

Comparative Analysis of the Chemical Constituents of Chrysanthemum morifolium with Different Drying Processes Integrating LC/GC–MS−Based, Non-Targeted Metabolomics

Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Plant Materials
2.2. Sample Preparation
2.3. Metabolomics Analysis Based on LC–MS
2.4. Metabolomics Analysis Based on GC–MS
2.5. LC/GC–MS Data Analysis
2.6. High−Performance Liquid Chromatography
3. Results
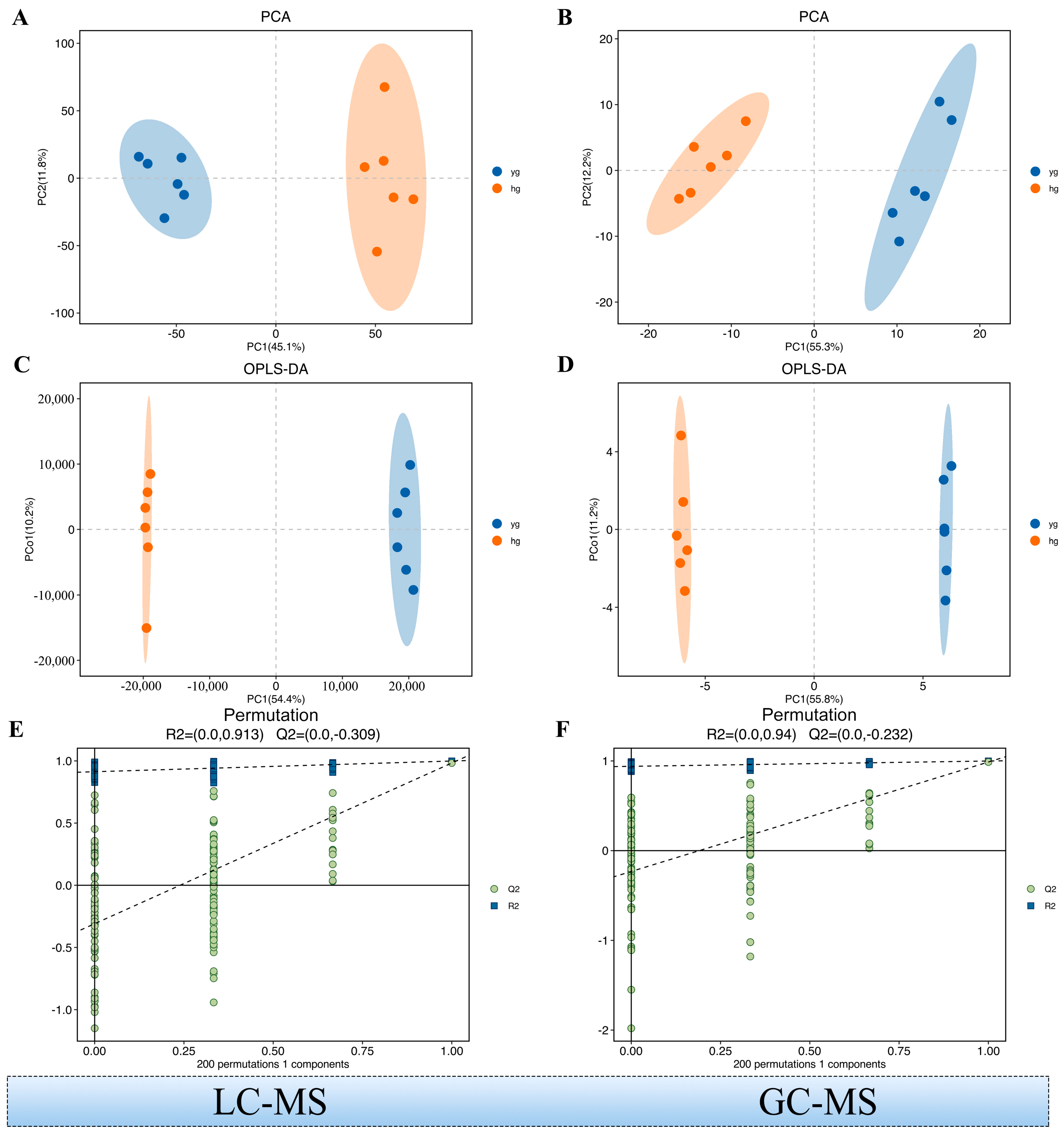
3.1. Identification of the Global Metabolite Profile Based on GC–MS and LC–MS Metabolomics
3.2. Multivariate Analysis of the Identified Metabolites
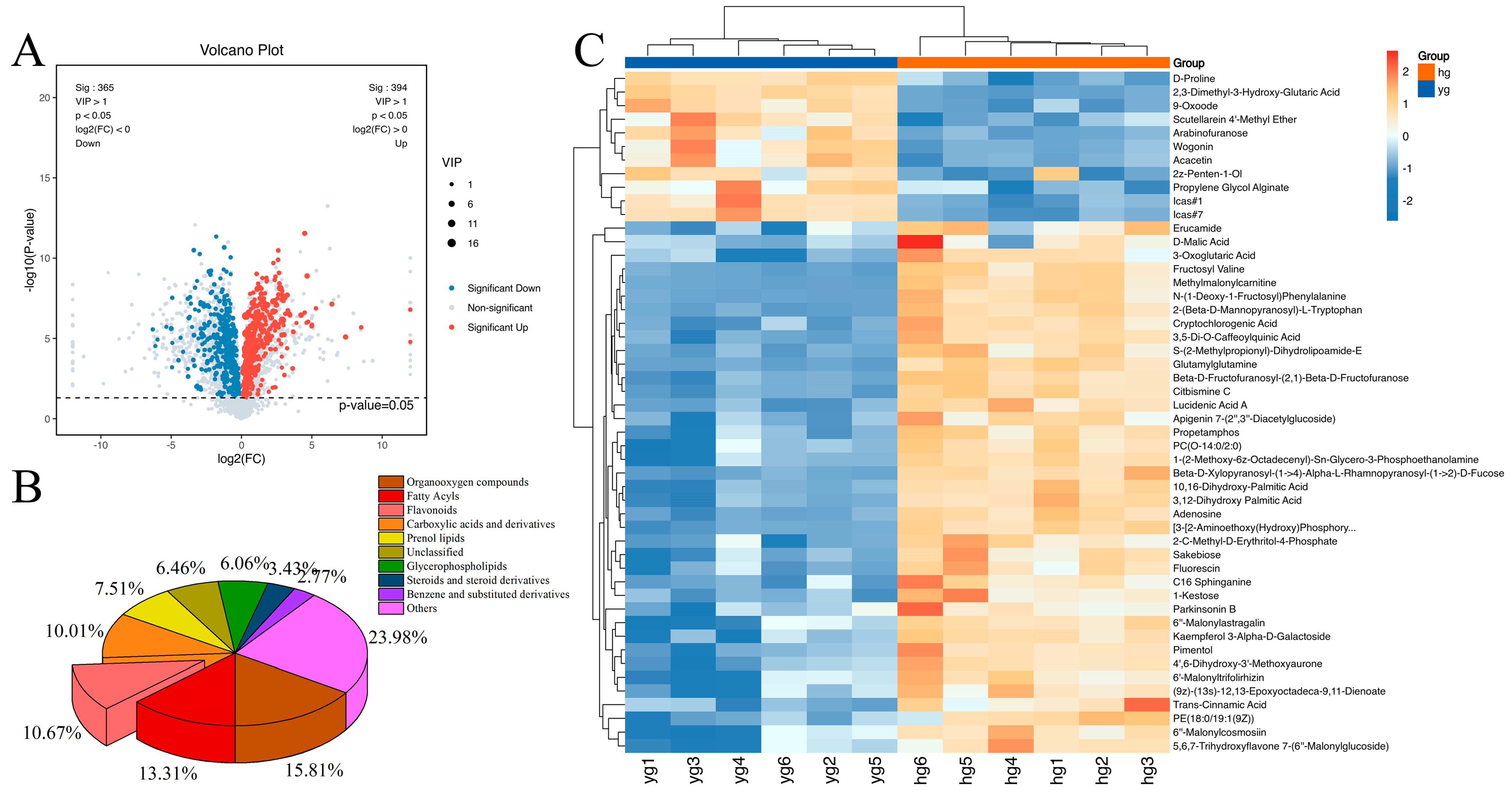
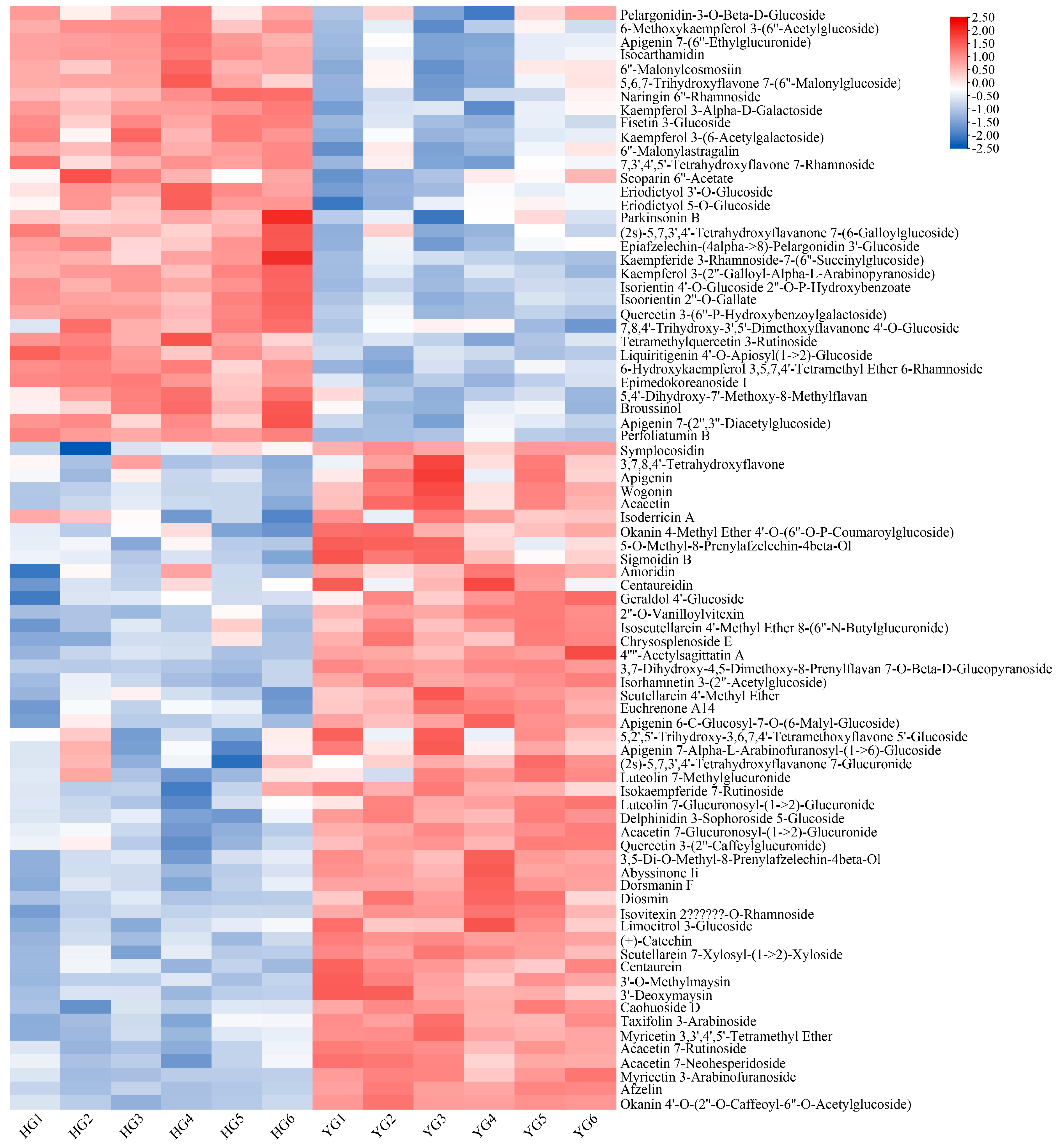
3.3. Screening and Classification of the Differential Metabolites
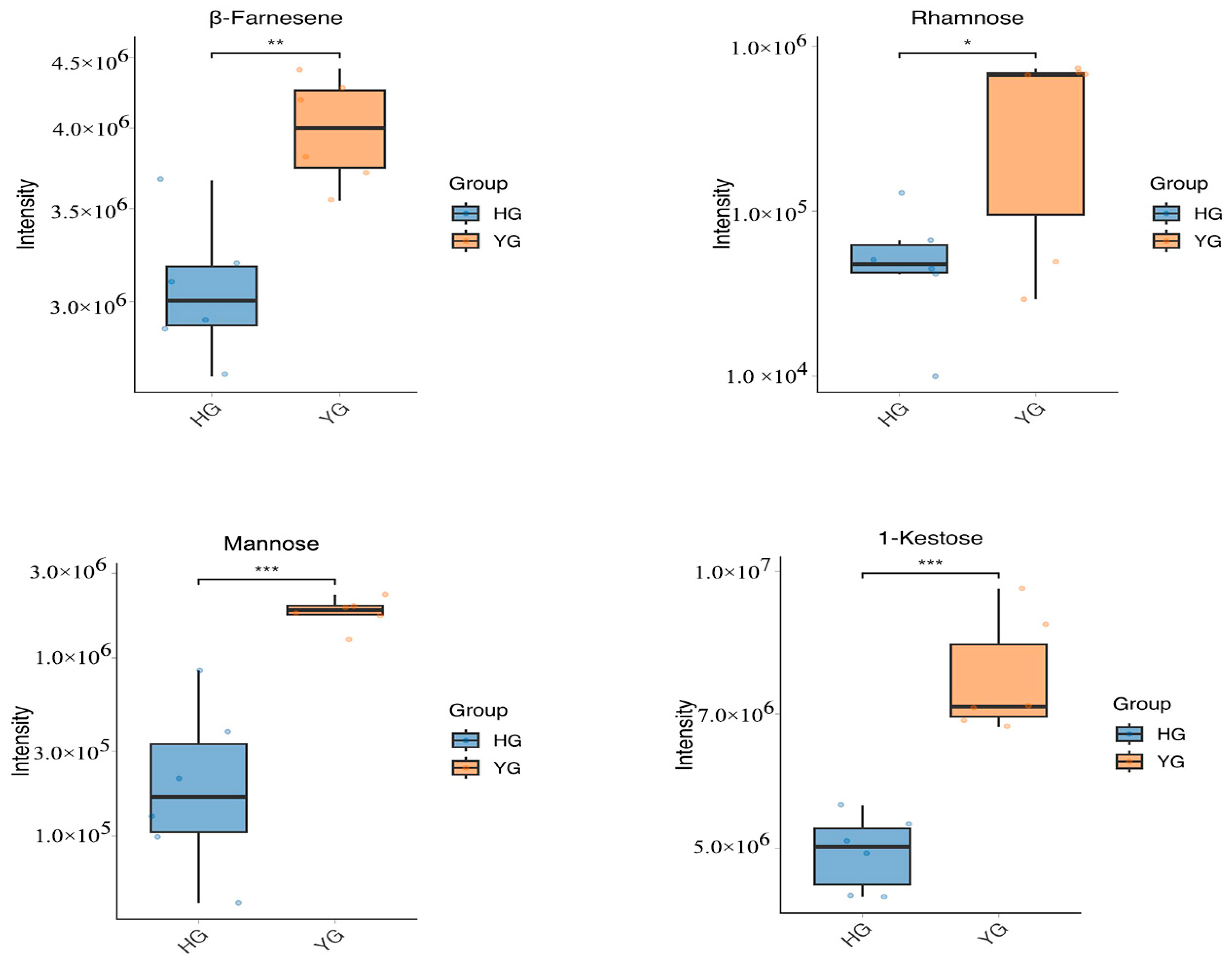
3.4. Determination of YG and HG Indicator Active Ingredients by HPLC
3.5. KEGG Classification and Enrichment Analysis of the Differential Metabolites
4. Discussion
4.1. The YG Samples Had a Higher Flavonoid Content than the HG Samples, Which May Endow C. morifolium with Better Anti-Oxidant Activity
4.2. YG Treatment Resulted in Higher Levels of Phenolic Acids, Which May Endow C. morifolium With Better Anti-Oxidant Activity
4.3. Other Active Compounds That Were Higher in the YG Group than in the HG Group
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- National Pharmacopoeia Committee. Pharmacopoeia of the People’s Republic of China; China Medical Science and Technology Press: Beijing, China, 2020; pp. 323–324. [Google Scholar]
- Shao, Y.; Sun, Y.; Li, D.; Chen, Y. Chrysanthemum indicum L.: A Comprehensive Review of its Botany, Phytochemistry and Pharmacology. Am. J. Chin. Med. 2020, 48, 871–897. [Google Scholar] [CrossRef] [PubMed]
- Hu, C. Chrysanthemum Morifolium Ramat (Juhua, Florists Chrysanthemum); Springer: Vienna, Austria, 2015. [Google Scholar]
- Dong, M.; Yu, D.; Duraipandiyan, V.; Abdullah Al-Dhabi, N. The Protective Effect of Chrysanthemum indicum Extract against Ankylosing Spondylitis in Mouse Models. BioMed Res. Int. 2017, 8206281. [Google Scholar] [CrossRef]
- Cheon, M.S.; Yoon, T.; Choi, G.; Moon, B.C.; Lee, A.Y.; Choo, B.K.; Kim, H.K. Chrysanthemum indicum Linné extract inhibits the inflammatory response by suppressing NF-κB and MAPKs activation in lipopolysaccharide-induced RAW 264.7 macrophages. J. Ethnopharmacol. 2009, 122, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Arokiyaraj, S.; Arasu, M.V.; Vincent, S.; Prakash, N.U.; Choi, S.H.; Oh, Y.K.; Choi, K.C.; Kim, K.H. Rapid green synthesis of silver nanoparticles from Chrysanthemum indicum L and its antibacterial and cytotoxic effects: An in vitro study. Int. J. Nanomed. 2014, 9, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.J.; Li, Y.; Su, S.L.; Wei, D.D.; Yan, H.; Guo, S.; Shang, E.X.; Sun, X.D.; Duan, J.A. Comparative Analysis of Chemical Composition and Antibacterial and Anti-Inflammatory Activities of the Essential Oils from Chrysanthemum morifolium of Different Flowering Stages and Different Parts. Evid Based Complement Altern. Med. 2022, 6, 5954963. [Google Scholar]
- Yang, M.; Sun, S.; Jia, X.; Wen, X.; Tian, X.; Niu, Y.; Wei, J.; Jin, Y.; Du, Y. Study on mechanism of hepatoprotective effect of Chrysanthemum morifolium Ramat. based on metabolomics with network analysis and network pharmacology. J Chromatogr. B Anal. Technol. Biomed. Life Sci. 2023, 1, 123711. [Google Scholar] [CrossRef]
- Zhang, B.; Li, M.; Shi, J.; Zeng, M.; Zhang, J.; Liu, J.; Zheng, X.; Feng, W. Six sesquiterpenoids from the stems and leaves of Chrysanthemum morifolium Ramat and their anti-asthma activities. Fitoterapia 2023, 171, 105633. [Google Scholar] [CrossRef]
- Matsuda, H.; Morikawa, T.; Toguchida, I.; Harima, S.; Yoshikawa, M. Medicinal flowers. VI. Absolute stereostructures of two new flavanone glycosides and a phenylbutanoid glycoside from the flowers of Chrysanthemum indicum L.: Their inhibitory activities for rat lens aldose reductase. Chem. Pharm. Bull. 2002, 50, 972–975. [Google Scholar] [CrossRef]
- Miyazawa, M.; Hisama, M. Antimutagenic activity of flavonoids from Chrysanthemum morifolium. Biosci. Biotechnol. Biochem. 2003, 67, 2091–2099. [Google Scholar] [CrossRef]
- Beninger, C.W.; Abou-Zaid, M.M.; Kistner, A.L.; Hallett, R.H.; Iqbal, M.J.; Grodzinski, B.; Hall, J.C. A flavanone and two phenolic acids from Chrysanthemum morifolium with phytotoxic and insect growth regulating activity. J. Chem. Ecol. 2004, 30, 589–606. [Google Scholar] [CrossRef]
- Lin, L.-Z.; Harnly, J.M. Identification of the phenolic components of chrysanthemum flower (Chrysanthemum morifolium Ramat). Food Chem. 2010, 1, 319–326. [Google Scholar] [CrossRef]
- Park, K.H.; Yang, M.S.; Park, M.K.; Kim, S.C.; Yang, C.H.; Park, S.J.; Lee, J.R. A new cytotoxic guaianolide from Chrysanthemum boreale. Fitoterapia 2009, 80, 54–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.S.; Zhou, J.; Kong, L.Y. Three new germacrane-type sesquiterpene stereoisomers from the flowers of Chrysanthemum indicum. Fitoterapia 2012, 83, 1675–1679. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.F.; Li, D.X.; Zhang, R.; Zhao, L.L.; Qiu, Z.; Du, Y.; Ji, S.; Tang, D.Q. Chemical Antioxidant Quality Markers of Chrysanthemum morifolium Using a Spectrum-Effect Approach. Front. Pharmacol. 2022, 7, 809482. [Google Scholar] [CrossRef]
- Peng, A.; Lin, L.; Zhao, M.; Sun, B. Classification of edible chrysanthemums based on phenolic profiles and mechanisms underlying the protective effects of characteristic phenolics on oxidatively damaged erythrocyte. Food Res. Int. 2019, 123, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhou, M.; Wang, L.; Ying, X.; Peng, J.; Jiang, M.; Bai, G.; Luo, G. Comparative evaluation of different cultivars of Flos Chrysanthemi by an anti-inflammatory-based NF-κB reporter gene assay coupled to UPLC-Q/TOF MS with PCA and ANN. J. Ethnopharmacol. 2015, 4, 387–395. [Google Scholar] [CrossRef]
- Housley, L.; Magana, A.A.; Hsu, A.; Beaver, L.M.; Wong, C.P.; Stevens, J.F.; Choi, J.; Jiang, Y.; Bella, D.; Williams, D.E.; et al. Untargeted Metabolomic Screen Reveals Changes in Human Plasma Metabolite Profiles Following Consumption of Fresh Broccoli Sprouts. Mol. Nutr. Food Res. 2018, 62, e1700665. [Google Scholar] [CrossRef]
- Cao, M.; Liu, Y.; Jiang, W.; Meng, X.; Zhang, W.; Chen, W.; Peng, D.; Xing, S. UPLC/MS-based untargeted metabolomics reveals the changes of metabolites profile of Salvia miltiorrhiza bunge during Sweating processing. Sci. Rep. 2020, 11, 19524. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, J.; Sun, Z.; Jiang, W.; Liu, Y.; Tang, J.; Meng, X.; Su, X.; Wu, L.; Wang, L.; et al. ICP-MS based metallomics and GC-MS based metabolomics reveals the physiological and metabolic responses of Dendrobium huoshanense plants exposed to Fe3O4 nanoparticles. Front. Nutr. 2022, 23, 1013756. [Google Scholar] [CrossRef]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Sun, J.; Wang, Z.; Lin, C.; Xia, H.; Yang, L.; Wang, S.; Sun, G. The hypolipidemic mechanism of chrysanthemum flavonoids and its main components, luteolin and luteoloside, based on the gene expression profile. Front. Nutr. 2022, 9, 952588. [Google Scholar] [CrossRef]
- Xue, G.M.; Xue, J.F.; Zhao, C.G.; Zhao, Z.Z.; Zhi, Y.L.; Du, K.; Li, H.W.; Sun, Y.J.; Feng, W.S. 1,10-seco guaianolide-type sesquiterpenoids from Chrysanthemum indicum. Asian Nat. Prod. Res. 2021, 23, 877–883. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.J.; Su, S.L.; Yan, H.; Guo, S.; Qian, D.W.; Duan, J.A. Evaluation of Anti-Inflammatory and Antioxidant Effectsof Chrysanthemum Stem and Leaf Extract on Zebrafish Inflammatory Bowel Disease Model. Molecules 2022, 27, 2114. [Google Scholar] [CrossRef]
- Tian, Z.; Jia, H.; Jin, Y.; Wang, M.; Kou, J.; Wang, C.; Rong, X.; Xie, X.; Han, G.; Pang, X. Chrysanthemum extract attenuates hepatotoxicity via inhibiting oxidative stress in vivo and in vitro. Food Nutr. Res. 2019, 15, 63. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; He, Z.; He, S.; Jing, P. Insights into the importance of dietary chrysanthemum flower (Chrysanthemum morifolium cv. Hangju)-wolfberry (Lycium barbarum fruit) combination in antioxidant and anti-inflammatory properties. Food Res. Int. 2019, 116, 810–818. [Google Scholar] [CrossRef]
- Zhuo, F.F.; Zhang, C.; Zhang, H.; Xia, Y.; Xue, G.M.; Yang, L.; Kong, L.Y. Chrysanthemulide A induces apoptosis through DR5 upregulation via JNK-mediated autophagosome accumulation in human osteosarcoma cells. Cell Physiol. 2019, 234, 13191–13208. [Google Scholar] [CrossRef]
- Yang, X.; Liu, Y.; Zhong, C.; Hu, J.; Xu, S.; Zhang, P.; He, L. Total flavonoids of Chrysanthemum indicum L inhibit acute pancreatitis through suppressing apoptosis and inflammation. BMC Complement Med. Ther. 2023, 23, 23. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.C.; Song, Y.; Xiao, P.; Zhong, Y.; Wu, P.; Xu, L. The genus Chrysanthemum: Phylogeny, biodiversity, phytometabolites, and chemodiversity. Front. Plant Sci. 2022, 11, 973197. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Huang, X.; Zhu, Y.; Li, J.; Miao, Y.; Du, H.; Liu, D. Comparison of Chemical Constituents and Pharmacological Effects of Different Varieties of Chrysanthemum Flos in China. Chem. Biodivers. 2021, 18, e2100206. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, C.; Zhou, J.; Zhou, F.; Gui, A.; Chu, H.; Shao, Q. Chrysanthemum morifolium as a traditional herb: A review of historical development, classification, phytochemistry, pharmacology and application. J. Ethnopharmacol. 2024, 10, 118198. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Li, L.; He, Y.; Zhang, X.; Zhao, Y.; Wang, Q.; Hong, G. Flavonoids in vegetables: Improvement of dietary flavonoids by metabolic engineering to promote health. Crit. Rev. Food Sci. Nutr. 2024, 64, 3220–3234. [Google Scholar] [CrossRef] [PubMed]
- Franza, L.; Carusi, V.; Nucera, E.; Pandolfi, F. Luteolin, inflammation and cancer: Special emphasis on gut microbiota. Biofactors 2021, 47, 181–189. [Google Scholar] [CrossRef]
- Nguyen, T.D. Rhamnosyltransferases in Chrysanthemum: Just a spoonful of sugar helps the flavonoid-based medicines abound. Plant Physiol. 2022, 28, 2061–2063. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.W.; Wei, M.; Feng, L.F.; Ding, L.; Wei, W.K.; Yang, J.F.; Lin, X.J.; Liang, H.L.; Zhan, R.T.; Ma, D.M. Rhamnosyltransferases involved in the biosynthesis of flavone rutinosides in Chrysanthemum species. Plant Physiol. 2022, 190, 2122–2136. [Google Scholar] [CrossRef]
- Sun, J.; Wang, Z.; Chen, L.; Sun, G. Hypolipidemic Effects and Preliminary Mechanism of Chrysanthemum Flavonoids, Its Main Components Luteolin and Luteoloside in Hyperlipidemia Rats. Antioxidants 2021, 10, 1309. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Xu, Z.; Li, M.; Wang, K.; Pang, S.; Ni, Y. The formation process of green substances in Chrysanthemum morifolium tea. Food Chem. 2020, 1, 127028. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Tian, Z.; Cui, Y.; Liu, Z.; Ma, X. Chlorogenic acid: A comprehensive review of the dietary sources, processing effects, bioavailability, beneficial properties, mechanisms of action, and future directions. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3130–3158. [Google Scholar] [CrossRef]
- Gong, J.; Chu, B.; Gong, L.; Fang, Z.; Zhang, X.; Qiu, S.; Wang, J.; Xiang, Y.; Xiao, G.; Yuan, H.; et al. Comparison of Phenolic Compounds and the Antioxidant Activities of Fifteen Chrysanthemum morifolium Ramat cv. ‘Hangbaiju’ in China. Antioxidants 2019, 20, 325. [Google Scholar] [CrossRef]
- Kuang, C.L.; Lv, D.; Shen, G.H.; Li, S.S.; Luo, Q.Y.; Zhang, Z.Q. Chemical composition and antimicrobial activities of volatile oil extracted from Chrysanthemum morifolium Ramat. Food Sci. Technol. 2018, 55, 2786–2794. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, W.; Chen, X.; Wang, W.; Köllner, T.G.; Chen, S.; Chen, F.; Chen, F. Diversity and Biosynthesis of Volatile Terpenoid Secondary Metabolites in the Chrysanthemum Genus. Crit. Rev. Plant Sci. 2021, 40, 422–445. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Zhao, P.; Ren, L.; Li, X.; Gao, W. Physicochemical characteristics and immunoregulatory activities of polysaccharides from five cultivars of Chrysanthemi Flos. Food Sci. Nutr. 2022, 10, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, X.; Chen, H.; Lu, J.; Chen, D.; Luo, C.; Cheng, X.; Jia, Y.; Huang, C. Transcriptome and Metabolome Analyses of the Flowers and Leaves of Chrysanthemum dichrum. Front. Genet. 2021, 31, 716163. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Positive Ion Mode | Negative Ion Mode |
|---|---|---|
| Spray voltage (V) | 3800 | −3000 |
| Capillary temperature (°C) | 320 | |
| Auxiliary gas heater temperature (°C) | 350 | |
| Sheath gas flow rate (arb) | 35 | |
| Auxiliary gas flow rate (arb) | 8 | |
| S−lens RF level | 50 | |
| Mass range (m/z) | 70–1050 | |
| Full MS resolution | 60,000 | |
| MS/MS resolution | 15,000 | |
| NCE/stepped NCE | 10, 20, 40 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, N.; Fan, J.; Li, G.; Guo, X.; Meng, X.; Wang, Y.; Duan, Y.; Ding, W.; Liu, K.; Liu, Y.; et al. Comparative Analysis of the Chemical Constituents of Chrysanthemum morifolium with Different Drying Processes Integrating LC/GC–MS−Based, Non-Targeted Metabolomics. Metabolites 2024, 14, 481. https://doi.org/10.3390/metabo14090481
Chen N, Fan J, Li G, Guo X, Meng X, Wang Y, Duan Y, Ding W, Liu K, Liu Y, et al. Comparative Analysis of the Chemical Constituents of Chrysanthemum morifolium with Different Drying Processes Integrating LC/GC–MS−Based, Non-Targeted Metabolomics. Metabolites. 2024; 14(9):481. https://doi.org/10.3390/metabo14090481
Chicago/Turabian StyleChen, Na, Jizhou Fan, Gang Li, Xuanxuan Guo, Xiao Meng, Yuqing Wang, Yingying Duan, Wanyue Ding, Kai Liu, Yaowu Liu, and et al. 2024. "Comparative Analysis of the Chemical Constituents of Chrysanthemum morifolium with Different Drying Processes Integrating LC/GC–MS−Based, Non-Targeted Metabolomics" Metabolites 14, no. 9: 481. https://doi.org/10.3390/metabo14090481
APA StyleChen, N., Fan, J., Li, G., Guo, X., Meng, X., Wang, Y., Duan, Y., Ding, W., Liu, K., Liu, Y., & Xing, S. (2024). Comparative Analysis of the Chemical Constituents of Chrysanthemum morifolium with Different Drying Processes Integrating LC/GC–MS−Based, Non-Targeted Metabolomics. Metabolites, 14(9), 481. https://doi.org/10.3390/metabo14090481