Freeze Tolerance in Sculpins (Pisces; Cottoidea) Inhabiting North Pacific and Arctic Oceans: Antifreeze Activity and Gene Sequences of the Antifreeze Protein


Abstract
:1. Introduction
2. Materials and Methods
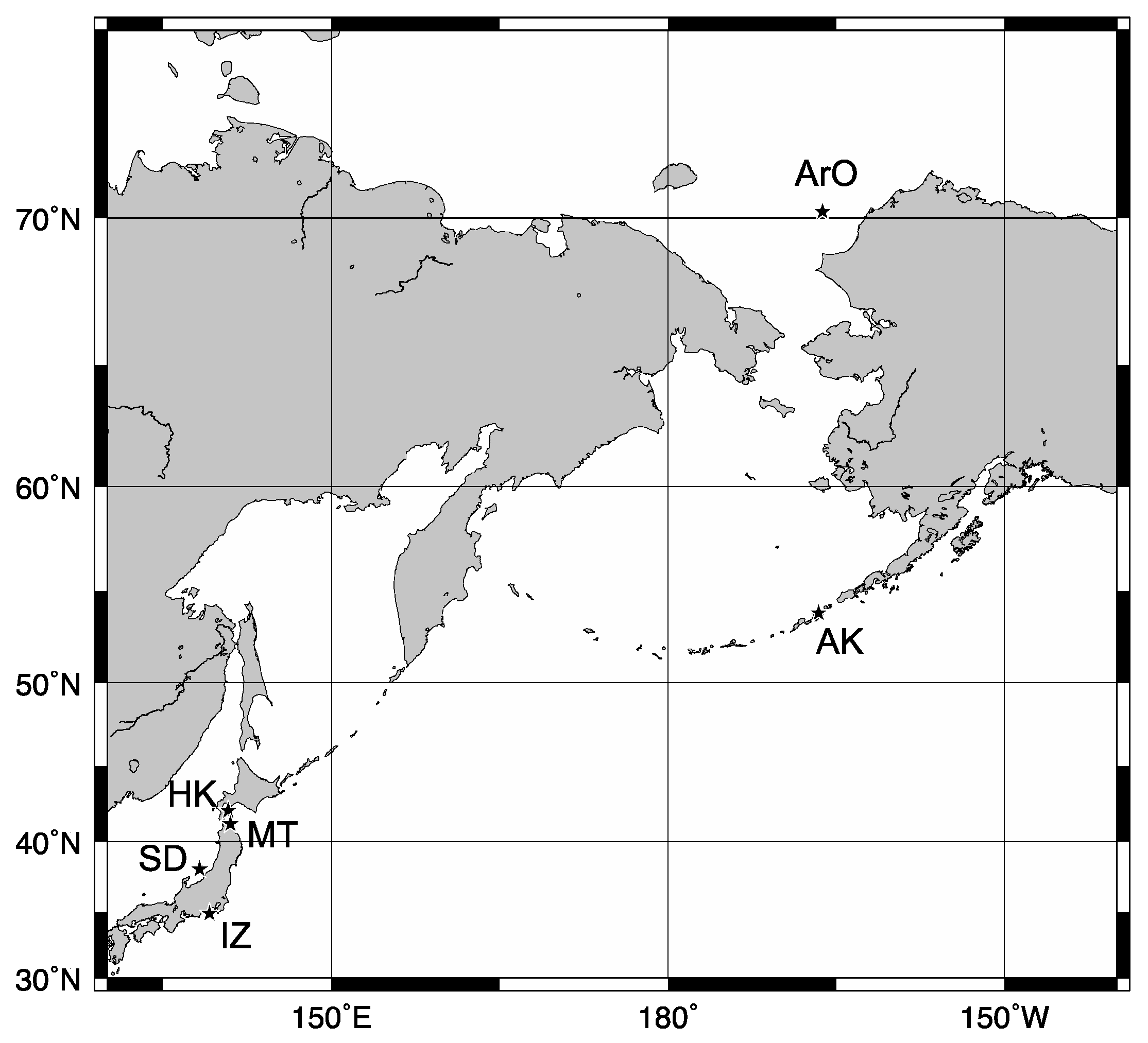
2.1. Sample Collections
2.2. Measurement of Antifreeze Activities
2.3. cDNA Syntheses, Cloning, and Sequencing
2.4. Bioinformatics Analyses
3. Results
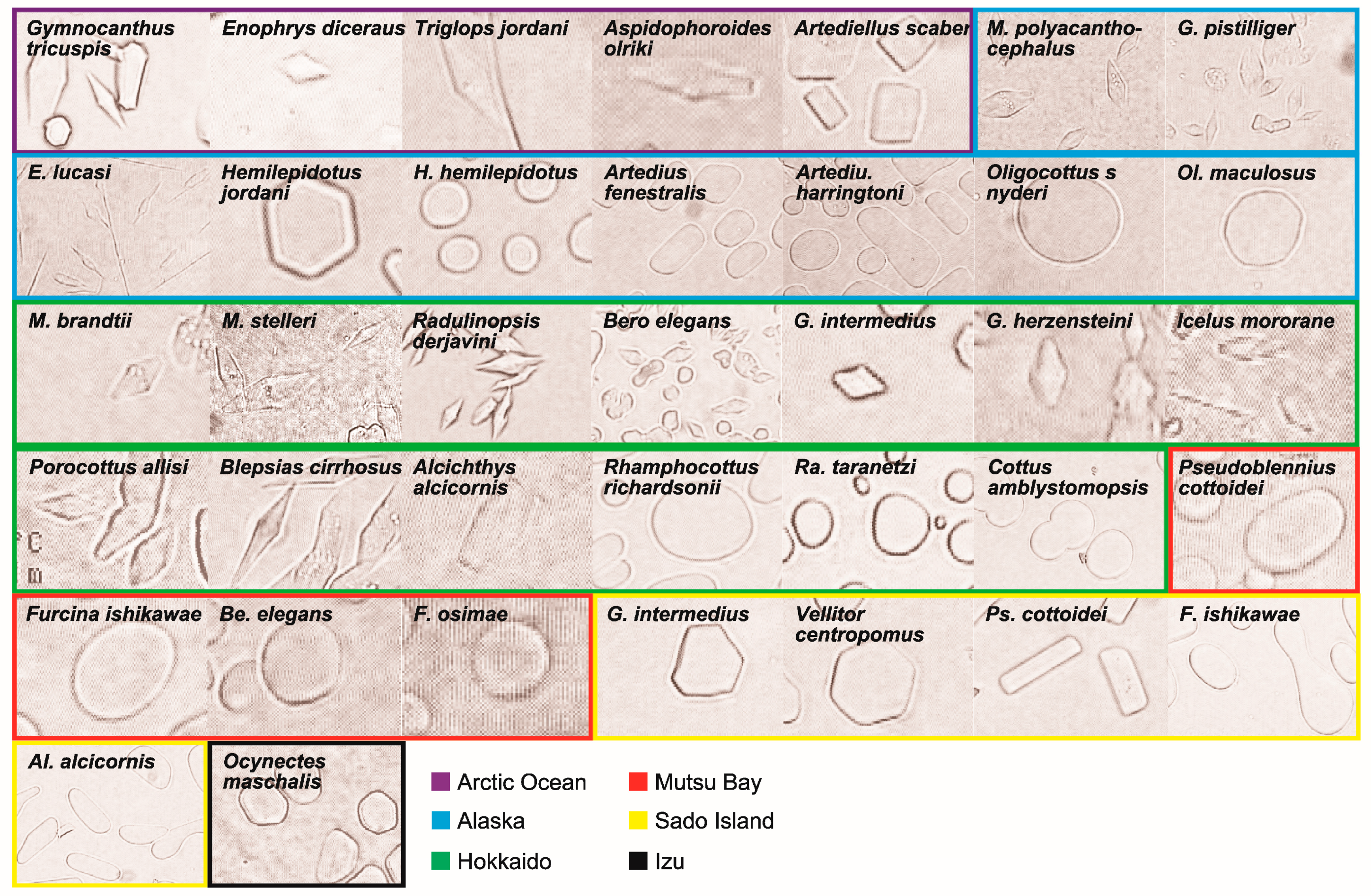
3.1. Antifreeze Activity
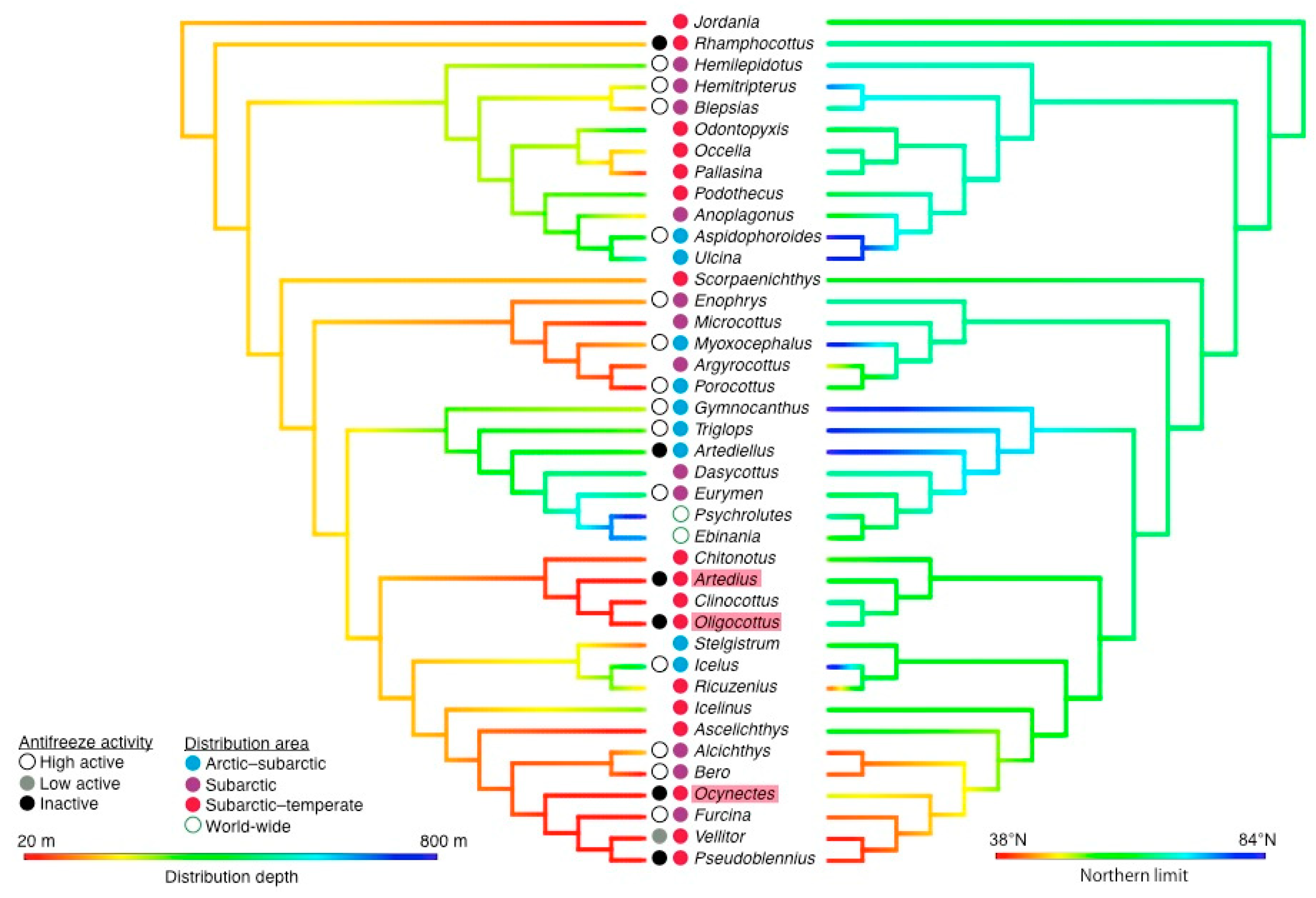
3.2. Genetic Structures of Cottidae Antifreeze Proteins
4. Discussion
4.1. Type I AFP Was Acquired before the Divergence within Cottoidea
4.2. Antifreeze Activity and Current Cold Adaptation
4.3. Distributional Expansion to Cold Regions and Acquisition of AFP
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, W.; Hui, J.H.L.; Williams, G.A.; Cartwright, S.R.; Tsang, L.M.; Chu, K.H. Comparative transcriptomics across populations offers new insights into the evolution of thermal resistance in marine snails. Mar. Biol. 2016, 163, 92. [Google Scholar] [CrossRef]
- Werner, I.; Linares-Casenave, J.; Van Eenennaam, J.P.; Doroshov, S.I. The effect of temperature stress on development and heat-shock protein expression in larval green sturgeon (Acipenser mirostris). Environ. Biol. Fishes 2006, 79, 191–200. [Google Scholar] [CrossRef]
- Davies, P.L. Ice-binding proteins: A remarkable diversity of structures for stopping and starting ice growth. Trends Biochem. Sci. 2014, 39, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, G.L.; Hew, C.L.; Davies, P.L. Antifreeze proteins of teleost fishes. Annu. Rev. Physiol. 2001, 63, 359–390. [Google Scholar] [CrossRef] [PubMed]
- DeVries, A.L. Role of glycopeptides and peptides in inhibition of crystallization of water in polar fishes. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1984, 304, 575–588. [Google Scholar]
- Cheng, C.H. Evolution of the diverse antifreeze proteins. Curr. Opin. Genet. Dev. 1988, 8, 715–720. [Google Scholar] [CrossRef]
- DeVries, A.L.; Wohlschlag, D.E. Freezing resistance in some antarctic fishes. Science 1969, 163, 1073–1075. [Google Scholar] [CrossRef]
- Duman, J.G.; DeVries, A.L. Isolation, characterization, and physical properties of protein antifreezes from the winter flounder, pseudopleuronectes americanus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1976, 54, 375–380. [Google Scholar] [CrossRef]
- Slaughter, D.; Fletcher, G.L.; Ananthanarayanan, V.S.; Hew, C.L. Antifreeze proteins from the sea raven, Hemitripterus americanus. Further evidence for diversity among fish polypeptide antifreezes. J. Biol. Chem. 1981, 256, 2022–2026. [Google Scholar]
- Hew, C.; Slaughter, D.; Joshi, S.; Fletcher, G.; Ananthanarayanan, V.S. Antifreeze polypeptides from the newfoundland ocean pout, Macrozoarces americanus: Presence of multiple and compositionally diverse components. J. Comp. Physiol. B 1984, 155, 81–88. [Google Scholar] [CrossRef]
- Li, X.-M.; Trinh, K.-Y.; Hew, C.L. Structure of an antifreeze polypeptide and its precursor from the ocean pout, Macrozoarces americanus. J. Biol. Chem. 1985, 260, 12904–12909. [Google Scholar] [PubMed]
- Raymond, J.A.; DeVries, A.L. Adsorption inhibition as a mechanism of freezing resistance in polar fishes. Proc. Natl. Acad. Sci. USA 1977, 74, 2589–2593. [Google Scholar] [CrossRef] [PubMed]
- Near, T.J.; Dornburg, A.; Kuhn, K.L.; Eastman, J.T.; Pennington, J.N.; Patarnello, T.; Zane, L.; Fernandez, D.A.; Jones, C.D. Ancient climate change, antifreeze, and the evolutionary diversification of antarctic fishes. Proc. Natl. Acad. Sci. USA 2012, 109, 3434–3439. [Google Scholar] [CrossRef]
- Stickley, C.E.; St John, K.; Koç, N.; Jordan, R.W.; Passchier, S.; Pearce, R.B.; Kearns, L.E. Evidence for middle eocene arctic sea ice from diatoms and ice-rafted debris. Nature 2009, 460, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, A.; Nishimiya, Y.; Tsuda, S.; Togashi, K.; Munehara, H. Gene expression of antifreeze protein in relation to historical distributions of myoxocephalus fish species. Mar. Biol. 2018, 165, 181. [Google Scholar] [CrossRef]
- Hobbs, R.S.; Fletcher, G.L. Epithelial dominant expression of antifreeze proteins in cunner suggests recent entry into a high freeze-risk ecozone. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 164, 111–118. [Google Scholar] [CrossRef]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V. Fishes of the World, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Mecklenburg, C.W.; Møller, P.R.; Steinke, D. Biodiversity of arctic marine fishes: Taxonomy and zoogeography. Mar. Biodivers. 2011, 41, 109–140. [Google Scholar] [CrossRef]
- Ng, N.F.; Khiet-Yen, T.; Hew, C.L. Structure of an antifreeze polypeptide precursor from the sea raven, Hemitripterus americanus. J. Biol. Chem. 1986, 261, 15690–15695. [Google Scholar]
- Nishimiya, Y.; Kondo, H.; Takamichi, M.; Sugimoto, H.; Suzuki, M.; Miura, A.; Tsuda, S. Crystal structure and mutational analysis of ca2+-independent type ii antifreeze protein from longsnout poacher, Brachyopsis rostratus. J. Mol. Biol. 2008, 382, 734–746. [Google Scholar] [CrossRef]
- Low, W.K.; Miao, M.; Ewart, K.V.; Yang, D.S.C.; Fletcher, G.L.; Hew, C.L. Skin-type antifreeze protein from the shorthorn sculpin, Myoxocephalus scorpius: Expression and characterization of a mr 9,700 recombinant protein. J. Biol. Chem. 1998, 273, 23098–23103. [Google Scholar] [CrossRef]
- Reisman, H.M.; Fletcher, G.L.; Kao, M.H.; Shears, M.A. Antifreeze proteins in the grubby sculpin, Myoxocephalus aenaeus and the tomcod, microgadus tomcod: Comparisons of seasonal cycles. Environ. Biol. Fishes 1987, 18, 295–301. [Google Scholar] [CrossRef]
- Low, W.K.; Lin, Q.; Stathakis, C.; Miao, M.; Fletcher, G.L.; Hew, C.L. Isolation and characterization of skin-type, type i antifreeze polypeptides from the longhorn sculpin, Myoxocephalus octodecemspinosus. J. Biol. Chem. 2001, 276, 11582–11589. [Google Scholar] [CrossRef]
- Duman, J.G.; DeVries, A.L. The role of macromolecular antifreezes in cold water fishes. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1974, 52, 193–199. [Google Scholar] [CrossRef]
- Mahatabuddin, S.; Tsuda, S. Applications of antifreeze proteins: Practical use of the quality products from japanese fishes. In Survival Strategies in Extreme Cold and Desiccation; Iwaya-Inoue, M., Sakurai, M., Uemura, M., Eds.; Springer: Heidelberg, Germany, 2018; pp. 321–337. [Google Scholar]
- Tsuda, S.; Miura, A. Antifreeze Proteins Originating in Fishes. U.S. Patent Application No. 10,104, 27 January 2005. [Google Scholar]
- Nishimiya, Y.; Kondo, H.; Yasui, M.; Sugimoto, H.; Noro, N.; Sato, R.; Suzuki, M.; Miura, A.; Tsuda, S. Crystallization and preliminary x-ray crystallographic analysis of ca2+-independent and ca2+-dependent species of the type ii antifreeze protein. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2006, 62, 538–541. [Google Scholar] [CrossRef]
- Mecklenburg, C.W.; Mecklenburg, T.A.; Thorsteinson, L.K. Fishes of Alaska; American Fisheries Society: North Bethesda, MD, USA, 2002; p. 1037. [Google Scholar]
- Nakabo, T. Fishes of Japan with Pictoral Keys to the Species, 3rd ed.; Tokai University Press: Kanagawa, Japan, 2013. [Google Scholar]
- DeVries, A.L. Biological antifreeze agents in coldwater fishes. Comp. Biochem. Physiol. A Physiol. 1982, 73, 627–640. [Google Scholar] [CrossRef]
- Mahatabuddin, S.; Hanada, Y.; Nishimiya, Y.; Miura, A.; Kondo, H.; Davies, P.L.; Tsuda, S. Concentration-dependent oligomerization of an alpha-helical antifreeze polypeptide makes it hyperactive. Sci. Rep. 2017, 7, 42501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, Y.; Miura, R.; Takemoto, Y.; Tsuda, S.; Kawahara, H.; Obata, H. Type ii antifreeze protein from a mid-latitude freshwater fish, Japanese smelt (Hypomesus nipponensis). Biosci. Biotechnol. Biochem. 2003, 67, 461–466. [Google Scholar] [CrossRef]
- Nishimiya, Y.; Sato, R.; Takamichi, M.; Miura, A.; Tsuda, S. Co-operative effect of the isoforms of type iii antifreeze protein expressed in notched-fin eelpout, Zoarces elongatus kner. FEBS J. 2005, 272, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Takamichi, M.; Nishimiya, Y.; Miura, A.; Tsuda, S. Effect of annealing time of an ice crystal on the activity of type III antifreeze protein. FEBS J. 2007, 274, 6469–6476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, P.L.; Hew, C.L. Biochemistry of fish antifreeze proeins. FASEB J. 1990, 4, 2460–2468. [Google Scholar] [CrossRef]
- Mahatabuddin, S.; Nishimiya, Y.; Miura, A.; Kondo, H.; Tsuda, S. Critical ice shaping concentration (CICS): A new parameter to evaluate the activity of antifreeze proteins. Cryobiol. Cryotechnol. 2016, 62, 95–103. [Google Scholar]
- Rahman, A.T.; Arai, T.; Yamauchi, A.; Miura, A.; Kondo, H.; Ohyama, Y.; Tsuda, S. Ice crystallization is strongly inhibited when antifreeze proteins bind to multiple ice planes. Sci. Rep. 2019, 9, 2212. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.A.; Hobbs, R.S.; Fletcher, G.L.; Davies, P.L. Helical antifreeze proteins have independently evolved in fishes on four occasions. PLoS ONE 2013, 8, e81285. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. Mafft multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. Trimal: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Tanabe, A.S. Kakusan4 and aminosan: Two programs for comparing nonpartitioned, proportional and separate models for combined molecular phylogenetic analyses of multilocus sequence data. Mol. Ecol. Resour. 2011, 11, 914–921. [Google Scholar] [CrossRef]
- Stamatakis, A. Raxml-vi-hpc: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. Fishbase. World Wide Web Electronic Publication. Available online: http://www.fishbase.org (accessed on 28 February 2019).
- Ewart, K.V.; Fletcher, G.L. Isolation and characterization of antifreeze proteins from smelt (osmerus mordax) and atlantic herring (clupea harengus harengus). Can. J. Zool. 1990, 68, 1652–1658. [Google Scholar] [CrossRef]
- Hayes, P.H.; Scott, G.; Ng, N.; Hew, C.; Davies, P. Cystine-rich type ii antifreeze protein precursor is initiated from the third aug codon of its mrna. J. Biol. Chem. 1989, 264, 18761–18767. [Google Scholar] [PubMed]
- Graham, L.A.; Lougheed, S.C.; Ewart, K.V.; Davies, P.L. Lateral transfer of a lectin-like antifreeze protein gene in fishes. PLoS ONE 2008, 3, e2616. [Google Scholar] [CrossRef] [PubMed]
- Sorhannus, U. Evolution of type ii antifreeze protein genes in teleost fish: A complex scenario involving lateral gene transfers and episodic directional selection. Evol. Bioinform. Online 2012, 8, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Briggs, J.C. Marine centres of origin as evolutionary engines. J. Biogeogr. 2003, 30, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Cziko, P.A.; Evans, C.W.; Cheng, C.H.; DeVries, A.L. Freezing resistance of antifreeze-deficient larval antarctic fish. J. Exp. Biol. 2006, 209, 407–420. [Google Scholar] [CrossRef]
- Desjardins, M.; Graham, L.A.; Davies, P.L.; Fletcher, G.L. Antifreeze protein gene amplification facilitated niche exploitation and speciation in wolffish. FEBS J. 2012, 279, 2215–2230. [Google Scholar] [CrossRef] [Green Version]
- Jung, A.; Johnson, P.; Eastman, J.; DeVries, A. Protein content and freezing avoidance properties of the subdermal extracellular matrix and serum of the antarctic snailfish, Paraliparis devriesi. Fish Physiol. Biochem. 1995, 14, 71–80. [Google Scholar] [CrossRef] [PubMed]
- DeMartini, E.E. Spatial aspects of reproduction in buffalo sculpin, Enophrys bison. Environ. Biol. Fishes 1978, 3, 331–336. [Google Scholar] [CrossRef]
- Munehara, H. Utilization of polychaete tubes as spawning substrate by the sea raven Hemitripterus villosus (scorpaeniformes). Environ. Biol. Fishes 1992, 33, 395–398. [Google Scholar] [CrossRef]
- Markevich, A. Spawning of the sea Ravenhemitripterus villosus in peter the great bay, Sea of Japan. Russ. J. Mar. Biol. 2000, 26, 283–285. [Google Scholar] [CrossRef]
- Mandic, M.; Ramon, M.L.; Gracey, A.Y.; Richards, J.G. Divergent transcriptional patterns are related to differences in hypoxia tolerance between the intertidal and the subtidal sculpins. Mol. Ecol. 2014, 23, 6091–6103. [Google Scholar] [CrossRef]
- Near, T.J.; Dornburg, A.; Eytan, R.I.; Keck, B.P.; Smith, W.L.; Kuhn, K.L.; Moore, J.A.; Price, S.A.; Burbrink, F.T.; Friedman, M.; et al. Phylogeny and tempo of diversification in the superradiation of spiny-rayed fishes. Proc. Natl. Acad. Sci. USA 2013, 110, 12738–12743. [Google Scholar] [CrossRef] [Green Version]
- Møller, P.R.; Gravlund, P. Phylogeny of the eelpout genus lycodes (pisces, zoarcidae) as inferred from mitochondrial cytochrome b and 12s rdna. Mol. Phylogen. Evol. 2003, 26, 369–388. [Google Scholar] [CrossRef]
- Kontula, T.; Väinölä, R. Relationships of palearctic and nearctic ‘glacial relict’ Myoxocephalus sculpins from mitochondrial DNA data. Mol. Ecol. 2003, 12, 3179–3184. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Date | SBT | N | Antifreeze Activity | ||
|---|---|---|---|---|---|---|
| High | Low | None | ||||
| Arctic Ocean | July 2013 | −0.7 | 5 | 4 | 0 | 1 |
| Alaska | March 2014 | 3.6 | 9 | 3 | 1 | 5 |
| Hokkaido | April 2014 | 4.4 | 13 | 7 | 3 | 3 |
| Mutsu Bay | December 2013 | 10.4 | 3 | 0 | 0 | 3 |
| Sado Island | February 2014 | 9.6 | 5 | 0 | 2 | 3 |
| Izu | March 2015 | 7.0 | 1 | 0 | 0 | 1 |
| Summary * | 32 | 14 | 6 | 16 | ||
| 1st Reaction | 2nd Reaction | ||
|---|---|---|---|
| Forward | Reverse | Forward | Reverse |
| 5’1 | 3’1 | 5’2 | 3’2 |
| LS-ST-P2 | 3’2 | ||
| Deg1 | 3’2 | ||
| 5’2 | 3’1 | LS-ST-P2 | 3’2 |
| Deg1 | 3’2 | ||
| LS-ST-P2 | 3’1 | Deg1 | 3’2 |
| Family | Genus | Species | Location | Depth | Date | Antifreeze Activity |
|---|---|---|---|---|---|---|
| Cottidae | Alcichthys | alcicornis | HK | Subtidal | April 2015 | Low |
| SD | February 2014 | None | ||||
| Artediellus | scaber | ArO | Subtidal | July 2013 | None | |
| Artedius | fenestralis | AK | Tidal | March 2014 | None | |
| harringtoni | AK | Tidal | March 2014 | None | ||
| Bero | elegans | HK | Subtidal | April 2014 | Low | |
| MT | December 2013 | None | ||||
| Cottus | amblystomopsis | HK | Stream | April 2014 | None | |
| Enophrys | lucasi | AK | Subtidal | March 2014 | High | |
| diceraus | ArO | Subtidal | July 2013 | High | ||
| Furcina | ishikawae | MT | Subtidal | December 2013 | None | |
| SD | February 2014 | None | ||||
| osimae | MT | Subtidal | December 2013 | None | ||
| Gymnocanthus | tricuspis | ArO | Subtidal | July 2013 | High | |
| pistilliger | AK | Subtidal | March 2014 | High | ||
| herzensteini | HK | Subtidal | April 2014 | High | ||
| Janurary 2015 | High | |||||
| intermedius | HK | Subtidal | April 2014 | High | ||
| March 2015 | High | |||||
| SD | February 2014 | Low | ||||
| Icelus | mororane | HK | Subtidal | March 2015 | High | |
| Myoxocephalus | polyacanthocephalus | AK | Subtidal | March 2014 | High | |
| stelleri | HK | Subtidal | November 2014 | None | ||
| Janurary 2015 | Low | |||||
| February 2015 | None | |||||
| March 2015 | High | |||||
| April 2015 | Low | |||||
| brandtii | HK | Subtidal | Janurary 2015 | None | ||
| February 2015 | None | |||||
| March 2015 | High | |||||
| April 2015 | High | |||||
| Oligocottus | snyderi | AK | Tidal | March 2014 | None | |
| maculosus | AK | Tidal | March 2014 | None | ||
| Ocynectes | maschalis | IZ | Tidal | April 2015 | None | |
| Porocottus | allisi | HK | Subtidal | March 2015 | High | |
| Pseudoblennius | cottoides | SD | Subtidal | November 2013 | None | |
| February 2014 | None | |||||
| MT | December 2013 | None | ||||
| Radulinopsis | taranetzi | HK | Subtidal | April 2014 | None | |
| derjavini | HK | Subtidal | April 2014 | High | ||
| Triglops | pingelii | ArO | Subtidal | July 2013 | High | |
| Vellitor | centropomus | SD | Subtidal | November 2013 | None | |
| February 2014 | Low | |||||
| Rhamphocottidae | Rhamphocottus | richardsonii | HK | Subtidal | April 2014 | None |
| Agonidae | Aspidophoroides | olrikii | ArO | Subtidal | July 2013 | High |
| Blepsias | cirrhosus | HK | Subtidal | April 2014 | High | |
| Hemilepidotus | hemilepidotus | AK | Subtidal | March 2014 | None | |
| jordani | AK | Subtidal | March 2014 | Low | ||
| 3 families | 20 genera | 32 species |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamazaki, A.; Nishimiya, Y.; Tsuda, S.; Togashi, K.; Munehara, H. Freeze Tolerance in Sculpins (Pisces; Cottoidea) Inhabiting North Pacific and Arctic Oceans: Antifreeze Activity and Gene Sequences of the Antifreeze Protein. Biomolecules 2019, 9, 139. https://doi.org/10.3390/biom9040139
Yamazaki A, Nishimiya Y, Tsuda S, Togashi K, Munehara H. Freeze Tolerance in Sculpins (Pisces; Cottoidea) Inhabiting North Pacific and Arctic Oceans: Antifreeze Activity and Gene Sequences of the Antifreeze Protein. Biomolecules. 2019; 9(4):139. https://doi.org/10.3390/biom9040139
Chicago/Turabian StyleYamazaki, Aya, Yoshiyuki Nishimiya, Sakae Tsuda, Koji Togashi, and Hiroyuki Munehara. 2019. "Freeze Tolerance in Sculpins (Pisces; Cottoidea) Inhabiting North Pacific and Arctic Oceans: Antifreeze Activity and Gene Sequences of the Antifreeze Protein" Biomolecules 9, no. 4: 139. https://doi.org/10.3390/biom9040139
APA StyleYamazaki, A., Nishimiya, Y., Tsuda, S., Togashi, K., & Munehara, H. (2019). Freeze Tolerance in Sculpins (Pisces; Cottoidea) Inhabiting North Pacific and Arctic Oceans: Antifreeze Activity and Gene Sequences of the Antifreeze Protein. Biomolecules, 9(4), 139. https://doi.org/10.3390/biom9040139