
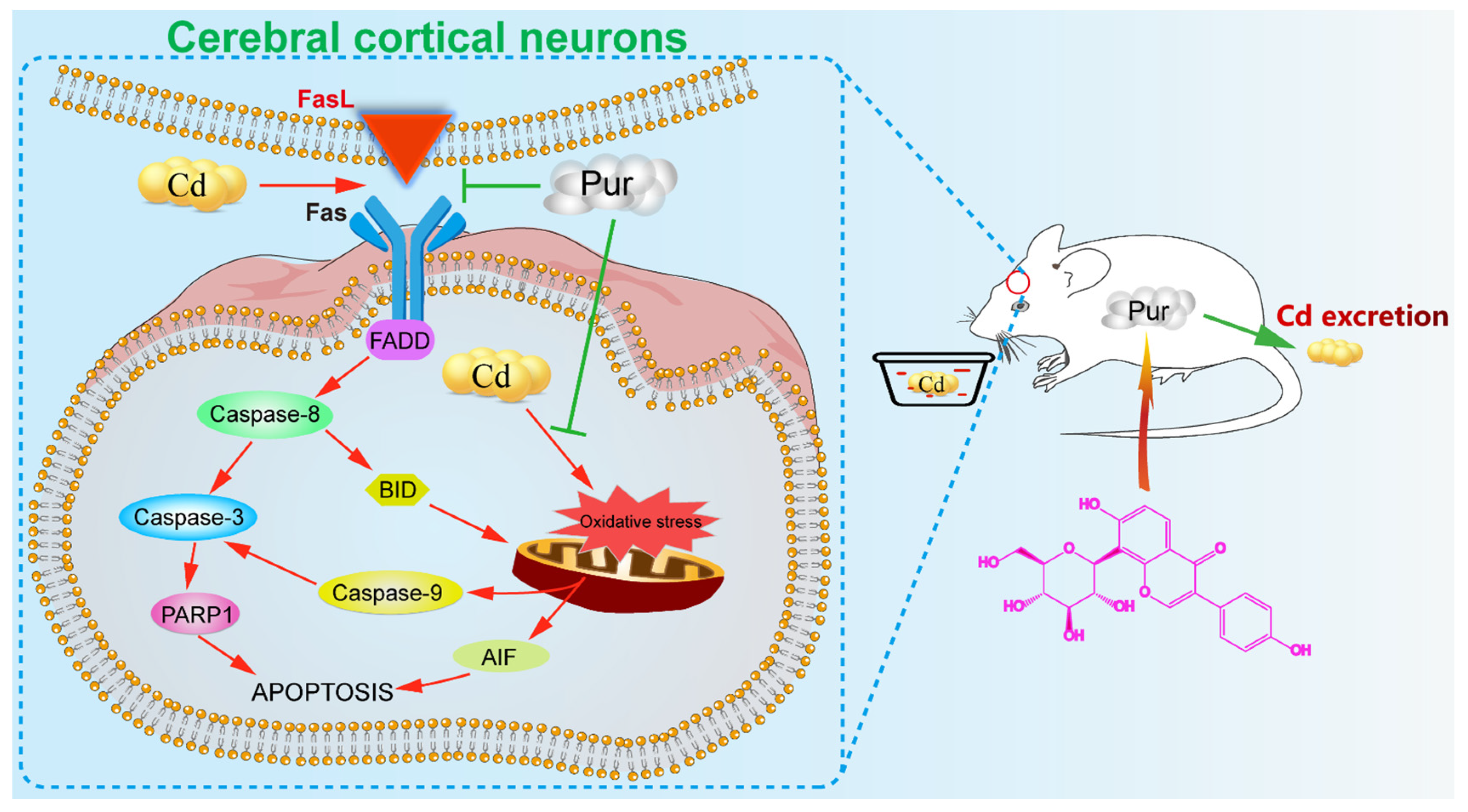
Puerarin Attenuates Cadmium-Induced Neuronal Injury via Stimulating Cadmium Excretion, Inhibiting Oxidative Stress and Apoptosis



 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals and Reagents
2.2. Animals and Treatments
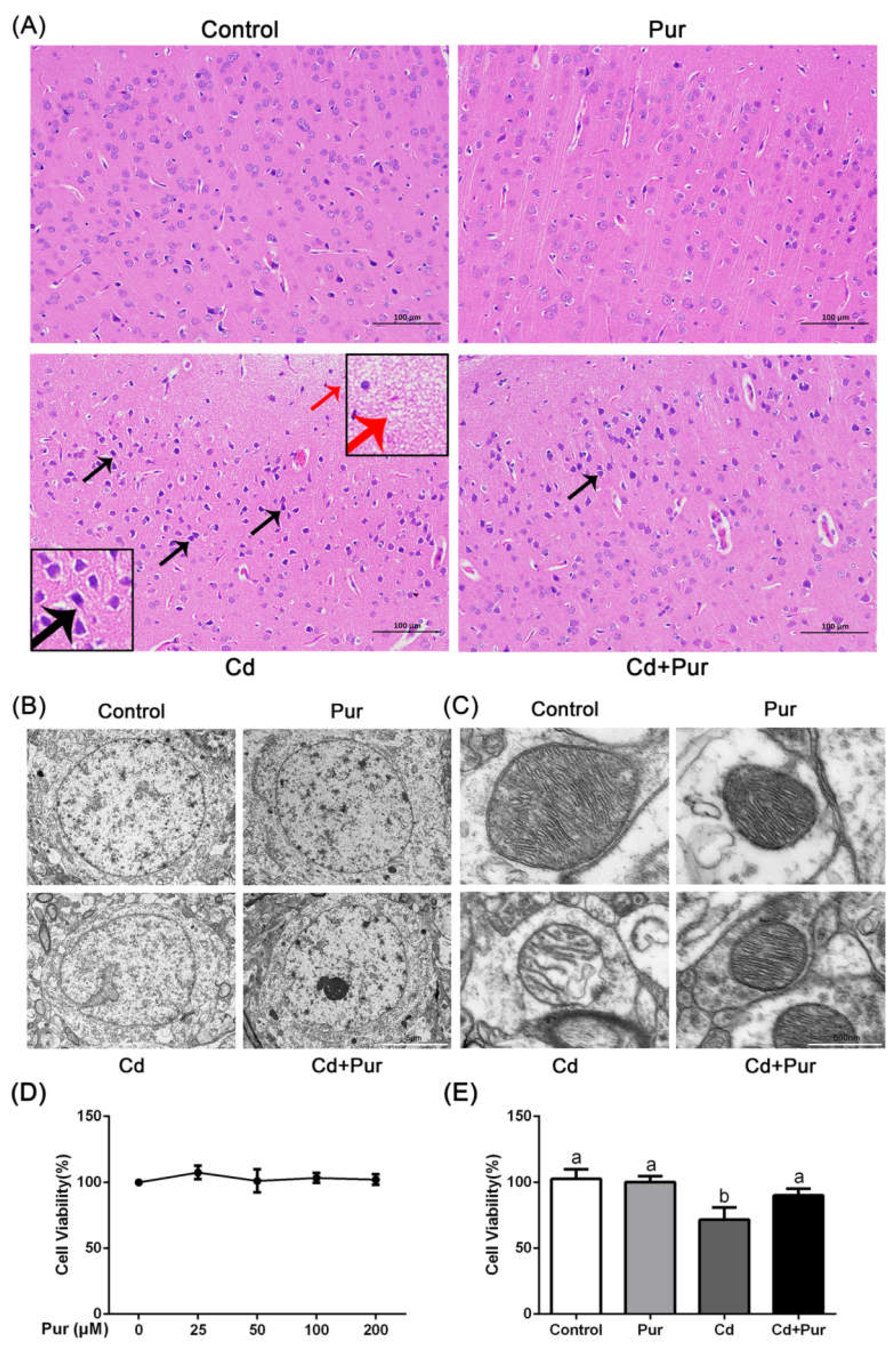
2.3. Hematoxylin and Eosin (H&E) Staining and Histological Analysis
2.4. Transmission Electron Microscopy
2.5. Cd Measurement
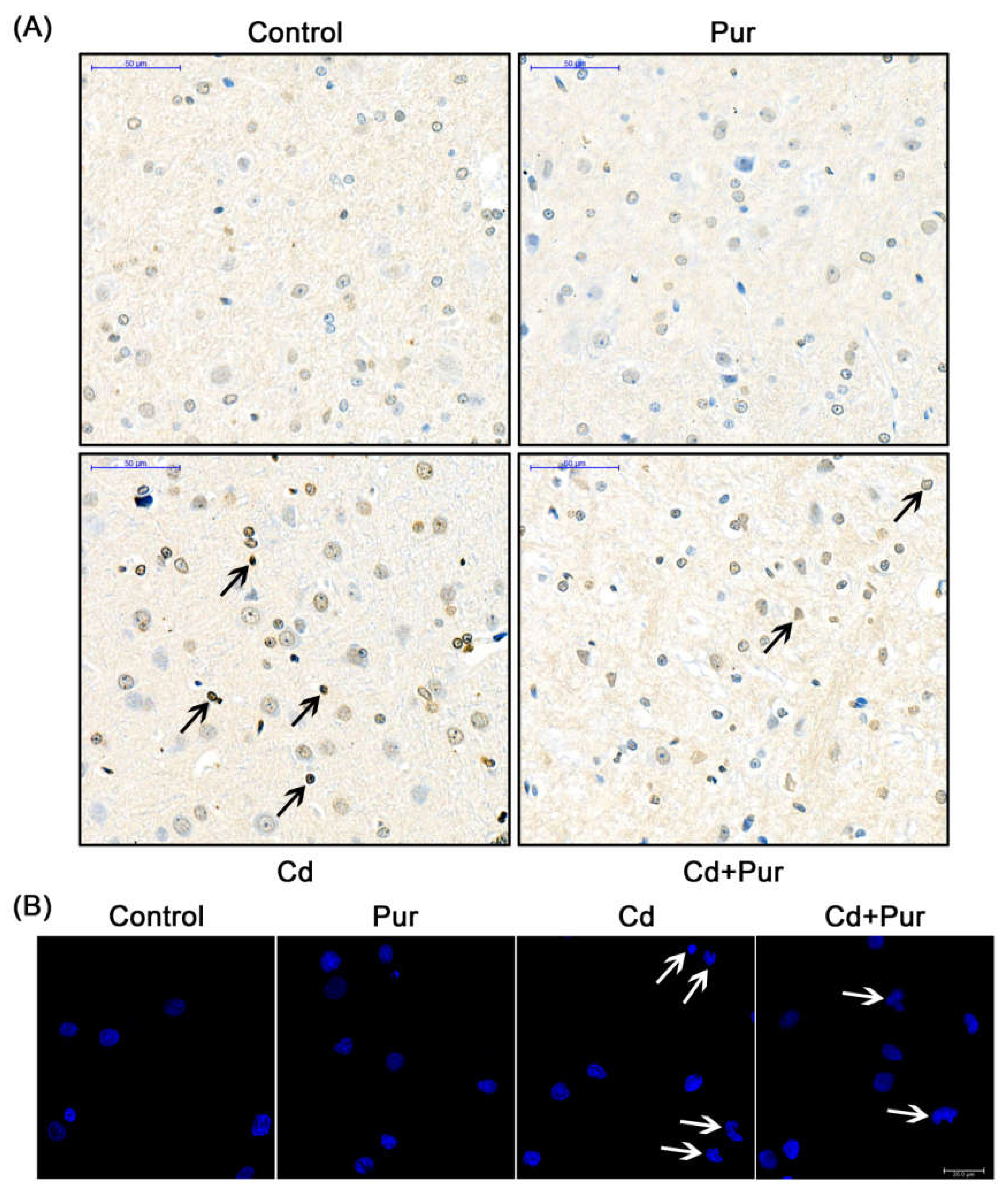
2.6. TUNEL Staining
2.7. Cell Isolation and Culture
2.8. Cell Viability Assay
2.9. DAPI Staining
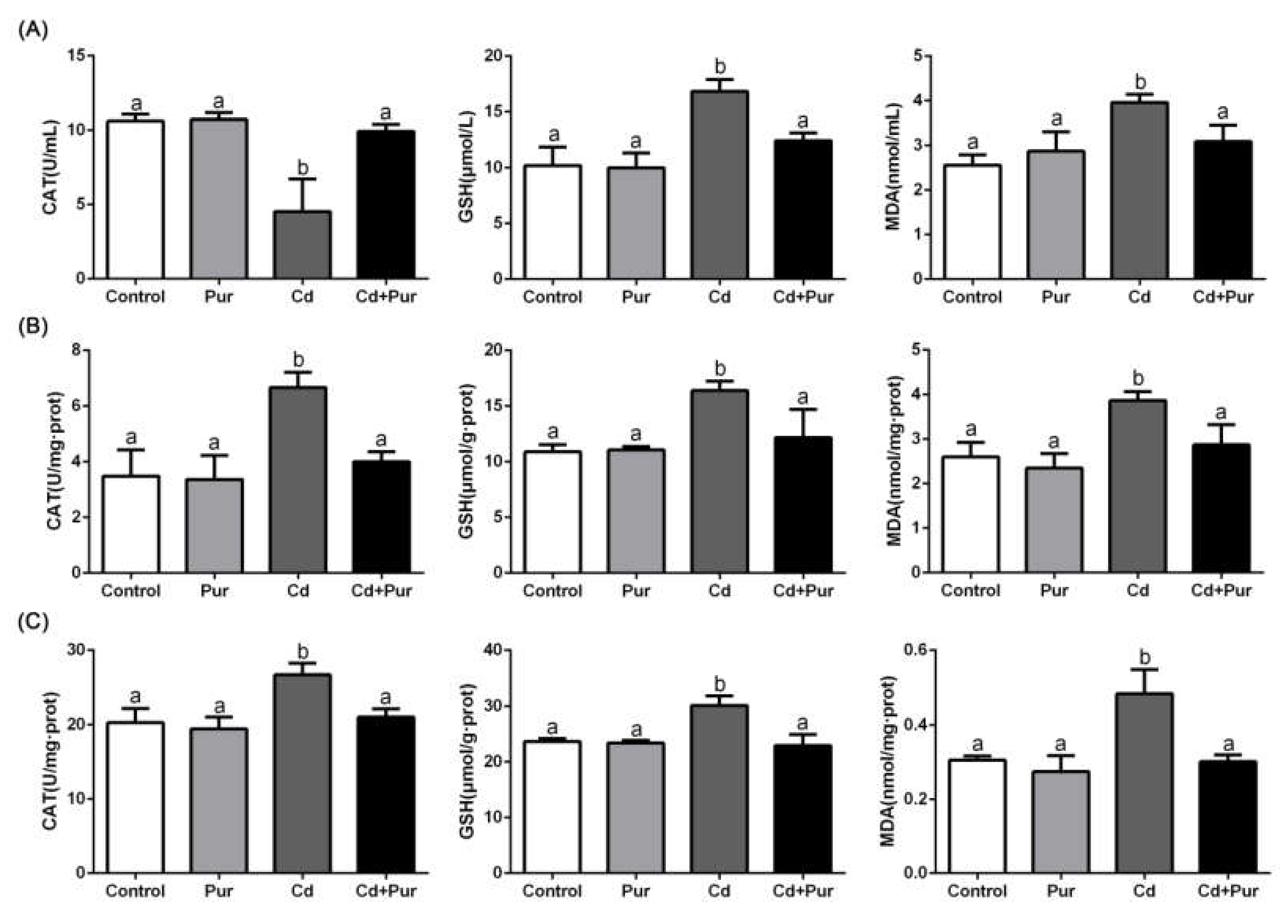
2.10. Oxidative Stress Assessment
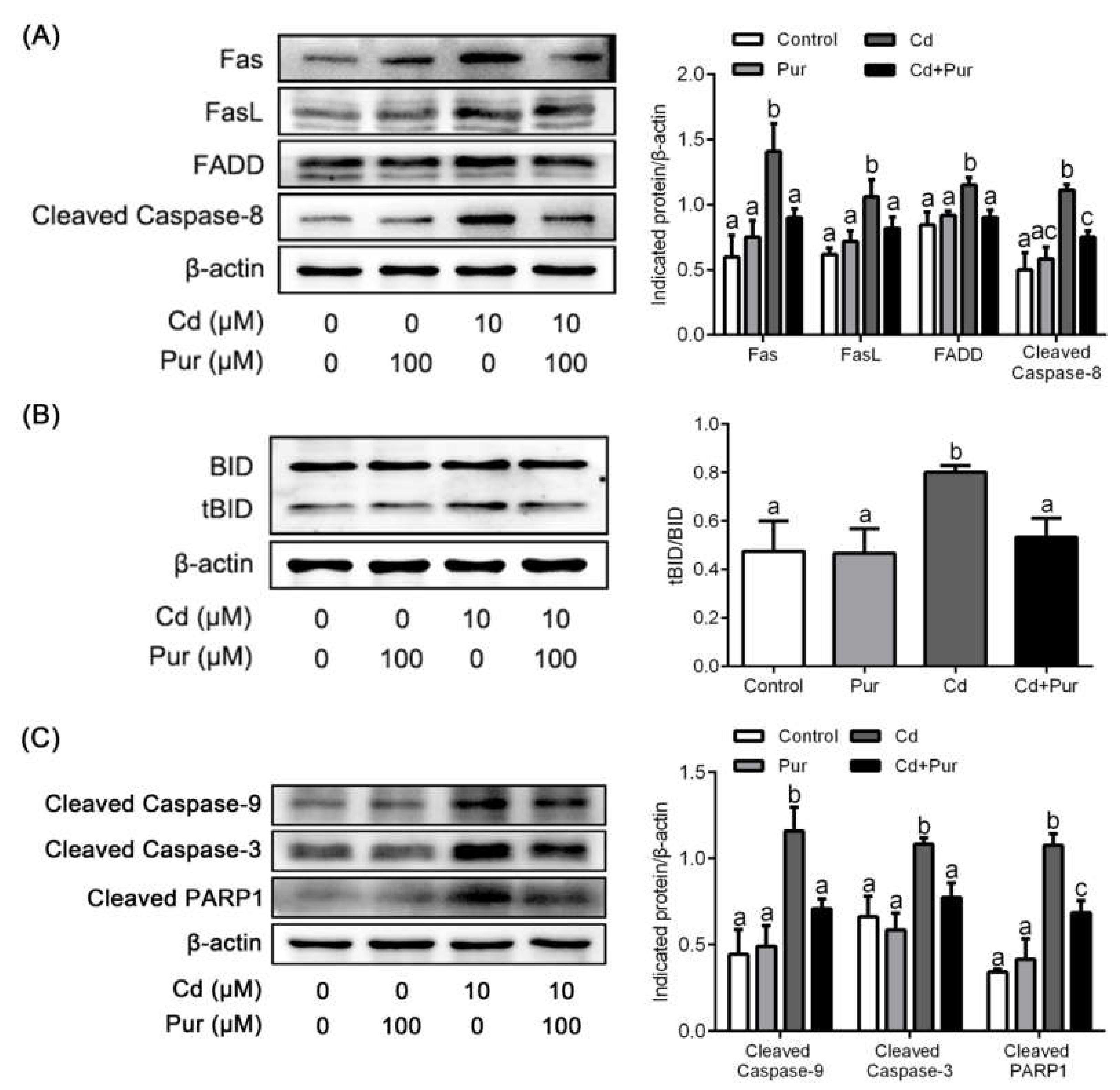
2.11. Western Blotting
2.12. Immunofluorescence Assays
2.13. Statistical Analyses
3. Results
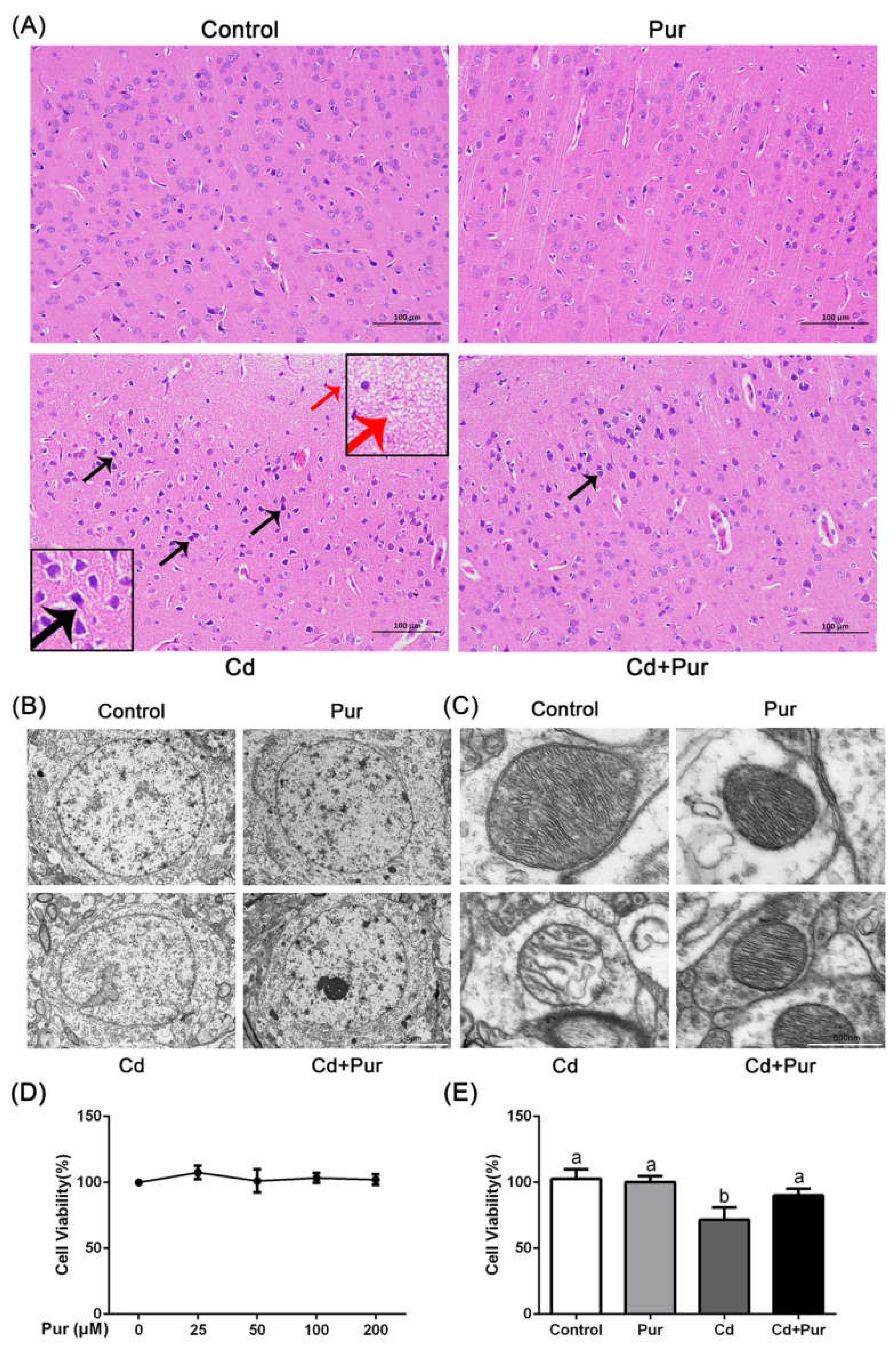
3.1. Pur Alleviates Cd-Induced Neuronal Injury
3.2. Pur Reduces Cd Levels in the Rat Cerebral Cortex and Stimulates Cd Excretion
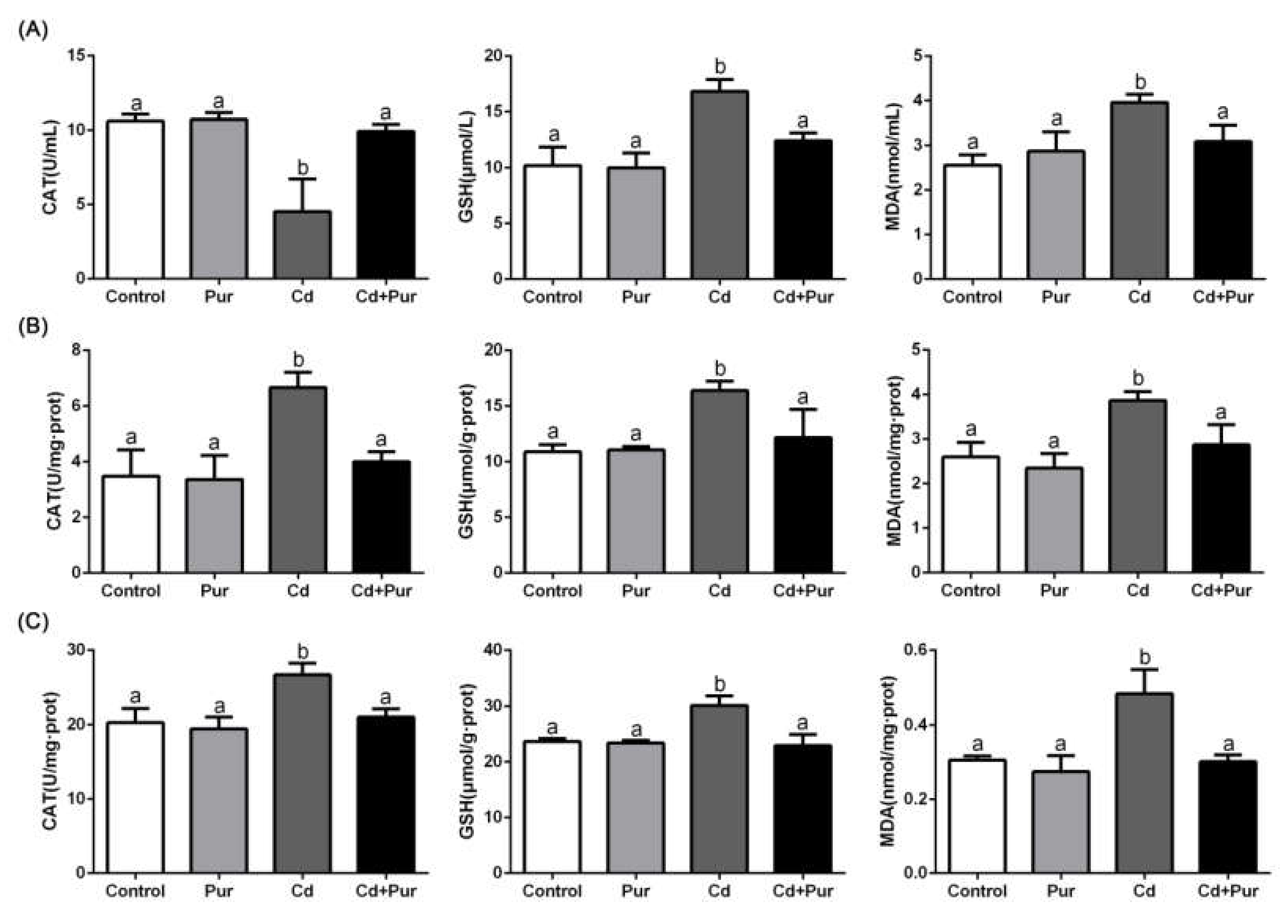
3.3. Pur Alleviates Cd-Induced Oxidative Stress in the Rat Serum and Cerebral Cortical Neurons
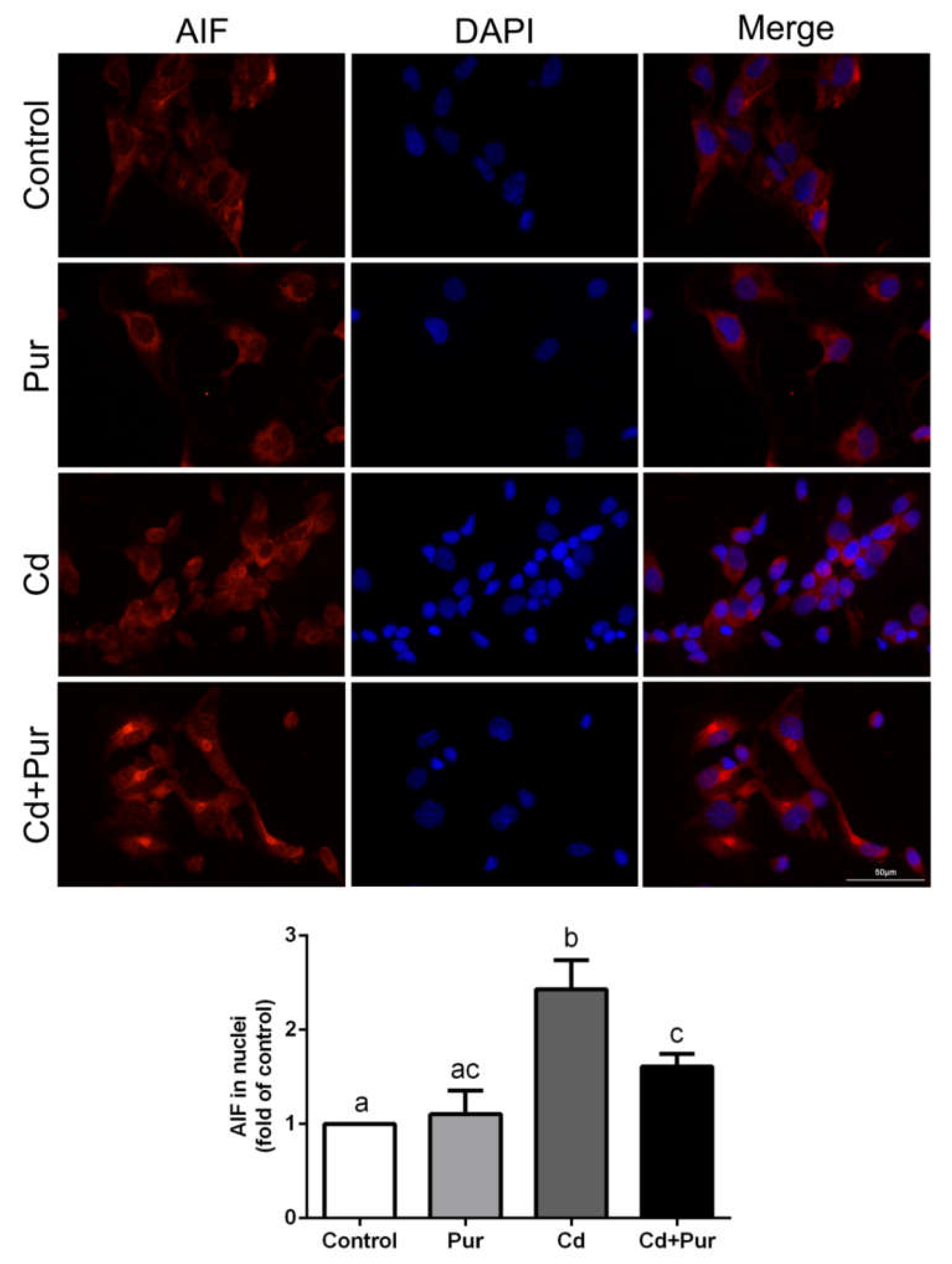
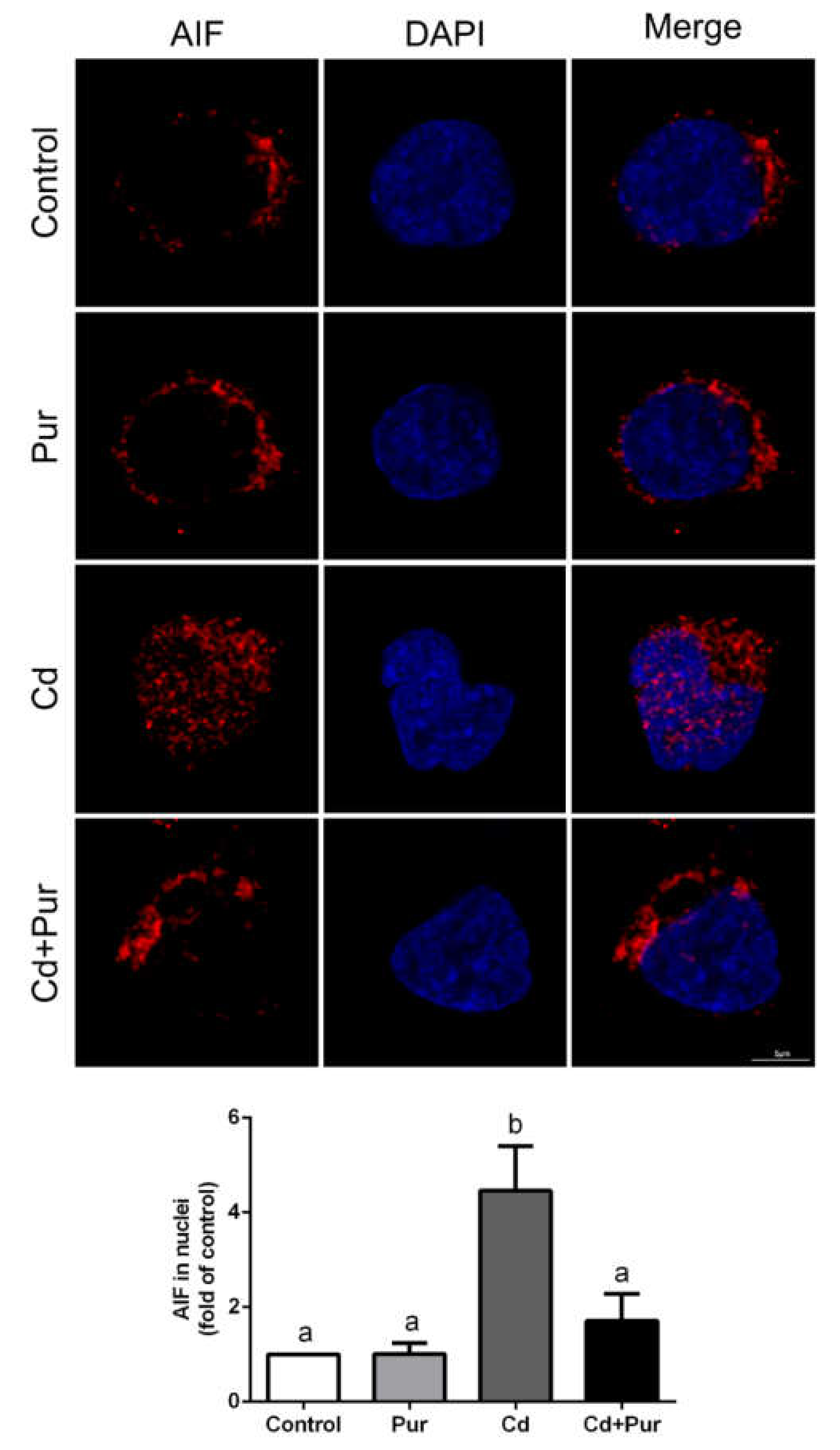
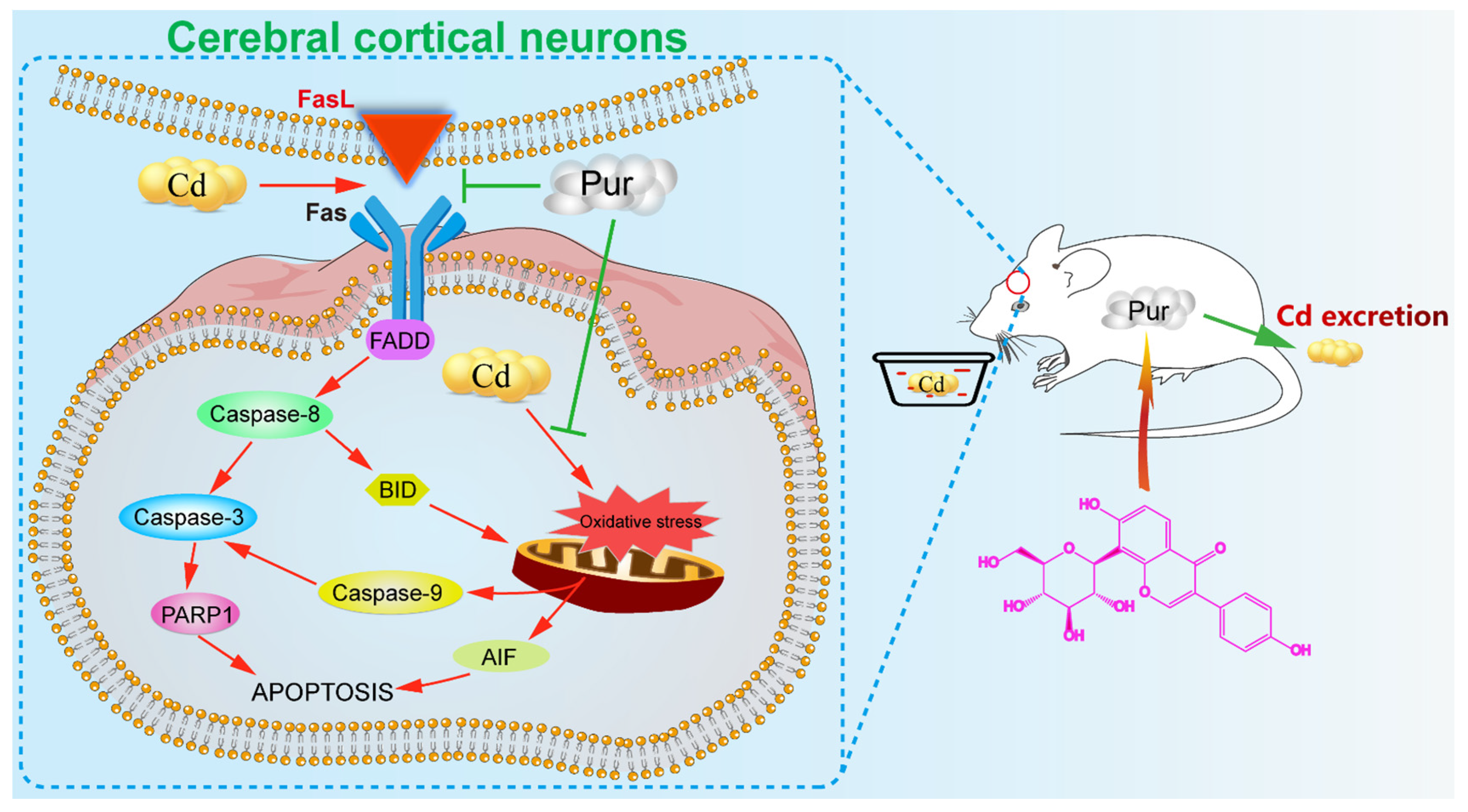
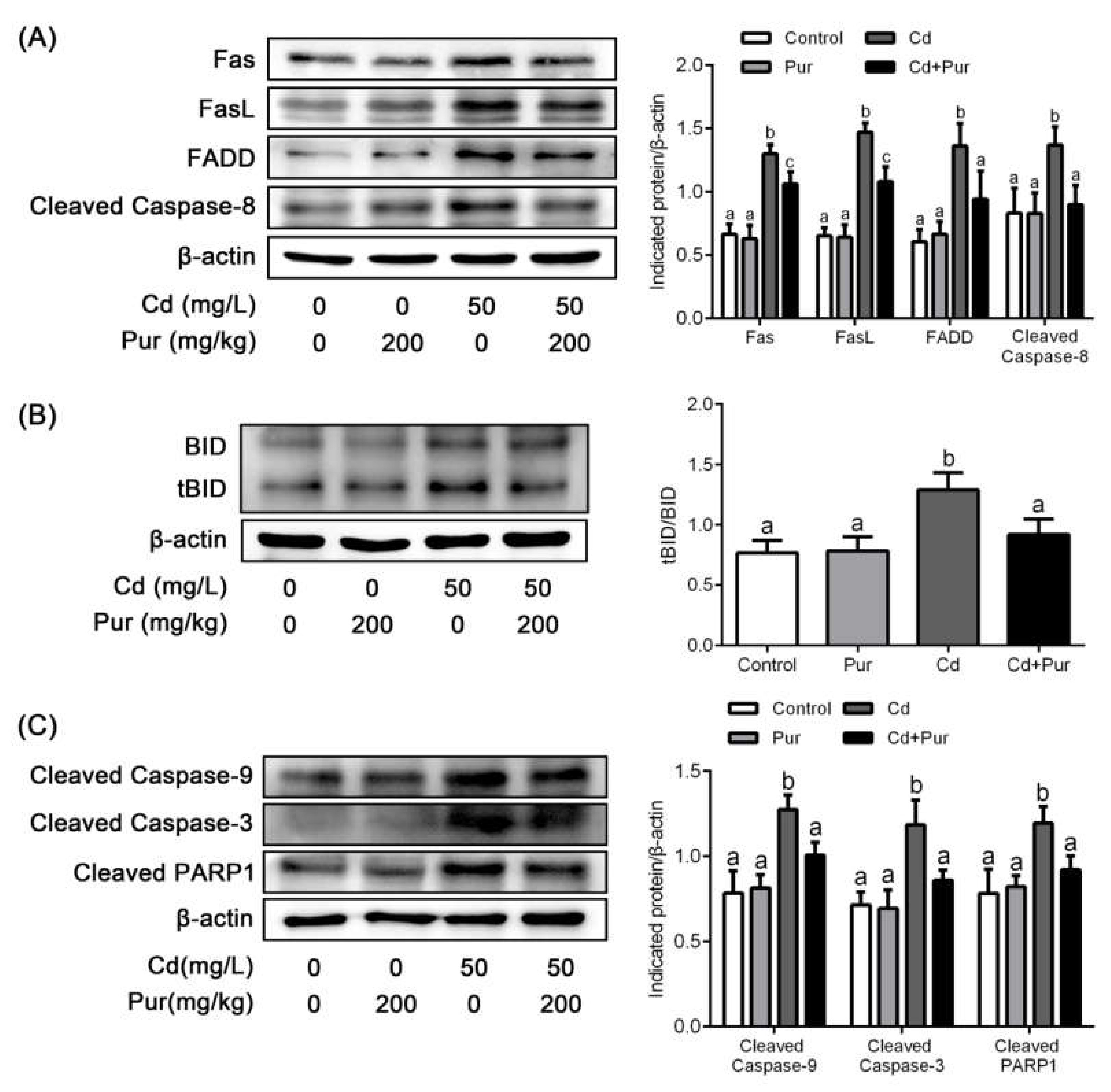
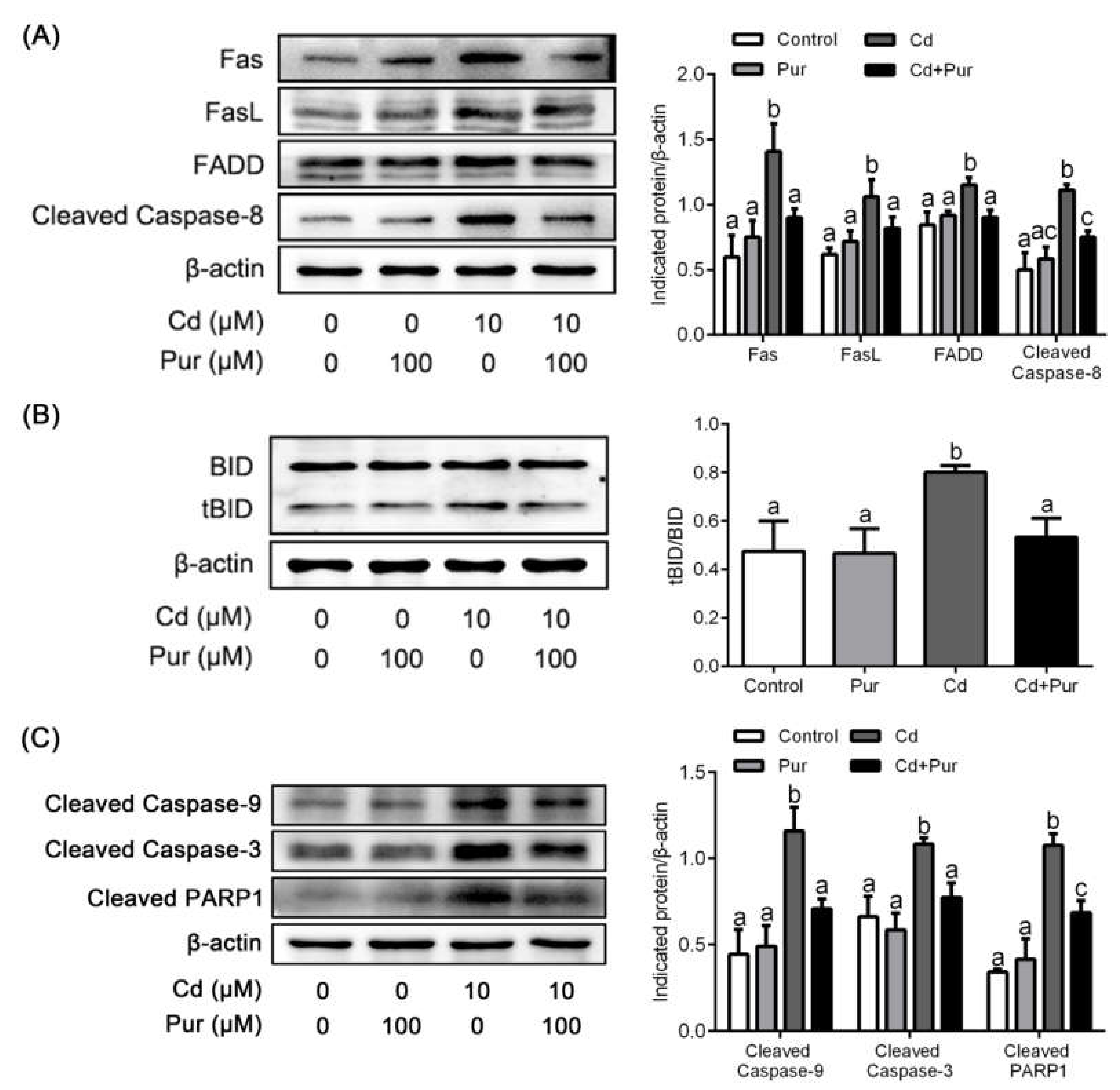
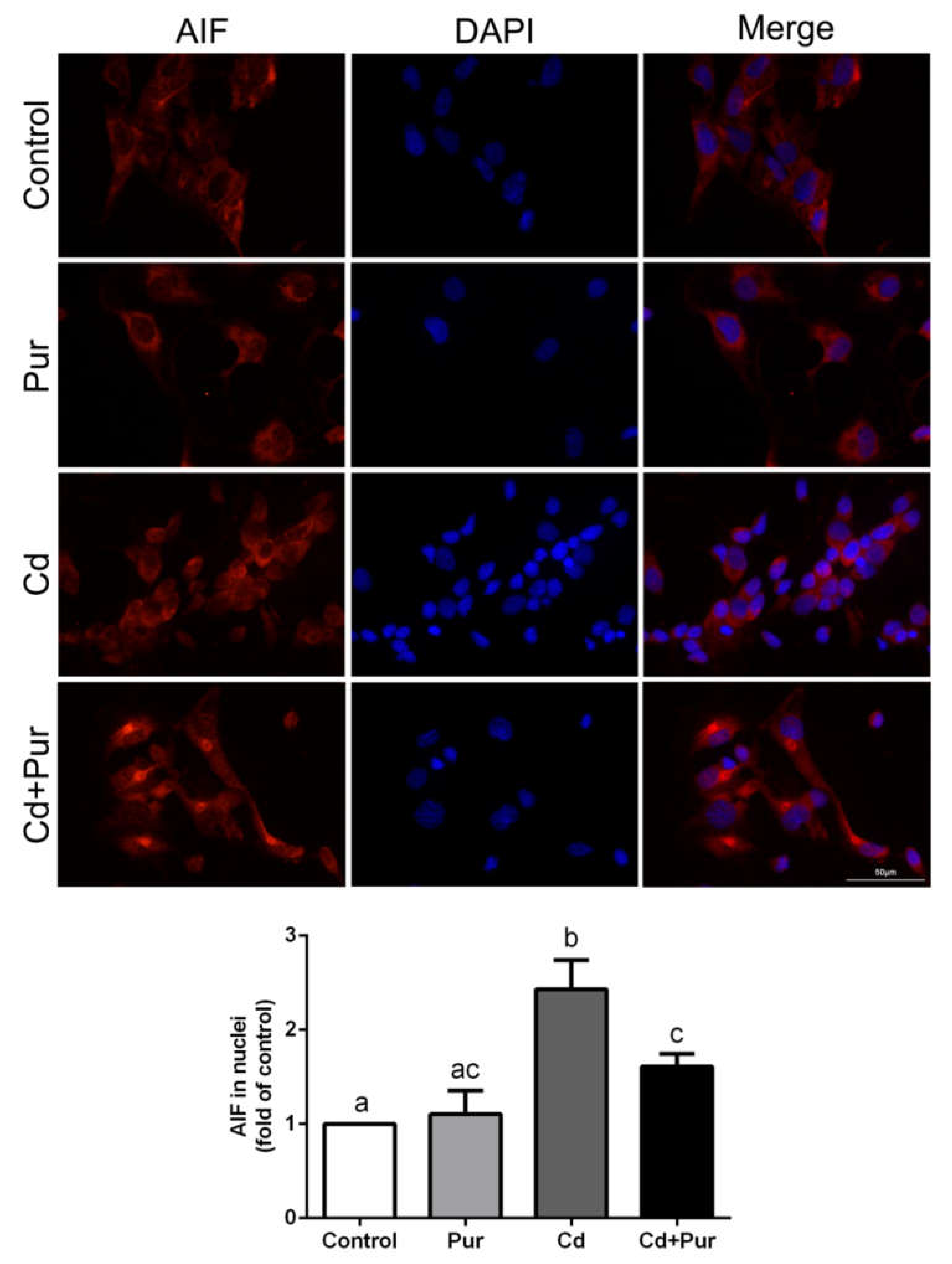
3.4. Pur Attenuates Cd-Induced Apoptosis via Inhibiting the Fas/FasL-Mediated Mitochondrial Apoptotic Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AIF | apoptosis-inducing factor |
| BID | Bcl-2-interacting domain |
| CAT | catalase |
| CCK-8 | cell counting kit-8 |
| Cd | cadmium |
| CMC-Na | Carboxymethylcellulose sodium |
| DAPI | 4′, 6-diamidino-2-phenylindole |
| FADD | Fas-associated death domain protein |
| FasL | Fas Ligand |
| GSH | glutathione |
| H&E | hematoxylin and eosin |
| MDA | malondialdehyde |
| PARP1 | poly (ADP-ribose) polymerase-1 |
| PBS | phosphate-buffered saline |
| Pur | puerarin |
| TBST | Tris-buffered saline containing 0.05% Tween-20 |
| TUNEL | terminal deoxynucleotidyl transferase (TdT)-mediated deoxyuridine triphosphate (dUTP) nick-end labeling |
References
- Jones, M.M.; Cherian, M.G. The search for chelate antagonists for chronic cadmium intoxication. Toxicology 1990, 62, 1–25. [Google Scholar] [CrossRef]
- Gong, Z.-G.; Wang, X.-Y.; Wang, J.-H.; Fan, R.-F.; Wang, L. Trehalose prevents cadmium-induced hepatotoxicity by blocking Nrf2 pathway, restoring autophagy and inhibiting apoptosis. J. Inorg. Biochem. 2019, 192, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Liu, G.; Long, M.; Yang, J.; Song, R.; Wang, Y.; Yuan, Y.; Bian, J.; Liu, X.; Gu, J.; et al. Treatment of cadmium-induced renal oxidative damage in rats by administration of alpha-lipoic acid. Environ. Sci. Pollut. Res. 2017, 24, 1832–1844. [Google Scholar] [CrossRef] [PubMed]
- Momeni, H.R.; Eskandari, N. Curcumin protects the testis against cadmium-induced histopathological damages and oxidative stress in mice. Hum. Exp. Toxicol. 2020, 39, 653–661. [Google Scholar] [CrossRef]
- He, S.; Zhuo, L.; Cao, Y.; Liu, G.; Zhao, H.; Song, R.; Liu, Z. Effect of cadmium on osteoclast differentiation during bone injury in female mice. Environ. Toxicol. 2020, 35, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Yang, J.; Chen, J.; Zhao, S.; Wang, T.; Zou, H.; Wang, Y.; Gu, J.; Liu, X.; Bian, J.; et al. Alpha-lipoic acid protects against cadmium-induced neuronal injury by inhibiting the endoplasmic reticulum stress eIF2alpha-ATF4 pathway in rat cortical neurons in vitro and in vivo. Toxicology 2018, 414, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.; Shukla, G.S.; Srimal, R.C. Cadmium-induced alterations in blood-brain barrier permeability and its possible correlation with decreased microvessel antioxidant potential in rat. Hum. Exp. Toxicol. 1996, 15, 400–405. [Google Scholar] [CrossRef]
- Mendez-Armenta, M.; Rios, C. Cadmium neurotoxicity. Environ. Toxicol. Pharmacol. 2007, 23, 350–358. [Google Scholar] [CrossRef]
- Yan, Y.; Bian, J.C.; Zhong, L.X.; Zhang, Y.; Sun, Y.; Liu, Z.P. Oxidative stress and apoptotic changes of rat cerebral cortical neurons exposed to cadmium in vitro. Biomed. Environ. Sci. 2012, 25, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.K.; Liu, X.Y.; Wang, Z.Y.; Qu, K.C.; Fan, R.F. Trehalose alleviates cadmium-induced brain damage by ameliorating oxidative stress, autophagy inhibition, and apoptosis. Met. Integr. Biometal Sci. 2019, 11, 2043–2051. [Google Scholar] [CrossRef]
- Huang, Q.; Shi, J.; Gao, B.; Zhang, H.Y.; Fan, J.; Li, X.J.; Fan, J.Z.; Han, Y.H.; Zhang, J.K.; Yang, L.; et al. Gastrodin: An ancient Chinese herbal medicine as a source for anti-osteoporosis agents via reducing reactive oxygen species. Bone 2015, 73, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Tang, N.Y.; Kao, S.T.; Hsieh, C.L. Ferulic Acid Administered at Various Time Points Protects against Cerebral Infarction by Activating p38 MAPK/p90RSK/CREB/Bcl-2 Anti-Apoptotic Signaling in the Subacute Phase of Cerebral Ischemia-Reperfusion Injury in Rats. PLoS ONE 2016, 11, e0155748. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Li, G.Q.; Li, K.M.; Razmovski-Naumovski, V.; Chan, K. Kudzu root: Traditional uses and potential medicinal benefits in diabetes and cardiovascular diseases. J. Ethnopharmacol. 2011, 134, 584–607. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Wang, A.; Liu, C.; Li, Y.; Shi, Y.; Zhou, M.S. Puerarin Improves Vascular Insulin Resistance and Cardiovascular Remodeling in Salt-Sensitive Hypertension. Am. J. Chin. Med. 2017, 45, 1169–1184. [Google Scholar] [CrossRef]
- Hsu, F.L.; Liu, I.M.; Kuo, D.H.; Chen, W.C.; Su, H.C.; Cheng, J.T. Antihyperglycemic effect of puerarin in streptozotocin-induced diabetic rats. J. Nat. Prod. 2003, 66, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.J.; Xie, J.J.; Hong, R.H.; Pan, H.S.; Zhang, F.G.; Liang, Y.M. Puerarin alleviates streptozotocin (STZ)-induced osteoporosis in rats through suppressing inflammation and apoptosis via HDAC1/HDAC3 signaling. Biomed. Pharmacother. 2019, 115, 108570. [Google Scholar] [CrossRef]
- Zhou, B.G.; Zhao, H.M.; Lu, X.Y.; Zhou, W.; Liu, F.C.; Liu, X.K.; Liu, D.Y. Effect of Puerarin Regulated mTOR Signaling Pathway in Experimental Liver Injury. Front. Pharmacol. 2018, 9, 1165. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Cao, X.L.; Liu, G.Q.; Zhou, T.; Yang, X.L.; Ma, B.X. The in silico and in vivo evaluation of puerarin against Alzheimer’s disease. Food Funct. 2019, 10, 799–813. [Google Scholar] [CrossRef]
- Kong, H.; Wang, X.; Shi, R.; Zhao, Y.; Cheng, J.; Yan, X.; Liu, X.; Wang, Y.; Zhang, M.; Wang, Q.; et al. Pharmacokinetics and Tissue Distribution Kinetics of Puerarin in Rats Using Indirect Competitive ELISA. Molecules 2017, 22, 939. [Google Scholar] [CrossRef]
- Su, A.S.; Zhang, J.W.; Zou, J. The anxiolytic-like effects of puerarin on an animal model of PTSD. Biomed. Pharmacother. 2019, 115, 108978. [Google Scholar] [CrossRef]
- Xing, G.; Dong, M.; Li, X.; Zou, Y.; Fan, L.; Wang, X.; Cai, D.; Li, C.; Zhou, L.; Liu, J.; et al. Neuroprotective effects of puerarin against beta-amyloid-induced neurotoxicity in PC12 cells via a PI3K-dependent signaling pathway. Brain Res. Bull. 2011, 85, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Xie, N.; Wang, C.; Lian, Y.; Wu, C.; Zhang, H.; Zhang, Q. Puerarin protects hippocampal neurons against cell death in pilocarpine-induced seizures through antioxidant and anti-apoptotic mechanisms. Cell. Mol. Neurobiol. 2014, 34, 1175–1182. [Google Scholar] [CrossRef]
- Om, A.S.; Shim, J.Y. Effect of daidzein in rats on cadmium excretion. Bull. Environ. Contam. Toxicol. 2007, 78, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Paik, M.K.; Lee, H.O.; Chung, H.S.; Yang, S.O.; Kim, J.H.; Om, A.S. Genistein may prevent cadmium-induced bone loss in ovariectomized rats. J. Med. Food 2003, 6, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Jiang, C.Y.; Xu, H.; Sun, Y.; Hu, F.F.; Bian, J.C.; Liu, X.Z.; Gu, J.H.; Liu, Z.P. Cadmium-induced apoptosis in primary rat cerebral cortical neurons culture is mediated by a calcium signaling pathway. PLoS ONE 2013, 8, e64330. [Google Scholar] [CrossRef]
- Wang, B.; Du, Y. Cadmium and its neurotoxic effects. Oxidative Med. Cell. Longev. 2013, 2013, 898034. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Jiang, C.; Hu, F.; Wang, Q.; Zhang, K.; Wang, Y.; Gu, J.; Liu, X.; Bian, J.; Liu, Z. The role of mitogen-activated protein kinase in cadmium-induced primary rat cerebral cortical neurons apoptosis via a mitochondrial apoptotic pathway. J. Trace Elements Med. Biol. Organ Soc. Miner. Trace Elem. 2015, 29, 275–283. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, Y.; Zhao, S.; Chen, J.; Yang, J.; Wang, T.; Zou, H.; Wang, Y.; Gu, J.; Liu, X.; et al. Cadmium-induced apoptosis in neuronal cells is mediated by Fas/FasL-mediated mitochondrial apoptotic signaling pathway. Sci. Rep. 2018, 8, 8837. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.-Y.; Chen, Y.; Xu, X.-Y. Progress on the pharmacological research of puerarin: A review. Chin. J. Nat. Med. 2014, 12, 407–414. [Google Scholar] [CrossRef]
- Xu, M.Y.; Wang, P.; Sun, Y.J.; Wu, Y.J. Metabolomic analysis for combined hepatotoxicity of chlorpyrifos and cadmium in rats. Toxicology 2017, 384, 50–58. [Google Scholar] [CrossRef]
- Reinhardt, C.A.; Pelli, D.A.; Sandvold, M. Cell detachment and growth of fibroblasts as parameters for cytotoxicity of inorganic metal salts in vitro. Cell Biol. Toxicol. 1985, 1, 33–43. [Google Scholar] [CrossRef]
- Li, H.; Wang, P.; Huang, F.; Jin, J.; Wu, H.; Zhang, B.; Wang, Z.; Shi, H.; Wu, X. Astragaloside IV protects blood-brain barrier integrity from LPS-induced disruption via activating Nrf2 antioxidant signaling pathway in mice. Toxicol. Appl. Pharmacol. 2018, 340, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.K.; Hyun, S.W.; Jung, Y.S. Yuzu and Hesperidin Ameliorate Blood-Brain Barrier Disruption during Hypoxia via Antioxidant Activity. Antioxidants 2020, 9, 843. [Google Scholar] [CrossRef]
- Neathery, M.W.; Miller, W.J.; Gentry, R.P.; Stake, P.E.; Blackmon, D.M. Cadmium-109 and methyl mercury-203 metabolism, tissue distribution, and secretion into milk of cows. J. Dairy Sci. 1974, 57, 1177–1183. [Google Scholar] [CrossRef]
- Khan, A.; Ikram, M.; Muhammad, T. Caffeine Modulates Cadmium-Induced Oxidative Stress, Neuroinflammation, and Cognitive Impairments by Regulating Nrf-2/HO-1 In Vivo and In Vitro. J. Clin. Med. 2019, 8, 680. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Ren, Q.; Zhang, J.; Ye, Y.; Zhang, Z.; Xu, Y.; Guo, M.; Ji, H.; Xu, C.; Gu, C.; et al. N-acetyl-L-cysteine protects against cadmium-induced neuronal apoptosis by inhibiting ROS-dependent activation of Akt/mTOR pathway in mouse brain. Neuropathol. Appl. Neurobiol. 2014, 40, 759–777. [Google Scholar] [CrossRef] [Green Version]
- Gan, L.; Johnson, J.A. Oxidative damage and the Nrf2-ARE pathway in neurodegenerative diseases. Biochim. Biophys. Acta 2014, 1842, 1208–1218. [Google Scholar] [CrossRef] [Green Version]
- Bhat, A.H.; Dar, K.B.; Anees, S.; Zargar, M.A.; Masood, A.; Sofi, M.A.; Ganie, S.A. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight. Biomed. Pharmacother. 2015, 74, 101–110. [Google Scholar] [CrossRef]
- Patel, M. Targeting Oxidative Stress in Central Nervous System Disorders. Trends Pharmacol. Sci. 2016, 37, 768–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Rees, H.D.; Weintraub, S.T.; Levey, A.I.; Chin, L.S.; Li, L. Oxidative modifications and aggregation of Cu, Zn-superoxide dismutase associated with Alzheimer and Parkinson diseases. J. Biol. Chem. 2005, 280, 11648–11655. [Google Scholar] [CrossRef] [Green Version]
- Blesa, J.; Trigo-Damas, I.; Quiroga-Varela, A.; Jackson-Lewis, V.R. Oxidative stress and Parkinson’s disease. Front. Neuroanat. 2015, 9, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Xie, N.; Li, L.; Zou, Y.; Zhang, X.; Dong, M. Puerarin alleviates cognitive impairment and oxidative stress in APP/PS1 transgenic mice. Int. J. Neuropsychopharmacol. 2014, 17, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Tong, T.; Wan, S.; Yan, T.; Ren, F.; Bi, K.; Jia, Y. Protective Effects of Puerarin against Abeta 1-42-Induced Learning and Memory Impairments in Mice. Planta Med. 2017, 83, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Lopez, E.; Arce, C.; Oset-Gasque, M.J.; Canadas, S.; Gonzalez, M.P. Cadmium induces reactive oxygen species generation and lipid peroxidation in cortical neurons in culture. Free. Radic. Biol. Med. 2006, 40, 940–951. [Google Scholar] [CrossRef]
- Ben, P.; Zhang, Z.; Xuan, C.; Sun, S.; Shen, L.; Gao, Y.; Cao, X.; Zhou, Y.; Lan, L.; Yin, Z.; et al. Protective Effect of L-Theanine on Cadmium-Induced Apoptosis in PC12 Cells by Inhibiting the Mitochondria-Mediated Pathway. Neurochem. Res. 2015, 40, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xu, B.; Liu, L.; Luo, Y.; Zhou, H.; Chen, W.; Shen, T.; Han, X.; Kontos, C.D.; Huang, S. Cadmium induction of reactive oxygen species activates the mTOR pathway, leading to neuronal cell death. Free. Radic. Biol. Med. 2011, 50, 624–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Budihardjo, I.; Zou, H.; Slaughter, C.; Wang, X. Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell 1998, 94, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Nakabayashi, J.; Sasaki, A. A mathematical model for apoptosome assembly: The optimal cytochrome c/Apaf-1 ratio. J. Theor. Biol. 2006, 242, 280–287. [Google Scholar] [CrossRef]
- Chaitanya, G.V.; Steven, A.J.; Babu, P.P. PARP-1 cleavage fragments: Signatures of cell-death proteases in neurodegeneration. Cell Commun. Signal. CCS 2010, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Sairanen, T.; Szepesi, R.; Karjalainen-Lindsberg, M.L.; Saksi, J.; Paetau, A.; Lindsberg, P.J. Neuronal caspase-3 and PARP-1 correlate differentially with apoptosis and necrosis in ischemic human stroke. Acta Neuropathol. 2009, 118, 541–552. [Google Scholar] [CrossRef]
- Pan, H.P.; Li, G. Protecting mechanism of puerarin on the brain neurocyte of rat in acute local ischemia brain injury and local cerebral ischemia-reperfusion injury. Yakugaku Zasshi J. Pharm. Soc. Jpn. 2008, 128, 1689–1698. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Wang, K.; Zhang, K.; Lin, X.; Zhu, L.; Zhou, F. Puerarin Protects Human Neuroblastoma SH-SY5Y Cells against Glutamate-Induced Oxidative Stress and Mitochondrial Dysfunction. J. Biochem. Mol. Toxicol. 2016, 30, 22–28. [Google Scholar] [CrossRef]
- van Loo, G.; Saelens, X.; van Gurp, M.; MacFarlane, M.; Martin, S.J.; Vandenabeele, P. The role of mitochondrial factors in apoptosis: A Russian roulette with more than one bullet. Cell Death Differ. 2002, 9, 1031–1042. [Google Scholar] [CrossRef] [PubMed]
- Susin, S.A.; Lorenzo, H.K.; Zamzami, N.; Marzo, I.; Snow, B.E.; Brothers, G.M.; Mangion, J.; Jacotot, E.; Costantini, P.; Loeffler, M.; et al. Molecular characterization of mitochondrial apoptosis-inducing factor. Nature 1999, 397, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Xu, X.; Ye, D.; Chen, W.; Gao, B.; Huang, Y. Caspase/AIF/apoptosis pathway: A new target of puerarin for diabetes mellitus therapy. Mol. Biol. Rep. 2019, 46, 4787–4797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ettinger, B.; Quesenberry, C.; Schroeder, D.A.; Friedman, G. Long-term postmenopausal estrogen therapy may be associated with increased risk of breast cancer: A cohort study. Menopause 2018, 25, 1191–1194. [Google Scholar] [CrossRef] [PubMed]
- Hammond, C.B.; Jelovsek, F.R.; Lee, K.L.; Creasman, W.T.; Parker, R.T. Effects of long-term estrogen replacement therapy. I. Metabolic effects. Am. J. Obstet. Gynecol. 1979, 133, 525–536. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Control | Pur | Cd | Cd + Pur |
|---|---|---|---|---|
| Cd consumption (mg/kg body weight/day) | — | — | 3.64 ± 0.52 a | 3.71 ± 0.48 a |
| Cerebral cortical Cd (μg/g) | 0.028 ± 0.008 a | 0.027 ± 0.007 a | 0.094 ± 0.014 b | 0.042 ± 0.005 a |
| Serum Cd (μg/L) | 0.35 ± 0.09 a | 0.32 ± 0.08 a | 1.52 ± 0.18 b | 0.87 ± 0.06 c |
| Urinary Cd (μg/L) | 1.59 ± 0.06 a | 1.66 ± 0.30 a | 36.46 ± 4.30 b | 63.96 ± 8.84 c |
| Fecal Cd (μg/g) | 0.56 ± 0.04 a | 0.62 ± 0.06 a | 148.34 ± 16.75 b | 219.96 ± 15.00 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, S.; Wang, L.; Zou, H.; Gu, J.; Song, R.; Bian, J.; Yuan, Y.; Liu, Z. Puerarin Attenuates Cadmium-Induced Neuronal Injury via Stimulating Cadmium Excretion, Inhibiting Oxidative Stress and Apoptosis. Biomolecules 2021, 11, 978. https://doi.org/10.3390/biom11070978
Wen S, Wang L, Zou H, Gu J, Song R, Bian J, Yuan Y, Liu Z. Puerarin Attenuates Cadmium-Induced Neuronal Injury via Stimulating Cadmium Excretion, Inhibiting Oxidative Stress and Apoptosis. Biomolecules. 2021; 11(7):978. https://doi.org/10.3390/biom11070978
Chicago/Turabian StyleWen, Shuangquan, Li Wang, Hui Zou, Jianhong Gu, Ruilong Song, Jianchun Bian, Yan Yuan, and Zongping Liu. 2021. "Puerarin Attenuates Cadmium-Induced Neuronal Injury via Stimulating Cadmium Excretion, Inhibiting Oxidative Stress and Apoptosis" Biomolecules 11, no. 7: 978. https://doi.org/10.3390/biom11070978
APA StyleWen, S., Wang, L., Zou, H., Gu, J., Song, R., Bian, J., Yuan, Y., & Liu, Z. (2021). Puerarin Attenuates Cadmium-Induced Neuronal Injury via Stimulating Cadmium Excretion, Inhibiting Oxidative Stress and Apoptosis. Biomolecules, 11(7), 978. https://doi.org/10.3390/biom11070978