Res. Int. 2010, 43, 2341–2348. [Google Scholar] [CrossRef] [Green Version]
- López, A.; Rico, M.; Rivero, A.; de Tangil, M.S. The effects of solvents on the phenolic contents and antioxidant activity of Stypocaulon scoparium algae extracts. Food Chem. 2011, 125, 1104–1109. [Google Scholar] [CrossRef]
- Belda, M.; Sanchez, D.; Bover, E.; Prieto, B.; Padrón, C.; Cejalvo, D.; Lloris, J.M. Extraction of polyphenols in Himanthalia elongata and determination by high performance liquid chromatography with diode array detector prior to its potential use against oxidative stress. J. Chromatogr. B 2016, 1033, 334–341. [Google Scholar] [CrossRef]
- Otero, P.; López-Martínez, M.I.; García-Risco, M. Application of pressurized liquid extraction (PLE) to obtain bioactive fatty acids and phenols from Laminaria ochroleuca collected in Galicia (NW Spain). J. Pharm. Biomed. Anal. 2019, 5, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; AbdElgawad, H.; Castellano, I.; Selim, S.; Beemster, G.T.S.; Asard, H.; Buia, M.C.; Palumbo, A. Effects of ocean acidification on the levels of primary and secondary metabolites in the brown macroalga Sargassum vulgare at different time scales. Sci. Total Environ. 2018, 643, 946. [Google Scholar] [CrossRef] [PubMed]
- Boonchum, W.; Peerapornpisal, Y.; Kanjanapothi, D.; Pekkoh, J.; Pumas, C.; Jamjai, U.; Amornlerdpison, D.; Noiraksar, T.; Vacharapiyasophon, P. Antioxidant activity of some seaweed from the Gulf of Thailand. Int. J. Agric. Biol. 2011, 13, 95–99. [Google Scholar]
- Rattaya, S.; Benjakul, S.; Prodpran, T. Extraction, antioxidative, and antimicrobial activities of brown seaweed extracts, Turbinaria ornata and Sargassum polycystum, grown in Thailand. Int. Aquat. Res. 2015, 7, 1–16. [Google Scholar] [CrossRef]
- Aoun, Z.B.; Said, R.B.; Farhat, F. Anti-inflammatory, antioxidant and antimicrobial activities of aqueous and organic extracts from Dictyopteris membranacea. Bot. Mar. 2010, 53, 259–264. [Google Scholar] [CrossRef]
- Mhadhebi, L.; Laroche-Clary, A.; Robert, J.; Bouraoui, A. Anti-inflammatory, anti-proliferative and anti-oxidant activities of organic extracts from the Mediterranean seaweed, Cystoseira crinita. Afr. J. Biotechnol. 2011, 10, 16682–16690. [Google Scholar] [CrossRef]
- Chakraborty, K.; Praveen, N.K.; Vijayan, K.K.; Rao, G.S. Evaluation of phenolic contents and antioxidant activities of brown seaweeds belonging to Turbinaria spp. (Phaeophyta, Sargassaceae) collected from Gulf of Mannar. Asian Pac. J. Trop. Biomed. 2013, 3, 8–16. [Google Scholar] [CrossRef]
- Ibañez, E.; Herrero, M.; Mendiola, J.A.; Castro-Puyana, M. Extraction and characterization of bioactive compounds with health benefits from marine resources: Macro and micro algae, cyanobacteria, and invertebrates. In Marine bioactive compounds; Hayes, M., Ed.; Springer: Boston, MA, USA, 2012; pp. 55–98. [Google Scholar]
- Machmudah, S.; Wahyudiono, K.H.; Goto, M. Supercritical fluids extraction of valuable compounds from algae: Future perspectives and challenges. Eng. J. 2018, 22, 13–30. [Google Scholar] [CrossRef]
- Cikoš, A.-M.; Jokić, S.; Šubarić, D.; Jerković, I. Overview on the application of modern methods for the extraction of bioactive compounds from marine macroalgae. Mar. Drugs 2018, 16, 348. [Google Scholar] [CrossRef]
- Grosso, C.; Valentão, P.; Ferreres, F.; Andrade, P.B. Alternative and efficient extraction methods for marine-derived compounds. Mar. Drugs 2015, 13, 3182–3230. [Google Scholar] [CrossRef]
- Setyaningsih, W.; Saputro, I.E.; Palma, M.; Barroso, C.G. Stability of 40 phenolic compounds during ultrasound-assisted extractions (UAE). In AIP Conference Proceedings (Vol. 1755, No. 1, p. 080009); AIP Publishing: Mellvile, NY, USA, 2016. [Google Scholar]
- Wijesinghe, W.A.J.P.; Won-Woo, L.; Young-Mog, K.; Young-Tae, K.; Se-Kwon, K.; Byong-Tae, J.; Jin-Soo, K.; Min-Soo, H.; Won-Kyo, J.; Ahn, G.; et al. Value-added fermentation of Ecklonia cava processing by-product and its antioxidant effect. J. Appl. Phycol. 2012, 24, 201–209. [Google Scholar] [CrossRef]


{kind=link}
| Algae Species | Collecting Location | Collecting Period | Solvent | Plant: Solvent Ratio | Extraction Mode | Total Phenolic Content | Reference |
|---|---|---|---|---|---|---|---|
| Dictyota ciliolata | Spain | - | 70% MeOH | - | - | 0.08% dw | [69] |
| Dictyota dichotoma | France | May 2007 | DCM, MeOH | - | PLE, 75 °C, 1500 psi | 18.8 mg PGE/g | [48] |
| India | December | MeOH | 1:5 | 48 h, RT | 0.02 mg PGE/g | [12] | |
| Malaysia | - | MeOH | 1:10 | 72 h, RT | 35.23 mg PGE/g | [66] | |
| Dictyota sp. 1 | Spain | - | 70% MeOH | - | - | 0.03% dw | [69] |
| Dictyota sp. 2 | Spain | - | 70% MeOH | - | - | 0.001% dw | [69] |
| Dictyopteris polypodioides | Tunisia | July 2006 | W | 1:10 | 24 h, 4 °C | 33.8 mg GAE/g dw | [77] |
| Chl | 84.96 mg GAE/g dw | ||||||
| EtOAc | 60.96 mg GAE/g dw | ||||||
| Spain | - | 70% MeOH | - | - | 0.09% dw | [69] | |
| Lobophora variegata | Spain | - | 70% MeOH | - | - | 1.20% dw | [69] |
| Padina antillarum | Malaysia | October–December 2005 | 20% MeOH | 1: 50 | Shaking | 20.4 mg GAE/g dw | [65] |
| 50% MeOH | 24.3 mg GAE/g dw | ||||||
| MeOH | 12.4 mg GAE/g dw | ||||||
| Padina pavonica | Tunisia | July 2015 | 50% EtOH | 1:5 | Shaking, 30 min, 50 °C | 7.06 mg PGE/g dw | [4] |
| Spain | - | 70% MeOH | - | - | 0.69% dw | [69] | |
| Lebanon | 2010 | MeOH | 1:10 | Shaking, 72 h, RT | 10.55 mg GAE/g | [64] | |
| Padina sp. | Australia | March 2016 | 70% EtOH | 1:50 | UAE, 1 h, 150 W, 30 °C | 124.65 mg GAE/g | [28] |
| Malaysia | - | MeOH | 1:10 | 72 h, RT | 33.11 mg PGE/g | [66] | |
| Spatoglossum schroederi | Brasil | December 2011– January 2012 | MeOH | 1:20 | Shaking, 24 h, RT | 11.75 mg GAE/g | [43] |
| ACE | 14.10 mg GAE/g | ||||||
| Chl | 6.84 mg GAE/g | ||||||
| Stypopodium zonale | Spain | - | 70% MeOH | - | - | 1.22% dw | [69] |
| Zonaria tournefortii | Algeria | December 2013, June and September 2014 | W | 1:100 | Shaking, 1 h, RT | 0.78 mg GAE/g | [39] |
| Spain | - | 70% MeOH | - | - | 1.06% dw | [69] |
| Algae Species | Collecting Location | Collecting Period | Solvent | Plant: Solvent Ratio | Extraction Mode | Total Phenolic Content | Reference |
|---|---|---|---|---|---|---|---|
| Anthophycus longifolius | India | - | MeOH | 1:6 | 3 h, 45 °C | 41.11 mg GAE/g | [38] |
| Ascophyllum nodosum | Ireland | - | 60% MeOH | 1:15 | 3 h, 40 °C | 4.5 mg GAE/g dw | [55] |
| Iceland | May 2007 | W | 1:20 | Shaking, 24 h, RT | 138 mg PGE/g | [51] | |
| 70% ACE | 159 mg PGE/g | ||||||
| Ireland | May 2014 | W | 1:20 | UAE, 35.61 Wcm−2, 15 min | 0.16 mg PGE/g dw | [58] | |
| 0.1 M HCl | 0.13 mg PGE/g dw | ||||||
| W | Shaking, 150 min, 70 °C | 0.17 mg PGE/g dw | |||||
| 0.1 M HCl | 0.11 mg PGE/g dw | ||||||
| March 2010 | W | 1:20 | Shaking, RT | 70.48 mg PGE/g | [56] | ||
| 80% EtOH | 1:10 | 66.26 mg PGE/g | |||||
| 80% ACE | 1:10 | 155.95 mg PGE/g | |||||
| W | - | PLE, 120 °C, 1500 psi | 93.44 mg PGE/g | ||||
| 80% EtOH | - | PLE, 100 °C, 1000 psi | 101.30 mg PGE/g | ||||
| 80% ACE | - | PLE, 60 °C, 1000 psi | 127.37 mg PGE/g | ||||
| China | - | 70% MeOH | 1:10 | MAE, 110 °C, 2.45 GHz, 15 min | 1.40 mg GAE/g dw | [82] | |
| 70% MeOH | 1:10 | Shaking, 4 h, RT | 0.51 mg GAE/g dw | ||||
| Bifurcaria bifurcata | France | September 2007 | MeOH | - | PLE, 75 °C, 1500 psi | 9.6 mg PGE/g | [48] |
| Carpophyllum flexuosum | New Zealand | July 2014 | MeOH, ACE | 1:8 | Shaking, RT | 86 mg PGE/g dw | [42] |
| W | 1:30 | MAE | 114 mg PGE/g dw | ||||
| Carpophyllum plumosum | New Zealand | July 2014 | MeOH, ACE | 1:8 | Shaking, RT | 75 mg PGE/g dw | [42] |
| Cystophora subfarcinata | Australia | November 2014 | MeOH, ACE | 1:8 | Shaking, RT | 22 mg PGE/g dw | [42] |
| Cystoseira abies-marina | Spain | - | W | 1:10 | PLE, 20 min, 100 °C, 1500 psi | 6.81 mg GAE/g | [70] |
| PLE, 20 min, 200 °C, 1500 psi | 48.09 mg GAE/g | ||||||
| Cystoseira compressa | Spain | - | 70% MeOH | - | - | 4.83% dw | [69] |
| Cystoseira crinita | Tunisia | June 2007 | Chl | - | 24 h | 402.44 mg GAE/g dw | [78] |
| EtOAc | 406.22 mg GAE/g dw | ||||||
| MeOH | 261.53 mg GAE/g dw | ||||||
| Cystoseira foeniculacea | Spain | - | 70% MeOH | - | - | 2.16% dw | [69] |
| Cystoseira sedoides | Tunisia | July 2015 | 50% EtOH | 1:5 | Shaking, 30 min, 50 °C | 26.45 mg PGE/g dw | [4] |
| Cystoseira tamariscifolia | France | April–June 2007 | DCM, MeOH | - | PLE, 75 °C, 1500 psi | 10.91 mg PGE/g | [48] |
| Fucus ceranoides | France | March 2006 | DCM, MeOH | - | PLE, 75 °C, 1500 psi | 54.7 mg PGE/g | [48] |
| Fucus serratus | France | March 2007 | DCM, MeOH | - | PLE, 75 °C, 1500 psi | 28.2 mg PGE/g | [48] |
| Iceland | March 2007 | W | 1:20 | Shaking, 24 h, RT | 169 mg PGE/g | [51] | |
| 70% ACE | 240 mg PGE/g | ||||||
| Ireland | - | 60% MeOH | 1:15 | 3 h, 40 °C | 4.0 mg GAE/g dw | [55] | |
| 2011 | W (cold) | 1:10 | Shaking, 24 h, RT | 81.93 mg GAE/g | [57] | ||
| 80% EtOH | 1:20 | 75.96 mg GAE/g | |||||
| 80% MeOH | 1:20 | 80.70 mg GAE/g | |||||
| W (hot) | 1:10 | Shaking, 24 h, 60 °C | 79.49 mg GAE/g | ||||
| Fucus spiralis | Spain | - | 70% MeOH | - | - | 2.17% dw | [69] |
| Ireland | May 2010 | W | 1:20 | Shaking, RT | 90.79 mg PGE/g | [56] | |
| 80% EtOH | 1:10 | Shaking, RT | 124.30 mg PGE/g | ||||
| 80% ACE | 1:10 | Shaking, RT | 204.40 mg PGE/g | ||||
| W | - | PLE, 120 °C, 1500 psi | 130.58 mg PGE/g | ||||
| 80% EtOH | - | PLE, 100 °C, 1000 psi | 142.81 mg PGE/g | ||||
| 80% ACE | 1:20 | PLE, 60 °C, 1000 psi | 187.55 mg PGE/g | ||||
| Fucus vesiculosus | France | - | 30–35% EtOH | 1:10 | Shaking, 4 h, RT | 277 mg PGE/g | [47] |
| 50–75% EtOH | Shaking, 2 h, RT | 163 mg PGE/g | |||||
| Ireland | - | 60% MeOH | 1:15 | 3 h, 40 °C | 2.5 mg GAE/g dw | [55] | |
| Iceland | March 2007 | W | 1:20 | Shaking, 24 h, RT | 17.6 mg PGE/g | [51] | |
| 70% ACE | 24.2 mg PGE/g | ||||||
| Denmark | September | W | 1:20 | Shaking, 24 h, 125 rpm, 20 °C | 134 mg GAE/g dw | [46] | |
| 80% EtOH | 165 mg GAE/g dw | ||||||
| Portugal | January 2016 | W | 1:20 | RT, 24 h | 14.8 mg GAE/g | [50] | |
| W | 1:20 | 90 °C, 30 min | 17.4 mg GAE/g | ||||
| 80% EtOH | 1:20 | RT, 24 h | 56.6 mg GAE/g | ||||
| 70% ACE | 1:20 | RT, 24 h | 39.1 mg GAE/g | ||||
| Halidrys siliquosa | France | May 2007 | DCM, MeOH | - | PLE, 75 °C, 1500 psi | 16.0 mg PGE/g | [48] |
| Hormosira banksii | Australia | March 2016 | 70% EtOH | 1:50 | Shaking, 12 h, 30 °C | 16.21 mg GAE/g | [41] |
| UAE, 60 min, 150 W, 30 °C | 23.12 mg GAE/g | [41] | |||||
| 158.82 mg GAE/g | [28] | ||||||
| Himanthalia elongata | Spain | - | 60% MeOH | 1:5 | Shaking, 2 h, 60 °C | 5.48 mg GAE/g | [72] |
| Ireland | June, September 2008 | 60% MeOH | 1:10 | Shaking, 2 h, 40 °C | 151.33 mg GAE/g | [53] | |
| - | 60% MeOH | 1:5 | Shaking, 2 h, 40 °C | 52.50 mg GAE/g | [54] | ||
| Kjellmaniella crassifolia | Japan | - | MeOH | 1:10 | 24 h, dark, RT | 9.90 mg PCE/g | [61] |
| EtOH | 8.01 mg PCE/g | ||||||
| ACE | 11.75 mg PCE/g | ||||||
| Chl | 17.82 mg PCE/g | ||||||
| EtOAc | 14.12 mg PCE/g | ||||||
| Hex | 16.67 mg PCE/g | ||||||
| Pelvetia canaliculata | Ireland | - | 60% MeOH | 1:15 | 3 h, 40 °C | 4.0 mg GAE/g dw | [55] |
| May 2010 | W | 1:20 | Shaking, RT | 41.13 mg PGE/g | [56] | ||
| 80% EtOH | 1:10 | Shaking, RT | 40.07 mg PGE/g | ||||
| 80% ACE | 1:10 | Shaking, RT | 168.82 mg PGE/g | ||||
| W | - | PLE, 120 °C, 1500 psi | 73.65 mg PGE/g | ||||
| 80% EtOH | - | PLE, 100 °C, 1000 psi | 61.89 mg PGE/g | ||||
| 80% ACE | - | PLE, 60 °C, 1000 psi | 68.24 mg PGE/g | ||||
| Saragassum aquifolium | Australia | March 2016 | 70% EtOH | 1:50 | UAE, 1 h, 150 W, 30 °C | 67.78 mg GAE/g | [28] |
| November 2014 | MeOH, ACE | 1:8 | Shaking, RT | 2 mg PGE/g dw | [42] | ||
| Sargassum binderi | Thailand | May 2007 | W | 1:10 | Shaking, 72 h, 25 °C | 0.267 mg GAE/g | [75] |
| EtOH | 0.063 mg GAE/g | ||||||
| Sargassum boveanum | Iran | May 2006 | W | 1:100 | 15 min, 121 °C | 17.0 mg CE/g dw | [52] |
| EtOH | 1:12.5 | Shaken, 4.5 h, 37 °C | 0.9 mg CE/g dw | ||||
| Sargassum desfontainesii | Spain | - | 70% MeOH | - | - | 1.68% dw | [69] |
| Sargassum flavicans | Australia | August 2014 | MeOH, ACE | 1:8 | Shaking, RT | 15 mg PGE/g dw | [42] |
| Sargassum fusiforme | China Japan | April-June 2014 | 30% EtOH | 1:50 | Shaking, 30 min, 25 °C | 880 mg PTC/g | [44] |
| - | 100% MeOH | 1:100 | Shaking, 24 h, 23 °C | 6.0 mg GAE/g | [62] | ||
| 70% ACE | 1:100 | Shaking, 30 min, 30 °C | 13.1 mg GAE/g | ||||
| 80% MeOH | 1:100 | Shaking, 1 h, 70 °C | 9.5 mg GAE/g | ||||
| MeOH:W:HAc (30:69:1) | 1:100 | Shaking, 50 min, 70 °C | 26.9 mg GAE/g | ||||
| W | 1:100 | Shaking, 10 min, 80°C | 34.5 mg GAE/g | ||||
| Sargassum furcatum | Spain | - | 70% MeOH | - | - | 2.97% dw | [69] |
| Sargassum horneri | Japan | 2007 | MeOH | 1:10 | 24 h, dark, RT | 9.90 mg PCE/g | [61] |
| Sargassum linearifolium | Australia | March 2016 | 70% EtOH | 1: 50 | UAE, 1 h, 150 W, 30 °C | 47.06 mg GAE/g | [28] |
| Sargassum muticum | Spain | - | W | 1:10 | PLE, 20 min, 100 °C, 1500 psi | 10.73 mg GAE/g | [70] |
| PLE, 20 min, 200 °C, 1500 psi | 58.67 mg GAE/g | ||||||
| France | July 2011 | W | - | PLE, 20 min, 1500 psi, 50 °C | 29.61 mg GAE/g | [49] | |
| EtOH | 94.20 mg GAE/g | ||||||
| 50% EtOH | 58.10 mg GAE/g | ||||||
| W | PLE, 20 min, 1500 psi, 125 °C | 52.26 mg GAE/g | |||||
| EtOH | 93.84 mg GAE/g | ||||||
| 50% EtOH | 76.62 mg GAE/g | ||||||
| W | PLE, 20 min, 1500 psi, 200 °C | 69.31 mg GAE/g | |||||
| EtOH | 93.16 mg GAE/g | ||||||
| 50% EtOH | 82.22 mg GAE/g | ||||||
| Saragassum polycystum | |||||||
| Sargassum plagiophyllum | India | - | MeOH | 1:6 | 3 h, 45 °C | 7.48 mg GAE/g | |
| Sargassum podacanthum | Australia | March 2016 | 70% EtOH | 1: 50 | UAE, 1 h, 150 W, 30 °C | 48.13 mg GAE/g | [28] |
| Sargassum polycystum | Thailand | - | MeOH | 1:30 | Shaking, 30 min, RT | 0.59 mg CE/g dw | [76] |
| EtOH | 0.32 mg CE/g dw | ||||||
| Malaysia | - | 50% EtOH | 1:10 | Shaking, 2 h, 65 °C | 0.37 mg GAE/g dw | [67] | |
| Malaysia | - | MeOH | 1:10 | 72 h, RT | 45.16 mg PGE/g | [66] | |
| India | - | MeOH | 1:6 | 3 h, 45 °C | 8.71 mg GAE/g | [38] | |
| Sargassum vestitum | Australia | March 2016 | 70% EtOH | 1: 50 | MAE | 58.20 mg GAE/g | [40] |
| Shaking, 12 h, 30 °C | 40.31 mg GAE/g | ||||||
| UAE, 1 h, 30 °C | 48.45 mg GAE/g | ||||||
| UAE, 1 h, 150 W, 30 °C | 141.91 mg GAE/g | [28] | |||||
| Sargassum vulgare | Spain | - | W | 1:10 | PLE, 20 min, 100 °C, 1500 psi | 26.43 mg GAE/g | [70] |
| PLE, 20 min, 200 °C, 1500 psi | 70.86 mg GAE/g | ||||||
| Lebanon | 2010 | MeOH | 1:10 | Shaking, 72 h, RT | 12.71 mg GAE/g | [64] | |
| Sirophysalis trinodis | Australia | August 2014 | MeOH, ACE | 1:8 | Shaking, RT | 25 mg PGE/g dw | [42] |
| India | December | MeOH | 1:5 | 48 h, RT | 0.15 mg PGE/g | [12] | |
| 80% EtOH | 62 mg PGE/g | ||||||
| Stephanocystis hakodatensis | Japan | 2007 | MeOH | 1:10 | 24 h, dark, RT | 31.33 mg PCE/g | [61] |
| Turbinaria conoides | Thailand | May 2007 | W | 1:10 | Shaking, 72 h, 25 °C | 1.12 mg GAE/g | [75] |
| EtOH | 0.09 mg GAE/g | ||||||
| India | - | MeOH | 1:10 | 3 h, 40–45 °C | 16.64 mg GAE/g | [79] | |
| Hex | 19.26 mg GAE/g | ||||||
| DCM | 51.47 mg GAE/g | ||||||
| EtOAc | 105.97 mg GAE/g | ||||||
| Turbinaria ornata | India | - | MeOH | 1:10 | 3 h, 40–45 °C | 3.42 mg GAE/g | [79] |
| Hex | 1.07 mg GAE/g | ||||||
| DCM | 12.72 mg GAE/g | ||||||
| EtOAc | 69.63 mg GAE/g | ||||||
| Thailand | - | MeOH | 1:30 | Shaking, 30 min, RT | 2.18 mg CE/g dw | [76] | |
| EtOH | 1.25 mg CE/g dw | ||||||
| Zonaria tournefortii | Algeria | December 2013, June and September 2014 | W | 1:100 | Shaking, 1 h, RT | 0.78 mg GAE/g | [39] |
| Spain | - | 70% MeOH | - | - | 1.06% dw | [69] |
| Algae Species | Collecting Location | Collecting Period | Solvent | Plant: Solvent Ratio | Extraction Mode | Total Phenolic Content | Reference |
|---|---|---|---|---|---|---|---|
| Laria crassifolia | Japan | 2007 | MeOH | 1:10 | 24 h, dark, RT | 5.94 mg PCE/g | [61] |
| EtOH | 7.21 mg PCE/g | ||||||
| ACE | 12.44 mg PCE/g | ||||||
| Chl | 12.93 mg PCE/g | ||||||
| EtOAc | 16.11 mg PCE/g | ||||||
| Hex | 21.13 mg PCE/g | ||||||
| Alaria esculenta | France | May 2007 | DCM, MeOH | - | PLE, 75 °C, 1500 psi | 20.3 mg PGE/g | [48] |
| Ecklonia cava | South Korea | May 2004 | MeOH | 1:50 | 72 h, 25 °C | 82.99 mg GAE/g dw | [68] |
| Korea | - | W | - | Boiling | 20.7 mg PGE/g | [63] | |
| 30% EtOH | - | 2 h, 50 °C | 45.3 mg PGE/g | ||||
| 80% EtOH | 1:50 | Shaking, 2 h, RT | 28.96 mg GAE/g | [84] | |||
| Ecklonia kurome | Japan | March 2006 | W | 1:20 | Shaking, 20 min, 75 °C | 97 mg PGE/g | [59] |
| 80% EtOH | 62 mg PGE/g | ||||||
| Ecklonia stolonifera | Japan | March 2006 | W | 1:20 | Shaking, 20 min, 75 °C | 74 mg PGE/g | [59] |
| 80% EtOH | Shaking, 20 min, 75 °C | 73 mg PGE/g | |||||
| - | W | 1:4 | - | 82.13 mg GAE/g | [60] | ||
| EtOH | - | 303.0 mg GAE/g | |||||
| Ecklonia bicyclis | Japan | - | MeOH | 1:100 | Shaking, 24 h, 23 °C | 9.5 mg GAE/g | [62] |
| 70% ACE | Shaking, 30 min, 30 °C | 84.1 mg GAE/g | |||||
| 80% MeOH | Shaking, 1 h, 70 °C | 143.2 mg GAE/g | |||||
| MeOH:W:HAc (30:69:1) | Shaking, 50 min, 70 °C | 192.8 mg GAE/g | |||||
| W | Shaking, 10 min, 80 °C | 192.6 mg GAE/g | |||||
| MeOH | 1:10 | 24 h, dark, RT | 7.77 mg PCE/g | [61] | |||
| EtOH | 1.87 mg PCE/g | ||||||
| ACE | 4.11 mg PCE/g | ||||||
| Chl | 14.41 mg PCE/g | ||||||
| EtOAc | 9.13 mg PCE/g | ||||||
| Hex | 16.75 mg PCE/g | ||||||
| Laminaria digitata | Ireland | June, September 2008 | 60% MeOH | 1:10 | Shaking, 2 h, 40 °C | 37.66 mg GAE/g | [53] |
| 2011 | W (cold) | 1:10 | Shaking, 24 h, RT | 2.24 mg GAE/g | [57] | ||
| 80% EtOH | 1:20 | 1.39 mg GAE/g | |||||
| 80% MeOH | 1:20 | 2.93 mg GAE/g | |||||
| W (hot) | 1:10 | Shaking, 24 h, 60 °C | 5.06 mg GAE/g | ||||
| - | 60% MeOH | 1:5 | Shaking, 2 h, 40 °C | 35.80 mg GAE/g | [54] | ||
| Laminaria hyperborea | Ireland | - | 60% MeOH | 1:15 | 3 h, 40 °C | 1.5 mg GAE/g dw | [55] |
| May 2014 | W | 1:20 | US, 35.61 Wcm−2, 15 min | 0.37 mg PGE/g dw | [58] | ||
| 0.1 M HCl | 0.34 mg PGE/g dw | ||||||
| W | Shaking, 2.5 h, 70 °C | 0.36 mg PGE/g dw | |||||
| 0.1 M HCl | 0.35 mg PGE/g dw | ||||||
| Laminaria ochroleuca | Spain | August 2017 | Hex | 1:20 | PLE, 10 min, 1450 bar, 80 °C | 6 mg GAE/g | [73] |
| EtOH | PLE, 10 min, 1450 bar, 160 °C | 83 mg GAE/g | |||||
| 50% EtOH | PLE, 10 min, 1450 bar, 160 °C | 173.65 mg GAE/g | |||||
| Lessonia nigrecens | China | - | 70% MeOH | 1:10 | MAE, 110°C, 2.45 GHz, 15 min | 1.07 mg GAE/g dw | [45] |
| 70% MeOH | 1:10 | Shaking, 4 h, RT | 0.78 mg GAE/g dw | ||||
| Lessonia trabeculata | China | - | 70% MeOH | 1:10 | MAE, 110 °C, 2.45 GHz, 15 min | 0.74 mg GAE/g dw | [45] |
| 70% MeOH | 1:10 | Shaking, 4 h, RT | 0.50 mg GAE/g dw | ||||
| Saccharina latissima | France | April 2016 | W | 1:20 | RT, 24 h | 4.7 mg GAE/g | [50] |
| W | 1:20 | 90 °C, 30 min | 7.8 mg GAE/g | ||||
| 80% EtOH | 1:20 | RT, 24 h | 1.9 mg GAE/g | ||||
| 70% ACE | 1:20 | RT, 24 h | 5.2 mg GAE/g | ||||
| Ireland | June, September 2008 | 60% MeOH | 1:10 | Shaking, 2 h, 40 °C | 66.75 mg GAE/g | [53] | |
| - | 1:5 | Shaking, 2 h, 40 °C | 43.50 mg GAE/g | [54] | |||
| Saccharina japonica | Japan | - | 100% MeOH | 1:100 | Shaking, 24 h, 23 °C | 0.7 mg GAE/g | [62] |
| 70% ACE | Shaking, 30 min, 30 °C | 8.8 mg GAE/g | |||||
| 80% MeOH | Shaking, 1 h, 70 °C | 14.9 mg GAE/g | |||||
| MeOH:W:HAc (30:69:1) | Shaking, 50 min, 70 °C | 8.5 mg GAE/g | |||||
| W | Shaking, 10 min, 80 °C | 8.7 mg GAE/g | |||||
| China | - | 70% MeOH | 1:10 | MAE, 110 °C, 2.45 GHz, 15 min | 0.73 mg GAE/g dw | [45] | |
| 70% MeOH | 1:10 | Shaking, 4 h, RT | 0.38 mg GAE/g dw | ||||
| Undaria pinnatifida | Spain | - | W | 1:10 | PLE, 20 min, 100 °C, 1500 psi | 3.79 mg GAE/g | [70] |
| PLE, 20 min, 200 °C, 1500 psi | 67.11 mg GAE/g | ||||||
| Japan | - | MeOH | 1:100 | Shaking, 24 h, 23 °C, | 1.3 mg GAE/g | [62] | |
| 70% ACE | Shaking, 30 min, 30 °C | 5.7 mg GAE/g | |||||
| 80% MeOH | Shaking, 1 h, 70 °C | 5.9 mg GAE/g | |||||
| MeOH:W:HAc (30:69:1) | Shaking, 50 min, 70 °C | 3.7 mg GAE/g | |||||
| W | Shaking, 10 min, 80 °C | 8.6 mg GAE/g |
| Algae Species | Order | Collecting Location | Collecting Period | Solvent | Plant: Solvent Ratio | Extraction Mode | Total Phenolic Content | Reference |
|---|---|---|---|---|---|---|---|---|
| Asperococcus bullosus | Ectocarpales | France | June 2007 | DCM, MeOH | - | PLE, 75 °C, 1500 psi | 11.1 mg PGE/g | [48] |
| Cladostephus spongiosum | Sphacelariales | Tunisia | July 2015 | 50% EtOH | 1:5 | Shaking, 30 min, 50 °C | 10.91 mg PGE/g dw | [4] |
| Desmarestia ligulata | Desmarestiales | France | May 2007 | DCM, MeOH | - | PLE, 75 °C, 1500 psi | 12.2 mg PGE/g | [48] |
| Halopteris scoparia | Sphacelariales | Algeria | December 2012, June and September 2014 | W | 1:100 | Shaking, 1 h, RT | 1.05 mg GAE/g | [39] |
| Spain | - | 70% MeOH | - | - | 0.16% dw | [69] | ||
| Spain | March-April 2008 | MeOH | 1:15 | Shaking, 2 h, RT | 3.29 mg GAE/g dw | [71] | ||
| EtOH | 2.92 mg GAE/g dw | |||||||
| W | 2.5 mg GAE/g dw | |||||||
| 50% MeOH | 1.23 mg GAE/g dw | |||||||
| Spain | March-April 2008 | MeOH | 1:15 | Shaking, 2 h, RT | 255.2 mg GAE/g dw | [71] | ||
| Saccorhiza polyschides | Tilopteridales | France | June 2007 | DCM, MeOH | - | PLE, 75 °C, 1500 psi | 16.6 mg PGE/g | [48] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Generalić Mekinić, I.; Skroza, D.; Šimat, V.; Hamed, I.; Čagalj, M.; Popović Perković, Z. Phenolic Content of Brown Algae (Pheophyceae) Species: Extraction, Identification, and Quantification. Biomolecules 2019, 9, 244. https://doi.org/10.3390/biom9060244
Generalić Mekinić I, Skroza D, Šimat V, Hamed I, Čagalj M, Popović Perković Z. Phenolic Content of Brown Algae (Pheophyceae) Species: Extraction, Identification, and Quantification. Biomolecules. 2019; 9(6):244. https://doi.org/10.3390/biom9060244
Chicago/Turabian StyleGeneralić Mekinić, Ivana, Danijela Skroza, Vida Šimat, Imen Hamed, Martina Čagalj, and Zvjezdana Popović Perković. 2019. "Phenolic Content of Brown Algae (Pheophyceae) Species: Extraction, Identification, and Quantification" Biomolecules 9, no. 6: 244. https://doi.org/10.3390/biom9060244
APA StyleGeneralić Mekinić, I., Skroza, D., Šimat, V., Hamed, I., Čagalj, M., & Popović Perković, Z. (2019). Phenolic Content of Brown Algae (Pheophyceae) Species: Extraction, Identification, and Quantification. Biomolecules, 9(6), 244. https://doi.org/10.3390/biom9060244