

Assessing Adventitious Matches of Non-Donors Related to True Contributors


Abstract
1. Introduction
2. Materials and Methods
2.1. DNA Mixtures
2.2. STRmixTM
2.3. FaSTRTM
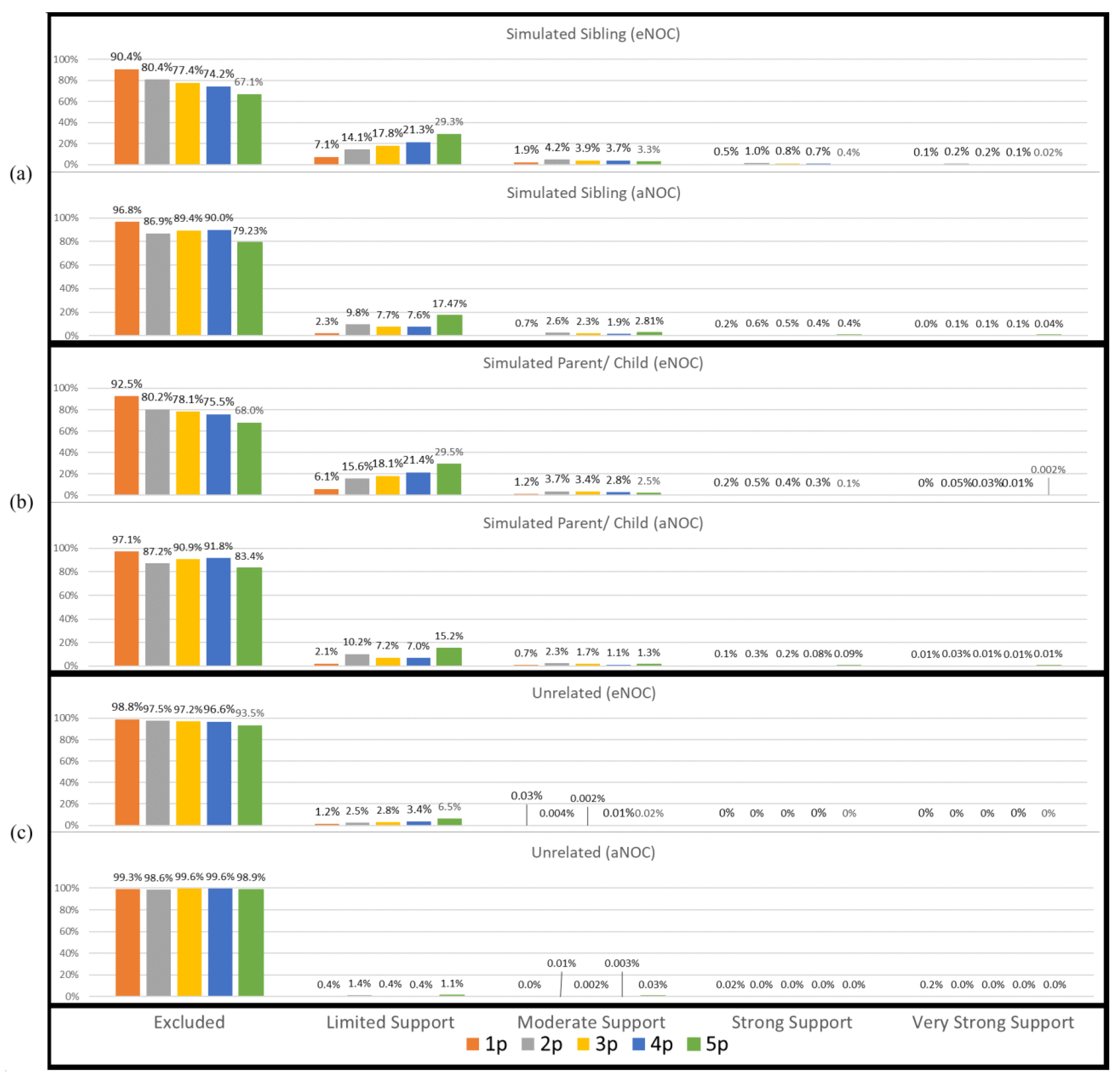
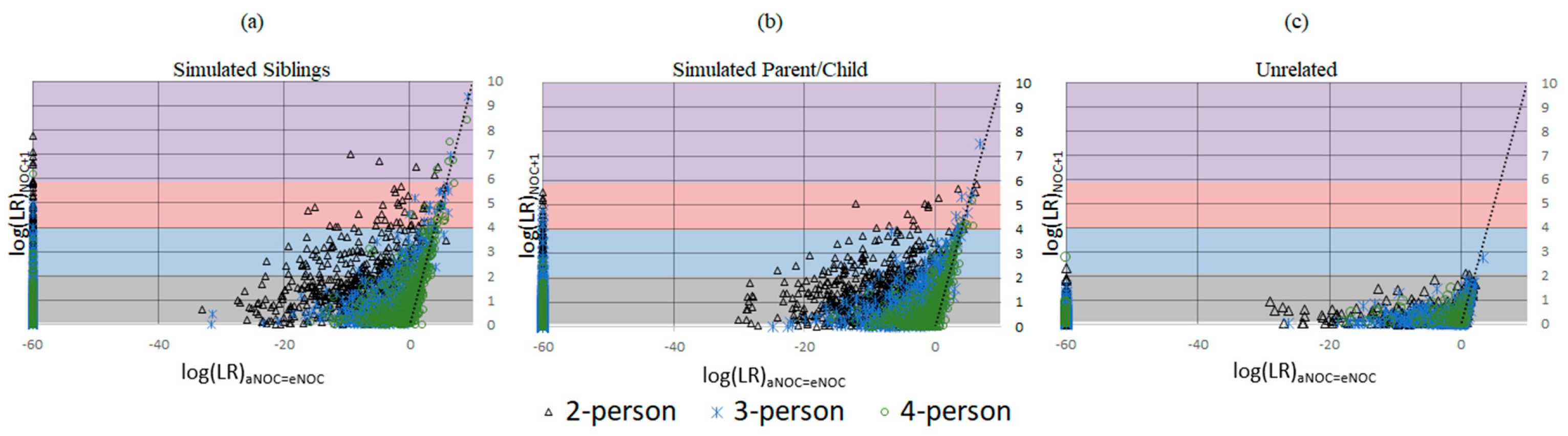
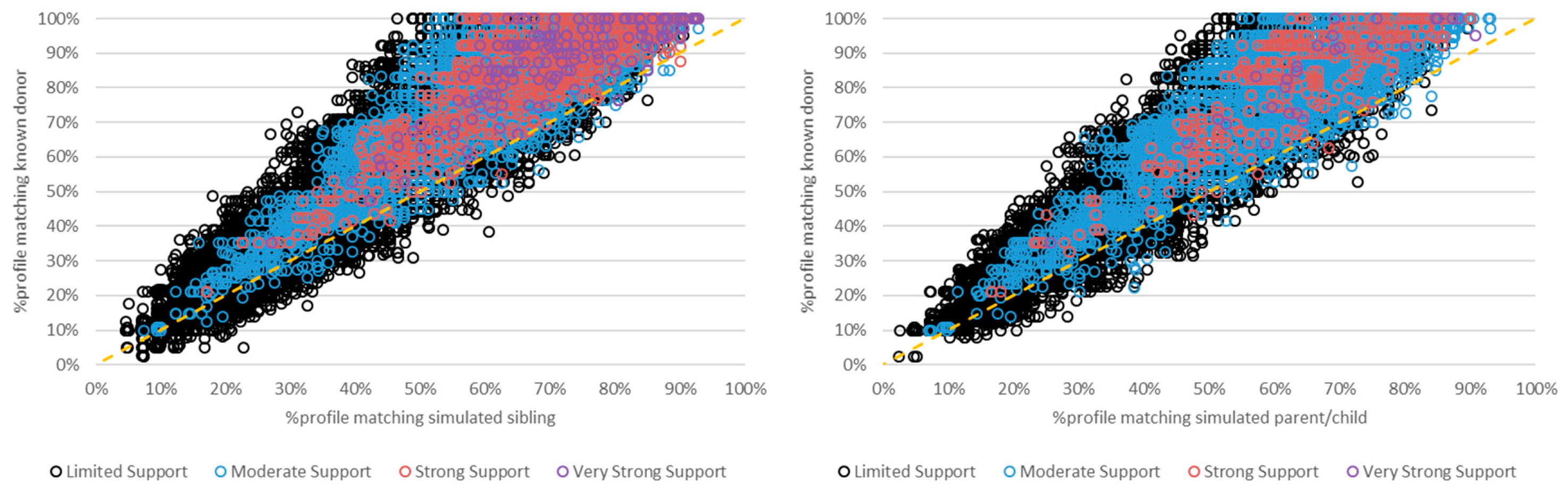
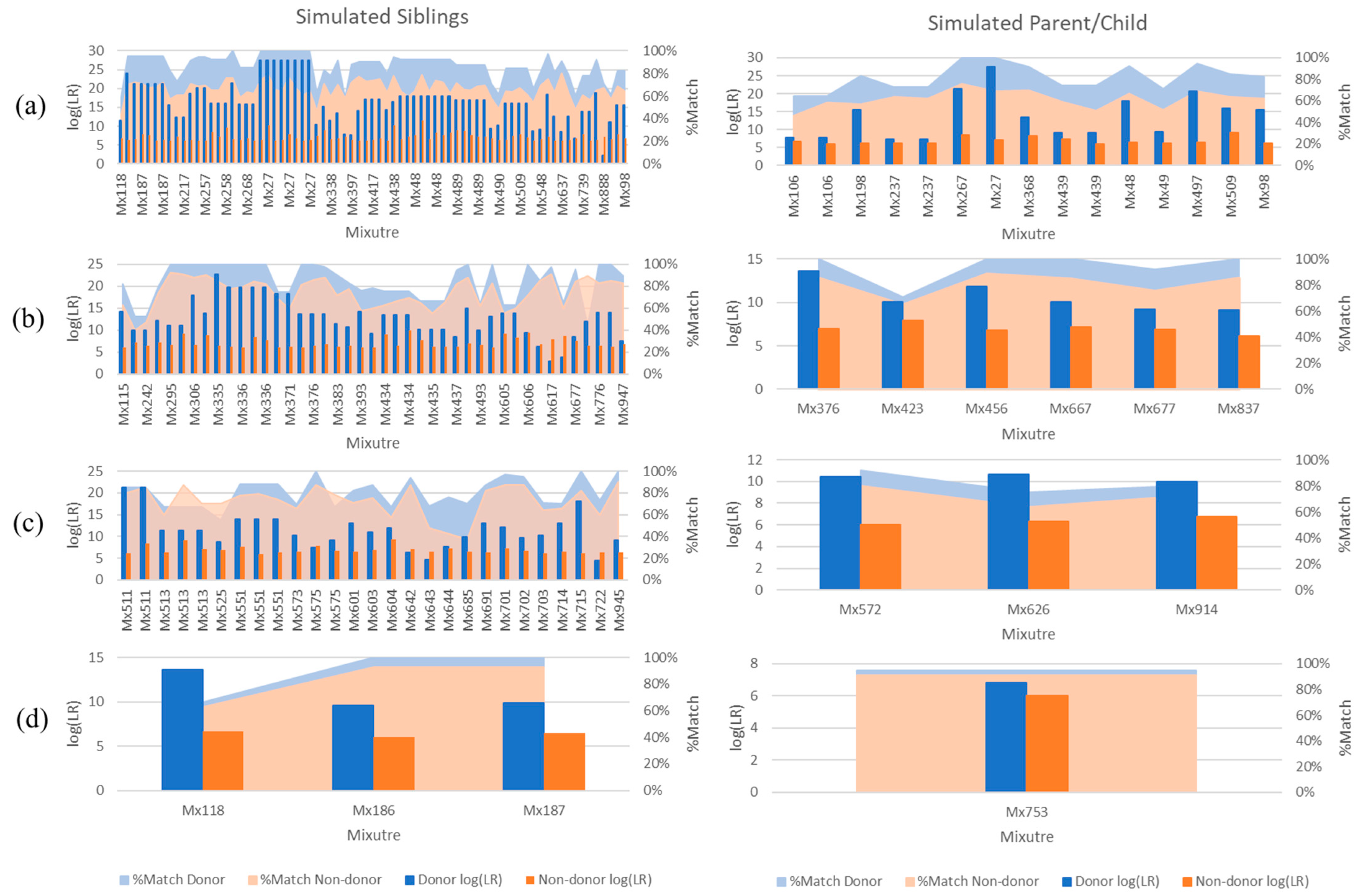
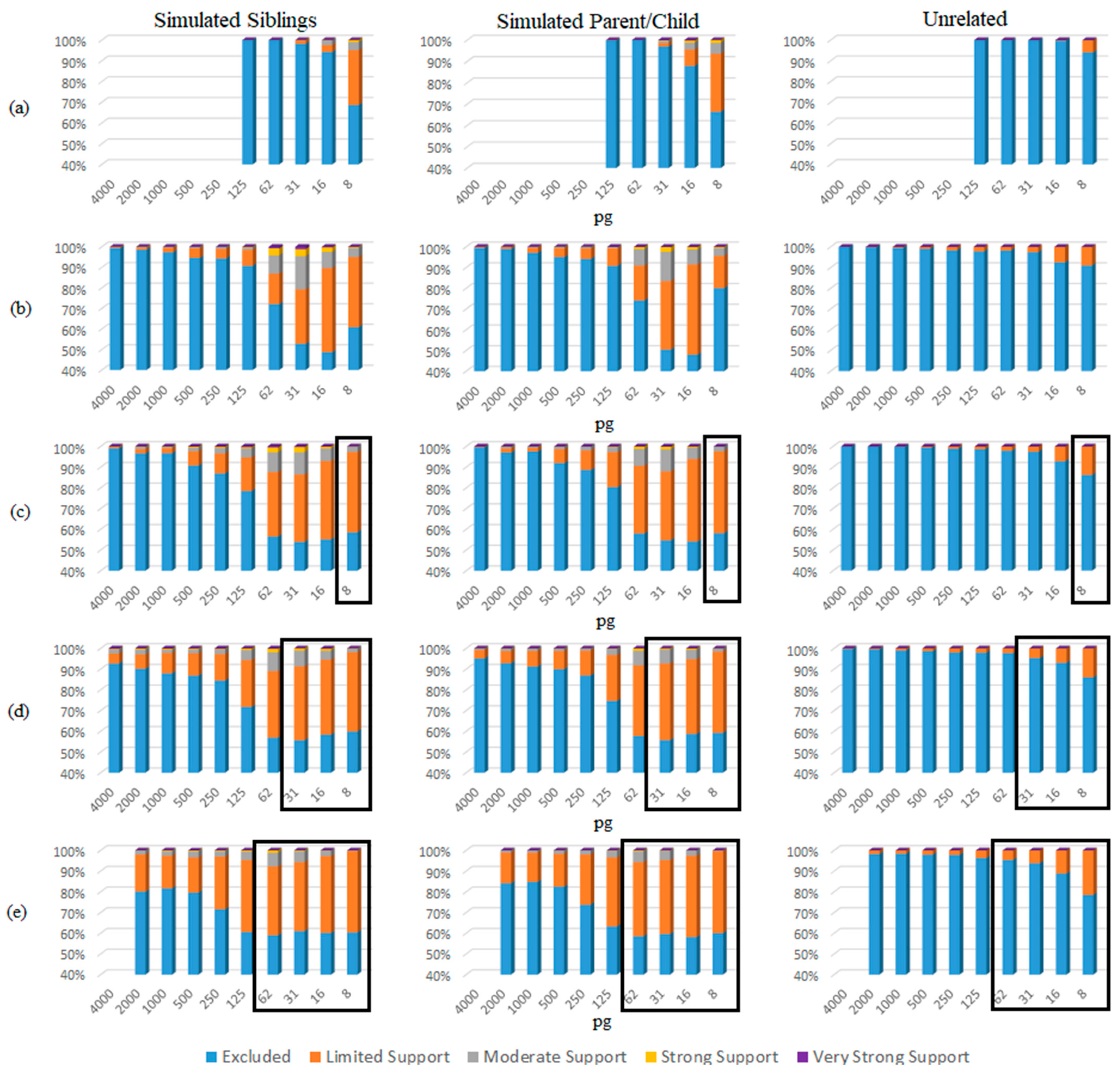
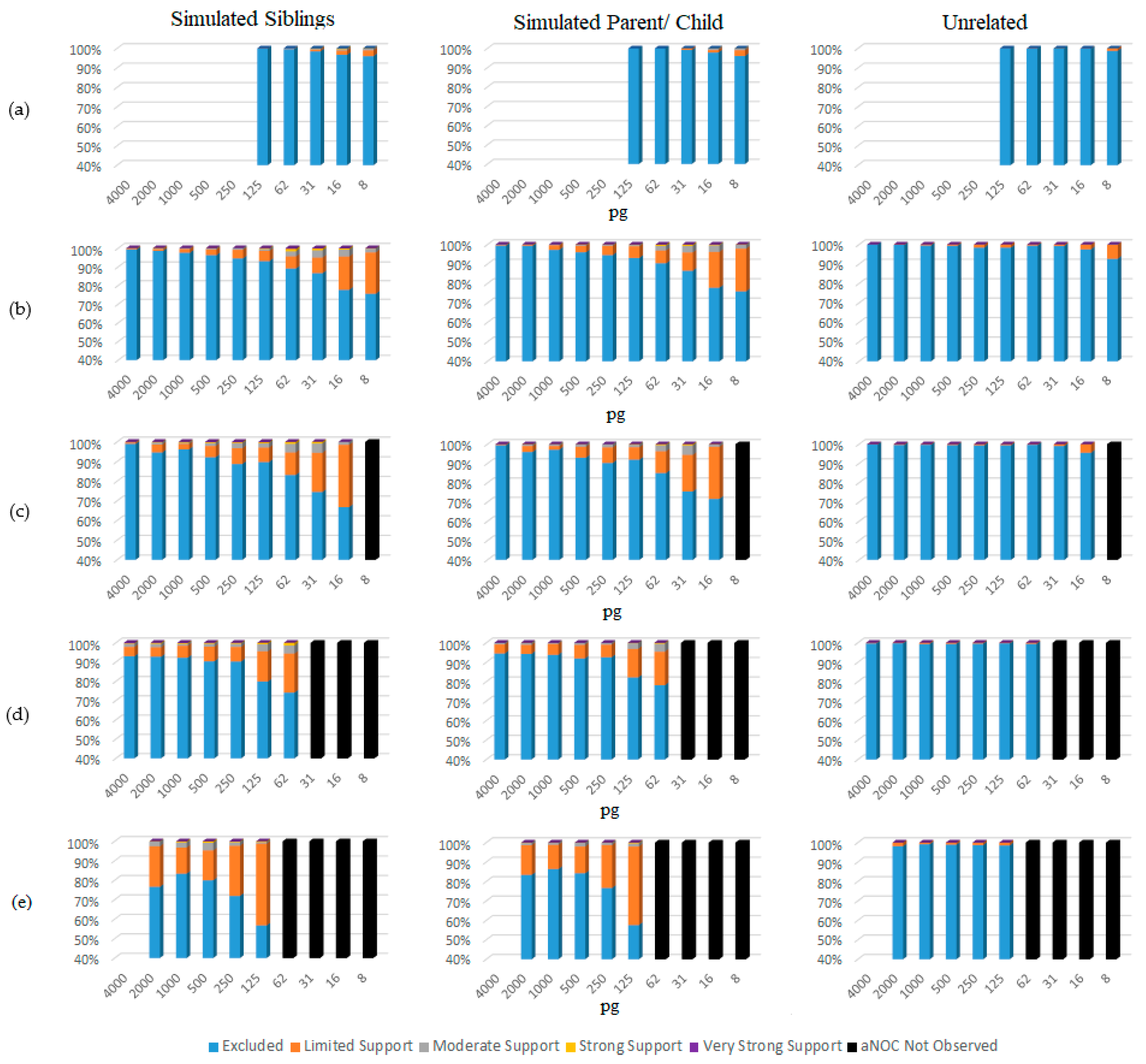
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, M.-H.; Bright, J.-A.; Pugh, S.N.; Buckleton, J.S. The interpretation of mixed DNA profiles from a mother, father, and child trio. Forensic Sci. Int. Genet. 2020, 44, 102175. [Google Scholar] [CrossRef] [PubMed]
- Kalafut, T.; Bright, J.-A.; Taylor, D.; Buckleton, J. Investigation into the effect of mixtures comprising related people on non-donor likelihood ratios, and potential practises to mitigate providing misleading opinions. Forensic Sci. Int. Genet. 2022, 59, 102691. [Google Scholar] [CrossRef] [PubMed]
- Kelly, H.; Coble, M.; Kruijver, M.; Wivell, R.; Bright, J. Exploring likelihood ratios assigned for siblings of the true mixture contributor as an alternate contributor. J. Forensic Sci. 2022, 67, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Oregon State Police Forensic Sciences Division Portland Metro Laboratory. Validation STR Casework Analysis Using GlobalFiler, the 3500xl, and STRmix. In Validation Study for STR Analysis; Oregon State Police Forensic Sciences Division Portland Metro Laboratory: Portland, OR, USA, 2016; Available online: https://indefenseof.us/uploads/Oregon-State-Police-Portland-Metro-Lab-DNA-Val-067-GlobalFiler-STRmix-Summary_Redacted.pdf (accessed on 1 July 2024).
- Kelly, H.; Bright, J.; Coble, M.D.; Buckleton, J.S. A description of the likelihood ratios in the probabilistic genotyping software STRmixTM. WIREs Forensic Sci. 2020, 2, e1377. [Google Scholar] [CrossRef]
- Huffman, K.; Ballantyne, J. Probabilistic Genotyping of Single Cell Replicates from Mixtures Involving First-Degree Relatives Prevents the False Inclusions of Non-Donor Relatives. Genes 2022, 13, 1658. [Google Scholar] [CrossRef] [PubMed]
- Sacramento County District Attorney’s Crime Laboratory. Internal Validation of STRmixTM V2.4. 2017. Available online: https://indefenseof.us/uploads/Sacramento-Cty-DA-STRmix-V2-4-internal-validation-summary.pdf (accessed on 1 July 2024).
- Los Angeles County Sheriff’s Department Scientific Services Bureau Biology Section. Validation of STRmixTM v. 2.5.11 Using the POWERPLEX FUSION 6C KIT. 2017. Available online: https://indefenseof.us/uploads/LASD-STRmix-2.5.11-Validation-Summary.pdf (accessed on 1 July 2024).
- Jefferson County Regional Crime Laboratory. Internal Validation of STRmixTM V2.6 for the Analysis of GlobalFilerTM Profiles. 2019. Available online: https://indefenseof.us/uploads/Jefferson-County-STRmix-Validation-V2.6-V2.6.3.pdf (accessed on 1 July 2024).
- Las Vegas Metropolitan Police Department. Internal Validation of STRmixTM v. 2.6 (QIAGEN Investigator 24plex QS with 3500xl). 2020. Available online: https://indefenseof.us/uploads/LVMPD-Summary.pdf (accessed on 1 July 2024).
- Wisconsin State Crime Laboratory. Internal Validation Summary for STRmixTM Probabilistic Genotyping Software. Available online: https://indefenseof.us/uploads/Wisconsin-STRmix-Validation-Summary-Part-1-Single-Single-Source-to-Three-Person-Mixtures.pdf (accessed on 1 July 2024).
- Buckleton, J.S.; Pugh, S.N.; Bright, J.-A.; Taylor, D.A.; Curran, J.M.; Kruijver, M.; Gill, P.; Budowle, B.; Cheng, K. Are low LRs reliable? Forensic Sci. Int. Genet. 2020, 49, 102350. [Google Scholar] [CrossRef] [PubMed]
- Benschop, C.C.; Nijveld, A.; Duijs, F.E.; Sijen, T. An assessment of the performance of the probabilistic genotyping software EuroForMix: Trends in likelihood ratios and analysis of Type I & II errors. Forensic Sci. Int. Genet. 2019, 42, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Bright, J.-A.; Richards, R.; Kruijver, M.; Kelly, H.; McGovern, C.; Magee, A.; McWhorter, A.; Ciecko, A.; Peck, B.; Baumgartner, C.; et al. Internal validation of STRmixTM—A multi laboratory response to PCAST. Forensic Sci. Int. Genet. 2018, 34, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Moretti, T.R.; Just, R.S.; Kehl, S.C.; Willis, L.E.; Buckleton, J.S.; Bright, J.-A.; Taylor, D.A.; Onorato, A.J. Onorato, Internal validation of STRmixTM for the interpretation of single source and mixed DNA profiles. Forensic Sci. Int. Genet. 2017, 29, 126–144. [Google Scholar] [CrossRef] [PubMed]
- Coble, M.D.; Bright, J.-A.; Buckleton, J.S.; Curran, J.M. Uncertainty in the number of contributors in the proposed new CODIS set. Forensic Sci. Int. Genet. 2015, 19, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Buckleton, J.S.; Bright, J.; Gittelson, S.; Moretti, T.R.; Onorato, A.J.; Bieber, F.R.; Budowle, B.; Taylor, D.A. The Probabilistic Genotyping Software STRmix: Utility and Evidence for its Validity. J. Forensic Sci. 2019, 64, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Kalafut, T.; Pugh, S.; Gill, P.; Abbas, S.; Semaan, M.; Mansour, I.; Curran, J.; Bright, J.; Hicks, T.; Wivell, R.; et al. A mixed DNA profile controversy revisited. J. Forensic Sci. 2022, 67, 128–135. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NOC | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| Samples | 78 | 230 | 236 | 232 | 190 |
| Simulated children | 50 | 50 | 50 | 50 | 50 |
| Simulated siblings | 50 | 50 | 50 | 50 | 50 |
| Hd true tests per relative type | 3900 | 23,000 | 35,400 | 46,400 | 47,500 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huffman, K.; Moreno, L. Assessing Adventitious Matches of Non-Donors Related to True Contributors. Biomolecules 2025, 15, 398. https://doi.org/10.3390/biom15030398
Huffman K, Moreno L. Assessing Adventitious Matches of Non-Donors Related to True Contributors. Biomolecules. 2025; 15(3):398. https://doi.org/10.3390/biom15030398
Chicago/Turabian StyleHuffman, Kaitlin, and Lilliana Moreno. 2025. "Assessing Adventitious Matches of Non-Donors Related to True Contributors" Biomolecules 15, no. 3: 398. https://doi.org/10.3390/biom15030398
APA StyleHuffman, K., & Moreno, L. (2025). Assessing Adventitious Matches of Non-Donors Related to True Contributors. Biomolecules, 15(3), 398. https://doi.org/10.3390/biom15030398