Promising Opportunities for Treating Neurodegenerative Diseases with Mesenchymal Stem Cell-Derived Exosomes



Abstract
1. Introduction
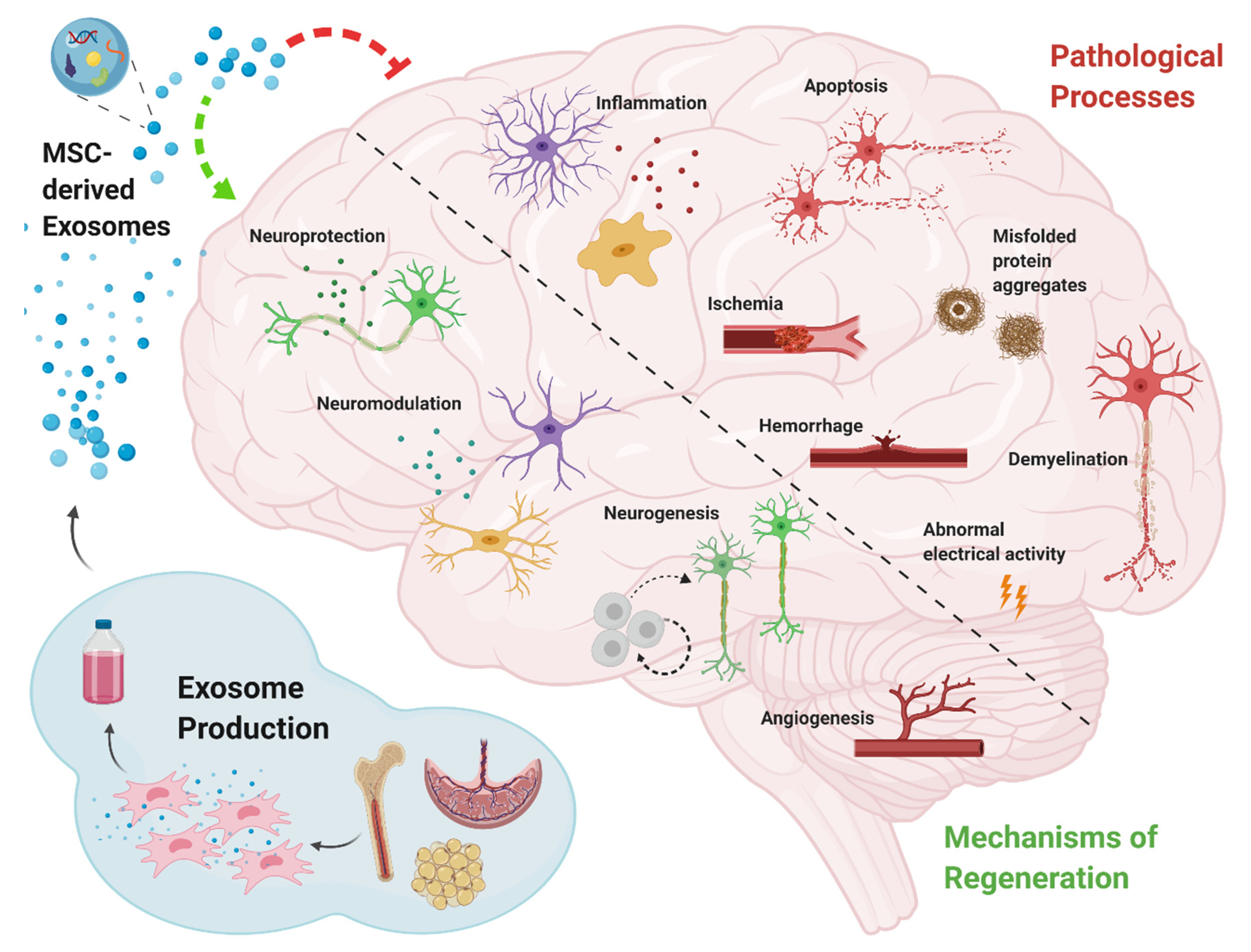
2. MSC-Derived Exosomes as a Therapeutic Tool
3. Clinical Outcomes following MSC-Derived Exosome Treatment in Neurodegenerative Animal Models
4. Mechanism of Action
5. Limitations of Current Knowledge
6. Toward MSC-Derived Exosome-Based Therapies
7. MSC-Derived Exosomes in Clinical Trials
8. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deans, R.J.; Moseley, A.B. Mesenchymal stem cells: Biology and potential clinical uses. Exp. Hematol. 2000, 28, 875–884. [Google Scholar] [CrossRef]
- Parekkadan, B.; Milwid, J.M. Mesenchymal Stem Cells as Therapeutics. Annu. Rev. Biomed. Eng. 2010, 12, 87–117. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, P.; Halabian, R.; Imani Fooladi, A.A. A revealing review of mesenchymal stem cells therapy, clinical perspectives and Modification strategies. Stem Cell Investig. 2019, 6, 34. [Google Scholar] [CrossRef]
- Volkman, R.; Offen, D. Concise Review: Mesenchymal Stem Cells in Neurodegenerative Diseases. Stem Cells 2017, 35, 1867–1880. [Google Scholar] [CrossRef] [PubMed]
- Joyce, N.; Annett, G.; Wirthlin, L.; Olson, S.; Bauer, G.; Nolta, J.A. Mesenchymal stem cells for the treatment of neurodegenerative disease. Regen. Med. 2010, 5, 933–946. [Google Scholar] [CrossRef]
- Tanna, T.; Sachan, V. Mesenchymal Stem Cells: Potential in Treatment of Neurodegenerative Diseases. Curr. Stem Cell Res. Ther. 2014, 9, 513–521. [Google Scholar] [CrossRef]
- Eliopoulos, N.; Stagg, J.; Lejeune, L.; Pommey, S.; Galipeau, J. Allogeneic marrow stromal cells are immune rejected by MHC class I- and class II-mismatched recipient mice. Blood 2005, 106, 4057–4065. [Google Scholar] [CrossRef]
- Zangi, L.; Margalit, R.; Reich-Zeliger, S.; Bachar-Lustig, E.; Beilhack, A.; Negrin, R.; Reisner, Y. Direct Imaging of Immune Rejection and Memory Induction by Allogeneic Mesenchymal Stromal Cells. Stem Cells 2009, 27, 2865–2874. [Google Scholar] [CrossRef]
- Ma, N.; Cheng, H.; Lu, M.; Liu, Q.; Chen, X.; Yin, G.; Zhu, H.; Zhang, L.; Meng, X.; Tang, Y.; et al. Magnetic Resonance Imaging with Superparamagnetic Iron Oxide Fails to Track the Long-term Fate of Mesenchymal Stem Cells Transplanted into Heart. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef]
- Cui, L.L.; Kerkelä, E.; Bakreen, A.; Nitzsche, F.; Andrzejewska, A.; Nowakowski, A.; Janowski, M.; Walczak, P.; Boltze, J.; Lukomska, B.; et al. The cerebral embolism evoked by intra-arterial delivery of allogeneic bone marrow mesenchymal stem cells in rats is related to cell dose and infusion velocity. Stem Cell Res. Ther. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Nauta, A.J.; Westerhuis, G.; Kruisselbrink, A.B.; Lurvink, E.G.A.; Willemze, R.; Fibbe, W.E. Donor-derived mesenchymal stem cells are immunogenic in an allogeneic host and stimulate donor graft rejection in a nonmyeloablative setting. Blood 2006, 108, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Reinders, M.E.J.; de Fijter, J.W.; Roelofs, H.; Bajema, I.M.; de Vries, D.K.; Schaapherder, A.F.; Claas, F.H.J.; van Miert, P.P.M.C.; Roelen, D.L.; van Kooten, C.; et al. Autologous Bone Marrow-Derived Mesenchymal Stromal Cells for the Treatment of Allograft Rejection After Renal Transplantation: Results of a Phase I Study. Stem Cells Transl. Med. 2013, 2, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Røsland, G.V.; Svendsen, A.; Torsvik, A.; Sobala, E.; McCormack, E.; Immervoll, H.; Mysliwietz, J.; Tonn, J.C.; Goldbrunner, R.; Lønning, P.E.; et al. Long-term cultures of bone marrow-derived human mesenchymal stem cells frequently undergo spontaneous malignant transformation. Cancer Res. 2009, 69, 5331–5339. [Google Scholar] [CrossRef]
- Miura, M.; Miura, Y.; Padilla-Nash, H.M.; Molinolo, A.A.; Fu, B.; Patel, V.; Seo, B.-M.; Sonoyama, W.; Zheng, J.J.; Baker, C.C.; et al. Accumulated Chromosomal Instability in Murine Bone Marrow Mesenchymal Stem Cells Leads to Malignant Transformation. Stem Cells 2006, 24, 1095–1103. [Google Scholar] [CrossRef]
- Eggenhofer, E.; Benseler, V.; Kroemer, A.; Popp, F.C.; Geissler, E.K.; Schlitt, H.J.; Baan, C.C.; Dahlke, M.H.; Hoogduijn, M.J. Mesenchymal stem cells are short-lived and do not migrate beyond the lungs after intravenous infusion. Front. Immunol. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Eggenhofer, E.; Luk, F.; Dahlke, M.H.; Hoogduijn, M.J. The life and fate of mesenchymal stem cells. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef]
- Rani, S.; Ryan, A.E.; Griffin, M.D.; Ritter, T. Mesenchymal stem cell-derived extracellular vesicles: Toward cell-free therapeutic applications. Mol. Ther. 2015, 23, 812–823. [Google Scholar] [CrossRef]
- Ng, K.S.; Kuncewicz, T.M.; Karp, J.M. Beyond hit-and-run: Stem cells leave a lasting memory. Cell Metab. 2015, 22, 541–543. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.-H.; Jeyaraj, M.; Qasim, M.; Kim, J.-H. Review of the Isolation, Characterization, Biological Function, and Multifarious Therapeutic Approaches of Exosomes. Cells 2019, 8, 307. [Google Scholar] [CrossRef]
- Sarvar, D.P.; Shamsasenjan, K.; Akbarzadehlaleh, P. Mesenchymal stem cell-derived exosomes: New opportunity in cell-free therapy. Adv. Pharm. Bull. 2016, 6, 293–299. [Google Scholar] [CrossRef]
- Yin, K.; Wang, S.; Zhao, R.C. Exosomes from mesenchymal stem/stromal cells: A new therapeutic paradigm. Biomark. Res. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Aghajani Nargesi, A.; Lerman, L.O.; Eirin, A. Mesenchymal stem cell-derived extracellular vesicles for kidney repair: Current status and looming challenges. Stem Cell Res. Ther. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Xiao, L.; Peng, J.; Qian, Y.Q.; Huang, C.C. Exosomes derived from bone marrow mesenchymal stem cells improve Osteoporosis through promoting osteoblast proliferation via MAPK pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3962–3970. [Google Scholar] [CrossRef]
- Li, Q.C.; Liang, Y.; Su, Z.B. Prophylactic treatment with MSC-derived exosomes attenuates traumatic acute lung injury in rats. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 316, L1107–L1117. [Google Scholar] [CrossRef]
- Zou, L.; Ma, X.; Lin, S.; Wu, B.; Chen, Y.; Peng, C. Bone marrow mesenchymal stem cell-derived exosomes protect against myocardial infarction by promoting autophagy. Exp. Ther. Med. 2019, 18, 2574. [Google Scholar] [CrossRef]
- Harrell, C.R.; Fellabaum, C.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Molecular Mechanisms Responsible for Therapeutic Potential of Mesenchymal Stem Cell-Derived Secretome. Cells 2019, 8, 467. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.C.; Yip, H.K.; Shao, P.L.; Wu, S.C.; Chen, K.H.; Chen, Y.T.; Yang, C.C.; Sun, C.K.; Kao, G.S.; Chen, S.Y.; et al. Combination of adipose-derived mesenchymal stem cells (ADMSC) and ADMSC-derived exosomes for protecting kidney from acute ischemia-reperfusion injury. Int. J. Cardiol. 2016, 216, 173–185. [Google Scholar] [CrossRef]
- Li, Z.; Liu, F.; He, X.; Yang, X.; Shan, F.; Feng, J. Exosomes derived from mesenchymal stem cells attenuate inflammation and demyelination of the central nervous system in EAE rats by regulating the polarization of microglia. Int. Immunopharmacol. 2019, 67, 268–280. [Google Scholar] [CrossRef]
- Riazifar, M.; Mohammadi, M.R.; Pone, E.J.; Yeri, A.; Lasser, C.; Segaliny, A.I.; McIntyre, L.L.; Shelke, G.V.; Hutchins, E.; Hamamoto, A.; et al. Stem Cell-Derived Exosomes as Nanotherapeutics for Autoimmune and Neurodegenerative Disorders. ACS Nano 2019, 13, 6670–6688. [Google Scholar] [CrossRef]
- Zhao, C.; Zhou, X.; Qiu, J.; Xin, D.; Li, T.; Chu, X.; Yuan, H.; Wang, H.; Wang, Z.; Wang, D. Exosomes derived from bone marrow mesenchymal stem cells inhibit complement activation in rats with spinal cord injury. Drug Des. Devel. Ther. 2019, 13, 3693–3704. [Google Scholar] [CrossRef]
- Huang, J.-H.; Yin, X.-M.; Xu, Y.; Xu, C.-C.; Lin, X.; Ye, F.-B.; Cao, Y.; Lin, F.-Y. Systemic Administration of Exosomes Released from Mesenchymal Stromal Cells Attenuates Apoptosis, Inflammation, and Promotes Angiogenesis after Spinal Cord Injury in Rats. J. Neurotrauma 2017, 34, 3388–3396. [Google Scholar] [CrossRef] [PubMed]
- Otero-Ortega, L.; Laso-García, F.; Frutos, M.C.G.-d.; Diekhorst, L.; Martínez-Arroyo, A.; Alonso-López, E.; García-Bermejo, M.L.; Rodríguez-Serrano, M.; Arrúe-Gonzalo, M.; Díez-Tejedor, E.; et al. Low dose of extracellular vesicles identified that promote recovery after ischemic stroke. Stem Cell Res. Ther. 2020, 11, 70. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gan, Y.; Xu, G.; Yin, G.; Liu, D. MSCs-Derived Exosomes Attenuate Acute Brain Injury and Inhibit Microglial Inflammation by Reversing CysLT2R-ERK1/2 Mediated Microglia M1 Polarization. Neurochem. Res. 2020, 45, 1180–1190. [Google Scholar] [CrossRef] [PubMed]
- Otero-Ortega, L.; Gómez de Frutos, M.C.; Laso-García, F.; Rodríguez-Frutos, B.; Medina-Gutiérrez, E.; López, J.A.; Vázquez, J.; Díez-Tejedor, E.; Gutiérrez-Fernández, M. Exosomes promote restoration after an experimental animal model of intracerebral hemorrhage. J. Cereb. Blood Flow Metab. 2018, 38, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Otero-Ortega, L.; Laso-García, F.; Del Carmen Gómez-De Frutos, M.; Rodríguez-Frutos, B.; Pascual-Guerra, J.; Fuentes, B.; Díez-Tejedor, E.; Gutiérrez-Fernández, M. White matter repair after extracellular vesicles administration in an experimental animal model of subcortical stroke. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Chen, K.H.; Chen, C.H.; Wallace, C.G.; Yuen, C.M.; Kao, G.S.; Chen, Y.L.; Shao, P.L.; Chen, Y.L.; Chai, H.T.; Lin, K.C.; et al. Intravenous administration of xenogenic adipose-derived mesenchymal stem cells (ADMSC) and ADMSC-derived exosomes markedly reduced brain infarct volume and preserved neurological function in rat after acute ischemic stroke. Oncotarget 2016, 7, 74537–74556. [Google Scholar] [CrossRef]
- Doeppner, T.R.; Herz, J.; Görgens, A.; Schlechter, J.; Ludwig, A.-K.; Radtke, S.; de Miroschedji, K.; Horn, P.A.; Giebel, B.; Hermann, D.M. Extracellular Vesicles Improve Post-Stroke Neuroregeneration and Prevent Postischemic Immunosuppression. Stem Cells Transl. Med. 2015, 4, 1131–1143. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Chu, X.; Yuan, H.; Qiu, J.; Zhao, C.; Xin, D.; Li, T.; Ma, W.; Wang, H.; Wang, Z.; et al. Mesenchymal stem cell derived EVs mediate neuroprotection after spinal cord injury in rats via the microRNA-21-5p/FasL gene axis. Biomed. Pharmacother. 2019, 115, 108818. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Y.; Gong, F.; Rong, Y.; Luo, Y.; Tang, P.; Zhou, Z.; Zhou, Z.; Xu, T.; Jiang, T.; et al. Exosomes derived from bone mesenchymal stem cells repair traumatic spinal cord injury by suppressing the activation of a1 neurotoxic reactive astrocytes. J. Neurotrauma 2019, 36, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Fei, K.; Tian, F.; Gao, C.; Song, Y. Adipose-derived mesenchymal stem cells attenuate ischemic brain injuries in rats by modulating miR-21-3p/MAT2B signaling transduction. Croat. Med. J. 2019, 60, 439–448. [Google Scholar] [CrossRef]
- Ni, H.; Yang, S.; Siaw-Debrah, F.; Hu, J.; Wu, K.; He, Z.; Yang, J.; Pan, S.; Lin, X.; Ye, H.; et al. Exosomes Derived From Bone Mesenchymal Stem Cells Ameliorate Early Inflammatory Responses Following Traumatic Brain Injury. Front. Neurosci. 2019, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhou, Y.; Zhang, R.; Wen, L.; Wu, K.; Li, Y.; Yao, Y.; Duan, R.; Jia, Y. Bone Mesenchymal Stem Cell-Derived Extracellular Vesicles Promote Recovery Following Spinal Cord Injury via Improvement of the Integrity of the Blood-Spinal Cord Barrier. Front. Neurosci. 2019, 13. [Google Scholar] [CrossRef]
- Sisa, C.; Kholia, S.; Naylor, J.; Sanchez, M.B.H.; Bruno, S.; Deregibus, M.C.; Camussi, G.; Inal, J.M.; Lange, S.; Hristova, M. Mesenchymal stromal cell derived extracellular vesicles reduce hypoxia-ischaemia induced perinatal injury. Front. Physiol. 2019, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- El-Mahalaway, A.M.; El-Azab, N.E.-E. The potential neuroprotective role of mesenchymal stem cell-derived exosomes in cerebellar cortex lipopolysaccharide-induced neuroinflammation in rats: A histological and immunohistochemical study. Ultrastruct. Pathol. 2020, 1–15. [Google Scholar] [CrossRef]
- Elia, C.A.; Tamborini, M.; Rasile, M.; Desiato, G.; Marchetti, S.; Swuec, P.; Mazzitelli, S.; Clemente, F.; Anselmo, A.; Matteoli, M.; et al. Intracerebral Injection of Extracellular Vesicles from Mesenchymal Stem Cells Exerts Reduced Aβ Plaque Burden in Early Stages of a Preclinical Model of Alzheimer’s Disease. Cells 2019, 8, 1059. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Jia, J.; Wang, Z. Mesenchymal Stem Cell-Derived Extracellular Vesicles Suppresses iNOS Expression and Ameliorates Neural Impairment in Alzheimer’s Disease Mice. J. Alzheimer’s Dis. 2018, 61, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Xiao, H.; Zhang, H.; Peng, H.; Yuan, H.; Xu, Y.; Zhang, G.; Hu, Z. Mesenchymal Stem Cell-Derived Extracellular Vesicles Ameliorates Hippocampal Synaptic Impairment after Transient Global Ischemia. Front. Cell. Neurosci. 2017, 11, 205. [Google Scholar] [CrossRef]
- Wang, L.; Pei, S.; Han, L.; Guo, B.; Li, Y.; Duan, R.; Yao, Y.; Xue, B.; Chen, X.; Jia, Y. Mesenchymal Stem Cell-Derived Exosomes Reduce A1 Astrocytes via Downregulation of Phosphorylated NFκB P65 Subunit in Spinal Cord Injury. Cell. Physiol. Biochem. 2018, 50, 1535–1559. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhang, H.; Yan, J.; Ma, X. An experimental study on the treatment of diabetes-induced cognitive disorder mice model with exosomes deriving from mesenchymal stem cells (MSCs). Pak. J. Pharm. Sci. 2019, 32, 1965–1970. [Google Scholar]
- Reza-Zaldivar, E.E.; Hernández-Sapiéns, M.A.; Gutiérrez-Mercado, Y.K.; Sandoval-Ávila, S.; Gomez-Pinedo, U.; Márquez-Aguirre, A.L.; Vázquez-Méndez, E.; Padilla-Camberos, E.; Canales-Aguirre, A.A. Mesenchymal stem cell-derived exosomes promote neurogenesis and cognitive function recovery in a mouse model of Alzheimer’s disease. Neural Regen. Res. 2019, 14, 1626–1634. [Google Scholar] [CrossRef]
- Ding, M.; Shen, Y.; Wang, P.; Xie, Z.; Xu, S.; Zhu, Z.Y.; Wang, Y.; Lyu, Y.; Wang, D.; Xu, L.; et al. Exosomes Isolated From Human Umbilical Cord Mesenchymal Stem Cells Alleviate Neuroinflammation and Reduce Amyloid-Beta Deposition by Modulating Microglial Activation in Alzheimer’s Disease. Neurochem. Res. 2018, 43, 2165–2177. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Rabea, A.A.; Ragae, A.; Sabry, D. The prospective role of mesenchymal stem cells exosomes on circumvallate taste buds in induced Alzheimer’s disease of ovariectomized albino rats: (Light and transmission electron microscopic study). Arch. Oral Biol. 2020, 110, 104596. [Google Scholar] [CrossRef] [PubMed]
- Moore, T.L.; Bowley, B.G.E.; Pessina, M.A.; Calderazzo, S.M.; Medalla, M.; Go, V.; Zhang, Z.G.; Chopp, M.; Finklestein, S.; Harbaugh, A.G.; et al. Mesenchymal derived exosomes enhance recovery of motor function in a monkey model of cortical injury. Restor. Neurol. Neurosci. 2019, 37, 347–362. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Seyfried, D.; Meng, Y.; Yang, D.; Schultz, L.; Chopp, M.; Seyfried, D. Multipotent mesenchymal stromal cell-derived exosomes improve functional recovery after experimental intracerebral hemorrhage in the rat. J. Neurosurg. 2019, 131, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Nalamolu, K.R.; Venkatesh, I.; Mohandass, A.; Klopfenstein, J.D.; Pinson, D.M.; Wang, D.Z.; Veeravallia, K.K. Exosomes treatment mitigates ischemic brain damage but does not improve post-stroke neurological outcome. Cell. Physiol. Biochem. 2019, 52, 1280–1291. [Google Scholar] [CrossRef] [PubMed]
- Thomi, G.; Joerger-Messerli, M.; Haesler, V.; Muri, L.; Surbek, D.; Schoeberlein, A. Intranasally Administered Exosomes from Umbilical Cord Stem Cells Have Preventive Neuroprotective Effects and Contribute to Functional Recovery after Perinatal Brain Injury. Cells 2019, 8, 855. [Google Scholar] [CrossRef] [PubMed]
- Drommelschmidt, K.; Serdar, M.; Bendix, I.; Herz, J.; Bertling, F.; Prager, S.; Keller, M.; Ludwig, A.K.; Duhan, V.; Radtke, S.; et al. Mesenchymal stem cell-derived extracellular vesicles ameliorate inflammation-induced preterm brain injury. Brain. Behav. Immun. 2017, 60, 220–232. [Google Scholar] [CrossRef]
- Ophelders, D.R.M.G.; Wolfs, T.G.A.M.; Jellema, R.K.; Zwanenburg, A.; Andriessen, P.; Delhaas, T.; Ludwig, A.-K.; Radtke, S.; Peters, V.; Janssen, L.; et al. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Protect the Fetal Brain After Hypoxia-Ischemia. Stem Cells Transl. Med. 2016, 5, 754–763. [Google Scholar] [CrossRef]
- Zhang, Y.; Chopp, M.; Meng, Y.; Katakowski, M.; Xin, H.; Mahmood, A.; Xiong, Y. Effect of exosomes derived from multipluripotent mesenchymal stromal cells on functional recovery and neurovascular plasticity in rats after traumatic brain injury. J. Neurosurg. 2015, 122, 856–867. [Google Scholar] [CrossRef]
- Kim, D.K.; Nishida, H.; An, S.Y.; Shetty, A.K.; Bartosh, T.J.; Prockop, D.J. Chromatographically isolated CD63+CD81+ extracellular vesicles from mesenchymal stromal cells rescue cognitive impairments after TBI. Proc. Natl. Acad. Sci. USA 2016, 113, 170–175. [Google Scholar] [CrossRef]
- Williams, A.M.; Dennahy, I.S.; Bhatti, U.F.; Halaweish, I.; Xiong, Y.; Chang, P.; Nikolian, V.C.; Chtraklin, K.; Brown, J.; Zhang, Y.; et al. Mesenchymal Stem Cell-Derived Exosomes Provide Neuroprotection and Improve Long-Term Neurologic Outcomes in a Swine Model of Traumatic Brain Injury and Hemorrhagic Shock. J. Neurotrauma 2019, 36, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Li, G.; Li, D.; Huang, W.; Zhang, R.; Zhang, H.; Duan, Y.; Wang, B. hucMSC derived exosomes promote functional recovery in spinal cord injury mice via attenuating inflammation. Mater. Sci. Eng. C 2018, 89, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Ruppert, K.A.; Nguyen, T.T.; Prabhakara, K.S.; Toledano Furman, N.E.; Srivastava, A.K.; Harting, M.T.; Cox, C.S.; Olson, S.D. Human Mesenchymal Stromal Cell-Derived Extracellular Vesicles Modify Microglial Response and Improve Clinical Outcomes in Experimental Spinal Cord Injury. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Xian, P.; Hei, Y.; Wang, R.; Wang, T.; Yang, J.; Li, J.; Di, Z.; Liu, Z.; Baskys, A.; Liu, W.; et al. Mesenchymal stem cell-derived exosomes as a nanotherapeutic agent for amelioration of inflammation-induced astrocyte alterations in mice. Theranostics 2019, 9, 5956–5975. [Google Scholar] [CrossRef] [PubMed]
- Longa, Q.; Upadhya, D.; Hattiangady, B.; Kim, D.K.; An, S.Y.; Shuai, B.; Prockop, D.J.; Shetty, A.K. Intranasal MSC-derived A1-exosomes ease inflammation, and prevent abnormal neurogenesis and memory dysfunction after status epilepticus. Proc. Natl. Acad. Sci. USA 2017, 114, E3536–E3545. [Google Scholar] [CrossRef]
- Perets, N.; Hertz, S.; London, M.; Offen, D. Intranasal administration of exosomes derived from mesenchymal stem cells ameliorates autistic-like behaviors of BTBR mice. Mol. Autism 2018, 9. [Google Scholar] [CrossRef]
- Liang, Y.; Duan, L.; Xu, X.; Li, X.; Liu, M.; Chen, H.; Lu, J.; Xia, J. Mesenchymal Stem Cell—Derived Exosomes for Treatment of Autism Spectrum Disorder. ACS Appl. Bio Mater. 2020. [Google Scholar] [CrossRef]
- Tracy, S.A.; Ahmed, A.; Tigges, J.C.; Ericsson, M.; Pal, A.K.; Zurakowski, D.; Fauza, D.O. A comparison of clinically relevant sources of mesenchymal stem cell-derived exosomes: Bone marrow and amniotic fluid. J. Pediatr. Surg. 2019, 54, 86–90. [Google Scholar] [CrossRef]
- Lopez-Verrilli, M.A.; Caviedes, A.; Cabrera, A.; Sandoval, S.; Wyneken, U.; Khoury, M. Mesenchymal stem cell-derived exosomes from different sources selectively promote neuritic outgrowth. Neuroscience 2016, 320, 129–139. [Google Scholar] [CrossRef]
- Xin, H.; Li, Y.; Cui, Y.; Yang, J.J.; Zhang, Z.G.; Chopp, M. Systemic Administration of Exosomes Released from Mesenchymal Stromal Cells Promote Functional Recovery and Neurovascular Plasticity After Stroke in Rats. J. Cereb. Blood Flow Metab. 2013, 33, 1711–1715. [Google Scholar] [CrossRef]
- Przedborski, S.; Vila, M.; Jackson-Lewis, V. Series Introduction: Neurodegeneration: What is it and where are we? J. Clin. Investig. 2003, 111, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Moon, G.J.; Sung, J.H.; Kim, D.H.; Kim, E.H.; Cho, Y.H.; Son, J.P.; Cha, J.M.; Bang, O.Y. Application of Mesenchymal Stem Cell-Derived Extracellular Vesicles for Stroke: Biodistribution and MicroRNA Study. Transl. Stroke Res. 2019, 10, 509–521. [Google Scholar] [CrossRef]
- Joo, H.S.; Suh, J.H.; Lee, H.J.; Bang, E.S.; Lee, J.M. Current knowledge and future perspectives on mesenchymal stem cell-derived exosomes as a new therapeutic agent. Int. J. Mol. Sci. 2020, 21, 727. [Google Scholar] [CrossRef]
- Reza-Zaldivar, E.E.; Hernández-Sapiéns, M.A.; Minjarez, B.; Gutiérrez-Mercado, Y.K.; Márquez-Aguirre, A.L.; Canales-Aguirre, A.A. Potential Effects of MSC-Derived Exosomes in Neuroplasticity in Alzheimer’s Disease. Front. Cell. Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Chopp, M.; Li, Y. Treatment of neural injury with marrow stromal cells. Lancet Neurol. 2002, 1, 92–100. [Google Scholar] [CrossRef]
- Katsuda, T.; Tsuchiya, R.; Kosaka, N.; Yoshioka, Y.; Takagaki, K.; Oki, K.; Takeshita, F.; Sakai, Y.; Kuroda, M.; Ochiya, T. Human adipose tissue-derived mesenchymal stem cells secrete functional neprilysin-bound exosomes. Sci. Rep. 2013, 3, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Zhang, G.; Zhang, L.; Hu, Y.; Zhang, K.; Sun, X.; Zhao, C.; Li, H.; Li, Y.M.; Zhao, J. Mesenchymal stem cells deliver exogenous miR-21 via exosomes to inhibit nucleus pulposus cell apoptosis and reduce intervertebral disc degeneration. J. Cell. Mol. Med. 2018, 22, 261–276. [Google Scholar] [CrossRef]
- Ma, J.F.; Zang, L.N.; Xi, Y.M.; Yang, W.J.; Zou, D. MiR-125a Rs12976445 Polymorphism is Associated with the Apoptosis Status of Nucleus Pulposus Cells and the Risk of Intervertebral Disc Degeneration. Cell. Physiol. Biochem. 2016, 38, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Baglio, S.R.; Rooijers, K.; Koppers-Lalic, D.; Verweij, F.J.; Pérez Lanzón, M.; Zini, N.; Naaijkens, B.; Perut, F.; Niessen, H.W.M.; Baldini, N.; et al. Human bone marrow- and adipose-mesenchymal stem cells secrete exosomes enriched in distinctive miRNA and tRNA species. Stem Cell Res. Ther. 2015, 6. [Google Scholar] [CrossRef]
- Vilaça-Faria, H.; Salgado, A.J.; Teixeira, F.G. Mesenchymal Stem Cells-derived Exosomes: A New Possible Therapeutic Strategy for Parkinson’s Disease? Cells 2019, 8, 118. [Google Scholar] [CrossRef]
- Göran Ronquist, K. Extracellular vesicles and energy metabolism. Clin. Chim. Acta 2019, 488, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Riau, A.K.; Ong, H.S.; Yam, G.H.F.; Mehta, J.S. Sustained Delivery System for Stem Cell-Derived Exosomes. Front. Pharmacol. 2019, 10, 1368. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Takahashi, Y.; Nishikawa, M.; Takakura, Y. Effect of exosome isolation methods on physicochemical properties of exosomes and clearance of exosomes from the blood circulation. Eur. J. Pharm. Biopharm. 2016, 98, 1–8. [Google Scholar] [CrossRef]
- McKelvey, K.J.; Powell, K.L.; Ashton, A.W.; Morris, J.M.; McCracken, S.A. Exosomes: Mechanisms of Uptake. J. Circ. Biomarkers 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Lin, E.Y.; Chiou, T.W.; Harn, H.J. Exosomes in clinical trial and their production in compliance with good manufacturing practice. Tzu Chi Med. J. 2020, 32, 113–120. [Google Scholar] [CrossRef]
- Betzer, O.; Perets, N.; Angel, A.; Motiei, M.; Sadan, T.; Yadid, G.; Offen, D.; Popovtzer, R. In Vivo Neuroimaging of Exosomes Using Gold 2 Nanoparticles. ACS Nano 2017, 11, 10883–10893. [Google Scholar] [CrossRef]
- Perets, N.; Betzer, O.; Shapira, R.; Brenstein, S.; Angel, A.; Sadan, T.; Ashery, U.; Popovtzer, R.; Offen, D. Golden Exosomes Selectively Target Brain Pathologies in Neurodegenerative and Neurodevelopmental Disorders. Nano Lett. 2019, 19, 3422–3431. [Google Scholar] [CrossRef]
- Hou, K.; Li, G.; Zhao, J.; Xu, B.; Zhang, Y.; Yu, J.; Xu, K. Bone mesenchymal stem cell-derived exosomal microRNA-29b-3p prevents hypoxic-ischemic injury in rat brain by activating the PTEN-mediated Akt signaling pathway. J. Neuroinflamm. 2020, 17, 1–15. [Google Scholar] [CrossRef]
- Geng, W.; Tang, H.; Luo, S.; Lv, Y.; Liang, D.; Kang, X.; Hong, W. Exosomes from mirna-126-modified adscs promotes functional recovery after stroke in rats by improving neurogenesis and suppressing microglia activation. Am. J. Transl. Res. 2019, 11, 780–792. [Google Scholar]
- Jiang, M.; Wang, H.; Jin, M.; Yang, X.; Ji, H.; Jiang, Y.; Zhang, H.; Wu, F.; Wu, G.; Lai, X.; et al. Exosomes from MiR-30d-5p-ADSCs Reverse Acute Ischemic Stroke-Induced, Autophagy-Mediated Brain Injury by Promoting M2 Microglial/Macrophage Polarization. Cell. Physiol. Biochem. 2018, 47, 864–878. [Google Scholar] [CrossRef]
- Yang, Y.; Ye, Y.; Kong, C.; Su, X.; Zhang, X.; Bai, W.; He, X. MiR-124 Enriched Exosomes Promoted the M2 Polarization of Microglia and Enhanced Hippocampus Neurogenesis After Traumatic Brain Injury by Inhibiting TLR4 Pathway. Neurochem. Res. 2019, 44, 811–828. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Yao, X.; Li, H.; Li, X.; Zhang, T.; Sun, Q.; Ji, C.; Chen, G. Role of Exosomes Derived from miR-133b Modified MSCs in an Experimental Rat Model of Intracerebral Hemorrhage. J. Mol. Neurosci. 2018, 64, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Katakowski, M.; Wang, F.; Qian, J.Y.; Liu, X.S.; Ali, M.M.; Buller, B.; Zhang, Z.G.; Chopp, M. MicroRNA cluster miR-17-92 Cluster in Exosomes Enhance Neuroplasticity and Functional Recovery after Stroke in Rats. Stroke 2017, 48, 747–753. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, J.; Wu, J.; Fan, Q.; Zhou, J.; Wu, J.; Liu, S.; Zang, J.; Ye, J.; Xiao, M.; et al. Exosome-mediated targeted delivery of miR-210 for angiogenic therapy after cerebral ischemia in mice. J. Nanobiotechnol. 2019, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, P.; Yao, X.; Li, H.; Shen, H.; Li, X.; Wu, J.; Lu, X. Exosomes Derived From miR-133b-Modified Mesenchymal Stem Cells Promote Recovery After Spinal Cord Injury. Front. Neurosci. 2018, 12, 845. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Jiang, X.; Shi, J.; Gao, S.; Zhu, Y.; Gu, T.; Shi, E. Exosomes derived from bone marrow mesenchymal stem cells overexpressing microRNA-25 protect spinal cords against transient ischemia. J. Thorac. Cardiovasc. Surg. 2019, 157, 508–517. [Google Scholar] [CrossRef]
- Yu, T.; Zhao, C.; Hou, S.; Zhou, W.; Wang, B.; Chen, Y. Exosomes secreted from miRNA-29b-modified mesenchymal stem cells repaired spinal cord injury in rats. Braz. J. Med. Biol. Res. 2019, 52. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Perets, N.; Betzer, O.; Ben-Shaul, S.; Sheinin, A.; Michaelevski, I.; Popovtzer, R.; Offen, D.; Levenberg, S. Intranasal Delivery of Mesenchymal Stem Cell Derived Exosomes Loaded with Phosphatase and Tensin Homolog siRNA Repairs Complete Spinal Cord Injury. ACS Nano 2019, 13, 10015–10028. [Google Scholar] [CrossRef]
- Hosseini Shamili, F.; Alibolandi, M.; Rafatpanah, H.; Abnous, K.; Mahmoudi, M.; Kalantari, M.; Taghdisi, S.M.; Ramezani, M. Immunomodulatory properties of MSC-derived exosomes armed with high affinity aptamer toward mylein as a platform for reducing multiple sclerosis clinical score. J. Control. Release 2019, 299, 149–164. [Google Scholar] [CrossRef]
- Liu, Y.; Fu, N.; Su, J.; Wang, X. Rapid Enkephalin Delivery Using Exosomes to Promote Neurons Recovery in Ischemic Stroke by Inhibiting Neuronal p53/Caspase-3|Semantic Scholar. BioMed Res. Int. 2019. Available online: https://www.semanticscholar.org/paper/Rapid-Enkephalin-Delivery-Using-Exosomes-to-Promote-Liu-Fu/3cc70c482c71952aea95127def334e101b231b75 (accessed on 24 April 2020).
- Huang, X.; Ding, J.; Li, Y.; Liu, W.; Ji, J.; Wang, H.; Wang, X. Exosomes derived from PEDF modified adipose-derived mesenchymal stem cells ameliorate cerebral ischemia-reperfusion injury by regulation of autophagy and apoptosis. Exp. Cell Res. 2018, 371, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Zhang, H.X.; He, C.P.; Fan, S.; Zhu, Y.L.; Qi, C.; Huang, N.P.; Xiao, Z.D.; Lu, Z.H.; Tannous, B.A.; et al. Surface functionalized exosomes as targeted drug delivery vehicles for cerebral ischemia therapy. Biomaterials 2018, 150, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Jia, Z.; Ma, K.; Zhang, J.; Dai, C.; Yao, Z.; Deng, W.; Su, J.; Wang, R.; Chen, X. Protective effect of BMSCs-derived exosomes mediated by BDNF on TBI via miR-216a-5p. Med. Sci. Monit. 2020, 26, e920855. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.H.; Guo, H.D.; Li, H.; Zhai, Y.; Gong, Z.B.; Wu, J.; Liu, J.S.; Dong, Y.R.; Hou, S.X.; Liu, J.R. RVG-modified exosomes derived from mesenchymal stem cells rescue memory deficits by regulating inflammatory responses in a mouse model of Alzheimer’s disease. Immun. Ageing 2019, 16, 10. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Kim, T.J.; Kang, L.; Kim, Y.J.; Kang, M.K.; Kim, J.; Ryu, J.H.; Hyeon, T.; Yoon, B.W.; Ko, S.B.; et al. Mesenchymal stem cell-derived magnetic extracellular nanovesicles for targeting and treatment of ischemic stroke. Biomaterials 2020, 243, 119942. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chopp, M.; Liu, X.S.; Katakowski, M.; Wang, X.; Tian, X.; Wu, D.; Zhang, Z.G. Exosomes Derived from Mesenchymal Stromal Cells Promote Axonal Growth of Cortical Neurons. Mol. Neurobiol. 2017, 54, 2659–2673. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Disease/Disorder | Reference | Animal Model | Cell Source | Dose | Route of Administration | Biological/Medical Improvement | Suggested Mechanism of Action |
|---|---|---|---|---|---|---|---|
| Alzheimer’s | [52] | Ovariectomized albino-rat | Rat BM | 100 μg | Intravenous (IV) | Improved in destructive structural changes in the taste buds and their innervations | Improved synaptophysin-immunoreactivity |
| Alzheimer’s | [45] | APP/PS1 mouse | Mouse BM | 22.4 µg | Intracerebral | Reduced amount of dystrophic neurites in both the cortex and hippocampus | Aβ plaque reduction |
| Alzheimer’s | [49] | Streptozotocin-induced mouse | Mouse BM | 0.5 μg/day for 5 days | Intraventricular | Recovered cognition impairment | Not mentioned |
| Alzheimer’s | [46] | APP/PS1 mouse | Mouse BM | 100 μg | Intracerebroventricular | Improved cognitive behavior, rescued impairment of CA1 synaptic transmission, and long-term potentiation | Suppression of Aβ induced iNOS expression |
| MS | [28] | EAE rat | Rat BM | 100/400 μg | IV | Decreased neural behavioral scores | Reduced demyelination and neuroinflammation |
| Stroke | [32] | Subcortical infarction rat | Rat adipose | 50/100/200 μg | IV | Improved functional outcomes associated with decreased cell death | Restored fiber tract connectivity, increased oligodendrocyte markers, and re-myelination |
| Stroke | [33] | MCAo rat | Rat BM | 120.68 μg | IV | Reduced neurological severity score; improved spatial learning and memory ability | Inhibited the expression of CysLT2R and NMLTC4 treated microglia; modulated the balance between M1 and M2 microglia; decreased pro-inflammatory cytokines secretion; increased anti-inflammatory and neurotrophic factors production |
| Stroke | [53] | Cortical injured monkey | Monkey BM | 4 × 1011 particles/kg | IV | Enhanced recovery of fine motor function | Not mentioned |
| Stroke | [40] | MCAo rat | Adipose (cell source not mentioned) | 3 treatments of 2.0 × 106 particles | IV | Reduced infarct volume; suppressed apoptosis | Improved BBB condition; suppressed inflammation; reduced abnormal high level of miR-21-3p |
| Stroke | [54] | Intracerebral hemorrhage injection rat | Rat BM | 100 µg | IV | Improved spatial learning, motor function, and sensory memory | Promoting endogenous angiogenesis and neurogenesis; increased white matter remodeling |
| Stroke | [70] | tMCAo rat | Rat BM | 30 µg | IV | Improved Neurological function | Promoted neurogenesis and angiogenesis via miR-184 and miR-210, respectively |
| Stroke | [34] | Intracerebral hemorrhage rat | Rat adipose | 100 µg | IV | Improved functional recovery; reduced infarct size | Increased fiber tract and axonal sprouting; enhanced oligodendrocyte formation and remyelination |
| Stroke | [47] | Transient global cerebral ischemia mouse | Mouse BM | 200 µg | Intracerebroventricular | Restored impaired basal synaptic transmission and synaptic plasticity, and improved spatial learning and memory | Inhibited pathogenic expression of COX-2 in the hippocampus |
| Stroke | [35] | Subcortical infarct rat | Rat adipose | 100 µg | IV | Improved functional recovery | Increased axonal sprouting and growth, oligodendrocyte formation, tract connectivity and remyelination |
| Stroke | [36] | MCAo rat | Mini-pig adipose | 100 µg | IV | Reduced brain infarct zone; improved neurological function | Suppressed inflammation; reduced ROS and oxidative stress generation; promoted angiogenesis |
| Stroke | [69] | MCAo rat | Rat BM | 100 µg | IV | Improved neurologic outcome | Enhanced neurite remodeling, neurogenesis, and angiogenesis |
| Neuroinflammation | [44] | LPS-induced rat | Rat BM | 200 µg | IV | Enhanced neuronal survival | Reduced oxidative stress; reduced inflammatory response |
| TBI | [41] | Controlled cortical impact (CCI) mouse | Rat BM | 30 µg | Retro-orbital | Improved functional recovery; reduced cortical lesion volume; attenuated cellular apoptosis | Inhibited early neuroinflammation through modulation of microglia/macrophages polarization |
| TBI | [59] | CCI rat | Rat BM | 100 µg | IV | Cognitive and sensorimotor improvement. | Promotion of endogenous angiogenesis and neurogenesis; and inflammation reduction. |
| SCI | [38] | Spinal cord hemisection rat | Rat BM | 100 µg | IV | Improved functional recovery and attenuated lesion size and apoptosis | Targeted inhibition of the FasL gene by miR-21-5p |
| SCI | [42] | Rat contusive SCI | Rat BM | 1 × 106 cells equivalents | IV | Reduced neuronal cell apoptosis, enhanced neuronal survival and regeneration, and improved motor function | Suppression of pericytes migration; and improved blood-spinal cord barrier integrity via NF-κB p65 signaling |
| SCI | [39] | Rat contusive SCI | Rat BM | 100 µg | IV | Suppressed glial scar formation; attenuated lesion size; promoted axonal regeneration; and improved functional behavioral recovery | Promoted blood vessel formation; reduced neuronal cells apoptosis; suppressed inflammation; and suppressed activation of A1 neurotoxic reactive astrocytes |
| SCI | [30] | Spinal cord hemisection injured rat | Rat BM | 100 µg | IV | Reduced disease severity | Inhibited complement mRNA synthesis and release; inhibited activation of NF-κB signaling by binding to microglia cells. |
| SCI | [48] | Rat contusive SCI | Rat BM | 1 × 106 cells equivalents | IV | Improved locomotor function; and the neuroprotective effect on residual neurons, synapses, and myelin sheath. | Reduced A1 astrocyte proportion by inhibiting NFκB activation; reductions in proinflammatory cytokine levels |
| SCI | [31] | Rat contusive SCI | Rat BM | 100 µg | IV | Attenuated lesion size and improved functional recovery | Attenuated cellular apoptosis and inflammation; promoted angiogenesis |
| Disease/Disorder | Reference | Animal Model | Cell Source | Dose | Root of Administration | Biological/Medical Improvement | Suggested Mechanism of Action |
|---|---|---|---|---|---|---|---|
| Alzheimer’s | [50] | Aβ-inoculated mouse | Human, purchased from ATCC | 10 µg | Intrahippocampal | Enhance neurogenesis and restore cognitive function | Not mentioned |
| Alzheimer’s | [51] | APP/PS1 mouse | Human umbilical cord | 30 µg | IV | Repair cognitive disfunctions | Help to clear Aβ deposition; and modulate the activation of microglia in the brain |
| MS | [29] | EAE mouse | Human BM | 150 μg | IV | Reduced disease severity | Reduced demyelination; decreased neuroinflammation; and upregulated the number of regulatory T cells |
| Stroke | [55] | MCAo rat | Human umbilical cord blood | 150 µg | IV | Attenuated infarct size; exacerbated the somatosensory and motor dysfunction | Not mentioned |
| Stroke | [37] | MCAo mouse | Human BM | Released by 2 × 106 MSCs | IV | Improved neurological impairment and long-term neuroprotection | Promoted neurogenesis and angiogenesis; prevented post-ischemic immunosuppression |
| Perinatal brain injury | [56] | A combination of a hypoxic-ischemic and an inflammatory insult in rat | Human Wharton’s jelly | 50 mg/kg | Intranasal (IN) | Improved long-term neurodevelopmental outcome | Prevented gray and white matter alterations |
| Perinatal brain injury | [43] | Rice-Vannucci mouse | Human BM | 1.25 × 109 particles/dose | IN | Improved short-term behavioral outcomes; reduced tissue volume loss and cell death | Reduced microglial activation |
| Perinatal brain injury | [57] | LPS-induced rat | Human BM | 1 × 108 cell equivalents/kg bodyweight | Intraperitoneal (IP) | Improved long-lasting cognitive functions | Ameliorated inflammation-induced neuronal cellular degeneration; reduced microgliosis; prevented reactive astrogliosis; and restored short-term myelination deficits and long-term microstructural abnormalities of the white matter |
| Perinatal brain injury | [58] | Transient umbilical cord occlusion in preterm ovine fetus | Human BM | Two boluses of 2.0 × 107 cell equivalents | IV | Reduced total number and duration of seizures; and preserved baroreceptor reflex sensitivity | Hypomyelination prevention |
| TBI | [61] | A combination of CCI and hemorrhagic shock swine | Human BM | 1 × 1013 particles | IV | Reduced the severity of neurological injury and improved neurocognitive recovery | Not mentioned |
| TBI | [60] | CCI mouse | Human BM | 30 µg | IV | Rescued pattern separation and spatial learning impairments | Immunomodulation |
| SCI | [62] | Mouse contusive SCI | Human umbilical cord | 20/200 µg | IV | Promoted locomotor functional recovery | Attenuated inflammation of the injury region |
| SCI | [63] | Spinal cord contusion rat | Human BM | 1 × 109 particles | IV | Improved locomotor recovery score; improved mechanical sensitivity | Diminished inflammatory response |
| SE | [64] | Pilocarpine mouse | Human umbilical cord | 30 µg | Intraventricular | Ameliorated learning and memory impairments | Reduced inflammatory responses associated with hippocampal astrocyte activation via Nrf2-NF-κB signaling pathway |
| SE | [65] | Pilocarpine mouse | Human BM | 30 µg | IN | Long-term preservation of normal hippocampal neurogenesis and cognitive and memory function | Diminished loss of glutamatergic and GABAergic neurons; and reduced inflammation in the hippocampus |
| Disease/Disorder | Reference | Animal Model | Cell Source | The Addition | Dose | Route of Administration | Biological/Medical Improvement | Suggested Mechanism of Action |
|---|---|---|---|---|---|---|---|---|
| Alzheimer’s | [92] | APP/PS1 mouse | Mouse BM | Rabies viral glycoprotein (RVG) | 4 boluses of 5 × 1011 particles | IV | Improved learning and memory function | Decreased plaque deposition and Aβ levels; reduced astrocytes activation; reduced pro-inflammatory mediators and raised anti-inflammatory factors |
| MS | [87] | EAE mouse | Mouse BM | LJM-3064 aptamer | 200 μg | IV | Reduced disease severity | Suppressed of inflammatory response; lowered demyelination lesion region |
| Stroke | [77] | MCAo rat | Rat BM | miR-29b-3p | 100 μg/kg/day for 3 days | Intracerebroventricular | Reduced infarct volume | Suppressed neuronal apoptosis and promoted angiogenesis through the downregulation of PTEN and activation of Akt signaling pathway |
| Stroke | [93] | MCAo rat | Human BM | Iron oxide nanoparticles (IONP) | 200 μg | IV | Decreased infarction volume and improved motor function | Promoted the anti-inflammatory response, angiogenesis, and anti-apoptosis |
| Stroke | [78] | MCAo rat | Rat adipose | miR-126 | Not mentioned | IV | Enhanced functional recovery | Inhibited microglial activation and inflammatory response; promoted neurogenesis and vasculogenesis |
| Stroke | [88] | Transient MCAo rat | Rat BM | Transferrin and enkephalin | One or two boluses of 5 × 104 | IV | Improved brain neuron density and neurological score | Decreased levels of LDH, p53, caspase-3, and NO |
| Stroke | [82] | MCAo rat | Mouse BM | c(RGDyK) peptide and miR-210 | 100 µg | IV | Enhanced survival rate | Promoted angiogenesis; up-regulation of integrin β3 and CD34 expression |
| Stroke | [90] | MCAo mice | Mouse BM | c(RGDyK) peptide and curcumin | 100 µg | IV | Reduced cellular apoptosis in the legion region | Suppressed inflammatory response |
| Stroke | [15] | Intracerebral hemorrhage rat | Rat BM | miR-133b | 100 μg | IV | Reduced apoptotic and neurodegenerative neurons | Inhibited RhoA and activation of ERK1/2/CREB pathway |
| Stroke | [89] | MCAo rat | Rat adipose | Pigment epithelium-derived factor (PEDF) | 100 μg/kg/day for 3 days | Lateral cerebral ventricle | Reduced infarct volume; suppressed neuronal apoptosis | Activated autophagy |
| Stroke | [79] | Modified MCAo rat | Rat adipose | miR-30d-5p | 80 μg | IV | Decreased cerebral injury area of infarction | Suppressed autophagy and promoted M2 microglia/macrophage polarization |
| Stroke | [81] | MCAo rat | Rat BM | miR-17-92 | 100 µg | IV | Improved neurological outcome | Increased neural remodeling including neurogenesis, oligodendrogenesis and neurite plasticity; inhibited PTEN, and subsequently increased the phosphorylation of PTEN downstream proteins, Akt, mTOR and GSK-3β |
| TBI | [91] | Electric cortical contusion impactor rat | Rat BM | BDNF | 100 µg | IV | Inhibit apoptosis | Inhibited inflammation and promoted neuronal regeneration; increased miR-216a-5p |
| TBI | [80] | Controlled cortex injury rat | Rat BM | miR-124 | 100 µg | IV | Improved neurological function recovery | Reduced production of pro-inflammatory cytokines; promoted M2 polarization of microglia; increased production of anti-inflammatory cytokines; enhanced neurogenesis in hippocampus |
| SCI | [86] | Complete spinal cord transection rat | Human BM | Phosphatase and tensin homolog (PTEN) siRNA | 5 boluses of 1.62 × 108 particles | IN | Elicited functional recovery; improved structural and electrophysiological function | Enhanced axonal growth and neovascularization; reduced microgliosis and astrogliosis |
| SCI | [84] | Spinal cord ischemia rat | Rat BM | miR-25 | 20 µg | Intrathecal | Improved MDI (motor deficit index); enhanced neuroprotection | Reduced pro-inflammatory cytokines; reduced oxidative stress markers |
| SCI | [85] | Rat contusive SCI | Rat BM | miR-29b | 100 µg | IV | Increased BBB score | Decreased contractile nerve cell numbers and GFAP positive neurons |
| SCI | [83] | Compression SCI rat | Rat BM | miR-133b | 100 μg | IV | Improved recovery of hindlimb locomotor function | Preserved neurons; promoted regeneration of axons; activated ERK1/2, STAT3, and CREB; inhibited RhoA expression |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guy, R.; Offen, D. Promising Opportunities for Treating Neurodegenerative Diseases with Mesenchymal Stem Cell-Derived Exosomes. Biomolecules 2020, 10, 1320. https://doi.org/10.3390/biom10091320
Guy R, Offen D. Promising Opportunities for Treating Neurodegenerative Diseases with Mesenchymal Stem Cell-Derived Exosomes. Biomolecules. 2020; 10(9):1320. https://doi.org/10.3390/biom10091320
Chicago/Turabian StyleGuy, Reut, and Daniel Offen. 2020. "Promising Opportunities for Treating Neurodegenerative Diseases with Mesenchymal Stem Cell-Derived Exosomes" Biomolecules 10, no. 9: 1320. https://doi.org/10.3390/biom10091320
APA StyleGuy, R., & Offen, D. (2020). Promising Opportunities for Treating Neurodegenerative Diseases with Mesenchymal Stem Cell-Derived Exosomes. Biomolecules, 10(9), 1320. https://doi.org/10.3390/biom10091320