
Dietary Nicotinamide Mononucleotide, a Key NAD+ Intermediate, Alleviates Body Fat Mass and Hypertriglyceridemia by Enhancing Energy Expenditure with Promotion of Fat Oxidation and Hepatic Lipolysis and Suppressing Hepatic Lipogenesis in db/db Mice



Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Diets
2.3. Animals
2.4. Respiratory Gas Analysis
2.5. Measurement of Plasma Biochemical Parameters
2.6. Measurement of Hepatic Contents of Lipids, Glycogen, NAD+, and NADH
2.7. Measurement of NAD+ and NADH Contents in Epididymal WAT
2.8. Measurement of Adiponectin Content in Mesenteric WAT
2.9. Measurement of Hepatic Fatty Acid Metabolism-Related Enzyme Activities
2.10. Measurement of mRNA Levels in Liver and Epididymal White Adipose Tissues
2.11. Statistical Analysis
3. Results
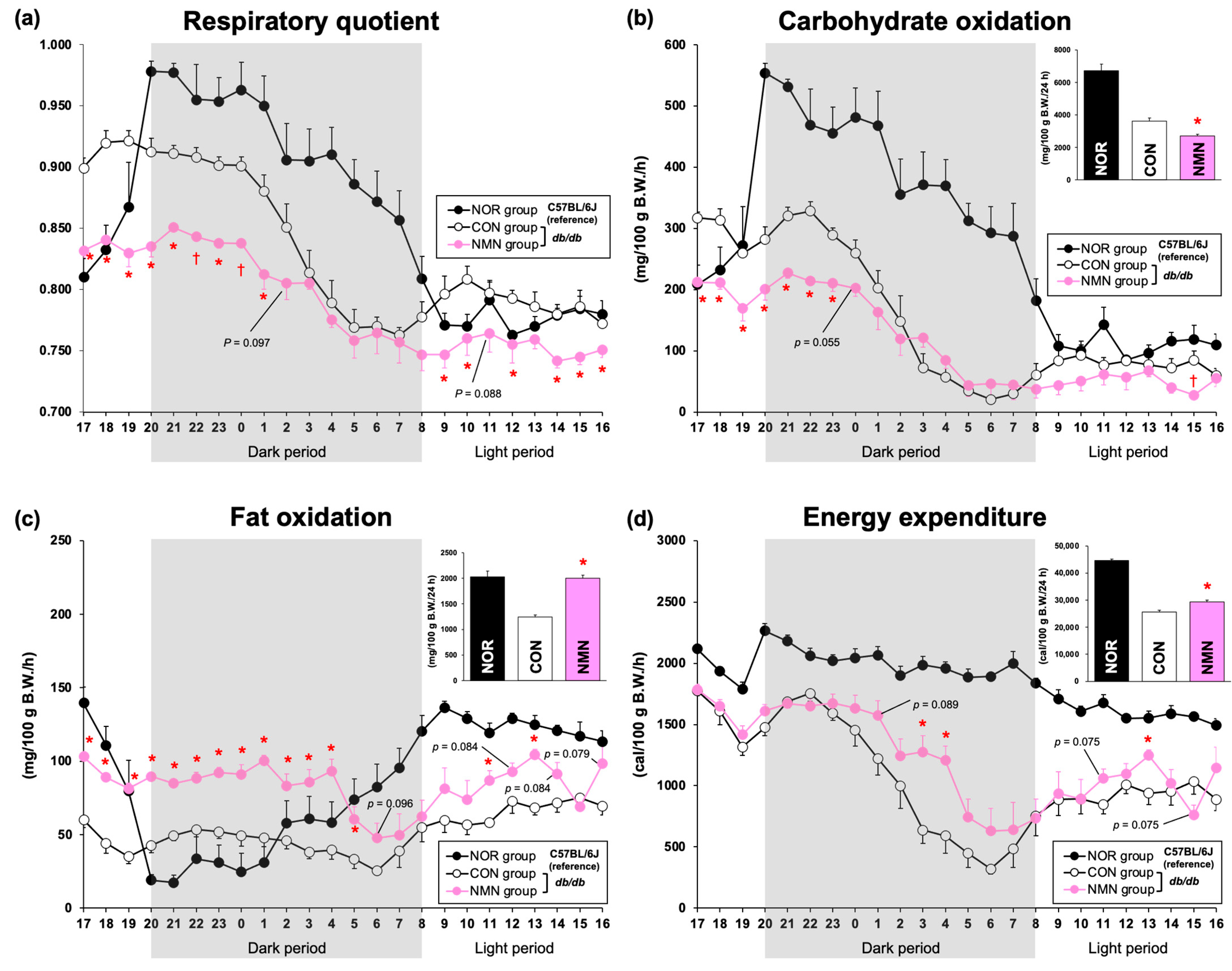
3.1. Effects of Dietary NMN Intake on Nutrients Oxidation in Obese Diabetic db/db Mice
3.2. Effects of Dietary NMN Intake on Morphometric Variables in Obese Diabetic db/db Mice
3.3. Effects of Dietary NMN Intake on Biochemical Parameters in Plasma, the Liver, Epididymal WAT, and Mesenteric WAT of Obese Diabetic db/db Mice
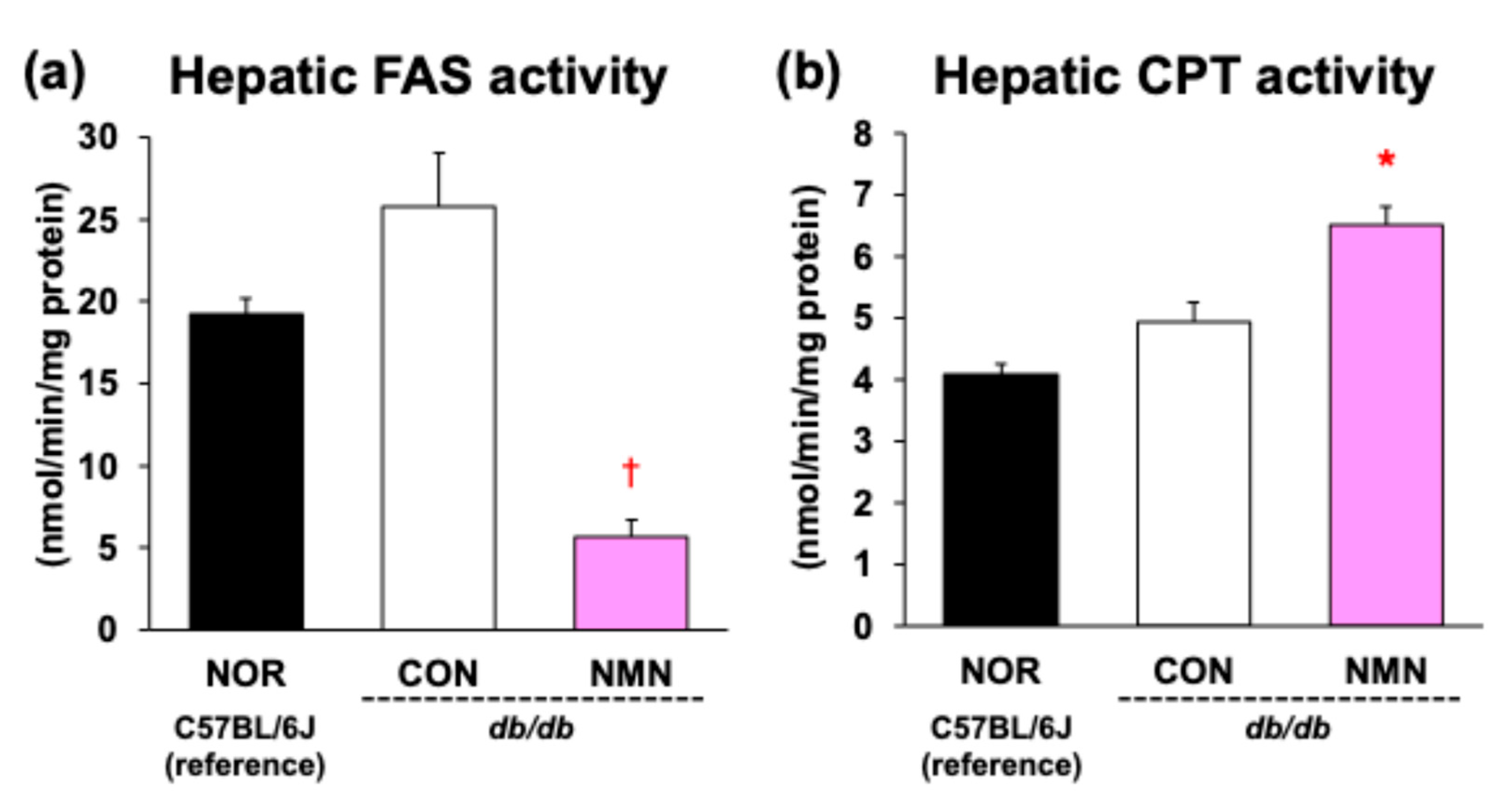
3.4. Effects of Dietary NMN Intake on Activities of Hepatic Enzymes Related to Fatty Acid Metabolism in Obese Diabetic db/db Mice
3.5. Effects of Dietary NMN Intake on mRNA Levels in the Liver and Epididymal WAT of Obese Diabetic db/db Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ALT | Alanine aminotransferase |
| BAT | Brown adipose tissue |
| ChE | Cholinesterase |
| Chol-E | Cholesterol esters |
| CPT | Carnitine palmitoyltransferase |
| FAS | Fatty acid synthase |
| F-Chol | Free cholesterol |
| HDL | High-density lipoprotein |
| NAD+ | Nicotinamide adenine dinucleotide |
| NADH | Nicotinamide adenine dinucleotide hydride |
| NMN | Nicotinamide mononucleotide |
| NOAEL | No-observable adverse effect level |
| PL | Phospholipids |
| RQ | Respiratory quotient |
| TG | Triglycerides |
| T-Chol | Total cholesterol |
| WAT | White adipose tissue |
References
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2005, 365, 1415–1428. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 14 January 2025).
- Nagao, K.; Wang, Y.M.; Inoue, N.; Han, S.Y.; Buang, Y.; Noda, T.; Kouda, N.; Okamatsu, H.; Yanagita, T. The 10trans, 12cis isomer of conjugated linoleic acid promotes energy metabolism in OLETF rats. Nutrition 2003, 19, 652–656. [Google Scholar] [CrossRef]
- Ikeda, I.; Konno, R.; Shimizu, T.; Ide, T.; Takahashi, N.; Kawada, T.; Nagao, K.; Inoue, N.; Yanagita, T.; Hamada, T.; et al. Campest-5-en-3-one, an oxidized derivative of campesterol, activates PPARalpha, promotes energy consumption and reduces visceral fat deposition in rats. Biochim. Biophys. Acta 2006, 1760, 800–807. [Google Scholar] [CrossRef]
- Inoue, N.; Matsunaga, Y.; Satoh, H.; Takahashi, M. Enhanced energy expenditure and fat oxidation in humans with high BMI scores by the ingestion of novel and non-pungent capsaicin analogues (capsinoids). Biosci. Biotechnol. Biochem. 2007, 71, 380–389. [Google Scholar] [CrossRef]
- Kim, M.; Goto, T.; Yu, R.; Uchida, K.; Tominaga, M.; Kano, Y.; Takahashi, N.; Kawada, T. Fish oil intake induces UCP1 upregulation in brown and white adipose tissue via the sympathetic nervous system. Sci. Rep. 2015, 5, 18013. [Google Scholar] [CrossRef]
- Nagao, K.; Jinnouchi, T.; Kai, S.; Yanagita, T. Pterostilbene, a dimethylated analog of resveratrol, promotes energy metabolism in obese rats. J. Nutr. Biochem. 2017, 43, 151–155. [Google Scholar] [CrossRef]
- Shirouchi, B.; Nagao, K.; Umegatani, M.; Shiraishi, A.; Morita, Y.; Kai, S.; Yanagita, T.; Ogawa, A.; Kadooka, Y.; Sato, M. Probiotic Lactobacillus gasseri SBT2055 improves glucose tolerance and reduces body weight gain in rats by stimulating energy expenditure. Br. J. Nutr. 2016, 116, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Xie, N.; Zhang, L.; Gao, W.; Huang, C.; Huber, P.E.; Zhou, X.; Li, C.; Shen, G.; Zou, B. NAD+ metabolism: Pathophysiologic mechanisms and therapeutic potential. Signal Transduct. Target Ther. 2020, 5, 227. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, J.; Mills, K.F.; Yoon, M.J.; Imai, S. Nicotinamide mononucleotide, a key NAD+ intermediate, treats the pathophysiology of diet- and age-induced diabetes in mice. Cell Metab. 2011, 14, 528–536. [Google Scholar] [CrossRef]
- Massudi, H.; Grant, R.; Braidy, N.; Guest, J.; Farnsworth, B.; Guillemin, G.J. Age-associated changes in oxidative stress and NAD+ metabolism in human tissue. PLoS ONE 2012, 7, e42357. [Google Scholar] [CrossRef]
- Gomes, A.P.; Price, N.L.; Ling, A.J.; Moslehi, J.J.; Montgomery, M.K.; Rajman, L.; White, J.P.; Teodoro, J.S.; Wrann, C.D.; Hubbard, B.P.; et al. Declining NAD+ induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging. Cell 2013, 155, 1624–1638. [Google Scholar] [CrossRef] [PubMed]
- Mouchiroud, L.; Houtkooper, R.H.; Moullan, N.; Katsyuba, E.; Ryu, D.; Cantó, C.; Mottis, A.; Jo, Y.S.; Viswanathan, M.; Schoonjans, K.; et al. The NAD+/Sirtuin Pathway Modulates Longevity through Activation of Mitochondrial UPR and FOXO Signaling. Cell 2013, 154, 430–441. [Google Scholar] [CrossRef]
- Stein, L.R.; Imai, S. Specific ablation of Nampt in adult neural stem cells recapitulates their functional defects during aging. EMBO J. 2014, 33, 1321–1340. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.H.; Lu, M.; Lee, B.Y.; Ugurbil, K.; Chen, W. In vivo NAD assay reveals the intracellular NAD contents and redox state in healthy human brain and their age dependences. Proc. Natl. Acad. Sci. USA 2015, 112, 2876–2881. [Google Scholar] [CrossRef] [PubMed]
- Uddin, G.M.; Youngson, N.A.; Doyle, B.M.; Sinclair, D.A.; Morris, M.J. Nicotinamide mononucleotide (NMN) supplementation ameliorates the impact of maternal obesity in mice: Comparison with exercise. Sci. Rep. 2017, 7, 15063. [Google Scholar] [CrossRef]
- Caton, P.W.; Kieswich, J.; Yaqoob, M.M.; Holness, M.J.; Sugden, M.C. Nicotinamide mononucleotide protects against pro-inflammatory cytokine-mediated impairment of mouse islet function. Diabetologia 2011, 54, 3083–3092. [Google Scholar] [CrossRef]
- Tarantini, S.; Valcarcel-Ares, M.N.; Toth, P.; Yabluchanskiy, A.; Tucsek, Z.; Kiss, T.; Hertelendy, P.; Kinter, M.; Ballabh, P.; Süle, Z.; et al. Nicotinamide mononucleotide (NMN) supplementation rescues cerebromicrovascular endothelial function and neurovascular coupling responses and improves cognitive function in aged mice. Redox Biol. 2019, 24, 101192. [Google Scholar] [CrossRef]
- Wang, X.; Hu, X.; Yang, Y.; Takata, T.; Sakurai, T. Nicotinamide mononucleotide protects against β-amyloid oligomer-induced cognitive impairment and neuronal death. Brain Res. 2016, 1643, 1–9. [Google Scholar] [CrossRef]
- Mills, K.F.; Yoshida, S.; Stein, L.R.; Grozio, A.; Kubota, S.; Sasaki, Y.; Redpath, P.; Migaud, M.E.; Apte, R.S.; Uchida, K.; et al. Long-Term Administration of Nicotinamide Mononucleotide Mitigates Age-Associated Physiological Decline in Mice. Cell Metab. 2016, 24, 795–806. [Google Scholar] [CrossRef]
- Hong, W.; Mo, F.; Zhang, Z.; Huang, M.; Wei, X. Nicotinamide Mononucleotide: A Promising Molecule for Therapy of Diverse Diseases by Targeting NAD+ Metabolism. Front. Cell Dev. Biol. 2020, 8, 246. [Google Scholar] [CrossRef]
- American Institute of Nutrition. Report of the American Institute of Nutrition ad hoc Committee on Standards for Nutritional Studies. J. Nutr. 1977, 107, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Cros, C.; Cannelle, H.; Laganier, L.; Grozio, A.; Canault, M. Safety evaluation after acute and sub-chronic oral administration of high purity nicotinamide mononucleotide (NMN-C®) in Sprague-Dawley rats. Food Chem. Toxicol. 2021, 150, 112060. [Google Scholar] [CrossRef]
- Chen, H.; Charlat, O.; Tartaglia, L.A.; Woolf, E.A.; Weng, X.; Ellis, S.J.; Lakey, N.D.; Culpepper, J.; Moore, K.J.; Breitbart, R.E.; et al. Evidence that the diabetes gene encodes the leptin receptor: Identification of a mutation in the leptin receptor gene in db/db mice. Cell 1996, 84, 491–495. [Google Scholar] [CrossRef]
- Lee, G.H.; Proenca, R.; Montez, J.M.; Carroll, K.M.; Darvishzadeh, J.G.; Lee, J.I.; Friedman, J.M. Abnormal splicing of the leptin receptor in diabetic mice. Nature 1996, 379, 632–635. [Google Scholar] [CrossRef]
- Hariri, N.; Thibault, L. High-fat diet-induced obesity in animal models. Nutr. Res. Rev. 2010, 23, 270–299. [Google Scholar] [CrossRef]
- Higuchi, M.; Okumura, M.; Mitsuta, S.; Shirouchi, B. Dietary Cholest-4-en-3-one, a Cholesterol Metabolite of Gut Microbiota, Alleviates Hyperlipidemia, Hepatic Cholesterol Accumulation, and Hyperinsulinemia in Obese, Diabetic db/db Mice. Metabolites 2024, 14, 321. [Google Scholar] [CrossRef] [PubMed]
- Niibo, M.; Shirouchi, B.; Umegatani, M.; Morita, Y.; Ogawa, A.; Sakai, F.; Kadooka, Y.; Sato, M. Probiotic Lactobacillus gasseri SBT2055 improves insulin secretion in a diabetic rat model. J. Dairy Sci. 2019, 102, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Shirouchi, B.; Kawahara, Y.; Kutsuna, Y.; Higuchi, M.; Okumura, M.; Mitsuta, S.; Nagao, N.; Tanaka, K. Oral Administration of Chaetoceros gracilis-A Marine Microalga-Alleviates Hepatic Lipid Accumulation in Rats Fed a High-Sucrose and Cholesterol-Containing Diet. Metabolites 2023, 13, 436. [Google Scholar] [CrossRef]
- Pfaffl, M.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Margier, M.; Kuehnemann, C.; Hulo, N.; Morales, J.; Ashok Kumaar, P.V.; Cros, C.; Cannelle, H.; Charmetant, J.; Verdin, E.; Canault, M.; et al. Nicotinamide Mononucleotide Administration Prevents Doxorubicin-Induced Cardiotoxicity and Loss in Physical Activity in Mice. Cells 2022, 12, 108. [Google Scholar] [CrossRef]
- Bai, L.B.; Yau, L.F.; Tong, T.T.; Chan, W.H.; Zhang, W.; Jiang, Z.H. Improvement of tissue-specific distribution and biotransformation potential of nicotinamide mononucleotide in combination with ginsenosides or resveratrol. Pharmacol. Res. Perspect. 2022, 10, e00986. [Google Scholar] [CrossRef]
- Niu, K.M.; Bao, T.; Gao, L.; Ru, M.; Li, Y.; Jiang, L.; Ye, C.; Wang, S.; Wu, X. The Impacts of Short-Term NMN Supplementation on Serum Metabolism, Fecal Microbiota, and Telomere Length in Pre-Aging Phase. Front. Nutr. 2021, 8, 756243. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Franczyk, M.P.; Chondronikola, M.; Qi, N.; Gunawardana, S.C.; Stromsdorfer, K.L.; Porter, L.C.; Wozniak, D.F.; Sasaki, Y.; Rensing, N.; et al. Adipose tissue NAD+ biosynthesis is required for regulating adaptive thermogenesis and whole-body energy homeostasis in mice. Proc. Natl. Acad. Sci. USA 2019, 116, 23822–23828. [Google Scholar] [CrossRef]
- Sahebkar, A.; Chew, G.T.; Watts, G.F. Recent advances in pharmacotherapy for hypertriglyceridemia. Prog. Lipid Res. 2014, 56, 47–66. [Google Scholar] [CrossRef]
- Matsuzawa, Y. The metabolic syndrome and adipocytokines. FEBS Lett. 2006, 580, 2917–2921. [Google Scholar] [CrossRef] [PubMed]
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.M.; Bragazzi, N.L. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients 2021, 13, 1180. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Kita, S.; Miyashita, K.; Iioka, M.; Murai, J.; Nakamura, T.; Nishizawa, H.; Fujishima, Y.; Morinaga, J.; Oike, Y.; et al. Identification and Clinical Associations of 3 Forms of Circulating T-cadherin in Human Serum. J. Clin. Endocrinol. Metab. 2021, 106, 1333–1344. [Google Scholar] [CrossRef]
- Kita, S.; Fukuda, S.; Maeda, N.; Shimomura, I. Native adiponectin in serum binds to mammalian cells expressing T-cadherin, but not AdipoRs or calreticulin. eLife 2019, 8, e48675. [Google Scholar] [CrossRef]
- You, Y.; Gao, Y.; Wang, H.; Li, J.; Zhang, X.; Zhu, Z.; Liu, N. Subacute Toxicity Study of Nicotinamide Mononucleotide via Oral Administration. Front. Pharmacol. 2020, 11, 604404. [Google Scholar] [CrossRef]
- Kreitzman, S.N.; Pedersen, M.; Budell, W.; Nichols, D.; Krissman, P.; Clements, M. Safety and effectiveness of weight reduction using a very-low-calorie formulated food. Arch. Intern. Med. 1984, 144, 747–750. [Google Scholar] [CrossRef] [PubMed]
- Friis, R.; Vaziri, N.D.; Akbarpour, F.; Afrasiabi, A. Effect of rapid weight loss with supplemented fasting on liver tests. J. Clin. Gastroenterol. 1987, 9, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Hoy, M.K.; Heshka, S.; Allison, D.B.; Grasset, E.; Blank, R.; Abiri, M.; Heymsfield, S.B. Reduced risk of liver-function-test abnormalities and new gallstone formation with weight loss on 3350-kJ (800-kcal) formula diets. Am. J. Clin. Nutr. 1994, 60, 249–254. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C57BL/6J | db/db | ||
|---|---|---|---|
| NOR Group | CON Group | NMN Group | |
| Initial B.W. (g) | 19.3 ± 0.3 | 27.5 ± 0.5 | 27.6 ± 0.6 |
| Final B.W. (g) | 24.0 ± 0.6 | 37.4 ± 0.9 | 29.7 ± 0.3 † |
| B.W. gain (g) | 4.73 ± 0.44 | 9.88 ± 0.80 | 2.14 ± 0.52 * |
| Food intake (g/day) | 2.88 ± 0.05 | 5.41 ± 0.02 | 5.36 ± 0.14 |
| NMN intake (mg/kg/day) | --- | --- | 902 ± 26 |
| Food efficiency (mg B.W. gain/g food intake) | |||
| 58.5 ± 4.5 | 65.2 ± 5.2 | 14.3 ± 3.3 * | |
| Water intake (mL/day) | 3.88 ± 0.13 | 16.5 ± 0.8 | 23.9 ± 1.9 * |
| Naso–anal length (cm) | 9.07 ± 0.15 | 9.03 ± 0.06 | 8.69 ± 0.05 * |
| Lee index | 318 ± 4 | 370 ± 2 | 356 ± 1 * |
| Organ weight (g) | |||
| Liver | 0.985 ± 0.026 | 2.44 ± 0.13 | 1.86 ± 0.07 * |
| Spleen | 0.0703 ± 0.0018 | 0.0331 ± 0.0013 | 0.0316 ± 0.0020 |
| Testis | 0.188 ± 0.006 | 0.141 ± 0.011 | 0.131 ± 0.011 |
| Kidneys | 0.328 ± 0.006 | 0.436 ± 0.009 | 0.453 ± 0.014 |
| Quadriceps femoris | 0.402 ± 0.029 | 0.185 ± 0.009 | 0.164 ± 0.009 |
| WAT weight (g) | |||
| Epididymal | 0.363 ± 0.027 | 1.63 ± 0.12 | 0.959 ± 0.064 * |
| Perirenal | 0.154 ± 0.024 | 0.725 ± 0.064 | 0.608 ± 0.027 |
| Mesenteric | 0.233 ± 0.017 | 1.32 ± 0.05 | 0.946 ± 0.019 * |
| Abdominal # | 0.750 ± 0.067 | 3.67 ± 0.17 | 2.51 ± 0.06 * |
| Subcutaneous | 0.550 ± 0.069 | 4.09 ± 0.12 | 2.50 ± 0.06 * |
| BAT weight (g) | 0.139 ± 0.010 | 0.309 ± 0.030 | 0.283 ± 0.028 |
| Feces weight (g/day) | 0.271 ± 0.010 | 0.519 ± 0.011 | 0.449 ± 0.040 |
| Rectal temperature (°C) | 36.6 ± 0.1 | 34.4 ± 0.1 | 35.9 ± 0.2 † |
| C57BL/6J | db/db | ||
|---|---|---|---|
| NOR Group | CON Group | NMN Group | |
| Plasma biochemical parameters | |||
| TG (mg/dL) | 74.1 ± 6.5 | 143 ± 15 | 67.7 ± 13.0 * |
| HDL TG (mg/dL) | 43.3 ± 3.0 | 69.9 ± 6.0 | 46.2 ± 5.7 * |
| Non-HDL TG (mg/dL) | 30.7 ± 4.9 | 72.8 ± 10.9 | 21.5 ± 7.9 * |
| T-Chol (mg/dL) | 133 ± 6 | 221 ± 9 | 262 ± 22 |
| HDL Chol (mg/dL) | 118 ± 5 | 202 ± 11 | 241 ± 19 (p = 0.096) |
| Non-HDL Chol (mg/dL) | 14.8 ± 3.6 | 19.3 ± 4.0 | 20.3 ± 7.6 |
| PL (mg/dL) | 243 ± 9 | 329 ± 16 | 362 ± 21 |
| Glucose (mg/dL) | 285 ± 33 | 611 ± 33 | 558 ± 40 |
| Insulin (ng/mL) | 0.0188 ± 0.0057 | 6.35 ± 1.43 | 5.90 ± 0.86 |
| Leptin (ng/mL) | 1.69 ± 0.39 | 61.5 ± 2.9 | 56.1 ± 1.9 |
| Adiponectin (µg/mL) | 26.8 ± 0.6 | 10.7 ± 0.4 | 19.6 ± 2.1 † |
| T-Cadherin (ng/mL) | |||
| 100 kDa + 130 kDa | 192 ± 9 | 144 ± 5 | 199 ± 13 * |
| 100 kDa | 166 ± 8 | 122 ± 4 | 165 ± 12 † |
| 130 kDa | 26.0 ± 2.5 | 21.3 ± 1.0 | 34.0 ± 1.5 * |
| ALT (IU/L) | 5.46 ± 0.36 | 24.4 ± 1.8 | 43.5 ± 2.8 * |
| ChE (IU/L) | 12.9 ± 1.0 | 38.7 ± 1.8 | 55.7 ± 2.4 * |
| Albumin (g/dL) | 3.14 ± 0.05 | 4.09 ± 0.06 | 2.99 ± 0.05 * |
| Hepatic biochemical parameters | |||
| TG (mg/liver) T-Chol (mg/liver) | 27.8 ± 3.2 | 408 ± 65 | 269 ± 24 (p = 0.097) |
| 6.43 ± 0.69 | 42.0 ± 3.1 | 38.4 ± 0.9 | |
| F-Chol (mg/liver) | 3.24 ± 0.16 | 9.98 ± 0.44 | 9.15 ± 0.37 |
| Chol-E (mg/liver) | 3.18 ± 0.59 | 32.0 ± 2.7 | 29.2 ± 1.1 |
| PL (mg/liver) | 29.4 ± 0.9 | 59.5 ± 2.3 | 53.5 ± 1.8 (p = 0.082) |
| Glycogen (mg/liver) | 3.26 ± 1.36 | 42.2 ± 7.6 | 20.1 ± 7.7 (p = 0.074) |
| NAD+ (nmol/liver) | 11.0 ± 1.8 | 21.5 ± 1.7 | 227 ± 48 † |
| NADH (nmol/liver) | 15.7 ± 1.2 | 33.4 ± 4.3 | 660 ± 145 † |
| Epididymal WAT | |||
| NAD+ (nmol/Epi WAT) | 4.02 ± 0.54 | 15.8 ± 2.4 | 7.50 ± 2.11 * |
| NADH (nmol/Epi WAT) | 0.287 ± 0.015 | 0.748 ± 0.051 | 0.510 ± 0.043 * |
| Mesenteric WAT | |||
| Adiponectin | 59.2 ± 4.6 | 61.3 ± 10.5 | 70.2 ± 10.8 |
| (mg/g protein) | |||
| C57BL/6J | db/db | ||
|---|---|---|---|
| NOR Group | CON Group | NMN Group | |
| Liver | (Arbitrary unit) | ||
| Genes related to fatty acid synthesis | |||
| Acaca | 56.6 ± 6.6 | 100 ± 9 | 53.5 ± 4.0 * |
| Fasn | 30.8 ± 4.1 | 100 ± 26 | 35.3 ± 8.9 (p = 0.055) |
| Srebf1 | 105 ± 16 | 100 ± 11 | 143 ± 17 (p = 0.060) |
| Nr1h3 | 110 ± 18 | 100 ± 7 | 87.6 ± 9.0 |
| Genes related to fatty acidβ-oxidation | |||
| Cpt1a | 117 ± 18 | 100 ± 7 | 87.6 ± 7.0 |
| Cpt2 | 102 ± 11 | 100 ± 6 | 123 ± 8 * |
| Genes related to NAD+ metabolism | |||
| Nmnat1 | 89.3 ± 10.2 | 100 ± 7 | 86.7 ± 8.7 |
| Sirt1 | 143 ± 8 | 100 ± 4 | 104 ± 6 |
| Genes related to adiponectin signaling | |||
| Adipor1 | 93.2 ± 10.9 | 100 ± 7 | 88.4 ± 6.7 |
| Adipor2 | 160 ± 10 | 100 ± 8 | 63.6 ± 3.3 * |
| Epididymal WAT | |||
| Gene related to fatty acid synthesis | |||
| Fasn | 91.5 ± 25.4 | 100 ± 27 | 66.9 ± 20.1 |
| Gene related to adipocytokine | |||
| Adipoq | 133 ± 13 | 100 ± 15 | 99.3 ± 21.4 |
| Genes related to thermogenesis | |||
| Ucp2 | 36.8 ± 6.4 | 100 ± 18 | 68.0 ± 9.0 |
| Adrb3 | 425 ± 92 | 100 ± 27 | 78.6 ± 14.7 |
| Prdm16 | 262 ± 52 | 100 ± 15 | 113 ± 42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shirouchi, B.; Mitsuta, S.; Higuchi, M.; Okumura, M.; Tanaka, K. Dietary Nicotinamide Mononucleotide, a Key NAD+ Intermediate, Alleviates Body Fat Mass and Hypertriglyceridemia by Enhancing Energy Expenditure with Promotion of Fat Oxidation and Hepatic Lipolysis and Suppressing Hepatic Lipogenesis in db/db Mice. Metabolites 2025, 15, 333. https://doi.org/10.3390/metabo15050333
Shirouchi B, Mitsuta S, Higuchi M, Okumura M, Tanaka K. Dietary Nicotinamide Mononucleotide, a Key NAD+ Intermediate, Alleviates Body Fat Mass and Hypertriglyceridemia by Enhancing Energy Expenditure with Promotion of Fat Oxidation and Hepatic Lipolysis and Suppressing Hepatic Lipogenesis in db/db Mice. Metabolites. 2025; 15(5):333. https://doi.org/10.3390/metabo15050333
Chicago/Turabian StyleShirouchi, Bungo, Sarasa Mitsuta, Mina Higuchi, Mai Okumura, and Kazunari Tanaka. 2025. "Dietary Nicotinamide Mononucleotide, a Key NAD+ Intermediate, Alleviates Body Fat Mass and Hypertriglyceridemia by Enhancing Energy Expenditure with Promotion of Fat Oxidation and Hepatic Lipolysis and Suppressing Hepatic Lipogenesis in db/db Mice" Metabolites 15, no. 5: 333. https://doi.org/10.3390/metabo15050333
APA StyleShirouchi, B., Mitsuta, S., Higuchi, M., Okumura, M., & Tanaka, K. (2025). Dietary Nicotinamide Mononucleotide, a Key NAD+ Intermediate, Alleviates Body Fat Mass and Hypertriglyceridemia by Enhancing Energy Expenditure with Promotion of Fat Oxidation and Hepatic Lipolysis and Suppressing Hepatic Lipogenesis in db/db Mice. Metabolites, 15(5), 333. https://doi.org/10.3390/metabo15050333