Nucleotide Transport and Metabolism in Diatoms


Abstract
1. Introduction
2. Nucleotide Synthesis in Diatom Cells
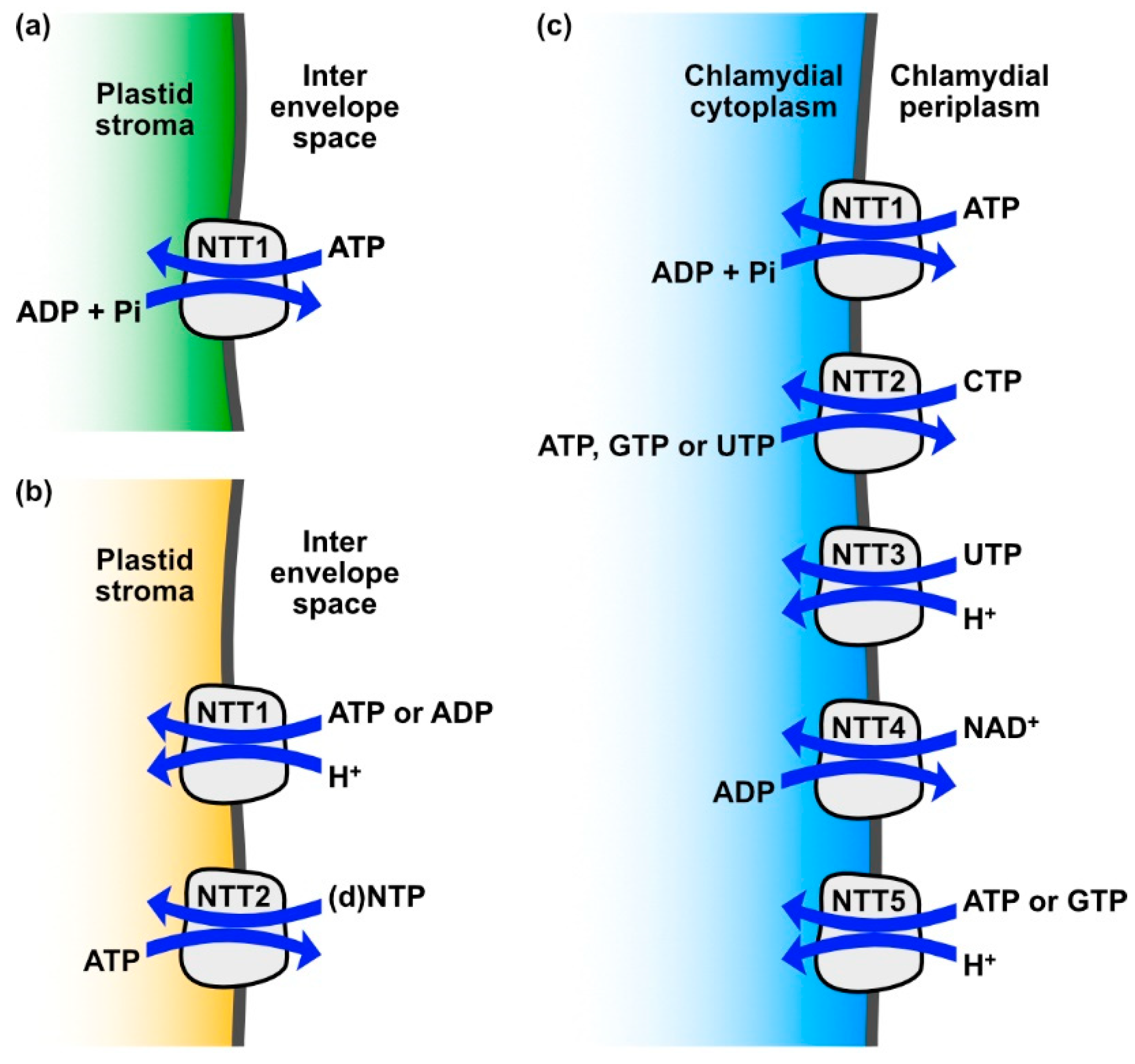
3. Nucleotide Transporters in Diatoms
4. Evolutionary Implications of the Distribution of NTTs across the Tree of Life
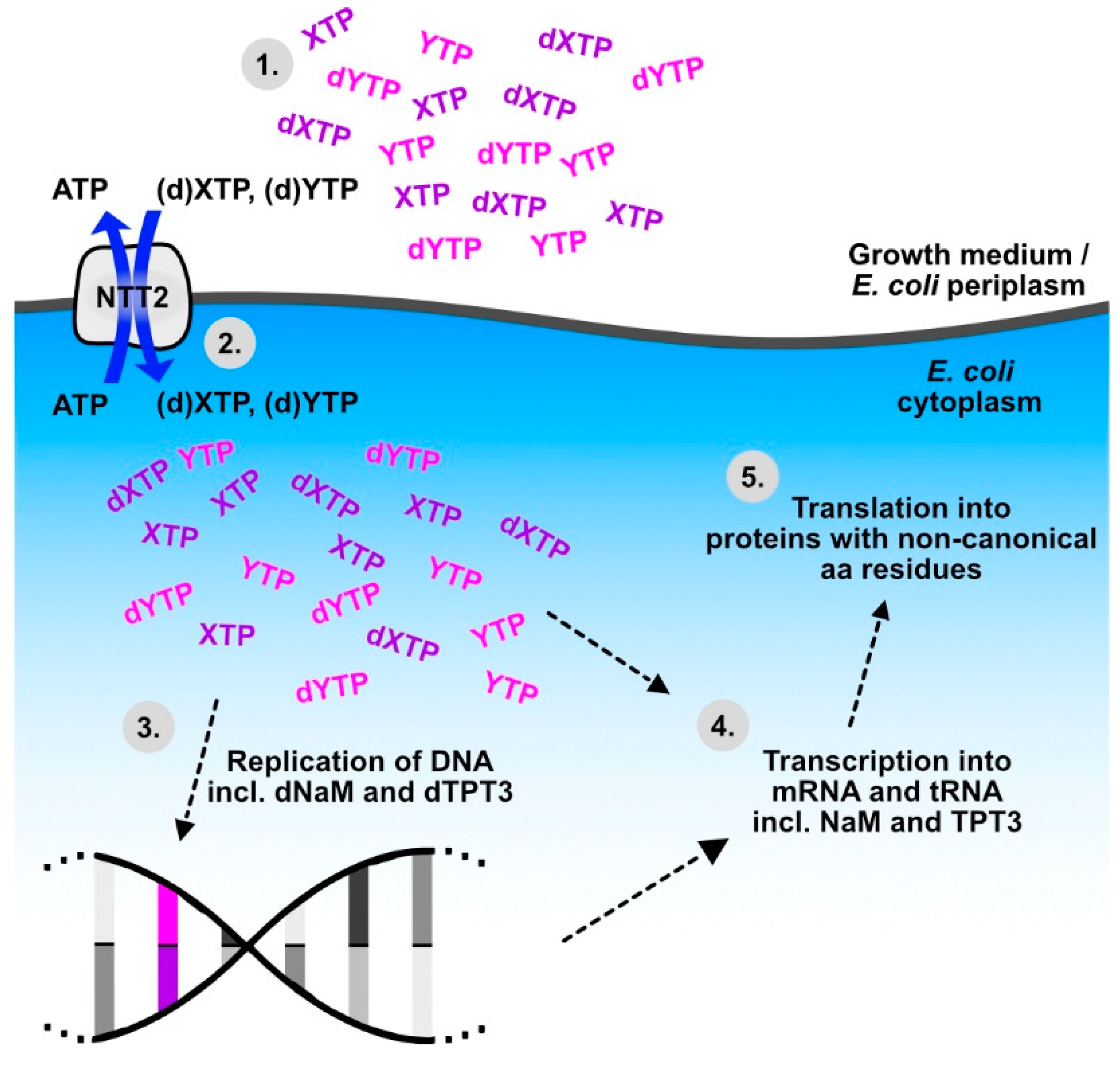
5. Biotechnological Applications of Diatom NTTs
6. Conclusions and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Field, C.B. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Armbrust, E.V. The life of diatoms in the world’s oceans. Nature 2009, 459, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G.; Katz, M.E.; Knoll, A.H.; Quigg, A.; Raven, J.A.; Schofield, O.; Taylor, F.J. The evolution of modern eukaryotic phytoplankton. Science 2004, 305, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Archibald, J.M. Genomic perspectives on the birth and spread of plastids. Proc. Natl. Acad. Sci. USA 2015. [Google Scholar] [CrossRef]
- Archibald, J.M. Endosymbiosis and Eukaryotic Cell Evolution. Curr. Biol. CB 2015, 25, R911–R921. [Google Scholar] [CrossRef]
- Brodie, J.; Ball, S.G.; Bouget, F.-Y.; Chan, C.X.; De Clerck, O.; Cock, J.M.; Gachon, C.; Grossman, A.R.; Mock, T.; Raven, J.A.; et al. Biotic interactions as drivers of algal origin and evolution. New Phytol. 2017, 216, 670–681. [Google Scholar] [CrossRef]
- Oborník, M. Endosymbiotic Evolution of Algae, Secondary Heterotrophy and Parasitism. Biomolecules 2019, 9, 266. [Google Scholar] [CrossRef]
- Baurain, D.; Brinkmann, H.; Petersen, J.; Rodriguez-Ezpeleta, N.; Stechmann, A.; Demoulin, V.; Roger, A.J.; Burger, G.; Lang, B.F.; Philippe, H. Phylogenomic evidence for separate acquisition of plastids in cryptophytes, haptophytes, and stramenopiles. Mol. Biol. Evol. 2010, 27, 1698–1709. [Google Scholar] [CrossRef]
- Dorrell, R.G.; Gile, G.; McCallum, G.; Méheust, R.; Bapteste, E.P.; Klinger, C.M.; Brillet-Guéguen, L.; Freeman, K.D.; Richter, D.J.; Bowler, C. Chimeric origins of ochrophytes and haptophytes revealed through an ancient plastid proteome. eLife 2017, 6, e23717. [Google Scholar] [CrossRef]
- Nowack, E.C.M.; Weber, A.P.M. Genomics-Informed Insights into Endosymbiotic Organelle Evolution in Photosynthetic Eukaryotes. Ann. Rev. Plant Biol. 2018, 69, 51–84. [Google Scholar] [CrossRef]
- Gould, S.B.; Waller, R.F.; McFadden, G.I. Plastid evolution. Ann. Rev. Plant Biol. 2008, 59, 491–517. [Google Scholar] [CrossRef] [PubMed]
- Kroth, P.G. Protein transport into secondary plastids and the evolution of primary and secondary plastids. Int. Rev. Cytol. 2002, 221, 191–255. [Google Scholar] [CrossRef] [PubMed]
- Flori, S.; Jouneau, P.-H.; Finazzi, G.; Maréchal, E.; Falconet, D. Ultrastructure of the Periplastidial Compartment of the Diatom Phaeodactylum tricornutum. Protist 2016, 167, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Gruber, A.; Weber, T.; Rio Bartulos, C.; Vugrinec, S.; Kroth, P.G. Intracellular distribution of the reductive and oxidative pentose phosphate pathways in two diatoms. J. Basic Microbiol. 2009, 49, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Moog, D.; Stork, S.; Zauner, S.; Maier, U.-G. In Silico and In Vivo Investigations of Proteins of a Minimized Eukaryotic Cytoplasm. Genome Biol. Evol. 2011, 3, 375–382. [Google Scholar] [CrossRef]
- Weber, T.; Gruber, A.; Kroth, P.G. The Presence and Localization of Thioredoxins in Diatoms, Unicellular Algae of Secondary Endosymbiotic Origin. Mol. Plant. 2009, 2, 468–477. [Google Scholar] [CrossRef]
- Ball, S.G.; Morell, M.K. From Bacterial Glycogen to Starch: Understanding the Biogenesis of the Plant Starch Granule. Ann. Rev. Plant Biol. 2003, 54, 207–233. [Google Scholar] [CrossRef]
- Deschamps, P.; Haferkamp, I.; d’Hulst, C.; Neuhaus, H.E.; Ball, S.G. The relocation of starch metabolism to chloroplasts: When, why and how. Trends Plant Sci. 2008, 13, 574–582. [Google Scholar] [CrossRef]
- Ball, S.; Colleoni, C.; Cenci, U.; Raj, J.N.; Tirtiaux, C. The evolution of glycogen and starch metabolism in eukaryotes gives molecular clues to understand the establishment of plastid endosymbiosis. J. Exp. Bot. 2011, 62, 1775–1801. [Google Scholar] [CrossRef]
- Haferkamp, I.; Fernie, A.R.; Neuhaus, H.E. Adenine nucleotide transport in plants: Much more than a mitochondrial issue. Trends Plant Sci. 2011, 16, 507–515. [Google Scholar] [CrossRef]
- Linka, N.; Hurka, H.; Lang, B.F.; Burger, G.; Winkler, H.H.; Stamme, C.; Urbany, C.; Seil, I.; Kusch, J.; Neuhaus, H.E. Phylogenetic relationships of non-mitochondrial nucleotide transport proteins in bacteria and eukaryotes. Gene 2003, 306, 27–35. [Google Scholar] [CrossRef]
- Weber, A.P.; Linka, N. Connecting the plastid: Transporters of the plastid envelope and their role in linking plastidial with cytosolic metabolism. Ann. Rev. Plant Biol. 2011, 62, 53–77. [Google Scholar] [CrossRef] [PubMed]
- Trentmann, O.; Jung, B.; Neuhaus, H.E.; Haferkamp, I. Nonmitochondrial ATP/ADP transporters accept phosphate as third substrate. J. Biol. Chem. 2008, 283, 36486–36493. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Haferkamp, I.; Lepetit, B.; Molchanova, M.; Hou, S.; Jeblick, W.; Río Bártulos, C.; Kroth, P.G. Reduced vacuolar β-1,3-glucan synthesis affects carbohydrate metabolism as well as plastid homeostasis and structure in Phaeodactylum tricornutum. Proc. Natl. Acad. Sci. USA 2018, 115, 4791–4796. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, C.; Buchel, C.; Fisahn, J.; Goss, R.; Jakob, T.; Laroche, J.; Lavaud, J.; Lohr, M.; Riebesell, U.; Stehfest, K.; et al. The regulation of carbon and nutrient assimilation in diatoms is significantly different from green algae. Protist 2006, 157, 91–124. [Google Scholar] [CrossRef]
- Schoefs, B.; Hu, H.; Kroth, P.G. The peculiar carbon metabolism in diatoms. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160405. [Google Scholar] [CrossRef]
- Nonoyama, T.; Kazamia, E.; Nawaly, H.; Gao, X.; Tsuji, Y.; Matsuda, Y.; Bowler, C.; Tanaka, T.; Dorrell, R.G. Metabolic Innovations Underpinning the Origin and Diversification of the Diatom Chloroplast. Biomolecules 2019, 9, 322. [Google Scholar] [CrossRef]
- Zrenner, R.; Stitt, M.; Sonnewald, U.; Boldt, R. Pyrimidine and purine biosynthesis and degradation in plants. Ann. Rev. Plant Biol. 2006, 57, 805–836. [Google Scholar] [CrossRef]
- Garavito, M.F.; Narváez-Ortiz, H.Y.; Zimmermann, B.H. Pyrimidine Metabolism: Dynamic and Versatile Pathways in Pathogens and Cellular Development. J. Genet. Genom. 2015, 42, 195–205. [Google Scholar] [CrossRef]
- Kruger, N.J.; von Schaewen, A. The oxidative pentose phosphate pathway: Structure and organisation. Curr. Opin. Plant Biol. 2003, 6, 236–246. [Google Scholar] [CrossRef]
- Witz, S.; Jung, B.; Fürst, S.; Möhlmann, T. De novo pyrimidine nucleotide synthesis mainly occurs outside of plastids, but a previously undiscovered nucleobase importer provides substrates for the essential salvage pathway in Arabidopsis. Plant Cell 2012, 24, 1549–1559. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Michels, A.K.; Wedel, N.; Kroth, P.G. Diatom plastids possess a phosphoribulokinase with an altered regulation and no oxidative pentose phosphate pathway. Plant. Physiol. 2005, 137, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Kroth, P.G.; Chiovitti, A.; Gruber, A.; Martin-Jezequel, V.; Mock, T.; Parker, M.S.; Stanley, M.S.; Kaplan, A.; Caron, L.; Weber, T.; et al. A model for carbohydrate metabolism in the diatom Phaeodactylum tricornutum deduced from comparative whole genome analysis. PLoS ONE 2008, 3, e1426. [Google Scholar] [CrossRef] [PubMed]
- Armbrust, E.V.; Berges, J.A.; Bowler, C.; Green, B.R.; Martinez, D.; Putnam, N.H.; Zhou, S.; Allen, A.E.; Apt, K.E.; Bechner, M.; et al. The genome of the diatom Thalassiosira pseudonana: Ecology, evolution, and metabolism. Science 2004, 306, 79–86. [Google Scholar] [CrossRef]
- Allen, A.E.; Dupont, C.L.; Obornik, M.; Horak, A.; Nunes-Nesi, A.; McCrow, J.P.; Zheng, H.; Johnson, D.A.; Hu, H.; Fernie, A.R.; et al. Evolution and metabolic significance of the urea cycle in photosynthetic diatoms. Nature 2011, 473, 203–207. [Google Scholar] [CrossRef]
- Bowler, C.; Allen, A.E.; Badger, J.H.; Grimwood, J.; Jabbari, K.; Kuo, A.; Maheswari, U.; Martens, C.; Maumus, F.; Otillar, R.P.; et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 2008, 456, 239–244. [Google Scholar] [CrossRef]
- Gruber, A.; Vugrinec, S.; Hempel, F.; Gould, S.B.; Maier, U.G.; Kroth, P.G. Protein targeting into complex diatom plastids: Functional characterisation of a specific targeting motif. Plant. Mol. Biol. 2007, 64, 519–530. [Google Scholar] [CrossRef]
- Kilian, O.; Kroth, P.G. Identification and characterization of a new conserved motif within the presequence of proteins targeted into complex diatom plastids. Plant. J. 2005, 41, 175–183. [Google Scholar] [CrossRef]
- Ast, M.; Gruber, A.; Schmitz-Esser, S.; Neuhaus, H.E.; Kroth, P.G.; Horn, M.; Haferkamp, I. Diatom plastids depend on nucleotide import from the cytosol. Proc. Natl. Acad. Sci. USA 2009, 106, 3621–3626. [Google Scholar] [CrossRef]
- Sakaguchi, T.; Nakajima, K.; Matsuda, Y. Identification of the UMP Synthase Gene by Establishment of Uracil Auxotrophic Mutants and the Phenotypic Complementation System in the Marine Diatom Phaeodactylum tricornutum. Plant Physiol. 2011, 156, 78–89. [Google Scholar] [CrossRef]
- Major, P.; Embley, T.M.; Williams, T.A. Phylogenetic Diversity of NTT Nucleotide Transport Proteins in Free-Living and Parasitic Bacteria and Eukaryotes. Genome Biol. Evol. 2017, 9, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.; Gruber, A.; Ast, M.; Schmitz-Esser, S.; Altensell, J.; Neuhaus, H.E.; Kroth, P.G.; Haferkamp, I. Shuttling of (deoxy-) purine nucleotides between compartments of the diatom Phaeodactylum tricornutum. New Phytol. 2017, 213, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Schober, A.F.; Río Bártulos, C.; Bischoff, A.; Lepetit, B.; Gruber, A.; Kroth, P.G. Organelle Studies and Proteome Analyses of Mitochondria and Plastids Fractions from the Diatom Thalassiosira pseudonana. Plant Cell Physiol. 2019, 60, 1811–1828. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, H.; Cantor, M.; Dusheyko, S.; Hua, S.; Poliakov, A.; Shabalov, I.; Smirnova, T.; Grigoriev, I.V.; Dubchak, I. The genome portal of the Department of Energy Joint Genome Institute: 2014 updates. Nucleic Acids Res. 2014, 42, D26–D31. [Google Scholar] [CrossRef]
- Bendtsen, J.D.; Nielsen, H.; von Heijne, G.; Brunak, S. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol 2004, 340, 783–795. [Google Scholar] [CrossRef]
- Gruber, A.; Rocap, G.; Kroth, P.G.; Armbrust, E.V.; Mock, T. Plastid proteome prediction for diatoms and other algae with secondary plastids of the red lineage. Plant. J. 2015, 81, 519–528. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Salvatore, M.; Emanuelsson, O.; Winther, O.; von Heijne, G.; Elofsson, A.; Nielsen, H. Detecting sequence signals in targeting peptides using deep learning. Life Sci. Alliance 2019, 2, e201900429. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar] [CrossRef]
- Sonnhammer, E.L.L.; von Heijne, G.; Krogh, A. A hidden Markov model for predicting transmembrane helices in protein sequences. In Proceedings of the Sixth International Conference on Intelligent Systems for Molecular Biology; Glasgow, J., Littlejohn, T., Major, F., Lathrop, R., Sankoff, D., Sensen, C., Eds.; AAAI Press: Menlo Park, CA, USA, 1998; pp. 175–182. [Google Scholar]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes11Edited by F. Cohen. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef]
- Gruber, A.; Kroth, P.G. Deducing intracellular distributions of metabolic pathways from genomic data. Methods Mol. Biol. 2014, 1083, 187–211. [Google Scholar] [CrossRef]
- Gschloessl, B.; Guermeur, Y.; Cock, J.M. HECTAR: A method to predict subcellular targeting in heterokonts. BMC Bioinform. 2008, 9, 393. [Google Scholar] [CrossRef]
- Gould, S.B.; Sommer, M.S.; Kroth, P.G.; Gile, G.H.; Keeling, P.J.; Maier, U.G. Nucleus-to-nucleus gene transfer and protein retargeting into a remnant cytoplasm of cryptophytes and diatoms. Mol. Biol. Evol. 2006, 23, 2413–2422. [Google Scholar] [CrossRef]
- Sommer, M.S.; Gould, S.B.; Lehmann, P.; Gruber, A.; Przyborski, J.M.; Maier, U.-G. Der1-mediated Preprotein Import into the Periplastid Compartment of Chromalveolates? Mol. Biol. Evol. 2007, 24, 918–928. [Google Scholar] [CrossRef]
- Gruber, A.; Kroth, P.G. Intracellular metabolic pathway distribution in diatoms and tools for genome-enabled experimental diatom research. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160402. [Google Scholar] [CrossRef] [PubMed]
- Mulholland, M.R.; Rocha, A.M.; Boneillo, G.E. Incorporation of Leucine and Thymidine by Estuarine Phytoplankton: Implications for Bacterial Productivity Estimates. Estuaries Coasts 2011, 34, 310–325. [Google Scholar] [CrossRef]
- Tjaden, J.; Winkler, H.H.; Schwöppe, C.; Van Der Laan, M.; Möhlmann, T.; Neuhaus, H.E. Two Nucleotide Transport Proteins in Chlamydia trachomatis, One for Net Nucleoside Triphosphate Uptake and the Other for Transport of Energy. J. Bacteriol. 1999, 181, 1196–1202. [Google Scholar] [PubMed]
- Haferkamp, I.; Schmitz-Esser, S.; Linka, N.; Urbany, C.; Collingro, A.; Wagner, M.; Horn, M.; Neuhaus, H.E. A candidate NAD+ transporter in an intracellular bacterial symbiont related to Chlamydiae. Nature 2004, 432, 622–625. [Google Scholar] [CrossRef]
- Heinz, E.; Hacker, C.; Dean, P.; Mifsud, J.; Goldberg, A.V.; Williams, T.A.; Nakjang, S.; Gregory, A.; Hirt, R.P.; Lucocq, J.M.; et al. Plasma membrane-located purine nucleotide transport proteins are key components for host exploitation by microsporidian intracellular parasites. PLoS Pathog. 2014, 10, e1004547. [Google Scholar] [CrossRef]
- Dean, P.; Sendra, K.M.; Williams, T.A.; Watson, A.K.; Major, P.; Nakjang, S.; Kozhevnikova, E.; Goldberg, A.V.; Kunji, E.R.S.; Hirt, R.P.; et al. Transporter gene acquisition and innovation in the evolution of Microsporidia intracellular parasites. Nat. Commun. 2018, 9, 1709. [Google Scholar] [CrossRef]
- Schmitz-Esser, S.; Linka, N.; Collingro, A.; Beier, C.L.; Neuhaus, H.E.; Wagner, M.; Horn, M. ATP/ADP Translocases: A Common Feature of Obligate Intracellular Amoebal Symbionts Related to Chlamydiae and Rickettsiae. J. Bacteriol. 2004, 186, 683–691. [Google Scholar] [CrossRef]
- Vávra, J.; Lukeš, J. Chapter Four—Microsporidia and ‘The Art of Living Together’. In Advances in Parasitology; Rollinson, D., Ed.; Academic Press: San Diego, CA, USA, 2013; Volume 82, pp. 253–319. [Google Scholar] [CrossRef]
- Tsaousis, A.D.; Kunji, E.R.S.; Goldberg, A.V.; Lucocq, J.M.; Hirt, R.P.; Embley, T.M. A novel route for ATP acquisition by the remnant mitochondria of Encephalitozoon cuniculi. Nature 2008, 453, 553–556. [Google Scholar] [CrossRef]
- Dean, P.; Hirt, R.P.; Embley, T.M. Microsporidia: Why Make Nucleotides if You Can Steal Them? PLoS Pathog. 2016, 12, e1005870. [Google Scholar] [CrossRef] [PubMed]
- Bailleul, B.; Berne, N.; Murik, O.; Petroutsos, D.; Prihoda, J.; Tanaka, A.; Villanova, V.; Bligny, R.; Flori, S.; Falconet, D.; et al. Energetic coupling between plastids and mitochondria drives CO2 assimilation in diatoms. Nature 2015, 524, 366. [Google Scholar] [CrossRef] [PubMed]
- Ewe, D.; Tachibana, M.; Kikutani, S.; Gruber, A.; Bartulos, C.R.; Konert, G.; Kaplan, A.; Matsuda, Y.; Kroth, P.G. The intracellular distribution of inorganic carbon fixing enzymes does not support the presence of a C4 pathway in the diatom Phaeodactylum tricornutum. Photosynth. Res. 2018, 137, 263–280. [Google Scholar] [CrossRef] [PubMed]
- Fabris, M.; Matthijs, M.; Rombauts, S.; Vyverman, W.; Goossens, A.; Baart, G.J.E. The metabolic blueprint of Phaeodactylum tricornutum reveals a eukaryotic Entner-Doudoroff glycolytic pathway. Plant J. 2012, 70, 1004–1014. [Google Scholar] [CrossRef]
- Rio Bartulos, C.; Rogers, M.B.; Williams, T.A.; Gentekaki, E.; Brinkmann, H.; Cerff, R.; Liaud, M.-F.; Hehl, A.B.; Yarlett, N.R.; Gruber, A.; et al. Mitochondrial Glycolysis in a Major Lineage of Eukaryotes. Genome Biol. Evol. 2018, 10, 2310–2325. [Google Scholar] [CrossRef]
- Ball, S.G.; Subtil, A.; Bhattacharya, D.; Moustafa, A.; Weber, A.P.M.; Gehre, L.; Colleoni, C.; Arias, M.-C.; Cenci, U.; Dauvillée, D. Metabolic Effectors Secreted by Bacterial Pathogens: Essential Facilitators of Plastid Endosymbiosis? Plant Cell 2013, 25, 7–21. [Google Scholar] [CrossRef]
- Cenci, U.; Bhattacharya, D.; Weber, A.P.M.; Colleoni, C.; Subtil, A.; Ball, S.G. Biotic Host–Pathogen Interactions As Major Drivers of Plastid Endosymbiosis. Trends Plant Sci. 2017, 22, 316–328. [Google Scholar] [CrossRef]
- Dagan, T.; Roettger, M.; Stucken, K.; Landan, G.; Koch, R.; Major, P.; Gould, S.B.; Goremykin, V.V.; Rippka, R.; Tandeau de Marsac, N.; et al. Genomes of Stigonematalean Cyanobacteria (Subsection V) and the Evolution of Oxygenic Photosynthesis from Prokaryotes to Plastids. Genome Biol. Evol. 2012, 5, 31–44. [Google Scholar] [CrossRef]
- Domman, D.; Horn, M.; Embley, T.M.; Williams, T.A. Plastid establishment did not require a chlamydial partner. Nat. Commun. 2015, 6, 6421. [Google Scholar] [CrossRef]
- Ball, S.G.; Bhattacharya, D.; Qiu, H.; Weber, A.P.M. Commentary: Plastid establishment did not require a chlamydial partner. Front. Cell. Infect. Microbiol. 2016, 6. [Google Scholar] [CrossRef]
- Haferkamp, I.; Schmitz-Esser, S. The Plant Mitochondrial Carrier Family: Functional and Evolutionary Aspects. Front. Plant Sci. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.A.; Haferkamp, I.; Keeling, P.J. An ADP/ATP-specific mitochondrial carrier protein in the microsporidian Antonospora locustae. J. Mol. Biol. 2008, 375, 1249–1257. [Google Scholar] [CrossRef]
- Thuswaldner, S.; Lagerstedt, J.O.; Rojas-Stütz, M.; Bouhidel, K.; Der, C.; Leborgne-Castel, N.; Mishra, A.; Marty, F.; Schoefs, B.; Adamska, I.; et al. Identification, Expression, and Functional Analyses of a Thylakoid ATP/ADP Carrier from Arabidopsis. J. Biol. Chem. 2007, 282, 8848–8859. [Google Scholar] [CrossRef] [PubMed]
- Gigolashvili, T.; Geier, M.; Ashykhmina, N.; Frerigmann, H.; Wulfert, S.; Krueger, S.; Mugford, S.G.; Kopriva, S.; Haferkamp, I.; Flügge, U.-I. The Arabidopsis thylakoid ADP/ATP carrier TAAC has an additional role in supplying plastidic phosphoadenosine 5′-phosphosulfate to the cytosol. Plant Cell 2012, 24, 4187–4204. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.-C.; Zimmermann, K.; Schorr, S.; Landini, M.; Klemens, P.A.W.; Altensell, J.; Jung, M.; Krause, E.; Nguyen, D.; Helms, V.; et al. AXER is an ATP/ADP exchanger in the membrane of the endoplasmic reticulum. Nat. Comm. 2018, 9, 3489. [Google Scholar] [CrossRef] [PubMed]
- Major, P.; Sendra, K.M.; Dean, P.; Williams, T.A.; Watson, A.K.; Thwaites, D.T.; Embley, T.M.; Hirt, R.P. A new family of cell surface located purine transporters in Microsporidia and related fungal endoparasites. eLife 2019, 8, e47037. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Principles of protein and lipid targeting in secondary symbiogenesis: Euglenoid, dinoflagellate, and sporozoan plastid origins and the eukaryote family tree. J. Eukaryot. Microbiol. 1999, 46, 347–366. [Google Scholar] [CrossRef]
- Petersen, J.; Ludewig, A.K.; Michael, V.; Bunk, B.; Jarek, M.; Baurain, D.; Brinkmann, H. Chromera velia, endosymbioses and the rhodoplex hypothesis--plastid evolution in cryptophytes, alveolates, stramenopiles, and haptophytes (CASH lineages). Genome Biol. Evol. 2014, 6, 666–684. [Google Scholar] [CrossRef]
- Derelle, R.; López-García, P.; Timpano, H.; Moreira, D. A Phylogenomic Framework to Study the Diversity and Evolution of Stramenopiles (=Heterokonts). Mol. Biol. Evol. 2016, 33, 2890–2898. [Google Scholar] [CrossRef]
- Malyshev, D.A.; Dhami, K.; Lavergne, T.; Chen, T.; Dai, N.; Foster, J.M.; Correa, I.R., Jr.; Romesberg, F.E. A semi-synthetic organism with an expanded genetic alphabet. Nature 2014, 509, 385–388. [Google Scholar] [CrossRef]
- Dien, V.T.; Morris, S.E.; Karadeema, R.J.; Romesberg, F.E. Expansion of the genetic code via expansion of the genetic alphabet. Curr. Opin. Chem. Biol. 2018, 46, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Eremeeva, E.; Herdewijn, P. Non canonical genetic material. Curr. Opin. Biotechnol. 2019, 57, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lamb, B.M.; Feldman, A.W.; Zhou, A.X.; Lavergne, T.; Li, L.; Romesberg, F.E. A semisynthetic organism engineered for the stable expansion of the genetic alphabet. Proc. Natl. Acad. Sci. USA 2017, 114, 1317–1322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ptacin, J.L.; Fischer, E.C.; Aerni, H.R.; Caffaro, C.E.; San Jose, K.; Feldman, A.W.; Turner, C.R.; Romesberg, F.E. A semi-synthetic organism that stores and retrieves increased genetic information. Nature 2017, 551, 644. [Google Scholar] [CrossRef] [PubMed]
- Feldman, A.W.; Fischer, E.C.; Ledbetter, M.P.; Liao, J.-Y.; Chaput, J.C.; Romesberg, F.E. A Tool for the Import of Natural and Unnatural Nucleoside Triphosphates into Bacteria. J. Am. Chem. Soc. 2018, 140, 1447–1454. [Google Scholar] [CrossRef]
- Pezo, V.; Hassan, C.; Louis, D.; Sargueil, B.; Herdewijn, P.; Marlière, P. Metabolic Recruitment and Directed Evolution of Nucleoside Triphosphate Uptake in Escherichia coli. ACS Synth. Biol. 2018, 7, 1565–1572. [Google Scholar] [CrossRef]
- Huang, W.; Daboussi, F. Genetic and metabolic engineering in diatoms. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160411. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Name | Protein ID 1 | Targeting Prediction 2 | ASAFAP Motif Identified by Ast et al. [39] | N-Terminal Elongation Identified by Major et al. [41] | Estimated Length 3 | Estimated Size 4/kDa | Predicted Number of Trans-Membrane Helices 5 | Experimental Localisations; Comments |
|---|---|---|---|---|---|---|---|---|
| PtNTT1 | 49533 | Plastid, high confidence | TEA-FAP (yes) | Yes | 555 | 60.11 | 11 | Targeted to the innermost plastid membrane via the stroma [39]; Ensembl identifier 6 Phatr3_J49533 |
| PtNTT2 | 45145 | Not plastid, SignalP positive | ISA-TSS (no) | Yes | 553 | 59.98 | 11 | Plastid and nuclear envelope, ER cisternae, due to similarity with TpNTT2, identical sub cellular localization was suggested [39]; Ensembl identifier 6 Phatr3_J11615 |
| PtNTT3 | 50189 | Not plastid, SignalP negative | No signal peptide | Yes | 666 | 73.05 | 11 | Ensembl identifier 6 Phatr3_J50189; this model excludes the first nine residues |
| PtNTT4 | 46794 | Not plastid, SignalP negative | No signal peptide | Yes | 591 | 66.42 | 8 | Ensembl identifier 6 Phatr3_J46794 |
| PtNTT5 | 54110 | Not plastid, SignalP negative | No signal peptide | Yes | 547 | 58.32 | 7 | Probably located in ER cisternae, C-terminus important for targeting [42]; Ensembl identifier 6 Phatr3_J54110 |
| PtNTT6 | 54907 | Plastid, low confidence | VRA-LLP (yes) | Yes | 557 | 61.47 | 7 | Ensembl identifier 6 Phatr3_J54907; this model excludes the first 90 residues, and 35 residues of CDS in a postulated intron in the C-terminal region |
| TpNTT1 | 26364 | Plastid, high confidence | THG-FSP (yes) | Yes | 525 | 57.44 | 11 | Targeted to the innermost plastid membrane via the stroma [39], identified in plastid fractions via mass spectrometry [43] |
| TpNTT2 | 24462 | Not plastid, SignalP negative | SSA-EML (no) | Yes | 632 | 69.53 | 10 | Presequence not sufficient for plastid targeting, full-length fusion protein might be located in plastid membrane [39] |
| TpNTT3 | 270249 | Not plastid, SignalP positive | TEA-ALP (no) | Yes | 692 | 75.17 | 10 | Identified in plastid fractions via mass spectrometry [43] |
| TpNTT4 | 270251 | Not plastid, SignalP negative | No signal peptide | Yes | 678 | 75.22 | 9 | |
| TpNTT5 | 9770 | Not plastid, SignalP negative | No signal peptide | Yes | 838 | 91.81 | 8 | |
| TpNTT6 | 24709 | Not plastid, SignalP positive | SLA-HQH (no) | Yes | 548 | 59.21 | 8 | |
| TpNTT7 | 270255 | Not plastid, SignalP negative | No signal peptide | Yes | 544 | 58.60 | 8 | |
| TpNTT8 | 270253 | Not plastid, SignalP negative | No SP | Yes | 589 | 63.59 | 10 |
| Group of Organisms | Species/Strain | Name | Activity | Role | References |
|---|---|---|---|---|---|
| Diatoms | Thalassiosira pseudonana | NTT1 | Symport of ADP or ATP with protons | Net import of full set of nucleotides into the plastid | [39] |
| NTT2 | Exchange of (d)NTP against ATP | ||||
| Phaeodactylum tricornutum | NTT1 | Symport of ADP or ATP with protons | |||
| NTT2 | Exchange of (d)NTP against ATP | ||||
| NTT5 | Exchange of GTP, GDP, dATP or dGTP against ATP or ADP | Provision of GTP, GDP, dA/GTP and chemical energy to the ER lumen | [42] | ||
| Chlamydiae | Chlamydia trachomatis1 | Npt2Ct | Symport of ATP, CTP, UTP and GTP with protons | Net import of full set of ribo-nucleotides into the symbiont | [57] |
| Protochlamydia amoebophila UWE25 2 | NTT2 | Antiport of UTP, ATP, GTP and CTP | Net import of ribo-nucleotides and NAD+ into the symbiont | [58] | |
| NTT3 | Symport of UTP with protons | ||||
| NTT4 | Exchange of NAD+ with ADP | ||||
| NTT5 | Symport of GTP or ATP with protons | ||||
| Microsporidia | Trachipleistophora hominis | NTT1, NTT2, NTT3 | Antiport of ATP/GTP against ADP/GDP | Energy parasitism, transport of NAD+ and NADH | [59,60] |
| NTT4 | Symport of GTP or ATP with protons | Net import of purine nucleotides |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gruber, A.; Haferkamp, I. Nucleotide Transport and Metabolism in Diatoms. Biomolecules 2019, 9, 761. https://doi.org/10.3390/biom9120761
Gruber A, Haferkamp I. Nucleotide Transport and Metabolism in Diatoms. Biomolecules. 2019; 9(12):761. https://doi.org/10.3390/biom9120761
Chicago/Turabian StyleGruber, Ansgar, and Ilka Haferkamp. 2019. "Nucleotide Transport and Metabolism in Diatoms" Biomolecules 9, no. 12: 761. https://doi.org/10.3390/biom9120761
APA StyleGruber, A., & Haferkamp, I. (2019). Nucleotide Transport and Metabolism in Diatoms. Biomolecules, 9(12), 761. https://doi.org/10.3390/biom9120761