
Phenolic-Rich Extracts from Avocado Fruit Residues as Functional Food Ingredients with Antioxidant and Antiproliferative Properties

 ,
, 
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Avocado Peel, Seed Coat and Seed Extracts
2.2. Chemical Characterization of the Extracts
2.3. Determination of Antioxidant Activity of Avocado Extracts
2.4. Profile of Phenolic Compounds in the Extracts
2.5. Macronutrients and Energy Content of Extract
2.6. Cell Culture, Cell Treatment and Determination of Citotoxicity
2.7. Cell Death Studies
2.8. Flow Cytometry Mitochondrial Membrane Potential Assay
2.9. Determination of Intracellular Levels of Reactive Oxygen Species (ROS)
2.10. Theoretical Absorption Percentage of Individual Phenolic Compounds
2.11. Statistical Analysis
3. Results and Discussion
3.1. Chemical Composition and Antioxidant Activity
3.2. Quantification of the Individual Phenolic Compounds in Avocado Peel, Seed Coat and Seed Extracts by UPLC-ESI-MS/MS
3.3. Macronutrients and Energy Value of Extracts
3.4. Effect of Peel, Seed Coat and Seed Avocado Extracts on Colonorectal Cancer Cells (Caco-2 Cells)
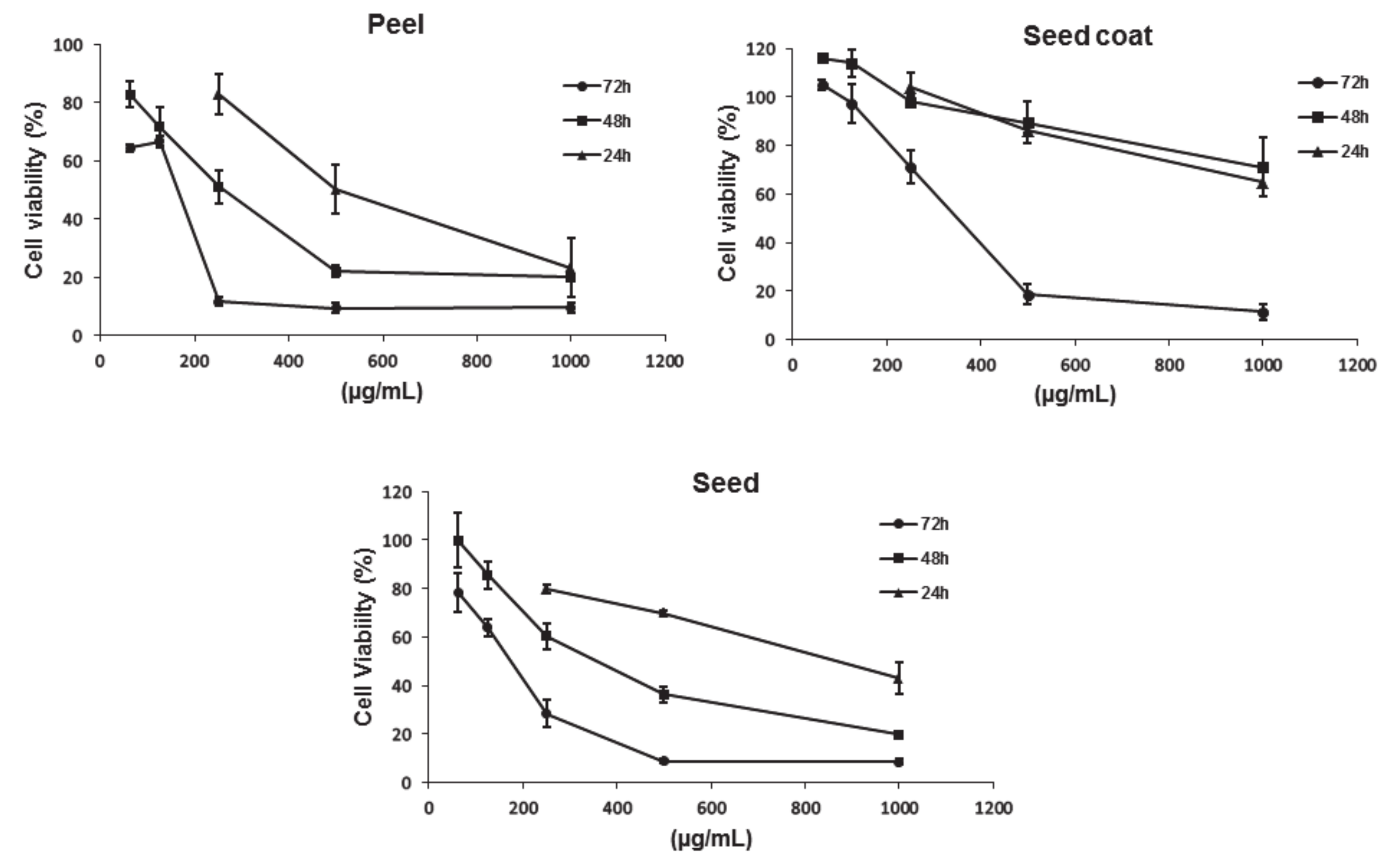
3.4.1. Antiproliferative Activity
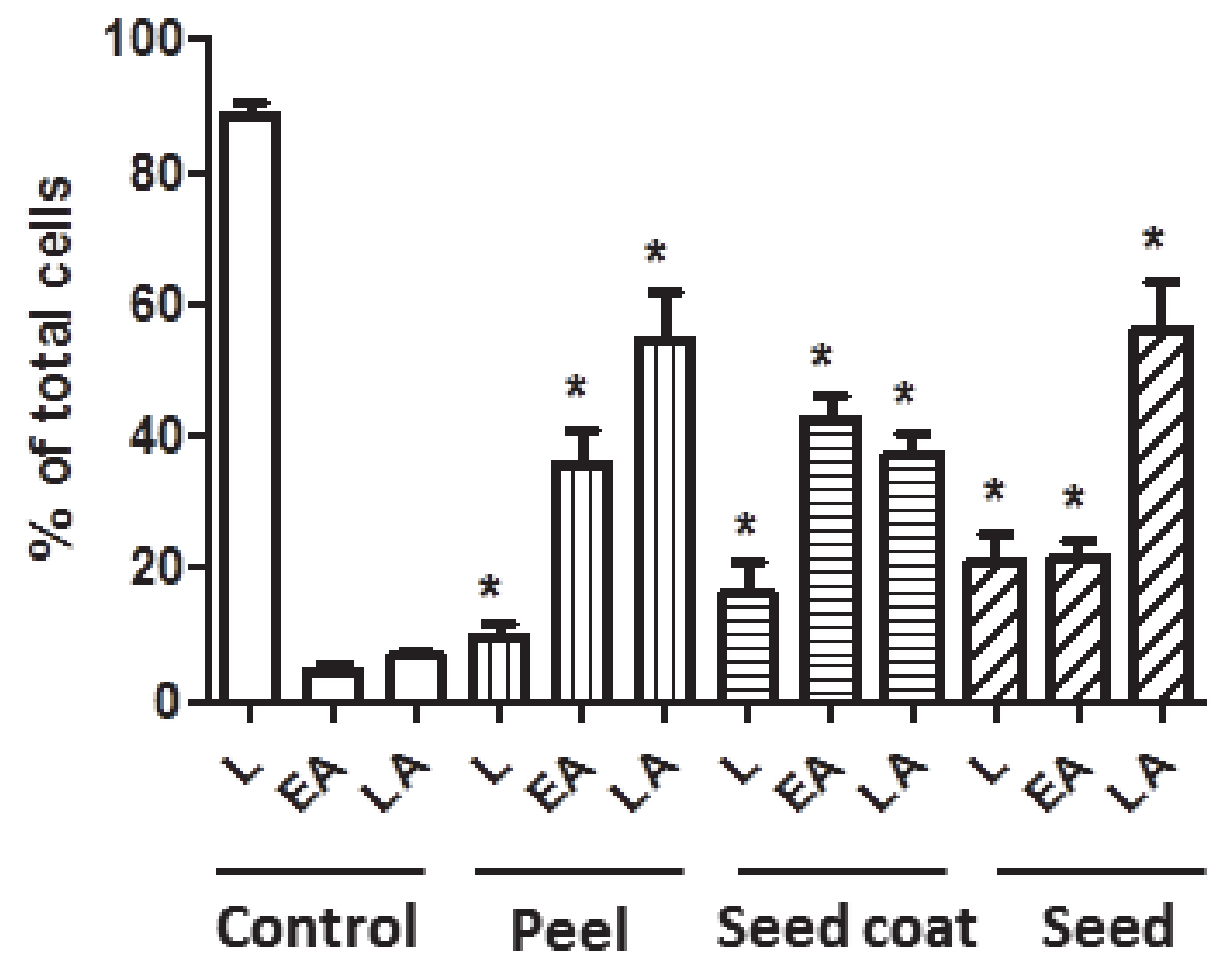
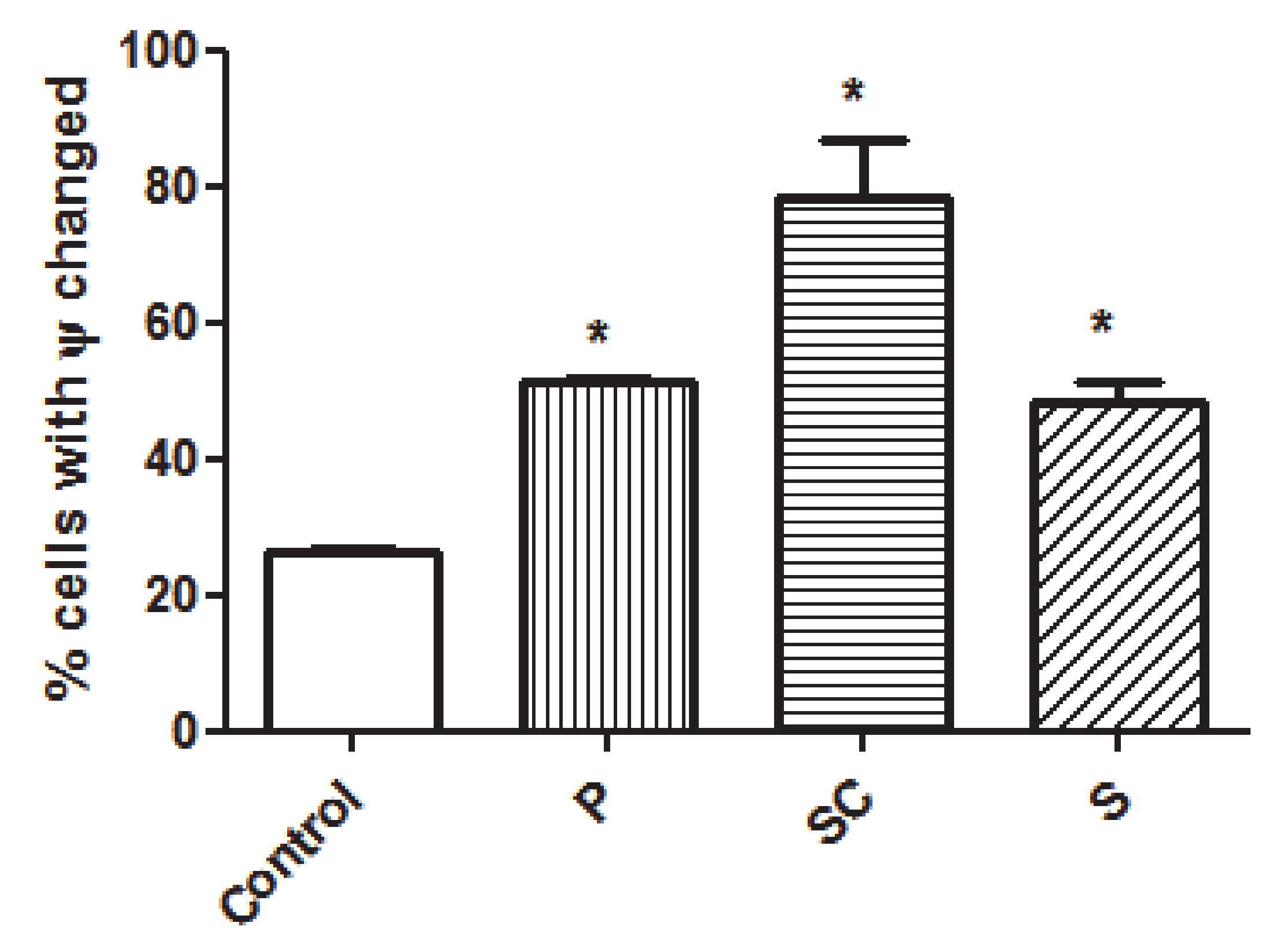
3.4.2. Apoptosis Analysis
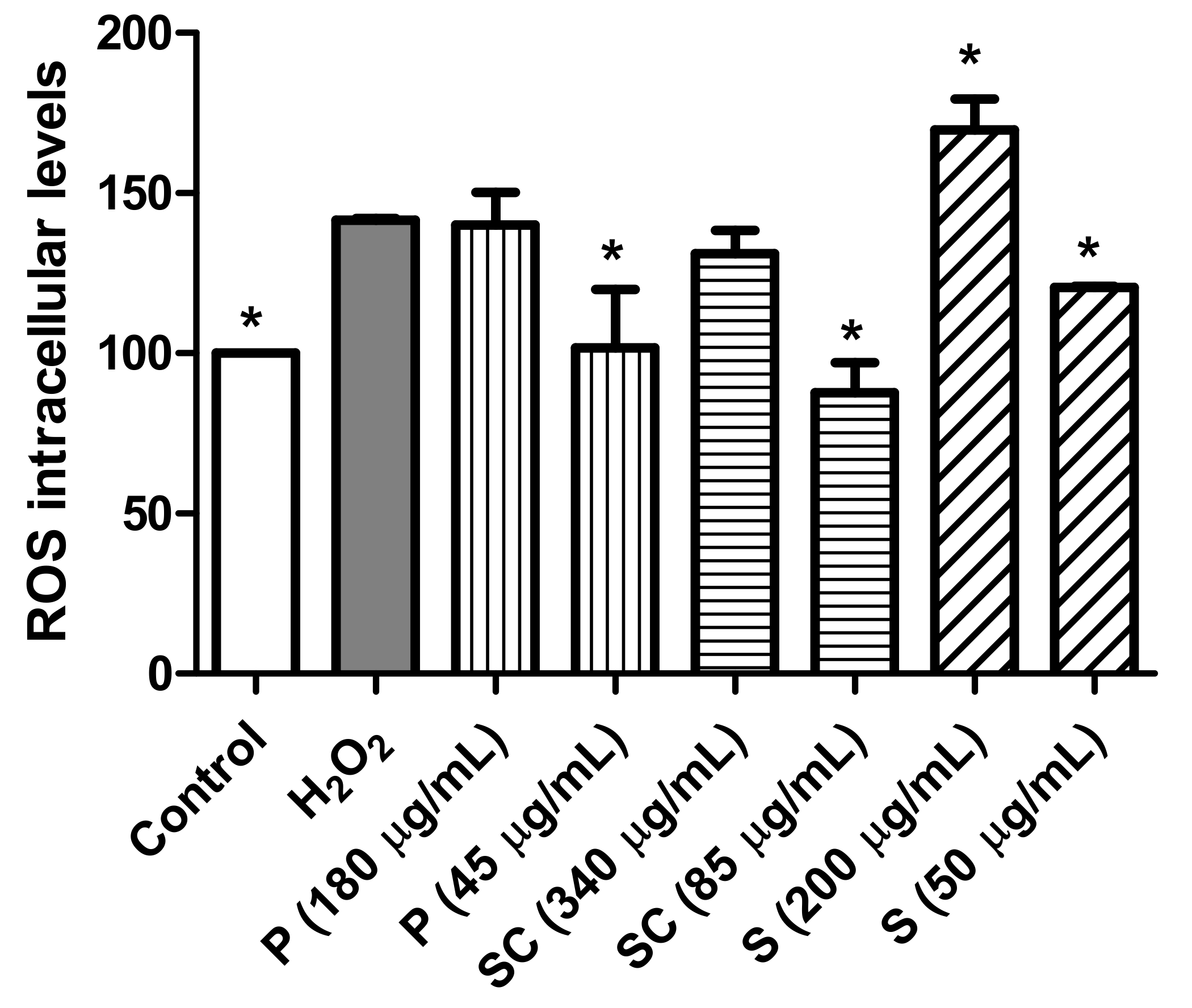
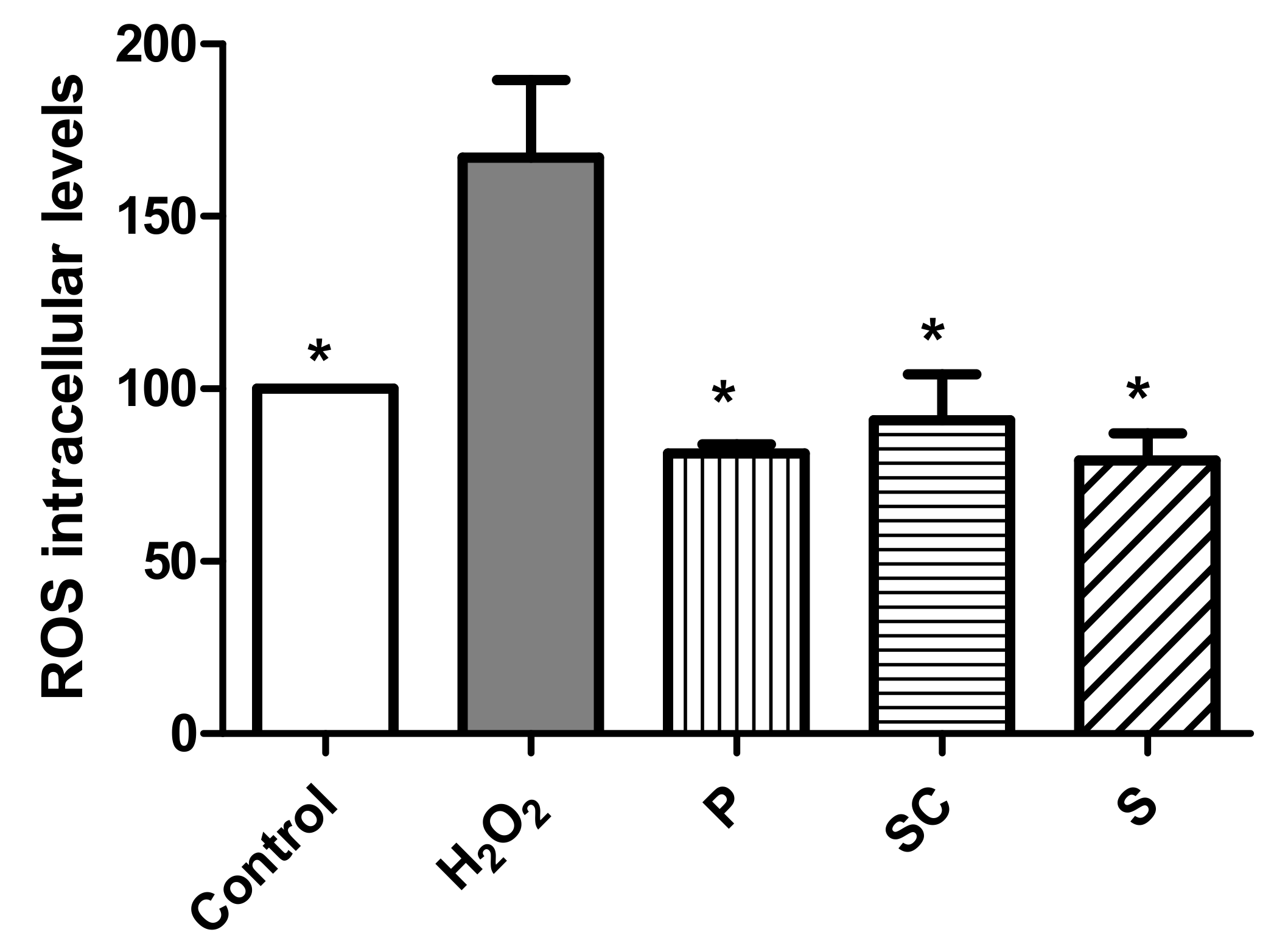
3.4.3. Effect of Avocado Extracts on ROS Intracellular Levels
3.4.4. Antioxidant Activity of Avocado Extracts on a Model Intestinal Barrier
3.4.5. Theoretical Absorption Percentage of Individual Phenolic Compounds (Based on Lipinski Parameters)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Velderrain-Rodríguez, G.R.; Salvia-Trujillo, L.; González-Aguilar, G.A.; Martín-Belloso, O. Interfacial activity of phenolic-rich extracts from avocado fruit waste: Influence on the colloidal and oxidative stability of emulsions and nanoemulsions. Innov. Food Sci. Emerg. Technol. 2021, 69, 102665. [Google Scholar] [CrossRef]
- Araújo, R.G.; Rodriguez-Jasso, R.M.; Ruiz, H.A.; Pintado, M.M.E.; Aguilar, C.N. Avocado by-products: Nutritional and functional properties. Trends Food Sci. Technol. 2018, 80, 51–60. [Google Scholar] [CrossRef]
- Alkhalaf, M.I.; Alansari, W.S.; Ibrahim, E.A.; Elhalwagy, M.E.A. Anti-oxidant, anti-inflammatory and anti-cancer activities of avocado (Persea americana) fruit and seed extract. J. King Saud. Univ. Sci. 2019, 31, 1358–1362. [Google Scholar] [CrossRef]
- Figueroa, J.G.; Borrás-Linares, I.; Del Pino-García, R.; Curiel, J.A.; Lozano-Sánchez, J.; Segura-Carretero, A. Functional ingredient from avocado peel: Microwave-assisted extraction, characterization and potential applications for the food industry. Food Chem. 2021, 352, 129300. [Google Scholar] [CrossRef] [PubMed]
- Cerda-Opazo, P.; Gotteland, M.; Oyarzun-Ampuero, F.A.; Garcia, L. Design, development and evaluation of nanoemulsion containing avocado peel extract with anticancer potential: A novel biological active ingredient to enrich food. Food Hydrocoll. 2021, 111, 106370. [Google Scholar] [CrossRef]
- Figueroa, J.G.; Borrás-Linares, I.; Lozano-Sánchez, J.; Segura-Carretero, A. Comprehensive characterization of phenolic and other polar compounds in the seed and seed coat of avocado by HPLC-DAD-ESI-QTOF-MS. Food Res. Int. 2018, 105, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Bahru, T.B.; Tadele, Z.H.; Ajebe, E.G. A Review on Avocado Seed: Functionality, Composition, Antioxidant and Antimicrobial Properties. Chem. Sci. Int. J. 2019, 27, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tavsan, Z.; Kayali, H.A. Flavonoids showed anticancer effects on the ovarian cancer cells: Involvement of reactive oxygen species, apoptosis, cell cycle and invasion. Biomed. Pharmacother. 2019, 116, 109004. [Google Scholar] [CrossRef]
- Lee, S.-G.; Yu, M.-H.; Lee, S.-P.; Lee, I.-S. Antioxidant activities and induction of apoptosis by methanol extracts from avocado. J. Korean Soc. Food Sci. Nutr. 2008, 37, 269–275. [Google Scholar] [CrossRef]
- Silvan, J.M.; Gutiérrez-Docio, A.; Moreno-Fernandez, S.; Alarcón-Cavero, T.; Prodanov, M.; Martinez-Rodriguez, A.J. Procyanidin-rich extract from grape seeds as a putative tool against Helicobacter pylori. Foods 2020, 9, 1370. [Google Scholar] [CrossRef]
- Li, M.; Feng, L.; Jiang, W.-D.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Zhou, X.-Q. Condensed tannins decreased the growth performance and impaired intestinal immune function in on-growing grass carp (Ctenopharyngodon idella). Br. J. Nutr. 2020, 123, 737–755. [Google Scholar] [CrossRef]
- Mazzucotelli, C.A.; González-Aguilar, G.A.; Villegas-Ochoa, M.A.; Domínguez-Avila, A.J.; Ansorena, M.R.; Di Scala, K.C. Chemical characterization and functional properties of selected leafy vegetables for innovative mixed salads. J. Food Biochem. 2018, 42, e12461. [Google Scholar] [CrossRef] [Green Version]
- Teixé-Roig, J.; Oms-Oliu, G.; Velderrain-Rodríguez, G.; Odriozola-Serrano, I.; Martín-Belloso, O. The effect of sodium carboxymethylcellulose on the stability and bioaccessibility of anthocyanin water-in-oil-in-water emulsions. Food Bioprocess Technol. 2018, 11, 2229–2241. [Google Scholar] [CrossRef]
- López-Gámez, G.; Elez-Martínez, P.; Quiles-Chuliá, A.; Martín-Belloso, O.; Hernando-Hernando, I.; Soliva-Fortuny, R. Effect of pulsed electric fields on carotenoid and phenolic bioaccessibility and their relationship with carrot structure. Food Funct. 2021, 12, 2772–2783. [Google Scholar] [CrossRef]
- Enneb, S.; Drine, S.; Bagues, M.; Triki, T.; Boussora, F.; Guasmi, F.; Nagaz, K.; Ferchichi, A. Phytochemical profiles and nutritional composition of squash (Cucurbita moschata D.) from Tunisia. South Afr. J. Bot. 2020, 130, 165–171. [Google Scholar] [CrossRef]
- Chua, G.K.; Tan, F.H.Y.; Chew, F.N.; Mohd-Hairul, A.R.; Ahmad, M.A.A. Food waste hydrolysate as fermentation medium: Comparison of pre-treatment methods. Mater. Today Proc. 2021, 42, 131–137. [Google Scholar] [CrossRef]
- Ferreira, I.; Gomes-Bispo, A.; Lourenço, H.; Matos, J.; Afonso, C.; Cardoso, C.; Castanheira, I.; Motta, C.; Prates, J.A.M.; Bandarra, N.M. The chemical composition and lipid profile of the chub mackerel (Scomber colias) show a strong seasonal dependence: Contribution to a nutritional evaluation. Biochimie 2020, 178, 181–189. [Google Scholar] [CrossRef]
- Demoliner, F.; de Britto Policarpi, P.; Ramos, J.C.; Bascuñan, V.L.A.F.; Ferrari, R.A.; Jachmanián, I.; de Francisco de Casas, A.; Vasconcelos, L.F.L.; Block, J.M. Sapucaia nut (Lecythis pisonis Cambess) and its by-products: A promising and underutilized source of bioactive compounds. Part I: Nutritional composition and lipid profile. Food Res. Int. 2018, 108, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Gascón, S.; Jiménez-Moreno, N.; Jiménez, S.; Quero, J.; Rodríguez-Yoldi, M.J.; Ancín-Azpilicueta, C. Nutraceutical composition of three pine bark extracts and their antiproliferative effect on Caco-2 cells. J. Funct. Foods 2018, 48, 420–429. [Google Scholar] [CrossRef]
- Allaoui, A.; Barranquero, C.; Yahia, S.; Herrera-Marcos, L.V.; Benomar, S.; Jridi, M.; Navarro, M.Á.; Rodriguez-Yoldi, M.J.; Nasri, M.; Osada, J.; et al. Fenugreek proteins and their hydrolysates prevent hypercholesterolemia and enhance the HDL antioxidant properties in rats. Nutr. Food Sci. 2018, 48, 973–989. [Google Scholar] [CrossRef] [Green Version]
- Quero, J.; Jiménez-Moreno, N.; Esparza, I.; Osada, J.; Cerrada, E.; Ancín-Azpilicueta, C.; Rodríguez-Yoldi, M.J. Grape Stem Extracts with Potential Anticancer and Antioxidant Properties. Antioxidants 2021, 10, 243. [Google Scholar] [CrossRef] [PubMed]
- Uzarski, J.S.; DiVito, M.D.; Wertheim, J.A.; Miller, W.M. Essential design considerations for the resazurin reduction assay to noninvasively quantify cell expansion within perfused extracellular matrix scaffolds. Biomaterials 2017, 129, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-de-Diego, C.; Marmol, I.; Perez, R.; Gascon, S.; Rodriguez-Yoldi, M.J.; Cerrada, E. The anticancer effect related to disturbances in redox balance on Caco-2 cells caused by an alkynyl gold(I) complex. J. Inorg. Biochem. 2017, 166, 108–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NCBI. PubChem Open Chemistry Database. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 12 June 2021).
- Ertl, P.; Schuffenhauer, A. Estimation of synthetic accessibility score of drug-like molecules based on molecular complexity and fragment contributions. J. Cheminform. 2009, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Colombo, R.; Papetti, A. Avocado (Persea americana Mill.) by-products and their impact: From bioactive compounds to biomass energy and sorbent material for removing contaminants. A review. Int. J. Food Sci. Technol. 2019, 54, 943–951. [Google Scholar] [CrossRef]
- De Souza, E.L.; de Albuquerque, T.M.R.; dos Santos, A.S.; Massa, N.M.L.; de Brito Alves, J.L. Potential interactions among phenolic compounds and probiotics for mutual boosting of their health-promoting properties and food functionalitie—A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1645–1659. [Google Scholar] [CrossRef]
- Coklar, H.; Akbulut, M. Changes in phenolic acids, flavonoids, anthocyanins, and antioxidant activities of Mahonia aquifolium berries during fruit development and elucidation of the phenolic biosynthetic pathway. Hortic. Environ. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Velderrain-Rodríguez, G.R.; Torres-Moreno, H.; Villegas-Ochoa, M.A.; Ayala-Zavala, J.F.; Robles-Zepeda, R.E.; Wall-Medrano, A.; González-Aguilar, G.A. Gallic Acid Content and an Antioxidant Mechanism Are Responsible for the Antiproliferative Activity of ‘Ataulfo’ Mango Peel on LS180 Cells. Molecules 2018, 23, 695. [Google Scholar] [CrossRef] [Green Version]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant Activity of Polyphenols, Flavonoids, Anthocyanins and Carotenoids: Updated Review of Mechanisms and Catalyzing Metals. Phytother. Res. PTR 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Pedan, V.; Stamm, E.; Do, T.; Holinger, M.; Reich, E. HPTLC fingerprint profile analysis of coffee polyphenols during different roast trials. J. Food Compos. Anal. 2020, 94, 103610. [Google Scholar] [CrossRef]
- Martinez-Micaelo, N.; González-Abuín, N.; Ardèvol, A.; Pinent, M.; Blay, M.T. Procyanidins and inflammation: Molecular targets and health implications. BioFactors 2012, 38, 257–265. [Google Scholar] [CrossRef]
- Kumar, K.; Issac, A.; Ninan, E.; Kuttan, R.; Maliakel, B. Enhanced anti-diabetic activity of polyphenol-rich de-coumarinated extracts of Cinnamomum cassia. J. Funct. Foods 2014, 10, 54–64. [Google Scholar] [CrossRef]
- Wang, M.; Li, S.; Lange, K.W.; Zhao, H. Focusing on the Pharmacological Effects of Iridoids and Crocetin and Its Ester Derivatives of Gardenia jasminoides. Curr. Pharmacol. Rep. 2019, 5, 150–162. [Google Scholar] [CrossRef]
- Dias, M.; Caleja, C.; Pereira, C.; Calhelha, R.C.; Kostic, M.; Sokovic, M.; Tavares, D.; Baraldi, I.J.; Barros, L.; Ferreira, I.C.F.R. Chemical composition and bioactive properties of byproducts from two different kiwi varieties. Food Res. Int. 2020, 127, 108753. [Google Scholar] [CrossRef]
- Pires, T.C.S.P.; Dias, M.I.; Barros, L.; Calhelha, R.C.; Alves, M.J.; Santos-Buelga, C.; Ferreira, I.C.F.R. Phenolic compounds profile, nutritional compounds and bioactive properties of Lycium barbarum L.: A comparative study with stems and fruits. Ind. Crop. Prod. 2018, 122, 574–581. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.R.; Amorim, M.; Vilas-Boas, A.; Tonon, R.V.; Cabral, L.M.C.; Pastrana, L.; Pintado, M. Impact of in vitro gastrointestinal digestion on the chemical composition, bioactive properties, and cytotoxicity of Vitis vinifera L. cv. Syrah grape pomace extract. Food Funct. 2019, 10, 1856–1869. [Google Scholar] [CrossRef]
- Costa, R.S.d.; Santos, O.V.d.; Lannes, S.C.d.S.; Casazza, A.A.; Aliakbarian, B.; Perego, P.; Ribeiro-Costa, R.M.; Converti, A.; Silva Júnior, J.O.C. Bioactive compounds and value-added applications of cupuassu (Theobroma grandiflorum Schum.) agroindustrial by-product. Food Sci. Technol. 2020, 40, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Villacís-Chiriboga, J.; Elst, K.; Van Camp, J.; Vera, E.; Ruales, J. Valorization of byproducts from tropical fruits: Extraction methodologies, applications, environmental, and economic assessment: A review (Part 1: General overview of the byproducts, traditional biorefinery practices, and possible applications). Compr. Rev. Food Sci. Food Saf. 2020, 19, 405–447. [Google Scholar] [CrossRef]
- Lu, Q.-Y.; Arteaga, J.R.; Zhang, Q.; Huerta, S.; Go, V.L.W.; Heber, D. Inhibition of prostate cancer cell growth by an avocado extract: Role of lipid-soluble bioactive substances. J. Nutr. Biochem. 2005, 16, 23–30. [Google Scholar] [CrossRef]
- Ding, H.; Chin, Y.-W.; Kinghorn, A.D.; D’Ambrosio, S.M. Chemopreventive characteristics of avocado fruit. Semin. Cancer Biol. 2007, 17, 386–394. [Google Scholar] [CrossRef]
- Ding, H.; Han, C.; Guo, D.; Chin, Y.-W.; Ding, Y.; Kinghorn, A.D.; D’Ambrosio, S.M. Selective Induction of Apoptosis of Human Oral Cancer Cell Lines by Avocado Extracts Via a ROS-Mediated Mechanism. Nutr. Cancer 2009, 61, 348–356. [Google Scholar] [CrossRef]
- D’Ambrosio, S.M.; Han, C.; Pan, L.; Douglas Kinghorn, A.; Ding, H. Aliphatic acetogenin constituents of avocado fruits inhibit human oral cancer cell proliferation by targeting the EGFR/RAS/RAF/MEK/ERK1/2 pathway. Biochem. Biophys. Res. Commun. 2011, 409, 465–469. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, N.S.; Barakat, H.; El-Hallouty, S.; Salem, D. Effect of the Water Extracts of Avocado Fruit and Cherimoya Leaf on Four Human Cancer Cell Lines and Vicia Faba Root Tip Cells. J. Agric. Sci. 2013, 5, 245. [Google Scholar] [CrossRef]
- Larijani, L.V.; Ghasemi, M.; AbedianKenari, S.; Naghshvar, F. Evaluating the effect of four extracts of avocado fruit on esophageal squamous carcinoma and colon adenocarcinoma cell lines in comparison with peripheral blood mononuclear cells. Acta Med. Iran. 2014, 52, 201–205. [Google Scholar]
- Yu, G.; Li, N.; Zhao, Y.; Wang, W.; Feng, X.L. Salidroside induces apoptosis in human ovarian cancer SKOV3 and A2780 cells through the p53 signaling pathway. Oncol. Lett. 2018, 15, 6513–6518. [Google Scholar] [CrossRef]
- Horvathova, E.; Mastihubova, M.; Karnisova Potocka, E.; Kis, P.; Galova, E.; Sevcovicova, A.; Klapakova, M.; Hunakova, L.; Mastihuba, V. Comparative study of relationship between structure of phenylethanoid glycopyranosides and their activities using cell-free assays and human cells cultured in vitro. Toxicol. Vitro 2019, 61, 104646. [Google Scholar] [CrossRef]
- Navarro-Salcedo, M.H.; Delgado-Saucedo, J.I.; Siordia-Sánchez, V.H.; González-Ortiz, L.J.; Castillo-Herrera, G.A.; Puebla-Pérez, A.M. Artemisia dracunculus extracts obtained by organic solvents and supercritical CO2 produce cytotoxic and antitumor effects in mice with L5178Y lymphoma. J. Med. Food 2017, 20, 1076–1082. [Google Scholar] [CrossRef]
- Wilsher, N.E.; Arroo, R.R.; Matsoukas, M.T.; Tsatsakis, A.M.; Spandidos, D.A.; Androutsopoulos, V.P. Cytochrome P450 CYP1 metabolism of hydroxylated flavones and flavonols: Selective bioactivation of luteolin in breast cancer cells. Food Chem. Toxicol. 2017, 110, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Ganai, S.A.; Sheikh, F.A.; Baba, Z.A.; Mir, M.A.; Mantoo, M.A.; Yatoo, M.A. Anticancer activity of the plant flavonoid luteolin against preclinical models of various cancers and insights on different signalling mechanisms modulated. Phytother. Res. 2021. [Google Scholar] [CrossRef]
- Jang, C.H.; Moon, N.; Oh, J.; Kim, J.-S. Luteolin Shifts Oxaliplatin-Induced Cell Cycle Arrest at G0/G1 to Apoptosis in HCT116 Human Colorectal Carcinoma Cells. Nutrients 2019, 11, 770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, M.E.; Jansen, E.S.; Sanchez, W.; Waterhouse, N.J. Flow cytometry based assays for the measurement of apoptosis-associated mitochondrial membrane depolarisation and cytochrome c release. Methods 2013, 61, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, B.F.; Xie, F.J.; Yang, W.L.; Cao, N. Luteolin induces mitochondrial apoptosis in HT29 cells by inhibiting the Nrf2/ARE signaling pathway. Exp. Ther. Med. 2020, 19, 2179–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Hu, R.; Wang, J.; An, Y.; Lu, L.; Long, C.; Yan, L. Salidroside Suppresses IL-1β-Induced Apoptosis in Chondrocytes via Phosphatidylinositol 3-Kinases (PI3K)/Akt Signaling Inhibition. Med. Sci. Monit. 2019, 25, 5833–5840. [Google Scholar] [CrossRef]
- Huang, L.; Jin, K.; Lan, H. Luteolin inhibits cell cycle progression and induces apoptosis of breast cancer cells through downregulation of human telomerase reverse transcriptase. Oncol. Lett. 2019, 17, 3842–3850. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, M.; McGrath, K.L.; Di Trapani, G.; Charoentong, P.; Shah, F.; King, M.M.; Clarke, F.M.; Tonissen, K.F. The thioredoxin system in breast cancer cell invasion and migration. Redox Biol. 2016, 8, 68–78. [Google Scholar] [CrossRef]
- Quero, J.; Mármol, I.; Cerrada, E.; Rodríguez-Yoldi, M.J. Insight into the potential application of polyphenol-rich dietary intervention in degenerative disease management. Food Funct. 2020, 11, 2805–2825. [Google Scholar] [CrossRef]
- Wing-Cheung Leung, H.; Kuo, C.-L.; Yang, W.-H.; Lin, C.-H.; Lee, H.-Z. Antioxidant enzymes activity involvement in luteolin-induced human lung squamous carcinoma CH27 cell apoptosis. Eur. J. Pharmacol. 2006, 534, 12–18. [Google Scholar] [CrossRef]
- Fei-Yan, F.; Li-Xuan, S.; Jiang, M. Catechins and Their Therapeutic Benefits to Inflammatory Bowel Disease. Molecules 2017, 22, 484. [Google Scholar]
- Yoo, J.-H.; Park, E.-J.; Kim, S.H.; Lee, H.-J. Gastroprotective Effects of Fermented Lotus Root against Ethanol/HCl-Induced Gastric Mucosal Acute Toxicity in Rats. Nutrients 2020, 12, 808. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Avila, J.A.; Wall-Medrano, A.; Velderrain-Rodríguez, G.R.; Chen, C.Y.O.; Salazar-López, N.J.; Robles-Sánchez, M.; González-Aguilar, G.A. Gastrointestinal interactions, absorption, splanchnic metabolism and pharmacokinetics of orally ingested phenolic compounds. Food Funct. 2017, 8, 15–38. [Google Scholar] [CrossRef]
- Li, H.-Z.; Ren, Z.; Reddy, N.; Hou, T.; Zhang, Z.-J. In silico evaluation of antimicrobial, antihyaluronidase and bioavailability parameters of rosmarinic acid in Perilla frutescens leaf extracts. SN Appl. Sci. 2020, 2, 1–14. [Google Scholar] [CrossRef]
- Hakkou, Z.; Maciuk, A.; Leblais, V.; Bouanani, N.E.; Mekhfi, H.; Bnouham, M.; Aziz, M.; Ziyyat, A.; Rauf, A.; Hadda, T.B.; et al. Antihypertensive and vasodilator effects of methanolic extract of Inula viscosa: Biological evaluation and POM analysis of cynarin, chlorogenic acid as potential hypertensive. Biomed. Pharmacother. 2017, 93, 62–69. [Google Scholar] [CrossRef]
- Hardjono, S.; Siswandono, S.; Andayani, R. Evaluation of N-benzoylthiourea derivatives as possible analgesic agents by predicting their physicochemical and pharmacokinetic properties, toxicity, and analgesic activity. Indonesian J. Biotechnol. 2017, 22, 76–85. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PEEL | SEED COAT | SEED | |
|---|---|---|---|
| Total phenolics (mMol GA/100 g of extract) | 309.95 ± 25.33 a | 208.87 ± 11.67 b | 232.36 ± 12.25 b |
| Total Flavonoids (mMol Cat. eq./100 g of extract) | 12.54 ± 0.52 a | 3.41 ± 0.36 b | 2.13 ± 0.22 c |
| Total anthocyanins (mMol Cyanidin-3-glucoside eq./100 g of extract) | 622.37 ± 17.26 a | 575.76 ± 20.18 b | 359.85 ± 12.61 c |
| PEEL | SEED COAT | SEED | |
|---|---|---|---|
| DPPH (mMol Trolox eq./100 g of extract) | 46.49 ± 4.04 a | 36.80 ± 11.03 ab | 32.51 ± 9.07 b |
| (EC50 µg/mL) | 72.64 ± 10.70 a | 82.13 ± 2.54 ab | 90.91 ± 3.59 b |
| ABTS (mMol Trolox eq./100 g of extract) | 504.60 ± 35.50 a | 391.49.68 ± 48.26 b | 210.88 ± 64.11 c |
| (EC50 µg/mL) | 181.68 ± 18.47 a | 260.29 ± 16.41 b | 318.68 ± 25.60 c |
| FRAP (mMol Trolox eq./100 g of extract) | 379,308.80 ± 18,262.41 b | 672,334.16 ± 35,505.87 a | 256,338.52 ± 37,812.61 c |
| No. | Phenolic Compound | Peel | Seed Coat | Seed |
|---|---|---|---|---|
| 1 | p-hydroxybenzoic acid | 0.38 ± 0.01 a | 0.19 ± 0.03 b | 0.12 ± 0.00 c |
| 2 | Vanillin | 0.10 ± 0.02 a | 10 ± 0.10 b | 0.02 ± 0.00 b |
| 3 | Vanillic acid | 0.02 ± 0.00 | n.d | n.d. |
| 4 | Syringic acid | 0.09 ± 0.02 | n.d | n.d. |
| 5 | Protocatechuic acid | 2.20 ± 0.23 a | n.d | 0.28 ± 0.12 b |
| 6 | Protocatechuic acid glucoside | 26.10 ± 0.26 a | 7.50 ± 0.12 b | 1.95 ± 0.51 c |
| 7 | Hydroxytyrosol | 0.18 ± 0.08 a | 0.26 ± 0.00 a | 0.18 ± 0.05 a |
| 8 | Hydroxytyrosol glucoside | 0.13 ± 0.00 b | 1.24 ± 0.08 a | 0.91 ± 0.22 a |
| 9 | Hydroxysalidroside | 0.101 ± 0.02 b | 1.24 ± 0.20 b | 5.30 ± 1.08 a |
| 10 | Hydroxytyrosol glucoside arabinoside | 2.54 ± 0.10 | 0.02 ± 0.00 | n.d. |
| 11 | Tyrosol glucoside | 8.30 ± 0.52 b | 51.40 ± 0.63 a | 10.40 ± 2.70 b |
| 12 | Salidroside | 0.53 ± 0.10 b | 35.60 ± 3.84 b | 148.40 ± 50.30 a |
| 13 | Tyrosol glucoside arabinoside | 62.90 ± 1.09 a | 1.85 ± 0.10 b | 0.75 ± 0.19 b |
| 14 | p-coumaric acid | 0.24 ± 0.011 b | 0.33 ± 0.01 a | 0.20 ± 0.02 c |
| 15 | Coumaric acid glucoside | 0.32 ± 0.11 | n.d | n.d. |
| 16 | Coumaroylquinic acid | 1.38 ± 0.05 c | 7.33 ± 0.03 a | 3.61 ± 1.35 b |
| 17 | Caffeic acid | 0.41 ± 0.11 b | 2.98 ± 0.30 a | 0.59 ± 0.01 b |
| 18 | Caffeic acid glucoside | 0.37 ± 0.06 a | 0.22 ± 0.02 b | 0.09 ± 0.01 c |
| 19 | Caffeic acid glucoside derivative | 0.07 ± 0.01 | n.d | n.d. |
| 20 | Dihydrocaffeic acid glucoside | 0.07 ± 0.00 c | 0.58 ± 0.02 a | 0.21 ± 0.04 b |
| 21 | Caffeoylshikimic acid | 0.26 ± 0.01 b | 2.11 ± 0.25 a | 2210 ± 0.76 a |
| 22 | 3-O-caffeoylquinic acid | 17.60 ± 0.26 c | 738.70 ± 59.80 a | 176.20 ± 53.40 b |
| 23 | 4-O-caffeoylquinic acid | 9.56 ± 0.07 b | 71.40 ± 7.14 a | 10.70 ± 4.12 b |
| 24 | 5-O-caffeoylquinic acid | 969.20 ± 9.21 a | 65.60 ± 7.81 b | 11.30 ± 4.26 c |
| 25 | Dicaffeoylquinic acid | 1.05 ± 0.01 a | 0.02 ± 0.00 b | n.d. |
| 26 | Ferulic acid | 0.25 ± 0.01 a | 0.04 ± 0.01 b | 0.05 ± 0.01 b |
| 27 | Ferulic acid glucoside | 4.14 ± 0.11 a | 3.97 ± 0.06 a | 1.52 ± 0.56 b |
| 28 | Dihydroferulic acid glucoside | 0.23 ± 0.01 a | 0.05 ± 0.00 b | 0.01 ± 0.00 c |
| 29 | 4-O-feruoylquinic acid | 0.48 ± 0.05 b | 1.54 ± 0.23 a | 0.22 ± 0.05 b |
| 30 | 5-O-feruoylquinic acid | 1.61 ± 0.05 a | 0.59 ± 0.04 b | 0.124 ± 0.06 c |
| 31 | 3-O-feruoylquinic acid | 0.00 ± 0.00 c | 11.50 ± 0.66 a | 2.83 ± 1.57 b |
| Phenolic acids | 1215.17 ± 24.28 a | 1011.57 ± 81.44 a | 378.21 ± 121.60 b | |
| 32 | Catechin | n.d. | 311.20 ± 43.10 a | 280.50 ± 0.14 a |
| 33 | Epicatechin | 1891.00 ± 75.70 a | 571.00 ± 91.00 b | 360.00 ± 140.60 b |
| 34 | Catechin glucoside | 2.40 ± 0.03 b | 10.80 ± 1.05 a | 3.85 ± 0.08 b |
| 35 | Epicatechin glucoside | 7.89 ± 0.16 a | 8.61 ± 0.34 a | 4.81 ± 0.66 b |
| 36 | Epigallocatechin | 6.27 ± 0.28 a | 1.36 ± 0.042 b | 1.86 ± 0.67 b |
| 37 | Epicatechin gallate | n.d. | 3.06 ± 0.11 a | 1.39 ± 0.35 b |
| 38 | Catechin derivative | 1.55 ± 0.32 c | 5.50 ± 0.00 a | 2.32 ± 0.27 b |
| 39 | Epicatechin derivative | 67.30 ± 9.96 a | 3.05 ± 0.01 b | 1.72 ± 0.16 b |
| 40 | Procyanidin dimer (type A) | 4.45 ± 0.085 c | 41.00 ± 0.49 a | 6.28 ± 0.88 b |
| 41 | Procyanidin dimer (type B) | 2262.00 ± 63.00 a | 332.50 ± 24.10 b | 207.70 ± 112.10 b |
| 42 | Procyanidin trimer (type A) | 9.27 ± 1.21 c | 401.70 ± 2.00 a | 231.40 ± 74.20 b |
| 43 | Procyanidin trimer (type B) | 383.20 ± 14.90 a | 104.60 ± 10.40 b | 11.60 ± 1.65 c |
| 44 | Procyanidin tetramer | 106.20 ± 4.18 a | 16.00 ± 1.12 b | 9.73 ± 2.91 b |
| 45 | Procyanidin pentamer | 1.09 ± 0.06 a | 0.67 ± 0.36 a | 0.54 ± 0.12 a |
| 46 | Procyanidin hexamer | 6.80 ± 0.02 a | 3.01 ± 0.00 b | 1.31 ± 0.14 c |
| 47 | Quercetin | 0.69 ± 0.018 a | 0.05 ± 0.01 c | 0.39 ± 0.00 b |
| 48 | Quercetin arabinoside | 0.70 ± 0.00 a | 0.11 ± 0.01 c | 0.61± 0.02 b |
| 49 | Quercetin glucoside | 20.30 ± 0.75 a | 3.95 ± 0.25 b | 4.510 ± 0.11 b |
| 50 | Quercetin rhmanoside | 2.07 ± 0.85 | n.d | n.d. |
| 51 | Quercetin glucuronide | 67.80 ± 0.18 a | 0.08 ± 0.01 b | 0.04 ± 0.00 b |
| 52 | Quercetin acetylglucoside | 2.99 ± 0.09 a | 0.17 ± 0.01 b | 0.02 ± 0.01 c |
| 53 | Quercetin arabinoside glucoside | 374.40 ± 22.70 a | 0.68 ± 0.19 b | 0.39 ± 0.08 b |
| 54 | Quercetin rutinoside | 6.73 ± 0.19 a | 0.36 ± 0.03 b | 0.22 ± 0.10 b |
| 55 | Quercetin diglucoside | 294.00 ± 1.35 a | 4.51 ± 0.28 b | 1.82 ± 0.17 c |
| 56 | Quercetin glucoside rhamnoside | 4.98 ± 1.12 | n.d | n.d. |
| 57 | Isorhamnetin | 0.01 ± 0.00 | n.d | n.d. |
| 58 | Isorhamnetin derivative | 2.19 ± 0.08 | n.d | n.d. |
| 59 | Isorhamnetin arabinoside | 0.04 ± 0.00 c | 0.98 ± 0.054 a | 0.48 ± 0.12 b |
| 60 | Isorhamnetin glucoside | 0.06 ± 0.00 b | 0.15 ± 0.01 a | 0.04 ± 0.01 b |
| 61 | Isorhamnetin glucuronide | 9.20 ± 0.06 a | 0.01 ± 0.00 b | n.d. |
| 62 | Isorhamnetin arabinoside glucoside | 0.10 ± 0.00 | n.d | n.d. |
| 63 | Kaempferol arabinoside | 0.26 ± 0.00 a | n.d | 0.09 ± 0.03 b |
| 64 | Kaempferol glucoside | 4.81 ± 0.00 a | 0.34 ± 0.04 c | 0.64 ± 0.10 b |
| 65 | Kaempferol rutinoside | 13.20 ± 0.06 | n.d | n.d. |
| 66 | Kaempferol arabinoside glucoside | 160.70 ± 0.00 a | 0.28 ± 0.01 b | 0.245 ± 0.02 b |
| 67 | Naringenin | 0.27 ± 0.08 a | 0.08 ± 0.00 b | 0.19 ± 0.01 a b |
| 68 | Naringenin glucoside | 0.59 ± 0.00 b | 1.11 ± 0.08 a | 0.00 ± 0.00 c |
| 69 | Sakuranetin | 0.24 ± 0.00 b | n.d | 0.86 ± 0.13 a |
| 70 | Luteolin | n.d. | 0.02 ± 0.00 b | 0.05 ± 0.01 a |
| 71 | Luteolin arabinoside glucoside | 6.07 ± 0.33 | n.d. | n.d. |
| Flavonoids | 5721.86 ± 208.82 a | 1826.99 ± 193.20 b | 1135.68 ± 507.45 b | |
| 72 | Penstemide | 2.82 ± 0.00 a | 1.91 ± 0.06 b | 0.62 ± 0.12 c |
| Terpenes | 2.820 ± 0.00 a | 1.91 ± 0.06 b | 0.62 ± 0.12 c | |
| Total phenolic compounds | 6836.35 ± 62.80 a | 3850137 ± 356.09 b | 1513.90 ± 578.20 c |
| Peel | Seed Coat | Seed | |
|---|---|---|---|
| Carbohydrates (%) | 36.39 ± 7.11 a | 23.92 ± 1.23 b | 28.13 ± 0.24 ab |
| Proteins (%) | 2.82 ± 0.30 a | 0.79 ± 0.01 b | 0.69 ± 0.00 b |
| Fat (%) | 7.68 ± 1.19 c | 13.41 ± 0.82 b | 21.55 ± 1.07 a |
| Moisture (%) | 14.45 ± 0.26 a | 15.14 ± 0.26 a | 11.09 ± 0.33 b |
| Ash (%) | 7.34 ± 0.34 a | 6.46 ± 0.12 b | 3.86 ± 0.03 c |
| Energy (kcal/100 g) | 226.06 ± 0.06 b | 219.62 ± 2.49 b | 309.30 ± 8.65 a |
| Identified Compound | MW | TPSA | Log P | No. Atoms | Hydrogen Bonds Acceptors | Hydrogen Bonds Donors | Rotatable Bonds | Molecular Volume (Å3) | Violations to LIRF | % ABS | log Papp | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | p-Hydroxybenzoic acid | 138.12 | 57.53 | 1.37 | 10 | 3 | 2 | 1 | 119.06 | 0 | 89.15 | 1.15 |
| 2 | Vanillin | 152.15 | 46.53 | 1.07 | 11 | 3 | 1 | 2 | 136.59 | 0 | 92.95 | 1.21 |
| 3 | Vanillic acid | 168.15 | 66.76 | 1.19 | 12 | 4 | 2 | 2 | 144.61 | 0 | 85.97 | 0.33 |
| 4 | Syringic acid | 198.17 | 76 | 1.20 | 14 | 5 | 2 | 3 | 170.15 | 0 | 82.78 | 0.49 |
| 5 | Protocatechuic acid | 154.12 | 77.75 | 0.88 | 11 | 4 | 3 | 1 | 127.08 | 0 | 82.18 | 0.49 |
| 6 | Protocatechuic acid glucoside | 302.24 | 156.91 | −1.37 | 21 | 9 | 6 | 3 | 242.40 | 1 | 54.87 | −0.66 |
| 7 | Hydroxytyrosol | 154.16 | 60.68 | 0.52 | 11 | 3 | 3 | 2 | 141.70 | 0 | 88.07 | 1.09 |
| 8 | Hydroxytyrosol glucoside | 316.31 | 139.84 | −1.19 | 22 | 8 | 6 | 5 | 273.82 | 1 | 60.76 | 0.14 |
| 9 | Hydroxysalidroside | 316.31 | 139.84 | −1.19 | 22 | 8 | 6 | 5 | 273.82 | 1 | 60.76 | 0.16 |
| 10 | Hydroxytyrosol glucoside arabinoside | 448.42 | 198.76 | −2.78 | 31 | 12 | 8 | 7 | 381.10 | 2 | 40.43 | −0.63 |
| 11 | Tyrosol glucoside | 300.31 | 119.61 | −0.70 | 21 | 7 | 5 | 5 | 265.80 | 0 | 67.73 | 0.46 |
| 12 | Salidroside | 300.31 | 119.61 | −0.70 | 21 | 7 | 5 | 5 | 265.80 | 0 | 67.73 | 0.46 |
| 13 | Tyrosol glucoside arabinoside | 432.42 | 178.53 | −2.06 | 30 | 11 | 7 | 7 | 373.08 | 2 | 47.41 | −0.44 |
| 14 | p-coumaric acid | 164.16 | 57.53 | 1.43 | 12 | 3 | 2 | 2 | 146.48 | 0 | 89.15 | 1.21 |
| 15 | Coumaric acid glucoside | 326.30 | 136.68 | −0.36 | 23 | 8 | 5 | 5 | 278.60 | 0 | 61.85 | −0.58 |
| 16 | Coumaroyl quinic acid | 338.31 | 144.52 | 0.28 | 24 | 8 | 5 | 5 | 288.60 | 0 | 59.14 | 0.80 |
| 17 | Caffeic acid | 180.16 | 77.75 | 0.94 | 13 | 4 | 3 | 2 | 154.50 | 0 | 82.18 | 0.63 |
| 18 | Caffeic Acid Glucoside | 342.30 | 156.91 | −1.07 | 24 | 9 | 6 | 5 | 286.62 | 1 | 54.87 | −0.67 |
| 19 | Caffeic Acid Glucoside derivative | 342.30 | 156.91 | −1.07 | 24 | 9 | 6 | 5 | 286.62 | 1 | 54.87 | −0.67 |
| 20 | Dihydrocaffeic acid glucoside | 330.29 | 156.91 | −1.33 | 23 | 9 | 6 | 5 | 276.00 | 1 | 54.87 | −0.08 |
| 21 | Caffeoylshikimic acid | 336.30 | 144.52 | 0.31 | 24 | 8 | 5 | 5 | 282.36 | 0 | 59.14 | −0.59 |
| 22 | 3-O-caffeoylquinic acid | 354.31 | 164.74 | −0.45 | 25 | 9 | 6 | 5 | 296.27 | 1 | 52.16 | −0.84 |
| 23 | 4-O-caffeoylquinic acid | 354.31 | 164.74 | −0.67 | 25 | 9 | 6 | 5 | 296.27 | 1 | 52.16 | −0.89 |
| 24 | 5-O-caffeoylquinic acid | 354.31 | 164.74 | −0.45 | 25 | 9 | 6 | 5 | 296.27 | 1 | 52.16 | −0.84 |
| 25 | Dicaffeoylquinic acid | 516.46 | 211.28 | 1.21 | 37 | 12 | 7 | 9 | 431.08 | 3 | 36.11 | −1.20 |
| 26 | Ferulic acid | 194.19 | 66.76 | 1.25 | 14 | 4 | 2 | 3 | 172.03 | 0 | 85.97 | 0.17 |
| 27 | Ferulic Acid Glucoside | 356.33 | 145.91 | −0.77 | 25 | 9 | 5 | 6 | 304.15 | 0 | 58.66 | −0.55 |
| 28 | Dihydroferulic acid glucoside | 356.33 | 145.91 | −0.77 | 25 | 9 | 5 | 6 | 304.15 | 0 | 58.66 | −0.55 |
| 29 | 4-O-feruloylquinic acid | 368.34 | 153.75 | −0.36 | 26 | 9 | 5 | 6 | 313.80 | 0 | 55.96 | −0.56 |
| 30 | 5-O-feruloylquinic acid | 368.34 | 153.75 | −0.14 | 26 | 9 | 5 | 6 | 313.80 | 0 | 55.96 | −0.56 |
| 31 | 3-O-feruloylquinic acid | 368.34 | 153.75 | −0.14 | 26 | 9 | 5 | 6 | 313.80 | 0 | 55.96 | −0.56 |
| 32 | Catechin | 290.27 | 110.37 | 1.37 | 21 | 6 | 5 | 1 | 244.14 | 0 | 70.92 | −0.28 |
| 33 | Epicatechin | 290.27 | 110.37 | 1.37 | 21 | 6 | 5 | 1 | 244.14 | 0 | 70.92 | −0.28 |
| 34 | Catechin glucoside | 452.41 | 189.53 | −0.89 | 32 | 11 | 8 | 4 | 376.26 | 2 | 43.61 | −0.88 |
| 35 | epicatechin glucoside | 452.41 | 189.53 | −0.34 | 32 | 11 | 8 | 4 | 376.26 | 2 | 43.61 | −0.92 |
| 36 | Epigallocatechin | 306.27 | 130.60 | 1.08 | 22 | 7 | 6 | 1 | 252.16 | 1 | 63.94 | −0.37 |
| 37 | Epicatechin gallate | 442.38 | 177.13 | 2.54 | 32 | 10 | 7 | 4 | 359.55 | 1 | 47.89 | −1.26 |
| 38 | Catechin derivative | 290.27 | 110.37 | 1.37 | 21 | 6 | 5 | 1 | 244.14 | 0 | 70.92 | −0.28 |
| 39 | Epicatechin derivative | 290.27 | 110.37 | 1.37 | 21 | 6 | 5 | 1 | 244.14 | 0 | 70.92 | −0.28 |
| 40 | Procyanidin dimer (type A) | 576.51 | 209.75 | 2.57 | 42 | 12 | 9 | 2 | 465.47 | 3 | 36.64 | −1.08 |
| 41 | Procyanidin dimer (type B) | 578.53 | 220.75 | 2.14 | 42 | 12 | 10 | 3 | 475.67 | 3 | 32.84 | −1.22 |
| 42 | Procyanidin trimer (type A) | 864.76 | 320.13 | 3.78 | 63 | 18 | 14 | 4 | 697.01 | 4 | -1.44 | −0.43 |
| 43 | Procyanidin trimer (type B) | 862.75 | 309.13 | 3.56 | 63 | 18 | 13 | 3 | 686.81 | 3 | 2.35 | −1.82 |
| 44 | Procyanidin tetramer | 1157.05 | 441.50 | 4.13 | 84 | 24 | 20 | 7 | 944.98 | 3 | -43.32 | −2.62 |
| 45 | Procyanidin pentamer | 1443.29 | 551.87 | 6.22 | 105 | 30 | 25 | 9 | 1170.27 | 4 | -81.40 | −0.43 |
| 46 | Procyanidin hexamer | 1701.56 | 632.78 | 8.54 | 124 | 34 | 29 | 14 | 1395.59 | 4 | -109.31 | −0.36 |
| 47 | Quercetin | 302.24 | 131.35 | 1.68 | 22 | 11 | 7 | 1 | 240.08 | 0 | 63.68 | −0.22 |
| 48 | Quercetin arabinoside | 434.35 | 190.28 | 0.06 | 31 | 11 | 7 | 3 | 347.36 | 2 | 43.35 | 0.15 |
| 49 | Quercetin glucoside | 464.38 | 210.50 | −0.36 | 33 | 12 | 8 | 4 | 372.21 | 2 | 36.38 | 0.27 |
| 50 | Quercetin rhamnoside | 448.38 | 190.28 | 0.64 | 32 | 11 | 7 | 3 | 363.95 | 2 | 43.35 | 0.048 |
| 51 | Quercetin glucuronide | 478.36 | 227.57 | −0.49 | 34 | 13 | 8 | 4 | 374.39 | 2 | 30.49 | −1.06 |
| 52 | Quercetin acetylglucoside | 506.42 | 216.58 | 0.34 | 36 | 13 | 7 | 6 | 408.72 | 3 | 34.28 | −0.00 |
| 53 | Quercetin arabinoside glucoside | 596.49 | 269.43 | −1.73 | 42 | 16 | 10 | 6 | 479.48 | 3 | 16.05 | −0.91 |
| 54 | Quercetin rutinoside | 610.52 | 269.43 | −1.06 | 43 | 16 | 10 | 6 | 496.07 | 3 | 16.05 | −1.59 |
| 55 | Quercetin diglucoside | 626.52 | 289.65 | −2.38 | 44 | 17 | 11 | 7 | 504.33 | 3 | 9.07 | −1.22 |
| 56 | Quercetin glucoside rhamnoside | 612.54 | 265.52 | −2.00 | 43 | 16 | 10 | 6 | 502.31 | 3 | 17.40 | −1.46 |
| 57 | Isorhamnetin | 316.26 | 120.36 | 1.99 | 23 | 7 | 4 | 0 | 257.61 | 0 | 67.48 | −0.00 |
| 58 | Isorhamnetin derivate | 316.26 | 120.36 | 1.99 | 23 | 7 | 4 | 0 | 257.61 | 0 | 67.48 | −0.00 |
| 59 | Isorhamnetin arabinoside | 448.38 | 179.28 | 0.37 | 32 | 11 | 6 | 2 | 364.89 | 2 | 47.15 | 0.37 |
| 60 | Isorhamnetin glucoside | 478.41 | 199.51 | −0.06 | 34 | 12 | 7 | 5 | 389.73 | 2 | 40.17 | 0.33 |
| 61 | Isorhamnetin glucoronide | 492.39 | 216.58 | −0.18 | 35 | 13 | 7 | 5 | 391.92 | 2 | 34.28 | −0.85 |
| 62 | Isorhamnetin arabinoside glucoside | 610.52 | 258.43 | −1.42 | 43 | 16 | 9 | 7 | 497.01 | 3 | 19.84 | −0.99 |
| 63 | Kaempferol arabinoside glycoside | 580.50 | 249.20 | −1.24 | 41 | 15 | 9 | 6 | 471.46 | 3 | 23.03 | −0.40 |
| 64 | Kaempferol glucoside | 448.38 | 190.28 | 0.12 | 32 | 11 | 7 | 4 | 364.19 | 2 | 43.35 | 0.35 |
| 65 | Kaempferol rutinoside | 594.54 | 249.20 | −0.57 | 42 | 15 | 9 | 6 | 488.05 | 3 | 23.03 | 0.18 |
| 66 | Naringenin | 272.26 | 86.99 | 2.12 | 20 | 5 | 3 | 1 | 230.26 | 0 | 78.99 | 1.02 |
| 67 | Naringenin glucoside | 434.40 | 177.13 | −0.04 | 31 | 10 | 7 | 3 | 361.42 | 1 | 47.89 | 0.41 |
| 68 | Sakuranetin | 286.28 | 76.00 | 2.65 | 21 | 5 | 2 | 2 | 247.79 | 0 | 82.78 | 1.36 |
| 69 | Luteolin | 286.24 | 111.12 | 1.97 | 21 | 6 | 4 | 1 | 232.07 | 0 | 70.66 | 0.96 |
| 70 | Luteolin arabinoside glucoside | 610.52 | 291.42 | −2.59 | 43 | 16 | 12 | 5 | 494.38 | 3 | 8,12 | −1.40 |
| 71 | Penstemide | 296.36 | 76 | 1.76 | 21 | 5 | 2 | 6 | 282.65 | 0 | 82.78 | 1.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velderrain-Rodríguez, G.R.; Quero, J.; Osada, J.; Martín-Belloso, O.; Rodríguez-Yoldi, M.J. Phenolic-Rich Extracts from Avocado Fruit Residues as Functional Food Ingredients with Antioxidant and Antiproliferative Properties. Biomolecules 2021, 11, 977. https://doi.org/10.3390/biom11070977
Velderrain-Rodríguez GR, Quero J, Osada J, Martín-Belloso O, Rodríguez-Yoldi MJ. Phenolic-Rich Extracts from Avocado Fruit Residues as Functional Food Ingredients with Antioxidant and Antiproliferative Properties. Biomolecules. 2021; 11(7):977. https://doi.org/10.3390/biom11070977
Chicago/Turabian StyleVelderrain-Rodríguez, Gustavo R., Javier Quero, Jesús Osada, Olga Martín-Belloso, and María Jesús Rodríguez-Yoldi. 2021. "Phenolic-Rich Extracts from Avocado Fruit Residues as Functional Food Ingredients with Antioxidant and Antiproliferative Properties" Biomolecules 11, no. 7: 977. https://doi.org/10.3390/biom11070977
APA StyleVelderrain-Rodríguez, G. R., Quero, J., Osada, J., Martín-Belloso, O., & Rodríguez-Yoldi, M. J. (2021). Phenolic-Rich Extracts from Avocado Fruit Residues as Functional Food Ingredients with Antioxidant and Antiproliferative Properties. Biomolecules, 11(7), 977. https://doi.org/10.3390/biom11070977