
Examining the Pathogenesis of MAFLD and the Medicinal Properties of Natural Products from a Metabolic Perspective


Abstract
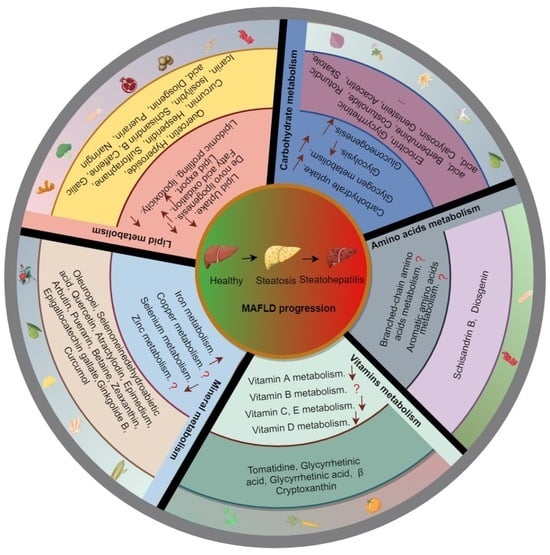
1. Introduction
2. Nutrient Metabolism and MAFLD
2.1. Macronutrient Metabolism and MAFLD
2.1.1. Lipid Metabolism and MAFLD
Lipid Uptake and MAFLD
De Novo Lipogenesis and MAFLD
Fatty Acid Oxidation and MAFLD
Lipid Export and MAFLD
Lipidomic Profiling, Lipotoxicity, and MAFLD
2.1.2. Carbohydrate Metabolism and MAFLD
Carbohydrate Uptake and MAFLD
Glycogen Metabolism and MAFLD
Glycolysis and MAFLD
Gluconeogenesis and MAFLD
2.1.3. Amino Acid Metabolism and MAFLD
Branched-Chain Amino Acid Metabolism and MAFLD
Aromatic Amino Acid Metabolism and MAFLD
2.2. Micronutrient Metabolism and MAFLD
2.2.1. Vitamin Metabolism and MAFLD
Vitamin A Metabolism and MAFLD
Vitamin B Metabolism and MAFLD
Vitamin C and Vitamin E Metabolism and MAFLD
Vitamin D Metabolism and MAFLD
2.2.2. Mineral Metabolism and MAFLD
Iron Metabolism and MAFLD
Copper Metabolism and MAFLD
Zinc Metabolism and MAFLD
Selenium Metabolism and MAFLD
3. Natural Products against MAFLD via the Modulation of Nutrient Metabolism
3.1. Natural Products and Lipid Metabolism in MAFLD
3.1.1. Natural Products and Lipid Uptake in MAFLD
3.1.2. Natural Products and DNL in MAFLD
3.1.3. Natural Products and Fatty Acid Oxidation in MAFLD
3.1.4. Natural Products and Lipid Export in MAFLD
3.2. Natural Products and Carbohydrate Metabolism in MAFLD
3.2.1. Natural Products and Carbohydrate Uptake in MAFLD
3.2.2. Natural Products and Glycogen Metabolism in MAFLD
3.2.3. Natural Products and Glycolysis in MAFLD
3.2.4. Natural Products and Gluconeogenesis in MAFLD
3.3. Natural Products and Amino Acid Metabolism in MAFLD
3.4. Natural Products and Micronutrient Metabolism in MAFLD
3.4.1. Natural Products and Vitamin Metabolism in MAFLD
3.4.2. Natural Products and Mineral Metabolism in MAFLD
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviations | Full names |
| ACC | Acetyl-CoA Carboxylase |
| ACSL4 | Acyl-CoA Synthetase Long-chain family member 4 |
| ACSL5 | Acyl-CoA Synthetase Long-chain family member 5 |
| AGPAT | Acylglycerol-3-Phosphate Acyl Transferase |
| AhR | Arylhydrocarbon Receptor |
| ApoB100 | Apolipoprotein B100 |
| BCATs | branched-chain aminotransferases |
| BCKDs | branched-chain α-ketoacid dehydrogenases |
| BMP4 | Bone morphogenetic protein 4 |
| CACT | Carnitine Acyl Carnitine Translocase |
| CCTα | Cytidine triphosphate: phosphocholine Cytidylyltransferase-α |
| CD36 | Cluster Determinant 36 |
| ChREBP | Carbohydrate Response Element Binding Protein |
| CP | ceruloplasmin |
| CPT1 | Carnitine Palmitoyl Transferase 1 |
| CPT2 | Carnitine Palmitoyl Transferase 2 |
| CYP4A | cytochrome P450 |
| DGAT | diacylglycerol acyltransferase |
| DLAT | dihydrolipoamide S-acetyltransferase |
| EGFR | epidermal cell growth factor receptor |
| EIF5A | Eukaryotic Initiation Factor 5A |
| FAO | fatty acid oxidation |
| FAS | Fatty Acid Synthase |
| FATPs | Fatty Acid Transporter Proteins |
| Fbpase | fructose-1,6-bisphosphatase |
| FGFR | fibroblast growth factor receptor |
| FMO2 | Flavin-containing Monooxygenase 2 |
| G6Pase | Glucose-6-phosphatase |
| GBE | glycogen branching enzyme |
| GCK | glucokinase |
| GLUTs | Glucose Transporter proteins |
| GPAT | Glycerol-3-Phosphate Acyl Transferase |
| GPCR | G protein-coupled receptor |
| GPXs | glutathione peroxidases |
| GSK3β | glycogen synthase kinase 3β |
| GYS | glycogen synthase |
| HIBCH | 3-hydroxyisobutyryl coenzyme A hydrolase |
| HK2 | Hexokinase 2 |
| HNF4α | Hepatocyte Nuclear Factor 4α |
| INSIG2 | Insulin-Induced Gene 2 |
| LXRs | Liver X Receptors |
| MCJ | Methylation-Controlled J protein |
| MMPs | matrix metalloproteinases |
| mTORC1 | mammalian Target of Rapamycin Complex 1 |
| MTTP | Microsomal Triglyceride Transfer Protein |
| PAPs | Phosphatidic Acid Phosphatases |
| PAR2 | Protease-Activated Receptor 2 |
| PEPCK | phosphoenolpyruvate carboxykinase |
| PFK1 | phosphofructokinase 1 |
| PFKFB3 | fructose-2,6-bisphosphatase 3 |
| PKM2 | pyruvate kinase type M2 |
| PPARα | Peroxisome proliferator-activated receptor α |
| PPARγ | Peroxisome Proliferator-Activated Receptor γ |
| RAR | retinoic acid receptor |
| RTKs | receptor tyrosine kinases |
| RXR | retinoid X receptor |
| SCAP | SREBP Cleavage-Activating Protein |
| SGLT2 | Sodium-dependent Glucose Transporter protein 2 |
| SMLR1 | Small Leucine-Rich protein 1 |
| SOD1 | superoxide dismutase 1 |
| SREBPs | Sterol Regulatory Element Binding Proteins |
| THRβ | Thyroid hormone receptor β subtype |
| TM4SF5 | Transmembrane 4L Six Family member 5 |
| TM6SF2 | Transmembrane 6 Superfamily member 2 |
| TXNRDs | thioredoxin reductases |
| VDR | vitamin D receptor |
| VLDL | Very Low-Density Lipoprotein |
| ZFPs | zinc finger proteins |
References
- Cobbina, E.; Akhlaghi, F. Non-Alcoholic Fatty Liver Disease (NAFLD)—Pathogenesis, Classification, and Effect on Drug Metabolizing Enzymes and Transporters. Drug Metab. Rev. 2017, 49, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Diehl, A.M.; Brunt, E.M.; Cusi, K.; Charlton, M.; Sanyal, A.J. The Diagnosis and Management of Non-Alcoholic Fatty Liver Disease: Practice Guideline by the American Association for the Study of Liver Diseases, American College of Gastroenterology, and the American Gastroenterological Association. Hepatology 2012, 55, 2005–2023. [Google Scholar] [CrossRef] [PubMed]
- Rong, L.; Zou, J.; Ran, W.; Qi, X.; Chen, Y.; Cui, H.; Guo, J. Advancements in the Treatment of Non-Alcoholic Fatty Liver Disease (NAFLD). Front. Endocrinol. 2022, 13, 1087260. [Google Scholar] [CrossRef] [PubMed]
- Eslam, M.; Sanyal, A.J.; George, J.; Sanyal, A.; Neuschwander-Tetri, B.; Tiribelli, C.; Kleiner, D.E.; Brunt, E.; Bugianesi, E.; Yki-Järvinen, H.; et al. MAFLD: A Consensus-Driven Proposed Nomenclature for Metabolic Associated Fatty Liver Disease. Gastroenterology 2020, 158, 1999–2014.e1. [Google Scholar] [CrossRef] [PubMed]
- Gofton, C.; Upendran, Y.; Zheng, M.-H.; George, J. MAFLD: How Is It Different from NAFLD? Clin. Mol. Hepatol. 2023, 29, S17–S31. [Google Scholar] [CrossRef] [PubMed]
- Lim, G.E.H.; Tang, A.; Ng, C.H.; Chin, Y.H.; Lim, W.H.; Tan, D.J.H.; Yong, J.N.; Xiao, J.; Lee, C.W.-M.; Chan, M.; et al. An Observational Data Meta-Analysis on the Differences in Prevalence and Risk Factors Between MAFLD vs NAFLD. Clin. Gastroenterol. Hepatol. 2023, 21, 619–629.e7. [Google Scholar] [CrossRef] [PubMed]
- Lei, F.; Qin, J.-J.; Song, X.; Liu, Y.-M.; Chen, M.-M.; Sun, T.; Huang, X.; Deng, K.-Q.; Zuo, X.; Yao, D.; et al. The Prevalence of MAFLD and Its Association with Atrial Fibrillation in a Nationwide Health Check-up Population in China. Front. Endocrinol. 2022, 13, 1007171. [Google Scholar] [CrossRef]
- Bechmann, L.P.; Hannivoort, R.A.; Gerken, G.; Hotamisligil, G.S.; Trauner, M.; Canbay, A. The Interaction of Hepatic Lipid and Glucose Metabolism in Liver Diseases. J. Hepatol. 2012, 56, 952–964. [Google Scholar] [CrossRef] [PubMed]
- Esler, W.P.; Bence, K.K. Metabolic Targets in Nonalcoholic Fatty Liver Disease. Cell. Mol. Gastroenterol. Hepatol. 2019, 8, 247–267. [Google Scholar] [CrossRef]
- Pickett-Blakely, O.; Young, K.; Carr, R.M. Micronutrients in Nonalcoholic Fatty Liver Disease Pathogenesis. Cell. Mol. Gastroenterol. Hepatol. 2018, 6, 451–462. [Google Scholar] [CrossRef]
- Eng, J.M.; Estall, J.L. Diet-Induced Models of Non-Alcoholic Fatty Liver Disease: Food for Thought on Sugar, Fat, and Cholesterol. Cells 2021, 10, 1805. [Google Scholar] [CrossRef]
- Cioffi, F.; Giacco, A.; Petito, G.; de Matteis, R.; Senese, R.; Lombardi, A.; de Lange, P.; Moreno, M.; Goglia, F.; Lanni, A.; et al. Altered Mitochondrial Quality Control in Rats with Metabolic Dysfunction-Associated Fatty Liver Disease (MAFLD) Induced by High-Fat Feeding. Genes 2022, 13, 315. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and Resupply of Pharmacologically Active Plant-Derived Natural Products: A Review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular Mechanisms of Hepatic Lipid Accumulation in Non-Alcoholic Fatty Liver Disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef]
- Softic, S.; Boucher, J.; Solheim, M.H.; Fujisaka, S.; Haering, M.-F.; Homan, E.P.; Winnay, J.; Perez-Atayde, A.R.; Kahn, C.R. Lipodystrophy Due to Adipose Tissue-Specific Insulin Receptor Knockout Results in Progressive NAFLD. Diabetes 2016, 65, 2187–2200. [Google Scholar] [CrossRef]
- Feng, Y.; Sun, W.; Sun, F.; Yin, G.; Liang, P.; Chen, S.; Liu, X.; Jiang, T.; Zhang, F. Biological Mechanisms and Related Natural Inhibitors of CD36 in Nonalcoholic Fatty Liver. Drug Des. Dev. Ther. 2022, 16, 3829–3845. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.-W.; Wang, J.; Guo, H.; Zhao, Y.-Y.; Sun, H.-H.; Li, Y.-F.; Lai, X.-Y.; Zhao, N.; Wang, X.; Xie, C.; et al. CD36 Facilitates Fatty Acid Uptake by Dynamic Palmitoylation-Regulated Endocytosis. Nat. Commun. 2020, 11, 4765. [Google Scholar] [CrossRef]
- Yang, S.; Jia, L.; Xiang, J.; Yang, G.; Qiu, S.; Kang, L.; Zheng, P.; Liang, Z.; Lu, Y. KLF10 Promotes Nonalcoholic Steatohepatitis Progression through Transcriptional Activation of zDHHC7. EMBO Rep 2022, 23, e54229. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, C.; Luo, X.; Wang, P.; Zhou, W.; Zhong, S.; Xie, Y.; Jiang, Y.; Yang, P.; Tang, R.; et al. CD36 Palmitoylation Disrupts Free Fatty Acid Metabolism and Promotes Tissue Inflammation in Non-Alcoholic Steatohepatitis. J. Hepatol. 2018, 69, 705–717. [Google Scholar] [CrossRef]
- Anderson, C.M.; Stahl, A. SLC27 Fatty Acid Transport Proteins. Mol. Asp. Med. 2013, 34, 516–528. [Google Scholar] [CrossRef]
- Qiu, P.; Wang, H.; Zhang, M.; Zhang, M.; Peng, R.; Zhao, Q.; Liu, J. FATP2-Targeted Therapies—A Role beyond Fatty Liver Disease. Pharmacol. Res. 2020, 161, 105228. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Yang, X.; Kuang, Q.; Wu, Y.; Tan, X.; Lan, J.; Qiang, Z.; Feng, T. HBx Induced Upregulation of FATP2 Promotes the Development of Hepatic Lipid Accumulation. Exp. Cell Res. 2023, 430, 113721. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Liu, J.; Wang, H.; Wen, K.; Ni, X.; Lin, Y.; Huang, J.; You, X.; Lei, Z.; Li, J.; et al. Nanoplastic Propels Diet-Induced NAFL to NASH via ER-Mitochondrial Tether-Controlled Redox Switch. J. Hazard. Mater. 2023, 465, 133142. [Google Scholar] [CrossRef] [PubMed]
- Auinger, A.; Valenti, L.; Pfeuffer, M.; Helwig, U.; Herrmann, J.; Fracanzani, A.L.; Dongiovanni, P.; Fargion, S.; Schrezenmeir, J.; Rubin, D. A Promoter Polymorphism in the Liver-Specific Fatty Acid Transport Protein 5 Is Associated with Features of the Metabolic Syndrome and Steatosis. Horm. Metab. Res. 2010, 42, 854–859. [Google Scholar] [CrossRef]
- Doege, H.; Baillie, R.A.; Ortegon, A.M.; Tsang, B.; Wu, Q.; Punreddy, S.; Hirsch, D.; Watson, N.; Gimeno, R.E.; Stahl, A. Targeted Deletion of FATP5 Reveals Multiple Functions in Liver Metabolism: Alterations in Hepatic Lipid Homeostasis. Gastroenterology 2006, 130, 1245–1258. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, B.; Doege, H.; Punreddy, S.; Wu, H.; Huang, X.; Kaushik, V.K.; Mozell, R.L.; Byrnes, J.J.; Stricker-Krongrad, A.; Chou, C.J.; et al. Mice Deleted for Fatty Acid Transport Protein 5 Have Defective Bile Acid Conjugation and Are Protected from Obesity. Gastroenterology 2006, 130, 1259–1269. [Google Scholar] [CrossRef] [PubMed]
- Mitsuyoshi, H.; Yasui, K.; Harano, Y.; Endo, M.; Tsuji, K.; Minami, M.; Itoh, Y.; Okanoue, T.; Yoshikawa, T. Analysis of Hepatic Genes Involved in the Metabolism of Fatty Acids and Iron in Nonalcoholic Fatty Liver Disease. Hepatol. Res. 2009, 39, 366–373. [Google Scholar] [CrossRef]
- Enooku, K.; Tsutsumi, T.; Kondo, M.; Fujiwara, N.; Sasako, T.; Shibahara, J.; Kado, A.; Okushin, K.; Fujinaga, H.; Nakagomi, R.; et al. Hepatic FATP5 Expression Is Associated with Histological Progression and Loss of Hepatic Fat in NAFLD Patients. J. Gastroenterol. 2020, 55, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Ameer, F.; Scandiuzzi, L.; Hasnain, S.; Kalbacher, H.; Zaidi, N. De Novo Lipogenesis in Health and Disease. Metabolism 2014, 63, 895–902. [Google Scholar] [CrossRef]
- Sanders, F.W.B.; Griffin, J.L. De Novo Lipogenesis in the Liver in Health and Disease: More than Just a Shunting Yard for Glucose. Biol. Rev. Camb. Philos. Soc. 2016, 91, 452–468. [Google Scholar] [CrossRef]
- Wang, Y.; Viscarra, J.; Kim, S.-J.; Sul, H.S. Transcriptional Regulation of Hepatic Lipogenesis. Nat. Rev. Mol. Cell Biol. 2015, 16, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Régnier, M.; Carbinatti, T.; Parlati, L.; Benhamed, F.; Postic, C. The Role of ChREBP in Carbohydrate Sensing and NAFLD Development. Nat. Rev. Endocrinol. 2023, 19, 336–349. [Google Scholar] [CrossRef]
- Febbraio, M.A.; Karin, M. “Sweet Death”: Fructose as a Metabolic Toxin That Targets the Gut-Liver Axis. Cell Metab. 2021, 33, 2316–2328. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, A.M.; Bentanachs, R.; Sala-Vila, A.; Lázaro, I.; Rodríguez-Morató, J.; Sánchez, R.M.; Alegret, M.; Roglans, N.; Laguna, J.C. ChREBP-Driven DNL and PNPLA3 Expression Induced by Liquid Fructose Are Essential in the Production of Fatty Liver and Hypertriglyceridemia in a High-Fat Diet-Fed Rat Model. Mol. Nutr. Food Res. 2022, 66, e2101115. [Google Scholar] [CrossRef]
- Geidl-Flueck, B.; Gerber, P.A. Fructose Drives de Novo Lipogenesis Affecting Metabolic Health. J. Endocrinol. 2023, 257, e220270. [Google Scholar] [CrossRef] [PubMed]
- Daniel, P.V.; Dogra, S.; Rawat, P.; Choubey, A.; Khan, A.S.; Rajak, S.; Kamthan, M.; Mondal, P. NF-κB P65 Regulates Hepatic Lipogenesis by Promoting Nuclear Entry of ChREBP in Response to a High Carbohydrate Diet. J. Biol. Chem. 2021, 296, 100714. [Google Scholar] [CrossRef]
- Nikolaou, K.C.; Godbersen, S.; Manoharan, M.; Wieland, S.; Heim, M.H.; Stoffel, M. Inflammation-Induced TRIM21 Represses Hepatic Steatosis by Promoting the Ubiquitination of Lipogenic Regulators. JCI Insight 2023, 8, e164694. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Iizuka, K.; Takao, K.; Horikawa, Y.; Kitamura, T.; Takeda, J. ChREBP-Knockout Mice Show Sucrose Intolerance and Fructose Malabsorption. Nutrients 2018, 10, 340. [Google Scholar] [CrossRef]
- Jang, J.; Hwang, S.; Oh, A.-R.; Park, S.; Yaseen, U.; Kim, J.G.; Park, S.; Jung, Y.; Cha, J.-Y. Fructose Malabsorption in ChREBP-Deficient Mice Disrupts the Small Intestine Immune Microenvironment and Leads to Diarrhea-Dominant Bowel Habit Changes. Inflamm. Res. 2023, 72, 769–782. [Google Scholar] [CrossRef]
- Shimano, H.; Sato, R. SREBP-Regulated Lipid Metabolism: Convergent Physiology—Divergent Pathophysiology. Nat. Rev. Endocrinol. 2017, 13, 710–730. [Google Scholar] [CrossRef]
- Ferré, P.; Foufelle, F. Hepatic Steatosis: A Role for de Novo Lipogenesis and the Transcription Factor SREBP-1c. Diabetes Obes. Metab. 2010, 12 (Suppl. S2), 83–92. [Google Scholar] [CrossRef]
- Li, N.; Li, X.; Ding, Y.; Liu, X.; Diggle, K.; Kisseleva, T.; Brenner, D.A. SREBP Regulation of Lipid Metabolism in Liver Disease, and Therapeutic Strategies. Biomedicines 2023, 11, 3280. [Google Scholar] [CrossRef] [PubMed]
- Ke, C.; Xiao, C.; Li, J.; Wu, X.; Zhang, Y.; Chen, Y.; Sheng, S.; Fu, Z.; Wang, L.; Ni, C.; et al. FMO2 Ameliorates Nonalcoholic Fatty Liver Disease by Suppressing ER-to-Golgi Transport of SREBP1. Hepatology 2023. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Qin, H.; Liao, M.; Zheng, E.; Luo, X.; Xiao, A.; Li, Y.; Chen, L.; Wei, L.; Zhao, L.; et al. CD36 Promotes de Novo Lipogenesis in Hepatocytes through INSIG2-Dependent SREBP1 Processing. Mol. Metab. 2022, 57, 101428. [Google Scholar] [CrossRef]
- Kim, J.Y.; Garcia-Carbonell, R.; Yamachika, S.; Zhao, P.; Dhar, D.; Loomba, R.; Kaufman, R.J.; Saltiel, A.R.; Karin, M. ER Stress Drives Lipogenesis and Steatohepatitis via Caspase-2 Activation of S1P. Cell 2018, 175, 133–145.e15. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Wang, L.Q.; Sladky, V.C.; Oh, T.G.; Liu, J.; Trinh, K.; Eichin, F.; Downes, M.; Hosseini, M.; Jacotot, E.D.; et al. PIDDosome-SCAP Crosstalk Controls High-Fructose-Diet-Dependent Transition from Simple Steatosis to Steatohepatitis. Cell Metab. 2022, 34, 1548–1560.e6. [Google Scholar] [CrossRef]
- Kawamura, S.; Matsushita, Y.; Kurosaki, S.; Tange, M.; Fujiwara, N.; Hayata, Y.; Hayakawa, Y.; Suzuki, N.; Hata, M.; Tsuboi, M.; et al. Inhibiting SCAP/SREBP Exacerbates Liver Injury and Carcinogenesis in Murine Nonalcoholic Steatohepatitis. J. Clin. Investig. 2022, 132, e151895. [Google Scholar] [CrossRef] [PubMed]
- Houten, S.M.; Violante, S.; Ventura, F.V.; Wanders, R.J.A. The Biochemistry and Physiology of Mitochondrial Fatty Acid β-Oxidation and Its Genetic Disorders. Annu. Rev. Physiol. 2016, 78, 23–44. [Google Scholar] [CrossRef]
- Adeva-Andany, M.M.; Carneiro-Freire, N.; Seco-Filgueira, M.; Fernández-Fernández, C.; Mouriño-Bayolo, D. Mitochondrial β-Oxidation of Saturated Fatty Acids in Humans. Mitochondrion 2019, 46, 73–90. [Google Scholar] [CrossRef]
- Kleiboeker, B.; Lodhi, I.J. Peroxisomal Regulation of Energy Homeostasis: Effect on Obesity and Related Metabolic Disorders. Mol. Metab. 2022, 65, 101577. [Google Scholar] [CrossRef]
- Reddy, J.K. Nonalcoholic Steatosis and Steatohepatitis. III. Peroxisomal Beta-Oxidation, PPAR Alpha, and Steatohepatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G1333–G1339. [Google Scholar] [CrossRef] [PubMed]
- Pyper, S.R.; Viswakarma, N.; Yu, S.; Reddy, J.K. PPARalpha: Energy Combustion, Hypolipidemia, Inflammation and Cancer. Nucl. Recept. Signal 2010, 8, e002. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.P.; Cunningham, R.P.; Meers, G.M.; Johnson, S.A.; Wheeler, A.A.; Ganga, R.R.; Spencer, N.M.; Pitt, J.B.; Diaz-Arias, A.; Swi, A.I.A.; et al. Compromised Hepatic Mitochondrial Fatty Acid Oxidation and Reduced Markers of Mitochondrial Turnover in Human NAFLD. Hepatology 2022, 76, 1452–1465. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Tian, Y.; Cao, Z.; Zhang, J.; Feng, T.; Tao, W.; Sun, H.; Wen, H.; Lu, X.; Zhu, Q.; et al. Cytoplasmic SIRT6-Mediated ACSL5 Deacetylation Impedes Nonalcoholic Fatty Liver Disease by Facilitating Hepatic Fatty Acid Oxidation. Mol. Cell 2022, 82, 4099–4115.e9. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Wang, Z.; Duan, R.; Yang, C.; Zhao, R.; Feng, Q.; Qin, Y.; Jiang, J.; Gu, S.; Lv, K.; et al. Therapeutic Targeting of Hepatic ACSL4 Ameliorates NASH in Mice. Hepatology 2022, 75, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Barbier-Torres, L.; Fortner, K.A.; Iruzubieta, P.; Delgado, T.C.; Giddings, E.; Chen, Y.; Champagne, D.; Fernández-Ramos, D.; Mestre, D.; Gomez-Santos, B.; et al. Silencing Hepatic MCJ Attenuates Non-Alcoholic Fatty Liver Disease (NAFLD) by Increasing Mitochondrial Fatty Acid Oxidation. Nat. Commun. 2020, 11, 3360. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Pang, J.; Tripathi, M.; Ho, J.P.; Widjaja, A.A.; Shekeran, S.G.; Cook, S.A.; Suzuki, A.; Diehl, A.M.; Petretto, E.; et al. Spermidine-Mediated Hypusination of Translation Factor EIF5A Improves Mitochondrial Fatty Acid Oxidation and Prevents Non-Alcoholic Steatohepatitis Progression. Nat. Commun. 2022, 13, 5202. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Wu, F.; Chen, M.; Li, Y.; You, M.; Zhang, Y.; Yang, P.; Wei, L.; Ruan, X.Z.; Zhao, L.; et al. Inhibition of Fatty Acid Translocase (FAT/CD36) Palmitoylation Enhances Hepatic Fatty Acid β-Oxidation by Increasing Its Localization to Mitochondria and Interaction with Long-Chain Acyl-CoA Synthetase 1. Antioxid. Redox Signal. 2022, 36, 1081–1100. [Google Scholar] [CrossRef] [PubMed]
- Alves-Bezerra, M.; Cohen, D.E. Triglyceride Metabolism in the Liver. Compr. Physiol. 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Van Zwol, W.; van de Sluis, B.; Ginsberg, H.N.; Kuivenhoven, J.A. VLDL Biogenesis and Secretion: It Takes a Village. Circ. Res. 2024, 134, 226–244. [Google Scholar] [CrossRef]
- Amin, N.B.; Saxena, A.R.; Somayaji, V.; Dullea, R. Inhibition of Diacylglycerol Acyltransferase 2 Versus Diacylglycerol Acyltransferase 1: Potential Therapeutic Implications of Pharmacology. Clin. Ther. 2023, 45, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Gluchowski, N.L.; Gabriel, K.R.; Chitraju, C.; Bronson, R.T.; Mejhert, N.; Boland, S.; Wang, K.; Lai, Z.W.; Farese, R.V.; Walther, T.C. Hepatocyte Deletion of Triglyceride-Synthesis Enzyme Acyl CoA: Diacylglycerol Acyltransferase 2 Reduces Steatosis without Increasing Inflammation or Fibrosis in Mice. Hepatology 2019, 70, 1972–1985. [Google Scholar] [CrossRef] [PubMed]
- Heeren, J.; Scheja, L. Metabolic-Associated Fatty Liver Disease and Lipoprotein Metabolism. Mol. Metab. 2021, 50, 101238. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yu, X.-H.; Ou, X.; Ouyang, X.-P.; Tang, C.-K. Hepatic Cholesterol Transport and Its Role in Non-Alcoholic Fatty Liver Disease and Atherosclerosis. Prog. Lipid Res. 2021, 83, 101109. [Google Scholar] [CrossRef] [PubMed]
- Newberry, E.P.; Hall, Z.; Xie, Y.; Molitor, E.A.; Bayguinov, P.O.; Strout, G.W.; Fitzpatrick, J.A.J.; Brunt, E.M.; Griffin, J.L.; Davidson, N.O. Liver-Specific Deletion of Mouse Tm6sf2 Promotes Steatosis, Fibrosis, and Hepatocellular Cancer. Hepatology 2021, 74, 1203–1219. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Smagris, E.; Martin, S.A.; Vale, G.; McDonald, J.G.; Fletcher, J.A.; Burgess, S.C.; Hobbs, H.H.; Cohen, J.C. Hepatic TM6SF2 Is Required for Lipidation of VLDL in a Pre-Golgi Compartment in Mice and Rats. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 879–899. [Google Scholar] [CrossRef]
- Van Zwol, W.; Rimbert, A.; Wolters, J.C.; Smit, M.; Bloks, V.W.; Kloosterhuis, N.J.; Huijkman, N.C.A.; Koster, M.H.; Tharehalli, U.; de Neck, S.M.; et al. Loss of Hepatic SMLR1 Causes Hepatosteatosis and Protects against Atherosclerosis Due to Decreased Hepatic VLDL Secretion. Hepatology 2023, 78, 1418–1432. [Google Scholar] [CrossRef] [PubMed]
- Van der Veen, J.N.; Kennelly, J.P.; Wan, S.; Vance, J.E.; Vance, D.E.; Jacobs, R.L. The Critical Role of Phosphatidylcholine and Phosphatidylethanolamine Metabolism in Health and Disease. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1558–1572. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.; Nuñez-García, M.; Fernández-Tussy, P.; Barbier-Torres, L.; Fernández-Ramos, D.; Gómez-Santos, B.; Buqué, X.; Lopitz-Otsoa, F.; Goikoetxea-Usandizaga, N.; Serrano-Macia, M.; et al. Targeting Hepatic Glutaminase 1 Ameliorates Non-Alcoholic Steatohepatitis by Restoring Very-Low-Density Lipoprotein Triglyceride Assembly. Cell Metab. 2020, 31, 605–622.e10. [Google Scholar] [CrossRef]
- Jiang, X.; Fulte, S.; Deng, F.; Chen, S.; Xie, Y.; Chao, X.; He, X.C.; Zhang, Y.; Li, T.; Li, F.; et al. Lack of VMP1 Impairs Hepatic Lipoprotein Secretion and Promotes Non-Alcoholic Steatohepatitis. J. Hepatol. 2022, 77, 619–631. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, A.; Ding, W.-X.; Ni, H.-M. VMP1 Regulates Hepatic Lipoprotein Secretion and NASH Independent of Autophagy. Autophagy 2023, 19, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Uehara, K.; Sostre-Colón, J.; Gavin, M.; Santoleri, D.; Leonard, K.-A.; Jacobs, R.L.; Titchenell, P.M. Activation of Liver mTORC1 Protects Against NASH via Dual Regulation of VLDL-TAG Secretion and De Novo Lipogenesis. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 1625–1647. [Google Scholar] [CrossRef]
- Hackl, M.T.; Fürnsinn, C.; Schuh, C.M.; Krssak, M.; Carli, F.; Guerra, S.; Freudenthaler, A.; Baumgartner-Parzer, S.; Helbich, T.H.; Luger, A.; et al. Brain Leptin Reduces Liver Lipids by Increasing Hepatic Triglyceride Secretion and Lowering Lipogenesis. Nat. Commun. 2019, 10, 2717. [Google Scholar] [CrossRef]
- Metz, M.; Beghini, M.; Wolf, P.; Pfleger, L.; Hackl, M.; Bastian, M.; Freudenthaler, A.; Harreiter, J.; Zeyda, M.; Baumgartner-Parzer, S.; et al. Leptin Increases Hepatic Triglyceride Export via a Vagal Mechanism in Humans. Cell Metab. 2022, 34, 1719–1731.e5. [Google Scholar] [CrossRef]
- Guo, Z.; Du, H.; Guo, Y.; Jin, Q.; Liu, R.; Yun, Z.; Zhang, J.; Li, X.; Ye, Y. Association between Leptin and NAFLD: A Two-Sample Mendelian Randomization Study. Eur. J. Med. Res. 2023, 28, 215. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Shapiro, M.D. Apolipoproteins in Vascular Biology and Atherosclerotic Disease. Nat. Rev. Cardiol. 2022, 19, 168–179. [Google Scholar] [CrossRef]
- Musso, G.; Saba, F.; Cassader, M.; Gambino, R. Lipidomics in Pathogenesis, Progression and Treatment of Nonalcoholic Steatohepatitis (NASH): Recent Advances. Prog. Lipid Res. 2023, 91, 101238. [Google Scholar] [CrossRef] [PubMed]
- McGlinchey, A.J.; Govaere, O.; Geng, D.; Ratziu, V.; Allison, M.; Bousier, J.; Petta, S.; de Oliviera, C.; Bugianesi, E.; Schattenberg, J.M.; et al. Metabolic Signatures across the Full Spectrum of Non-Alcoholic Fatty Liver Disease. JHEP Rep. 2022, 4, 100477. [Google Scholar] [CrossRef]
- Paul, B.; Lewinska, M.; Andersen, J.B. Lipid Alterations in Chronic Liver Disease and Liver Cancer. JHEP Rep. 2022, 4, 100479. [Google Scholar] [CrossRef]
- Bertran, L.; Capellades, J.; Abelló, S.; Durán-Bertran, J.; Aguilar, C.; Martinez, S.; Sabench, F.; Correig, X.; Yanes, O.; Auguet, T.; et al. LC/MS-Based Untargeted Metabolomics Study in Women with Nonalcoholic Steatohepatitis Associated with Morbid Obesity. Int. J. Mol. Sci. 2023, 24, 9789. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Y.; Wang, Z.; Zhang, X.; Zhang, Y.; Lin, J.; Du, Y.; Wang, S.; Si, D.; Bao, J.; et al. Integration of Metabolomics, Lipidomics, and Proteomics Reveals the Metabolic Characterization of Nonalcoholic Steatohepatitis. J. Proteome Res. 2023, 22, 2577–2592. [Google Scholar] [CrossRef] [PubMed]
- Bellanti, F.; Villani, R.; Facciorusso, A.; Vendemiale, G.; Serviddio, G. Lipid Oxidation Products in the Pathogenesis of Non-Alcoholic Steatohepatitis. Free Radic. Biol. Med. 2017, 111, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Svegliati-Baroni, G.; Pierantonelli, I.; Torquato, P.; Marinelli, R.; Ferreri, C.; Chatgilialoglu, C.; Bartolini, D.; Galli, F. Lipidomic Biomarkers and Mechanisms of Lipotoxicity in Non-Alcoholic Fatty Liver Disease. Free Radic. Biol. Med. 2019, 144, 293–309. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Ma, X.; Jing, Y.; Shen, M.; Ma, X.; Zhu, J.; Liu, H.; Zhang, G.; Chen, F. Ufmylation on UFBP1 Alleviates Non-Alcoholic Fatty Liver Disease by Modulating Hepatic Endoplasmic Reticulum Stress. Cell Death Dis. 2023, 14, 584. [Google Scholar] [CrossRef] [PubMed]
- Han, C.Y.; Rho, H.S.; Kim, A.; Kim, T.H.; Jang, K.; Jun, D.W.; Kim, J.W.; Kim, B.; Kim, S.G. FXR Inhibits Endoplasmic Reticulum Stress-Induced NLRP3 Inflammasome in Hepatocytes and Ameliorates Liver Injury. Cell Rep. 2018, 24, 2985–2999. [Google Scholar] [CrossRef]
- Latif, M.U.; Schmidt, G.E.; Mercan, S.; Rahman, R.; Gibhardt, C.S.; Stejerean-Todoran, I.; Reutlinger, K.; Hessmann, E.; Singh, S.K.; Moeed, A.; et al. NFATc1 Signaling Drives Chronic ER Stress Responses to Promote NAFLD Progression. Gut 2022, 71, 2561–2573. [Google Scholar] [CrossRef]
- Xia, S.-W.; Wang, Z.-M.; Sun, S.-M.; Su, Y.; Li, Z.-H.; Shao, J.-J.; Tan, S.-Z.; Chen, A.-P.; Wang, S.-J.; Zhang, Z.-L.; et al. Endoplasmic Reticulum Stress and Protein Degradation in Chronic Liver Disease. Pharmacol. Res. 2020, 161, 105218. [Google Scholar] [CrossRef] [PubMed]
- Nassir, F. NAFLD: Mechanisms, Treatments, and Biomarkers. Biomolecules 2022, 12, 824. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Tian, R.; She, Z.; Cai, J.; Li, H. Role of Oxidative Stress in the Pathogenesis of Nonalcoholic Fatty Liver Disease. Free Radic. Biol. Med. 2020, 152, 116–141. [Google Scholar] [CrossRef]
- Petersen, M.C.; Vatner, D.F.; Shulman, G.I. Regulation of Hepatic Glucose Metabolism in Health and Disease. Nat. Rev. Endocrinol. 2017, 13, 572–587. [Google Scholar] [CrossRef]
- Chadt, A.; Al-Hasani, H. Glucose Transporters in Adipose Tissue, Liver, and Skeletal Muscle in Metabolic Health and Disease. Pflugers Arch. 2020, 472, 1273–1298. [Google Scholar] [CrossRef] [PubMed]
- Shearer, A.M.; Wang, Y.; Fletcher, E.K.; Rana, R.; Michael, E.S.; Nguyen, N.; Abdelmalek, M.F.; Covic, L.; Kuliopulos, A. PAR2 Promotes Impaired Glucose Uptake and Insulin Resistance in NAFLD through GLUT2 and Akt Interference. Hepatology 2022, 76, 1778–1793. [Google Scholar] [CrossRef] [PubMed]
- Chun, H.J.; Kim, E.R.; Lee, M.; Choi, D.H.; Kim, S.H.; Shin, E.; Kim, J.-H.; Cho, J.W.; Han, D.H.; Cha, B.-S.; et al. Increased Expression of Sodium-Glucose Cotransporter 2 and O-GlcNAcylation in Hepatocytes Drives Non-Alcoholic Steatohepatitis. Metabolism 2023, 145, 155612. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Sasaki, Y.; Asahiyama, M.; Yano, W.; Takizawa, T.; Kamiya, W.; Matsumura, Y.; Anai, M.; Osawa, T.; Fruchart, J.-C.; et al. Selective PPARα Modulator Pemafibrate and Sodium-Glucose Cotransporter 2 Inhibitor Tofogliflozin Combination Treatment Improved Histopathology in Experimental Mice Model of Non-Alcoholic Steatohepatitis. Cells 2022, 11, 720. [Google Scholar] [CrossRef] [PubMed]
- DeBosch, B.J.; Chen, Z.; Saben, J.L.; Finck, B.N.; Moley, K.H. Glucose Transporter 8 (GLUT8) Mediates Fructose-Induced de Novo Lipogenesis and Macrosteatosis. J. Biol. Chem. 2014, 289, 10989–10998. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Kim, E.; Shin, E.-A.; Shon, J.C.; Sun, H.; Kim, J.E.; Jung, J.W.; Lee, H.; Pinanga, Y.; Song, D.-G.; et al. Crosstalk between TM4SF5 and GLUT8 Regulates Fructose Metabolism in Hepatic Steatosis. Mol. Metab. 2022, 58, 101451. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Um, H.; Park, J.; Jung, J.W.; Kim, J.E.; Lee, H.; Shin, E.-A.; Pinanga, Y.; Lee, H.; Nam, S.H.; et al. TM4SF5-Dependent Crosstalk between Hepatocytes and Macrophages to Reprogram the Inflammatory Environment. Cell Rep. 2021, 37, 110018. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Xin, X.; Ma, J.; Tan, C.; Osna, N.; Mahato, R.I. Therapeutic Targets, Novel Drugs, and Delivery Systems for Diabetes Associated NAFLD and Liver Fibrosis. Adv. Drug Deliv. Rev. 2021, 176, 113888. [Google Scholar] [CrossRef] [PubMed]
- Adeva-Andany, M.M.; González-Lucán, M.; Donapetry-García, C.; Fernández-Fernández, C.; Ameneiros-Rodríguez, E. Glycogen Metabolism in Humans. BBA Clin. 2016, 5, 85–100. [Google Scholar] [CrossRef]
- Soon, G.S.T.; Torbenson, M. The Liver and Glycogen: In Sickness and in Health. Int. J. Mol. Sci. 2023, 24, 6133. [Google Scholar] [CrossRef]
- An, L.; Shi, Q.; Zhu, Y.; Wang, H.; Peng, Q.; Wu, J.; Cheng, Y.; Zhang, W.; Yi, Y.; Bao, Z.; et al. Bone Morphogenetic Protein 4 (BMP4) Promotes Hepatic Glycogen Accumulation and Reduces Glucose Level in Hepatocytes through mTORC2 Signaling Pathway. Genes Dis. 2021, 8, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Thymiakou, E.; Tzardi, M.; Kardassis, D. Impaired Hepatic Glucose Metabolism and Liver-α-Cell Axis in Mice with Liver-Specific Ablation of the Hepatocyte Nuclear Factor 4α (Hnf4a) Gene. Metabolism 2023, 139, 155371. [Google Scholar] [CrossRef] [PubMed]
- Emma, M.R.; Augello, G.; Cusimano, A.; Azzolina, A.; Montalto, G.; McCubrey, J.A.; Cervello, M. GSK-3 in Liver Diseases: Friend or Foe? Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118743. [Google Scholar] [CrossRef] [PubMed]
- Samuel, V.T.; Shulman, G.I. Nonalcoholic Fatty Liver Disease as a Nexus of Metabolic and Hepatic Diseases. Cell Metab. 2018, 27, 22–41. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory Metabolism: Glycolysis, the TCA Cycle and Mitochondrial Electron Transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Hao, L.; Hu, X. Natural Products Target Glycolysis in Liver Disease. Front. Pharmacol. 2023, 14, 1242955. [Google Scholar] [CrossRef]
- Drummer, C.; Saaoud, F.; Jhala, N.C.; Cueto, R.; Sun, Y.; Xu, K.; Shao, Y.; Lu, Y.; Shen, H.; Yang, L.; et al. Caspase-11 Promotes High-Fat Diet-Induced NAFLD by Increasing Glycolysis, OXPHOS, and Pyroptosis in Macrophages. Front. Immunol. 2023, 14, 1113883. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.; Liu, J.; Zhang, D.; Xie, R.; Wang, L.; Hong, J. Glycolysis in Chronic Liver Diseases: Mechanistic Insights and Therapeutic Opportunities. Cells 2023, 12, 1930. [Google Scholar] [CrossRef]
- Dong, T.; Hu, G.; Fan, Z.; Wang, H.; Gao, Y.; Wang, S.; Xu, H.; Yaffe, M.B.; Vander Heiden, M.G.; Lv, G.; et al. Activation of GPR3-β-Arrestin2-PKM2 Pathway in Kupffer Cells Stimulates Glycolysis and Inhibits Obesity and Liver Pathogenesis. Nat. Commun. 2024, 15, 807. [Google Scholar] [CrossRef]
- Bates, J.; Vijayakumar, A.; Ghoshal, S.; Marchand, B.; Yi, S.; Kornyeyev, D.; Zagorska, A.; Hollenback, D.; Walker, K.; Liu, K.; et al. Acetyl-CoA Carboxylase Inhibition Disrupts Metabolic Reprogramming during Hepatic Stellate Cell Activation. J. Hepatol. 2020, 73, 896–905. [Google Scholar] [CrossRef]
- Bruschi, F.V.; Tardelli, M.; Einwallner, E.; Claudel, T.; Trauner, M. PNPLA3 I148M Up-Regulates Hedgehog and Yap Signaling in Human Hepatic Stellate Cells. Int. J. Mol. Sci. 2020, 21, 8711. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Shen, Y.; Meng, Y.; Chen, J.; Zhang, Y.; Zeng, S.; Xu, H. Suv39h1 Contributes to Activation of Hepatic Stellate Cells in Non-Alcoholic Fatty Liver Disease by Enabling Anaerobic Glycolysis. Life Sci. 2024, 341, 122498. [Google Scholar] [CrossRef] [PubMed]
- Fondevila, M.F.; Novoa, E.; Gonzalez-Rellan, M.J.; Fernandez, U.; Heras, V.; Porteiro, B.; Parracho, T.; Dorta, V.; Riobello, C.; da Silva Lima, N.; et al. P63 Controls Metabolic Activation of Hepatic Stellate Cells and Fibrosis via an HER2-ACC1 Pathway. Cell Rep. Med. 2024, 5, 101401. [Google Scholar] [CrossRef] [PubMed]
- Du, D.; Liu, C.; Qin, M.; Zhang, X.; Xi, T.; Yuan, S.; Hao, H.; Xiong, J. Metabolic Dysregulation and Emerging Therapeutical Targets for Hepatocellular Carcinoma. Acta Pharm. Sin. B 2022, 12, 558–580. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, S.; Chen, J.; Su, Z. Unraveling the Regulation of Hepatic Gluconeogenesis. Front. Endocrinol. 2018, 9, 802. [Google Scholar] [CrossRef] [PubMed]
- Wewer Albrechtsen, N.J.; Holst, J.J.; Cherrington, A.D.; Finan, B.; Gluud, L.L.; Dean, E.D.; Campbell, J.E.; Bloom, S.R.; Tan, T.M.-M.; Knop, F.K.; et al. 100 Years of Glucagon and 100 More. Diabetologia 2023, 66, 1378–1394. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.J. Regulation of Hepatic Lipid and Glucose Metabolism by INSP3R1. Diabetes 2022, 71, 1834–1841. [Google Scholar] [CrossRef] [PubMed]
- Vily-Petit, J.; Soty-Roca, M.; Silva, M.; Raffin, M.; Gautier-Stein, A.; Rajas, F.; Mithieux, G. Intestinal Gluconeogenesis Prevents Obesity-Linked Liver Steatosis and Non-Alcoholic Fatty Liver Disease. Gut 2020, 69, 2193–2202. [Google Scholar] [CrossRef] [PubMed]
- Onyango, A.N. Excessive Gluconeogenesis Causes the Hepatic Insulin Resistance Paradox and Its Sequelae. Heliyon 2022, 8, e12294. [Google Scholar] [CrossRef]
- Chen, K.-W.; Chen, Y.-S.; Chen, P.-J.; Yeh, S.-H. Androgen Receptor Functions in Pericentral Hepatocytes to Decrease Gluconeogenesis and Avoid Hyperglycemia and Obesity in Male Mice. Metabolism 2022, 135, 155269. [Google Scholar] [CrossRef]
- Hu, Y.; Li, J.; Li, X.; Wang, D.; Xiang, R.; Liu, W.; Hou, S.; Zhao, Q.; Yu, X.; Xu, M.; et al. Hepatocyte-Secreted FAM3D Ameliorates Hepatic Steatosis by Activating FPR1-hnRNP U-GR-SCAD Pathway to Enhance Lipid Oxidation. Metabolism 2023, 146, 155661. [Google Scholar] [CrossRef] [PubMed]
- Neinast, M.; Murashige, D.; Arany, Z. Branched Chain Amino Acids. Annu. Rev. Physiol. 2019, 81, 139–164. [Google Scholar] [CrossRef] [PubMed]
- Le Couteur, D.G.; Solon-Biet, S.M.; Cogger, V.C.; Ribeiro, R.; de Cabo, R.; Raubenheimer, D.; Cooney, G.J.; Simpson, S.J. Branched Chain Amino Acids, Aging and Age-Related Health. Ageing Res. Rev. 2020, 64, 101198. [Google Scholar] [CrossRef]
- Dimou, A.; Tsimihodimos, V.; Bairaktari, E. The Critical Role of the Branched Chain Amino Acids (BCAAs) Catabolism-Regulating Enzymes, Branched-Chain Aminotransferase (BCAT) and Branched-Chain α-Keto Acid Dehydrogenase (BCKD), in Human Pathophysiology. Int. J. Mol. Sci. 2022, 23, 4022. [Google Scholar] [CrossRef] [PubMed]
- Gaggini, M.; Carli, F.; Rosso, C.; Buzzigoli, E.; Marietti, M.; Della Latta, V.; Ciociaro, D.; Abate, M.L.; Gambino, R.; Cassader, M.; et al. Altered Amino Acid Concentrations in NAFLD: Impact of Obesity and Insulin Resistance. Hepatology 2018, 67, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Chen, R.; Kong, L.; Wei, P.; Liu, Z.; Wang, X.; Hao, H.; Lu, Y.; Hu, W. Effects of Serum Branched-Chain Amino Acids on Nonalcoholic Fatty Liver Disease and Subsequent Cardiovascular Disease. Hepatol. Int. 2022, 16, 1424–1434. [Google Scholar] [CrossRef]
- Grzych, G.; Vonghia, L.; Bout, M.-A.; Weyler, J.; Verrijken, A.; Dirinck, E.; Chevalier Curt, M.J.; Van Gaal, L.; Paumelle, R.; Francque, S.; et al. Plasma BCAA Changes in Patients with NAFLD Are Sex Dependent. J. Clin. Endocrinol. Metab. 2020, 105, 2311–2321. [Google Scholar] [CrossRef]
- Lischka, J.; Schanzer, A.; Hojreh, A.; Ba Ssalamah, A.; Item, C.B.; de Gier, C.; Walleczek, N.-K.; Metz, T.F.; Jakober, I.; Greber-Platzer, S.; et al. A Branched-Chain Amino Acid-Based Metabolic Score Can Predict Liver Fat in Children and Adolescents with Severe Obesity. Pediatr. Obes. 2021, 16, e12739. [Google Scholar] [CrossRef]
- Grenier-Larouche, T.; Coulter Kwee, L.; Deleye, Y.; Leon-Mimila, P.; Walejko, J.M.; McGarrah, R.W.; Marceau, S.; Trahan, S.; Racine, C.; Carpentier, A.C.; et al. Altered Branched-Chain α-Keto Acid Metabolism Is a Feature of NAFLD in Individuals with Severe Obesity. JCI Insight 2022, 7, e159204. [Google Scholar] [CrossRef]
- Bjune, M.S.; Lawrence-Archer, L.; Laupsa-Borge, J.; Sommersten, C.H.; McCann, A.; Glastad, R.C.; Johnston, I.G.; Kern, M.; Blüher, M.; Mellgren, G.; et al. Metabolic Role of the Hepatic Valine/3-Hydroxyisobutyrate (3-HIB) Pathway in Fatty Liver Disease. EBioMedicine 2023, 91, 104569. [Google Scholar] [CrossRef]
- Honda, T.; Ishigami, M.; Luo, F.; Lingyun, M.; Ishizu, Y.; Kuzuya, T.; Hayashi, K.; Nakano, I.; Ishikawa, T.; Feng, G.-G.; et al. Branched-Chain Amino Acids Alleviate Hepatic Steatosis and Liver Injury in Choline-Deficient High-Fat Diet Induced NASH Mice. Metabolism 2017, 69, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhao, S.; Yan, W.; Xia, Y.; Chen, X.; Wang, W.; Zhang, J.; Gao, C.; Peng, C.; Yan, F.; et al. Branched Chain Amino Acids Cause Liver Injury in Obese/Diabetic Mice by Promoting Adipocyte Lipolysis and Inhibiting Hepatic Autophagy. EBioMedicine 2016, 13, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Galarregui, C.; Cantero, I.; Marin-Alejandre, B.A.; Monreal, J.I.; Elorz, M.; Benito-Boillos, A.; Herrero, J.I.; de la O, V.; Ruiz-Canela, M.; Hermsdorff, H.H.M.; et al. Dietary Intake of Specific Amino Acids and Liver Status in Subjects with Nonalcoholic Fatty Liver Disease: Fatty Liver in Obesity (FLiO) Study. Eur. J. Nutr. 2021, 60, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- De Mello, V.D.; Sehgal, R.; Männistö, V.; Klåvus, A.; Nilsson, E.; Perfilyev, A.; Kaminska, D.; Miao, Z.; Pajukanta, P.; Ling, C.; et al. Serum Aromatic and Branched-Chain Amino Acids Associated with NASH Demonstrate Divergent Associations with Serum Lipids. Liver Int. 2021, 41, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.P.; Dupuy, C. Thyroid Hormone Biosynthesis and Release. Mol. Cell Endocrinol. 2017, 458, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.A.; Singh, B.K.; Yen, P.M. Direct Effects of Thyroid Hormones on Hepatic Lipid Metabolism. Nat. Rev. Endocrinol. 2018, 14, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Qin, P.; Li, X.-N.; Wu, J.; Lu, J.; Zhu, W.-F.; Diao, Q.-Q.; Xu, N.-Z.; Zhang, Q. Free Triiodothyronine Is Associated with Hepatic Steatosis and Liver Stiffness in Euthyroid Chinese Adults with Non-Alcoholic Fatty Liver Disease. Front. Endocrinol. 2021, 12, 711956. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Li, L.; Liu, Z.; Cao, L.; Chen, X.; Suo, C.; Zhang, T. The Association between Thyroid Hormones and MAFLD Is Mediated by Obesity and Metabolic Disorders and Varies among MAFLD Subtypes. Dig. Liver Dis. 2023, 55, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Sano, A.; Kakazu, E.; Hamada, S.; Inoue, J.; Ninomiya, M.; Iwata, T.; Tsuruoka, M.; Sato, K.; Masamune, A. Steatotic Hepatocytes Release Mature VLDL Through Methionine and Tyrosine Metabolism in a Keap1-Nrf2-Dependent Manner. Hepatology 2021, 74, 1271–1286. [Google Scholar] [CrossRef]
- Bhave, S.; Ho, H.K. Exploring the Gamut of Receptor Tyrosine Kinases for Their Promise in the Management of Non-Alcoholic Fatty Liver Disease. Biomedicines 2021, 9, 1776. [Google Scholar] [CrossRef]
- Ma, Q.; Chen, J.; Zhou, X.; Hu, L.; Sun, Y.; Wang, Z.; Yue, Z.; Shan, A. Dietary Supplementation with Aromatic Amino Acids Decreased Triglycerides and Alleviated Hepatic Steatosis by Stimulating Bile Acid Synthesis in Mice. Food Funct. 2021, 12, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.-H.; Xin, F.-Z.; Xue, Y.; Hu, Z.; Han, Y.; Ma, F.; Zhou, D.; Liu, X.-L.; Cui, A.; Liu, Z.; et al. Indole-3-Propionic Acid Inhibits Gut Dysbiosis and Endotoxin Leakage to Attenuate Steatohepatitis in Rats. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.-B. Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. Int. J. Tryptophan Res. 2017, 10, 1178646917691938. [Google Scholar] [CrossRef] [PubMed]
- Blaner, W.S.; Li, Y.; Brun, P.-J.; Yuen, J.J.; Lee, S.-A.; Clugston, R.D. Vitamin A Absorption, Storage and Mobilization. Subcell. Biochem. 2016, 81, 95–125. [Google Scholar] [CrossRef]
- Blaner, W.S. Vitamin A Signaling and Homeostasis in Obesity, Diabetes, and Metabolic Disorders. Pharmacol. Ther. 2019, 197, 153–178. [Google Scholar] [CrossRef] [PubMed]
- Carazo, A.; Macáková, K.; Matoušová, K.; Krčmová, L.K.; Protti, M.; Mladěnka, P. Vitamin A Update: Forms, Sources, Kinetics, Detection, Function, Deficiency, Therapeutic Use and Toxicity. Nutrients 2021, 13, 1703. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.; Bartuzi, P.; Heegsma, J.; Dekker, D.; Kloosterhuis, N.; de Bruin, A.; Jonker, J.W.; van de Sluis, B.; Faber, K.N. Impaired Hepatic Vitamin A Metabolism in NAFLD Mice Leading to Vitamin A Accumulation in Hepatocytes. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 309–325.e3. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Liu, Y.; He, S.; Zeng, J.; Li, P.; Ma, C.; Ping, F.; Zhang, H.; Xu, L.; Li, W.; et al. Negative Association between Antioxidant Vitamin Intake and Non-Alcoholic Fatty Liver Disease in Chinese Non-Diabetic Adults: Mediation Models Involving Superoxide Dismutase. Free Radic. Res. 2020, 54, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Coelho, J.M.; Cansanção, K.; Perez, R.d.M.; Leite, N.C.; Padilha, P.; Ramalho, A.; Peres, W. Association between Serum and Dietary Antioxidant Micronutrients and Advanced Liver Fibrosis in Non-Alcoholic Fatty Liver Disease: An Observational Study. PeerJ 2020, 8, e9838. [Google Scholar] [CrossRef]
- Jeon, D.; Son, M.; Shim, J. Dynamics of Serum Retinol and Alpha-Tocopherol Levels According to Non-Alcoholic Fatty Liver Disease Status. Nutrients 2021, 13, 1720. [Google Scholar] [CrossRef]
- Xiao, M.-L.; Zhong, H.-L.; Lin, H.-R.; Liu, C.-Y.; Yan, Y.; Ke, Y.-B.; Chen, Y.-M. Higher Serum Vitamin A Is Associated with a Worsened Progression of Non-Alcoholic Fatty Liver Disease in Adults: A Prospective Study. Food Funct. 2022, 13, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Nulali, J.; Zhang, C.; Chen, Y.; Cheng, J.; Shi, X.; Zhu, C.; Lu, Y.; Wang, N.; Han, B. The Association between Serum Vitamin A and NAFLD among US Adults Varied in Different BMI Groups: A Cross-Sectional Study. Food Funct. 2023, 14, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.; Dullaart, R.P.F.; Schreuder, T.C.M.A.; Blokzijl, H.; Faber, K.N. Disturbed Vitamin A Metabolism in Non-Alcoholic Fatty Liver Disease (NAFLD). Nutrients 2017, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Chen, G. The Interactions of Insulin and Vitamin A Signaling Systems for the Regulation of Hepatic Glucose and Lipid Metabolism. Cells 2021, 10, 2160. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.-H.; Melis, M.; Lu, C.; Rappa, A.; Zhang, T.; Jessurun, J.; Gross, S.S.; Gudas, L.J. A Retinoic Acid Receptor Β2 Agonist Attenuates Transcriptome and Metabolome Changes Underlying Nonalcohol-Associated Fatty Liver Disease. J. Biol. Chem. 2021, 297, 101331. [Google Scholar] [CrossRef]
- Cassim Bawa, F.N.; Hu, S.; Gopoju, R.; Shiyab, A.; Mongan, K.; Xu, Y.; Pan, X.; Clark, A.; Wang, H.; Zhang, Y. Adipocyte Retinoic Acid Receptor α Prevents Obesity and Steatohepatitis by Regulating Energy Expenditure and Lipogenesis. Obesity 2024, 32, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Hanna, M.; Jaqua, E.; Nguyen, V.; Clay, J. B Vitamins: Functions and Uses in Medicine. Perm. J. 2022, 26, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Kalyesubula, M.; Mopuri, R.; Asiku, J.; Rosov, A.; Yosefi, S.; Edery, N.; Bocobza, S.; Moallem, U.; Dvir, H. High-Dose Vitamin B1 Therapy Prevents the Development of Experimental Fatty Liver Driven by Overnutrition. Dis. Model. Mech. 2021, 14, dmm048355. [Google Scholar] [CrossRef] [PubMed]
- Masschelin, P.M.; Saha, P.; Ochsner, S.A.; Cox, A.R.; Kim, K.H.; Felix, J.B.; Sharp, R.; Li, X.; Tan, L.; Park, J.H.; et al. Vitamin B2 Enables Regulation of Fasting Glucose Availability. eLife 2023, 12, e84077. [Google Scholar] [CrossRef]
- Kirkland, J.B.; Meyer-Ficca, M.L. Niacin. Adv. Food Nutr. Res. 2018, 83, 83–149. [Google Scholar] [CrossRef]
- Linder, K.; Willmann, C.; Kantartzis, K.; Machann, J.; Schick, F.; Graf, M.; Kümmerle, S.; Häring, H.-U.; Fritsche, A.; Stefan, N.; et al. Dietary Niacin Intake Predicts the Decrease of Liver Fat Content During a Lifestyle Intervention. Sci. Rep. 2019, 9, 1303. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Zhou, Y.; Pang, N.; Yang, L. Dietary Niacin Intake and Mortality among Individuals with Nonalcoholic Fatty Liver Disease. JAMA Netw. Open 2024, 7, e2354277. [Google Scholar] [CrossRef] [PubMed]
- Ganji, S.H.; Kashyap, M.L.; Kamanna, V.S. Niacin Inhibits Fat Accumulation, Oxidative Stress, and Inflammatory Cytokine IL-8 in Cultured Hepatocytes: Impact on Non-Alcoholic Fatty Liver Disease. Metabolism 2015, 64, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Lezama, I.; Fuentes-Figueroa, M.Á.; Ramos-Tovar, E.; Márquez-Quiroga, L.V.; Ortiz-Fernández, A.; Vargas-Pozada, E.E.; Arellanes-Robledo, J.; Tsutsumi, V.; Muriel, P. Nicotinic Acid Attenuates Experimental Non-Alcoholic Steatohepatitis by Inhibiting the NLRP3 Inflammasome/Pyroptosis Pathway. Biochem. Pharmacol. 2023, 216, 115762. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Cao, Z.; Lai, X.; Shi, Y.; Zhou, N. Niacin Ameliorates Hepatic Steatosis by Inhibiting De Novo Lipogenesis Via a GPR109A-Mediated PKC-ERK1/2-AMPK Signaling Pathway in C57BL/6 Mice Fed a High-Fat Diet. J. Nutr. 2020, 150, 672–684. [Google Scholar] [CrossRef] [PubMed]
- Mejía, S.Á.; Gutman, L.A.B.; Camarillo, C.O.; Navarro, R.M.; Becerra, M.C.S.; Santana, L.D.; Cruz, M.; Pérez, E.H.; Flores, M.D. Nicotinamide Prevents Sweet Beverage-Induced Hepatic Steatosis in Rats by Regulating the G6PD, NADPH/NADP+ and GSH/GSSG Ratios and Reducing Oxidative and Inflammatory Stress. Eur. J. Pharmacol. 2018, 818, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Loza-Medrano, S.S.; Baiza-Gutman, L.A.; Manuel-Apolinar, L.; García-Macedo, R.; Damasio-Santana, L.; Martínez-Mar, O.A.; Sánchez-Becerra, M.C.; Cruz-López, M.; Ibáñez-Hernández, M.A.; Díaz-Flores, M. High Fructose-Containing Drinking Water-Induced Steatohepatitis in Rats Is Prevented by the Nicotinamide-Mediated Modulation of Redox Homeostasis and NADPH-Producing Enzymes. Mol. Biol. Rep. 2020, 47, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.N.G.d.S.; Silvares, R.R.; Flores, E.E.I.; Rodrigues, K.L.; Daliry, A. Pyridoxamine Improves Metabolic and Microcirculatory Complications Associated with Nonalcoholic Fatty Liver Disease. Microcirculation 2020, 27, e12603. [Google Scholar] [CrossRef] [PubMed]
- Ferro, Y.; Carè, I.; Mazza, E.; Provenzano, F.; Colica, C.; Torti, C.; Romeo, S.; Pujia, A.; Montalcini, T. Protein and Vitamin B6 Intake Are Associated with Liver Steatosis Assessed by Transient Elastography, Especially in Obese Individuals. Clin. Mol. Hepatol. 2017, 23, 249–259. [Google Scholar] [CrossRef]
- Yuan, S.; Chen, J.; Dan, L.; Xie, Y.; Sun, Y.; Li, X.; Larsson, S.C. Homocysteine, Folate, and Nonalcoholic Fatty Liver Disease: A Systematic Review with Meta-Analysis and Mendelian Randomization Investigation. Am. J. Clin. Nutr. 2022, 116, 1595–1609. [Google Scholar] [CrossRef]
- Chen, H.-K.; Luo, J.; Li, X.-J.; Liao, W.-Z.; Hu, Y.-Q.; Guo, X.-G. Serum Folate Associated with Nonalcoholic Fatty Liver Disease and Advanced Hepatic Fibrosis. Sci. Rep. 2023, 13, 12933. [Google Scholar] [CrossRef] [PubMed]
- Sid, V.; Siow, Y.L.; Karmin, O. Role of Folate in Nonalcoholic Fatty Liver Disease. Can. J. Physiol. Pharmacol. 2017, 95, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Mardinoglu, A.; Wu, H.; Bjornson, E.; Zhang, C.; Hakkarainen, A.; Räsänen, S.M.; Lee, S.; Mancina, R.M.; Bergentall, M.; Pietiläinen, K.H.; et al. An Integrated Understanding of the Rapid Metabolic Benefits of a Carbohydrate-Restricted Diet on Hepatic Steatosis in Humans. Cell Metab. 2018, 27, 559–571.e5. [Google Scholar] [CrossRef] [PubMed]
- Bakir, M.B.; Salama, M.A.; Refaat, R.; Ali, M.A.; Khalifa, E.A.; Kamel, M.A. Evaluating the Therapeutic Potential of One-Carbon Donors in Nonalcoholic Fatty Liver Disease. Eur. J. Pharmacol. 2019, 847, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shen, J.; Yang, X.; Sun, Q.; Yang, X. Folic Acid Reduced Triglycerides Deposition in Primary Chicken Hepatocytes. J. Agric. Food Chem. 2018, 66, 13162–13172. [Google Scholar] [CrossRef] [PubMed]
- Salman, M.; Kamel, M.A.; El-Nabi, S.E.H.; Ismail, A.H.A.; Ullah, S.; Al-Ghamdi, A.; Hathout, H.M.R.; El-Garawani, I.M. The Regulation of HBP1, SIRT1, and SREBP-1c Genes and the Related microRNAs in Non-Alcoholic Fatty Liver Rats: The Association with the Folic Acid Anti-Steatosis. PLoS ONE 2022, 17, e0265455. [Google Scholar] [CrossRef] [PubMed]
- Xin, F.-Z.; Zhao, Z.-H.; Zhang, R.-N.; Pan, Q.; Gong, Z.-Z.; Sun, C.; Fan, J.-G. Folic Acid Attenuates High-Fat Diet-Induced Steatohepatitis via Deacetylase SIRT1-Dependent Restoration of PPARα. World J. Gastroenterol. 2020, 26, 2203–2220. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zheng, B.; Xu, S.; Zhao, Z.; Liu, W.; Wang, T.; Yuan, M.; Sun, X.; Tan, Y.; Xu, Q.; et al. Mitochondrial Folate Metabolism-Mediated α-Linolenic Acid Exhaustion Masks Liver Fibrosis Resolution. J. Biol. Chem. 2023, 299, 104909. [Google Scholar] [CrossRef] [PubMed]
- Boachie, J.; Zammit, V.; Saravanan, P.; Adaikalakoteswari, A. Metformin Inefficiency to Lower Lipids in Vitamin B12 Deficient HepG2 Cells Is Alleviated via Adiponectin-AMPK Axis. Nutrients 2023, 15, 5046. [Google Scholar] [CrossRef]
- Tripathi, M.; Singh, B.K.; Zhou, J.; Tikno, K.; Widjaja, A.; Sandireddy, R.; Arul, K.; Abdul Ghani, S.A.B.; Bee, G.G.B.; Wong, K.A.; et al. Vitamin B12 and Folate Decrease Inflammation and Fibrosis in NASH by Preventing Syntaxin 17 Homocysteinylation. J. Hepatol. 2022, 77, 1246–1255. [Google Scholar] [CrossRef]
- Talari, H.R.; Molaqanbari, M.R.; Mokfi, M.; Taghizadeh, M.; Bahmani, F.; Tabatabaei, S.M.H.; Sharifi, N. The Effects of Vitamin B12 Supplementation on Metabolic Profile of Patients with Non-Alcoholic Fatty Liver Disease: A Randomized Controlled Trial. Sci. Rep. 2022, 12, 14047. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Zhang, W.; He, Z.; Yang, H.; Gao, J.; Wu, P.; Ma, Z.F. Dietary Vitamin C Intake Is Associated with Improved Liver Function and Glucose Metabolism in Chinese Adults. Front. Nutr. 2021, 8, 779912. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.-Q.; Li, H.-X.; Tan, W.-L.; Yang, L.; Ma, X.-W.; Li, W.-X.; Wang, Q.-B.; Shang, C.-Z.; Chen, Y.-J. Association of Serum Vitamin C with NAFLD and MAFLD among Adults in the United States. Front. Nutr. 2021, 8, 795391. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Li, Y.; Ma, B.; Lei, S.; Wang, X. Iron Metabolism Mediates the Relationship between Vitamin C and Hepatic Steatosis and Fibrosis in NAFLD. Front. Nutr. 2022, 9, 952056. [Google Scholar] [CrossRef] [PubMed]
- Ipsen, D.H.; Tveden-Nyborg, P.; Lykkesfeldt, J. Does Vitamin C Deficiency Promote Fatty Liver Disease Development? Nutrients 2014, 6, 5473–5499. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Ahn, J.; Shin, S.S.; Yoon, M. Ascorbic Acid Inhibits Visceral Obesity and Nonalcoholic Fatty Liver Disease by Activating Peroxisome Proliferator-Activated Receptor α in High-Fat-Diet-Fed C57BL/6J Mice. Int. J. Obes. 2019, 43, 1620–1630. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Zhao, L.; Meng, C.; Zhao, X.; Liu, Y.; Shi, R.; Han, X.; Wang, T.; Li, J. Prophylactic and Therapeutic Effects of Different Doses of Vitamin C on High-Fat-Diet-Induced Non-Alcoholic Fatty Liver Disease in Mice. Biomed. Pharmacother. 2020, 131, 110792. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-W.; Lee, Y.-J.; Baek, S.-M.; Kang, K.-K.; Kim, T.-U.; Yim, J.-H.; Kim, H.-Y.; Han, S.-H.; Choi, S.-K.; Park, S.-J.; et al. Mega-Dose Vitamin C Ameliorates Nonalcoholic Fatty Liver Disease in a Mouse Fast-Food Diet Model. Nutrients 2022, 14, 2195. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Luo, X.; Wang, Y.; He, Z.; Li, X.; Wu, K.; Zhang, Y.; Yang, Y.; Ji, J.; Luo, X. Ascorbic Acid Attenuates Cell Stress by Activating the Fibroblast Growth Factor 21/Fibroblast Growth Factor Receptor 2/Adiponectin Pathway in HepG2 Cells. Mol. Med. Rep. 2019, 20, 2450–2458. [Google Scholar] [CrossRef]
- Lee, S.-W.; Baek, S.-M.; Kang, K.-K.; Lee, A.-R.; Kim, T.-U.; Choi, S.-K.; Roh, Y.-S.; Hong, I.-H.; Park, S.-J.; Kim, T.-H.; et al. Vitamin C Deficiency Inhibits Nonalcoholic Fatty Liver Disease Progression through Impaired de Novo Lipogenesis. Am. J. Pathol. 2021, 191, 1550–1563. [Google Scholar] [CrossRef]
- Scorletti, E.; Creasy, K.T.; Vujkovic, M.; Vell, M.; Zandvakili, I.; Rader, D.J.; Schneider, K.M.; Schneider, C.V. Dietary Vitamin E Intake Is Associated with a Reduced Risk of Developing Digestive Diseases and Nonalcoholic Fatty Liver Disease. Am. J. Gastroenterol. 2022, 117, 927–930. [Google Scholar] [CrossRef]
- Qi, X.; Guo, J.; Li, Y.; Fang, C.; Lin, J.; Chen, X.; Jia, J. Vitamin E Intake Is Inversely Associated with NAFLD Measured by Liver Ultrasound Transient Elastography. Sci. Rep. 2024, 14, 2592. [Google Scholar] [CrossRef] [PubMed]
- Podszun, M.C.; Frank, J. Impact of Vitamin E on Redox Biomarkers in Non-Alcoholic Fatty Liver Disease. Redox Biol. 2021, 42, 101937. [Google Scholar] [CrossRef] [PubMed]
- Nagashimada, M.; Ota, T. Role of Vitamin E in Nonalcoholic Fatty Liver Disease. IUBMB Life 2019, 71, 516–522. [Google Scholar] [CrossRef]
- Barchetta, I.; Cimini, F.A.; Cavallo, M.G. Vitamin D and Metabolic Dysfunction-Associated Fatty Liver Disease (MAFLD): An Update. Nutrients 2020, 12, 3302. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Song, Y.; Wang, Y.; Hong, H. Vitamin D/Vitamin D Receptor Pathway in Non-Alcoholic Fatty Liver Disease. Expert Opin. Ther. Targets 2023, 27, 1145–1157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Moon, R.; Thorne, J.L.; Moore, J.B. NAFLD and Vitamin D: Evidence for Intersection of microRNA-Regulated Pathways. Nutr. Res. Rev. 2023, 36, 120–139. [Google Scholar] [CrossRef]
- Yuan, S.; Larsson, S.C. Inverse Association Between Serum 25-Hydroxyvitamin D and Nonalcoholic Fatty Liver Disease. Clin. Gastroenterol. Hepatol. 2023, 21, 398–405.e4. [Google Scholar] [CrossRef] [PubMed]
- Du, T.; Xiang, L.; Zhang, J.; Yang, C.; Zhao, W.; Li, J.; Zhou, Y.; Ma, L. Vitamin D Improves Hepatic Steatosis in NAFLD via Regulation of Fatty Acid Uptake and β-Oxidation. Front. Endocrinol. 2023, 14, 1138078. [Google Scholar] [CrossRef]
- Zhang, J.-J.; Yu, H.-C.; Li, Y.; Zhang, Y.-B.; Geng, T.-T.; Lu, Q.; Liao, Y.-F.; Guo, K.-Q.; Du, L.; Ruan, H.-L.; et al. Association between Serum 25-Hydroxy Vitamin D Concentrations and Mortality among Individuals with Metabolic Dysfunction-Associated Fatty Liver Disease: A Prospective Cohort Study. Am. J. Clin. Nutr. 2022, 116, 1409–1417. [Google Scholar] [CrossRef]
- Reda, D.; Elshopakey, G.E.; Albukhari, T.A.; Almehmadi, S.J.; Refaat, B.; Risha, E.F.; Mahgoub, H.A.; El-Boshy, M.E.; Abdelhamid, F.M. Vitamin D3 Alleviates Nonalcoholic Fatty Liver Disease in Rats by Inhibiting Hepatic Oxidative Stress and Inflammation via the SREBP-1-c/PPARα-NF-κB/IR-S2 Signaling Pathway. Front. Pharmacol. 2023, 14, 1164512. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Jeong, H.-J.; Lee, Y.J.; Park, H.Y.; Koo, S.K.; Lim, J.H. Vitamin D Ameliorates Age-Induced Nonalcoholic Fatty Liver Disease by Increasing the Mitochondrial Contact Site and Cristae Organizing System (MICOS) 60 Level. Exp. Mol. Med. 2024, 56, 142–155. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-L.; Chen, L.; Yang, J.; Zhao, S.-S.; Jin, S.; Ao, N.; Yang, J.; Liu, H.-X.; Du, J. Vitamin D Alleviates Non-Alcoholic Fatty Liver Disease via Restoring Gut Microbiota and Metabolism. Front. Microbiol. 2023, 14, 1117644. [Google Scholar] [CrossRef]
- Bogdan, A.R.; Miyazawa, M.; Hashimoto, K.; Tsuji, Y. Regulators of Iron Homeostasis: New Players in Metabolism, Cell Death, and Disease. Trends Biochem. Sci. 2016, 41, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, Y.; Zhang, Z.; Xie, J.; Yu, C.; Xu, L.; Li, Y. Iron Status and NAFLD among European Populations: A Bidirectional Two-Sample Mendelian Randomization Study. Nutrients 2022, 14, 5237. [Google Scholar] [CrossRef] [PubMed]
- Valenti, L.; Corradini, E.; Adams, L.A.; Aigner, E.; Alqahtani, S.; Arrese, M.; Bardou-Jacquet, E.; Bugianesi, E.; Fernandez-Real, J.-M.; Girelli, D.; et al. Consensus Statement on the Definition and Classification of Metabolic Hyperferritinaemia. Nat. Rev. Endocrinol. 2023, 19, 299–310. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Song, G.; Xing, H.; Yang, L.; Han, K.; Chang, Y.-Z. A High-Fructose Diet in Rats Induces Systemic Iron Deficiency and Hepatic Iron Overload by an Inflammation Mechanism. J. Food Biochem. 2021, 45, e13578. [Google Scholar] [CrossRef] [PubMed]
- Altamura, S.; Müdder, K.; Schlotterer, A.; Fleming, T.; Heidenreich, E.; Qiu, R.; Hammes, H.-P.; Nawroth, P.; Muckenthaler, M.U. Iron Aggravates Hepatic Insulin Resistance in the Absence of Inflammation in a Novel Db/Db Mouse Model with Iron Overload. Mol. Metab. 2021, 51, 101235. [Google Scholar] [CrossRef]
- Li, H.; Hu, L.; Wang, L.; Wang, Y.; Shao, M.; Chen, Y.; Wu, W.; Wang, L. Iron Activates cGAS-STING Signaling and Promotes Hepatic Inflammation. J. Agric. Food Chem. 2022, 70, 2211–2220. [Google Scholar] [CrossRef]
- Gao, H.; Jin, Z.; Bandyopadhyay, G.; Wang, G.; Zhang, D.; Rocha, K.C.E.; Liu, X.; Zhao, H.; Kisseleva, T.; Brenner, D.A.; et al. Aberrant Iron Distribution via Hepatocyte-Stellate Cell Axis Drives Liver Lipogenesis and Fibrosis. Cell Metab. 2022, 34, 1201–1213.e5. [Google Scholar] [CrossRef]
- Tsuchiya, H.; Ebata, Y.; Sakabe, T.; Hama, S.; Kogure, K.; Shiota, G. High-Fat, High-Fructose Diet Induces Hepatic Iron Overload via a Hepcidin-Independent Mechanism Prior to the Onset of Liver Steatosis and Insulin Resistance in Mice. Metabolism 2013, 62, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An Iron-Dependent Form of Nonapoptotic Cell Death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Byrne, C.D.; Targher, G.; Wang, F.; Zheng, M.-H. Ferroptosis and Metabolic Dysfunction-Associated Fatty Liver Disease: Is There a Link? Liver Int. 2022, 42, 1496–1502. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, Z.; Geng, J.; Li, L.; Feng, X. An Overview of Ferroptosis in Non-Alcoholic Fatty Liver Disease. Biomed. Pharmacother. 2022, 153, 113374. [Google Scholar] [CrossRef] [PubMed]
- Tsurusaki, S.; Tsuchiya, Y.; Koumura, T.; Nakasone, M.; Sakamoto, T.; Matsuoka, M.; Imai, H.; Yuet-Yin Kok, C.; Okochi, H.; Nakano, H.; et al. Hepatic Ferroptosis Plays an Important Role as the Trigger for Initiating Inflammation in Nonalcoholic Steatohepatitis. Cell Death Dis. 2019, 10, 449. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Minikes, A.M.; Jiang, X. Ferroptosis at the Intersection of Lipid Metabolism and Cellular Signaling. Mol. Cell 2022, 82, 2215–2227. [Google Scholar] [CrossRef] [PubMed]
- Pope, L.E.; Dixon, S.J. Regulation of Ferroptosis by Lipid Metabolism. Trends Cell Biol. 2023, 33, 1077–1087. [Google Scholar] [CrossRef]
- Stockwell, B.R. Ferroptosis Turns 10: Emerging Mechanisms, Physiological Functions, and Therapeutic Applications. Cell 2022, 185, 2401–2421. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Stockwell, B.R.; Conrad, M. Ferroptosis: Mechanisms, Biology and Role in Disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 266–282. [Google Scholar] [CrossRef]
- Li, Y. Copper Homeostasis: Emerging Target for Cancer Treatment. IUBMB Life 2020, 72, 1900–1908. [Google Scholar] [CrossRef]
- Morrell, A.; Tallino, S.; Yu, L.; Burkhead, J.L. The Role of Insufficient Copper in Lipid Synthesis and Fatty-Liver Disease. IUBMB Life 2017, 69, 263–270. [Google Scholar] [CrossRef]
- Song, M.; Vos, M.B.; McClain, C.J. Copper-Fructose Interactions: A Novel Mechanism in the Pathogenesis of NAFLD. Nutrients 2018, 10, 1815. [Google Scholar] [CrossRef] [PubMed]
- Aigner, E.; Strasser, M.; Haufe, H.; Sonnweber, T.; Hohla, F.; Stadlmayr, A.; Solioz, M.; Tilg, H.; Patsch, W.; Weiss, G.; et al. A Role for Low Hepatic Copper Concentrations in Nonalcoholic Fatty Liver Disease. Am. J. Gastroenterol. 2010, 105, 1978–1985. [Google Scholar] [CrossRef]
- Song, M.; Schuschke, D.A.; Zhou, Z.; Chen, T.; Pierce, W.M.; Wang, R.; Johnson, W.T.; McClain, C.J. High Fructose Feeding Induces Copper Deficiency in Sprague-Dawley Rats: A Novel Mechanism for Obesity Related Fatty Liver. J. Hepatol. 2012, 56, 433–440. [Google Scholar] [CrossRef]
- Heffern, M.C.; Park, H.M.; Au-Yeung, H.Y.; Van de Bittner, G.C.; Ackerman, C.M.; Stahl, A.; Chang, C.J. In Vivo Bioluminescence Imaging Reveals Copper Deficiency in a Murine Model of Nonalcoholic Fatty Liver Disease. Proc. Natl. Acad. Sci. USA 2016, 113, 14219–14224. [Google Scholar] [CrossRef]
- Jiang, X.; Hu, R.; Huang, Y.; Xu, Y.; Zheng, Z.; Shi, Y.; Miao, J.; Liu, Y. Fructose Aggravates Copper-Deficiency-Induced Non-Alcoholic Fatty Liver Disease. J. Nutr. Biochem. 2023, 119, 109402. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Yuan, Y.; Xu, S.; Lu, S.; Gu, J.; Wang, Y.; Wang, Y.; Zhang, X.; Chen, S.; Li, J.; et al. Downregulation of Hepatic Ceruloplasmin Ameliorates NAFLD via SCO1-AMPK-LKB1 Complex. Cell Rep. 2022, 41, 111498. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Wang, N.; Lu, S.; Xiong, J.; Yuan, Y.; Liu, J.; Chen, S. Targeting Hepatic Ceruloplasmin Mitigates Nonalcoholic Steatohepatitis by Modulating Bile Acid Metabolism. J. Mol. Cell Biol. 2023, 15, mjad060. [Google Scholar] [CrossRef]
- Zhong, C.-C.; Zhao, T.; Hogstrand, C.; Chen, F.; Song, C.-C.; Luo, Z. Copper (Cu) Induced Changes of Lipid Metabolism through Oxidative Stress-Mediated Autophagy and Nrf2/PPARγ Pathways. J. Nutr. Biochem. 2022, 100, 108883. [Google Scholar] [CrossRef]
- Zhong, C.-C.; Zhao, T.; Hogstrand, C.; Song, C.-C.; Zito, E.; Tan, X.-Y.; Xu, Y.-C.; Song, Y.-F.; Wei, X.-L.; Luo, Z. Copper Induces Liver Lipotoxicity Disease by Up-Regulating Nrf2 Expression via the Activation of MTF-1 and Inhibition of SP1/Fyn Pathway. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166752. [Google Scholar] [CrossRef]
- Tsvetkov, P.; Coy, S.; Petrova, B.; Dreishpoon, M.; Verma, A.; Abdusamad, M.; Rossen, J.; Joesch-Cohen, L.; Humeidi, R.; Spangler, R.D.; et al. Copper Induces Cell Death by Targeting Lipoylated TCA Cycle Proteins. Science 2022, 375, 1254–1261. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Chen, X.; Kroemer, G. Cuproptosis: A Copper-Triggered Modality of Mitochondrial Cell Death. Cell Res. 2022, 32, 417–418. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.-J.; He, R.-R. Cuproptosis: Copper-Induced Regulated Cell Death. Sci. China Life Sci. 2022, 65, 1680–1682. [Google Scholar] [CrossRef]
- Zhao, Z.; Luan, T.; Wan, J.; Du, H.; Hu, J.; Liu, H.; Gong, X.; Kuang, G.; Wang, B. Elucidating Cuproptosis-Associated Genes in the Progression from Nash to HCC Using Bulk and Single-Cell RNA Sequencing Analyses and Experimental Validation. Medicina 2023, 59, 1639. [Google Scholar] [CrossRef]
- Ouyang, G.; Wu, Z.; Liu, Z.; Pan, G.; Wang, Y.; Liu, J.; Guo, J.; Liu, T.; Huang, G.; Zeng, Y.; et al. Identification and Validation of Potential Diagnostic Signature and Immune Cell Infiltration for NAFLD Based on Cuproptosis-Related Genes by Bioinformatics Analysis and Machine Learning. Front. Immunol. 2023, 14, 1251750. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Fang, Z.; Yang, K.; Ji, Y.; Yu, X.; Guo, Z.; Dong, Z.; Zhu, T.; Liu, C. Identification and Validation of Cuproptosis-Related Molecular Clusters in Non-Alcoholic Fatty Liver Disease. J. Cell Mol. Med. 2024, 28, e18091. [Google Scholar] [CrossRef]
- Himoto, T.; Masaki, T. Associations between Zinc Deficiency and Metabolic Abnormalities in Patients with Chronic Liver Disease. Nutrients 2018, 10, 88. [Google Scholar] [CrossRef] [PubMed]
- Asprouli, E.; Kalafati, I.P.; Sakellari, A.; Karavoltsos, S.; Vlachogiannakos, J.; Revenas, K.; Kokkinos, A.; Dassenakis, M.; Dedoussis, G.V.; Kalogeropoulos, N. Evaluation of Plasma Trace Elements in Different Stages of Nonalcoholic Fatty Liver Disease. Biol. Trace Elem. Res. 2019, 188, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Ishigami, M.; Ishizu, Y.; Kuzuya, T.; Honda, T.; Ishikawa, T.; Toyoda, H.; Kumada, T.; Fujishiro, M. Correlation of Serum Zinc Levels with Pathological and Laboratory Findings in Patients with Nonalcoholic Fatty Liver Disease. Eur. J. Gastroenterol. Hepatol. 2020, 32, 748–753. [Google Scholar] [CrossRef]
- Kim, M.C.; Lee, J.I.; Kim, J.H.; Kim, H.J.; Cho, Y.K.; Jeon, W.K.; Kim, B.I.; Sohn, W. Serum Zinc Level and Hepatic Fibrosis in Patients with Nonalcoholic Fatty Liver Disease. PLoS ONE 2020, 15, e0240195. [Google Scholar] [CrossRef]
- Katayama, K. Zinc and Protein Metabolism in Chronic Liver Diseases. Nutr. Res. 2020, 74, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Zhang, Z.; Liu, S.; Aluo, Z.; Zhang, L.; Yu, L.; Li, Y.; Song, Z.; Zhou, L. Zinc Supplementation Alleviates Lipid and Glucose Metabolic Disorders Induced by a High-Fat Diet. J. Agric. Food Chem. 2020, 68, 5189–5200. [Google Scholar] [CrossRef] [PubMed]
- Bolatimi, O.E.; Head, K.Z.; Luo, J.; Gripshover, T.C.; Lin, Q.; Adiele, N.V.; Watson, W.H.; Wilkerson, C.; Cai, L.; Cave, M.C.; et al. Can Zinc Supplementation Attenuate High Fat Diet-Induced Non-Alcoholic Fatty Liver Disease? Int. J. Mol. Sci. 2023, 24, 1763. [Google Scholar] [CrossRef] [PubMed]
- Fathi, M.; Alavinejad, P.; Haidari, Z.; Amani, R. The Effect of Zinc Supplementation on Steatosis Severity and Liver Function Enzymes in Overweight/Obese Patients with Mild to Moderate Non-Alcoholic Fatty Liver Following Calorie-Restricted Diet: A Double-Blind, Randomized Placebo-Controlled Trial. Biol. Trace Elem. Res. 2020, 197, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Loft, A.; Alfaro, A.J.; Schmidt, S.F.; Pedersen, F.B.; Terkelsen, M.K.; Puglia, M.; Chow, K.K.; Feuchtinger, A.; Troullinaki, M.; Maida, A.; et al. Liver-Fibrosis-Activated Transcriptional Networks Govern Hepatocyte Reprogramming and Intra-Hepatic Communication. Cell Metab. 2021, 33, 1685–1700.e9. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gao, L.; Jiang, C.; Chen, J.; Qin, Z.; Zhong, F.; Yan, Y.; Tong, R.; Zhou, M.; Yuan, A.; et al. The Transcription Factor Zinc Fingers and Homeoboxes 2 Alleviates NASH by Transcriptional Activation of Phosphatase and Tensin Homolog. Hepatology 2022, 75, 939–954. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Sun, N.; Du, H.; He, Y.; Pan, K.; Liu, X.; Lu, X.; Wei, J.; Liao, M.; Duan, C. Mouse Promyelocytic Leukemia Zinc Finger Protein (PLZF) Regulates Hepatic Lipid and Glucose Homeostasis Dependent on SIRT1. Front. Pharmacol. 2022, 13, 1039726. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.; Shi, Y.; Zhang, H.; Zhangyuan, G.; Wang, F.; Li, S.; Chen, C.; Zhang, J.; Wang, H.; Zhang, W.; et al. A TNFα/Miz1-Positive Feedback Loop Inhibits Mitophagy in Hepatocytes and Propagates Non-Alcoholic Steatohepatitis. J. Hepatol. 2023, 79, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, J.; Yang, X.; Shen, C.; Huang, J.; Zhang, D.; Liu, N.; Liu, C.; Zhong, Y.; Chen, Y.; et al. Hepatic Zbtb18 (Zinc Finger and BTB Domain Containing 18) Alleviates Hepatic Steatohepatitis via FXR (Farnesoid X Receptor). Signal Transduct. Target. Ther. 2024, 9, 20. [Google Scholar] [CrossRef]
- Molière, S.; Jaulin, A.; Tomasetto, C.-L.; Dali-Youcef, N. Roles of Matrix Metalloproteinases and Their Natural Inhibitors in Metabolism: Insights into Health and Disease. Int. J. Mol. Sci. 2023, 24, 10649. [Google Scholar] [CrossRef]
- Himoto, T.; Masaki, T. Current Trends of Essential Trace Elements in Patients with Chronic Liver Diseases. Nutrients 2020, 12, 2084. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-H.; Wu, J.; Xu, Y.; Ding, C.-K.C.; Mestre, A.A.; Lin, C.-C.; Yang, W.-H.; Chi, J.-T. Zinc Transporter ZIP7 Is a Novel Determinant of Ferroptosis. Cell Death Dis. 2021, 12, 198. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.-H.; Tian, H.; Mao, L.; Li, D.-Y.; Lin, J.-Q.; Hu, H.-S.; Huang, S.-C.; Zhang, C.-J.; Mei, X.-F. Zinc Attenuates Ferroptosis and Promotes Functional Recovery in Contusion Spinal Cord Injury by Activating Nrf2/GPX4 Defense Pathway. CNS Neurosci. Ther. 2021, 27, 1023–1040. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Duntas, L.H.; Rayman, M.P. The Role of Selenium in Type-2 Diabetes Mellitus and Its Metabolic Comorbidities. Redox Biol. 2022, 50, 102236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, X.; Wei, Y. Selenium and Selenoproteins in Health. Biomolecules 2023, 13, 799. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; He, X.; Chen, X.; Li, Y.; Ke, Y. SeP Is Elevated in NAFLD and Participates in NAFLD Pathogenesis through AMPK/ACC Pathway. J. Cell. Physiol. 2021, 236, 3800–3807. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Li, S.; Zhang, K.; Li, J.; Han, Y.; Zhan, T.; Zhao, Q.; Guo, X.; Zhang, J. Selenium Deficiency-Induced Redox Imbalance Leads to Metabolic Reprogramming and Inflammation in the Liver. Redox Biol. 2020, 36, 101519. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, B.; Wu, P.; Chu, Y.; Gui, S.; Zheng, Y.; Chen, X. Dietary Selenium Alleviated Mouse Liver Oxidative Stress and NAFLD Induced by Obesity by Regulating the KEAP1/NRF2 Pathway. Antioxidants 2022, 11, 349. [Google Scholar] [CrossRef]
- Xu, L.; Lu, Y.; Wang, N.; Feng, Y. The Role and Mechanisms of Selenium Supplementation on Fatty Liver-Associated Disorder. Antioxidants 2022, 11, 922. [Google Scholar] [CrossRef]
- Day, K.; Seale, L.A.; Graham, R.M.; Cardoso, B.R. Selenotranscriptome Network in Non-Alcoholic Fatty Liver Disease. Front. Nutr. 2021, 8, 744825. [Google Scholar] [CrossRef]
- Ursini, F.; Maiorino, M. Lipid Peroxidation and Ferroptosis: The Role of GSH and GPx4. Free Radic. Biol. Med. 2020, 152, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Reja, M.; Makar, M.; Visaria, A.; Marino, D.; Rustgi, V. Increased Serum Selenium Levels Are Associated with Reduced Risk of Advanced Liver Fibrosis and All-Cause Mortality in NAFLD Patients: National Health and Nutrition Examination Survey (NHANES) III. Ann. Hepatol. 2020, 19, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Seo, Y.A.; Park, S.K. Serum Selenium and Non-Alcoholic Fatty Liver Disease (NAFLD) in U.S. Adults: National Health and Nutrition Examination Survey (NHANES) 2011–2016. Environ. Res. 2021, 197, 111190. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tan, L.; Liu, Z.; Shi, R. The Association between Non-Alcoholic Fatty Liver Disease (NAFLD) and Advanced Fibrosis with Blood Selenium Level Based on the NHANES 2017–2018. Ann. Med. 2022, 54, 2259–2268. [Google Scholar] [CrossRef]
- Lee, W.; Koo, H.R.; Choi, Y.-J.; Choi, J.G.; Oh, M.S.; Jin, X.; Choo, M.; Park, S.; Jeong, Y.-S.; Chung, S.-J.; et al. Z-Ligustilide and n-Butylidenephthalide Isolated from the Aerial Parts of Angelica tenuissima Inhibit Lipid Accumulation In Vitro and In Vivo. Planta Med. 2019, 85, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhang, N.; Aldhahrani, A.; Soliman, M.M.; Zhang, L.; Zhou, F. Puerarin Ameliorates Nonalcoholic Fatty Liver in Rats by Regulating Hepatic Lipid Accumulation, Oxidative Stress, and Inflammation. Front. Immunol. 2022, 13, 956688. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; Zhang, L.; La, X.; Wu, H.; Yang, R.; Li, H.; Li, Z. Ferulic Acid and P-Coumaric Acid Synergistically Attenuate Non-Alcoholic Fatty Liver Disease through HDAC1/PPARG-Mediated Free Fatty Acid Uptake. Int. J. Mol. Sci. 2022, 23, 15297. [Google Scholar] [CrossRef] [PubMed]
- Nie, K.; Gao, Y.; Chen, S.; Wang, Z.; Wang, H.; Tang, Y.; Su, H.; Lu, F.; Dong, H.; Fang, K. Diosgenin Attenuates Non-Alcoholic Fatty Liver Disease in Type 2 Diabetes through Regulating SIRT6-Related Fatty Acid Uptake. Phytomedicine 2023, 111, 154661. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Sun, S.; Feng, Y.; Li, X.; Yin, G.; Liang, P.; Yu, W.; Meng, D.; Zhang, X.; Liu, H.; et al. Diosgenin Attenuates Nonalcoholic Hepatic Steatosis through the Hepatic FXR-SHP-SREBP1C/PPARα/CD36 Pathway. Eur. J. Pharmacol. 2023, 952, 175808. [Google Scholar] [CrossRef]
- Pengnet, S.; Sumarithum, P.; Phongnu, N.; Prommaouan, S.; Kantip, N.; Phoungpetchara, I.; Malakul, W. Naringin Attenuates Fructose-Induced NAFLD Progression in Rats through Reducing Endogenous Triglyceride Synthesis and Activating the Nrf2/HO-1 Pathway. Front. Pharmacol. 2022, 13, 1049818. [Google Scholar] [CrossRef]
- Zhong, Y.; Li, Z.; Jin, R.; Yao, Y.; He, S.; Lei, M.; Wang, X.; Shi, C.; Gao, L.; Peng, X. Diosgenin Ameliorated Type II Diabetes-Associated Nonalcoholic Fatty Liver Disease through Inhibiting De Novo Lipogenesis and Improving Fatty Acid Oxidation and Mitochondrial Function in Rats. Nutrients 2022, 14, 4994. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, C.; Song, Y.; Chen, L.; Chen, X.; Zheng, G.; Yang, Y.; Cao, P.; Qiu, Z. Gallic Acid Impairs Fructose-Driven de Novo Lipogenesis and Ameliorates Hepatic Steatosis via AMPK-Dependent Suppression of SREBP-1/ACC/FASN Cascade. Eur. J. Pharmacol. 2023, 940, 175457. [Google Scholar] [CrossRef]
- Xing, Y.; Ren, X.; Li, X.; Sui, L.; Shi, X.; Sun, Y.; Quan, C.; Xiu, Z.; Dong, Y. Baicalein Enhances the Effect of Acarbose on the Improvement of Nonalcoholic Fatty Liver Disease Associated with Prediabetes via the Inhibition of De Novo Lipogenesis. J. Agric. Food Chem. 2021, 69, 9822–9836. [Google Scholar] [CrossRef] [PubMed]
- Gnoni, A.; Di Chiara Stanca, B.; Giannotti, L.; Gnoni, G.V.; Siculella, L.; Damiano, F. Quercetin Reduces Lipid Accumulation in a Cell Model of NAFLD by Inhibiting De Novo Fatty Acid Synthesis through the Acetyl-CoA Carboxylase 1/AMPK/PP2A Axis. Int. J. Mol. Sci. 2022, 23, 1044. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Sun, Y.; Chen, L.; Hu, J.; Meng, Y.; Yuan, M.; Wang, Q.; Li, S.; Zheng, G.; Qiu, Z. Caffeine Ameliorates AKT-Driven Nonalcoholic Steatohepatitis by Suppressing De Novo Lipogenesis and MyD88 Palmitoylation. J. Agric. Food Chem. 2022, 70, 6108–6122. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Fang, Y.; Liu, W.; Wang, J.; Zhao, J.; Tang, H.; Yin, Y.; Hu, Y.; Peng, J. Inhibition on XBP1s-Driven Lipogenesis by Qushi Huayu Decoction Contributes to Amelioration of Hepatic Steatosis Induced by Fructose. J. Ethnopharmacol. 2023, 301, 115806. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.-S.; Zhang, S.-F.; Luo, G.; Cheng, B.C.-Y.; Zhang, C.; Wang, Y.-W.; Qiu, X.-Y.; Zhou, X.-H.; Wang, Q.-G.; Song, X.-L.; et al. Schisandrin B Mitigates Hepatic Steatosis and Promotes Fatty Acid Oxidation by Inducing Autophagy through AMPK/mTOR Signaling Pathway. Metabolism 2022, 131, 155200. [Google Scholar] [CrossRef]
- Liu, X.; Hu, M.; Ye, C.; Liao, L.; Ding, C.; Sun, L.; Liang, J.; Chen, Y. Isosilybin Regulates Lipogenesis and Fatty Acid Oxidation via the AMPK/SREBP-1c/PPARα Pathway. Chem. Biol. Interact. 2022, 368, 110250. [Google Scholar] [CrossRef]
- Hai, Y.; Zuo, L.; Wang, M.; Zhang, R.; Wang, M.; Ren, L.; Yang, C.; Wang, J. Icariin Alleviates Nonalcoholic Fatty Liver Disease in Polycystic Ovary Syndrome by Improving Liver Fatty Acid Oxidation and Inhibiting Lipid Accumulation. Molecules 2023, 28, 517. [Google Scholar] [CrossRef]
- Luo, Z.; Li, M.; Yang, Q.; Zhang, Y.; Liu, F.; Gong, L.; Han, L.; Wang, M. Ferulic Acid Prevents Nonalcoholic Fatty Liver Disease by Promoting Fatty Acid Oxidation and Energy Expenditure in C57BL/6 Mice Fed a High-Fat Diet. Nutrients 2022, 14, 2530. [Google Scholar] [CrossRef]
- Wu, Y.-K.; Ren, Z.-N.; Zhu, S.-L.; Wu, Y.-Z.; Wang, G.; Zhang, H.; Chen, W.; He, Z.; Ye, X.-L.; Zhai, Q.-X. Sulforaphane Ameliorates Non-Alcoholic Fatty Liver Disease in Mice by Promoting FGF21/FGFR1 Signaling Pathway. Acta Pharmacol. Sin. 2022, 43, 1473–1483. [Google Scholar] [CrossRef] [PubMed]
- Nie, T.; Wang, X.; Li, A.; Shan, A.; Ma, J. The Promotion of Fatty Acid β-Oxidation by Hesperidin via Activating SIRT1/PGC1α to Improve NAFLD Induced by a High-Fat Diet. Food Funct. 2024, 15, 372–386. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhang, Y.; Chen, Q.; He, Y.; Zhou, H.; Wan, H.; Yang, J. Formononetin Protects against Inflammation Associated with Cerebral Ischemia-Reperfusion Injury in Rats by Targeting the JAK2/STAT3 Signaling Pathway. Biomed. Pharmacother. 2022, 149, 112836. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, R.P.; Moore, M.P.; Moore, A.N.; Healy, J.C.; Roberts, M.D.; Rector, R.S.; Martin, J.S. Curcumin Supplementation Mitigates NASH Development and Progression in Female Wistar Rats. Physiol. Rep. 2018, 6, e13789. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Sheng, F.; Zou, L.; Xiao, J.; Li, P. Hyperoside Attenuates Non-Alcoholic Fatty Liver Disease in Rats via Cholesterol Metabolism and Bile Acid Metabolism. J. Adv. Res. 2021, 34, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Xiong, T.; Liu, P.; Guo, X.; Xiao, L.; Zhou, F.; Tang, Y.; Yao, P. Quercetin Ameliorates HFD-Induced NAFLD by Promoting Hepatic VLDL Assembly and Lipophagy via the IRE1a/XBP1s Pathway. Food Chem. Toxicol. 2018, 114, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Seo, H.; Kwon, D.; Yuk, D.Y.; Jung, Y.-S. Taurine Ameliorates Tunicamycin-Induced Liver Injury by Disrupting the Vicious Cycle between Oxidative Stress and Endoplasmic Reticulum Stress. Life 2022, 12, 354. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wu, W.; Sheng, L.; Jiang, B.; Li, X.; Cai, K. Chrysin Ameliorates Hepatic Steatosis Induced by a Diet Deficient in Methionine and Choline by Inducing the Secretion of Hepatocyte Nuclear Factor 4α-Dependent Very Low-Density Lipoprotein. J. Biochem. Mol. Toxicol. 2020, 34, e22497. [Google Scholar] [CrossRef]
- Yang, S.; Cao, S.; Li, C.; Zhang, J.; Liu, C.; Qiu, F.; Kang, N. Berberrubine, a Main Metabolite of Berberine, Alleviates Non-Alcoholic Fatty Liver Disease via Modulating Glucose and Lipid Metabolism and Restoring Gut Microbiota. Front. Pharmacol. 2022, 13, 913378. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, L.L.; Tang, W.J.; Tang, B. Astragaloside IV Inhibits Protein Tyrosine Phosphatase 1B and Improves Insulin Resistance in Insulin-Resistant HepG2 Cells and Triglyceride Accumulation in Oleic Acid (OA)-Treated HepG2 Cells. J. Ethnopharmacol. 2021, 268, 113556. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Aguilera, Y.; Martin-Cabrejas, M.A.; Gonzalez de Mejia, E. Phytochemicals from the Cocoa Shell Modulate Mitochondrial Function, Lipid and Glucose Metabolism in Hepatocytes via Activation of FGF21/ERK, AKT, and mTOR Pathways. Antioxidants 2022, 11, 136. [Google Scholar] [CrossRef]
- Gai, H.; Zhou, F.; Zhang, Y.; Ai, J.; Zhan, J.; You, Y.; Huang, W. Coniferaldehyde Ameliorates the Lipid and Glucose Metabolism in Palmitic Acid-Induced HepG2 Cells via the LKB1/AMPK Signaling Pathway. J. Food Sci. 2020, 85, 4050–4060. [Google Scholar] [CrossRef]
- Hong, S.-H.; Hong, Y.; Lee, M.; Keum, B.-R.; Kim, G.-H. Natural Product Skatole Ameliorates Lipotoxicity-Induced Multiple Hepatic Damage under Hyperlipidemic Conditions in Hepatocytes. Nutrients 2023, 15, 1490. [Google Scholar] [CrossRef]
- Liou, C.-J.; Wu, S.-J.; Shen, S.-C.; Chen, L.-C.; Chen, Y.-L.; Huang, W.-C. Acacetin Protects against Non-Alcoholic Fatty Liver Disease by Regulating Lipid Accumulation and Inflammation in Mice. Int. J. Mol. Sci. 2022, 23, 4687. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Lin, L.; Yuan, Q.-Q.; Wang, X.-Y.; Zhang, Q.; Zhang, X.-M.; Tang, K.-C.; Guo, M.-Y.; Dong, T.-Y.; Han, W.; et al. Bavachin Protects against Diet-Induced Hepatic Steatosis and Obesity in Mice. Acta Pharmacol. Sin. 2023, 44, 1416–1428. [Google Scholar] [CrossRef]
- Zamani-Garmsiri, F.; Hashemnia, S.M.R.; Shabani, M.; Bagherieh, M.; Emamgholipour, S.; Meshkani, R. Combination of Metformin and Genistein Alleviates Non-Alcoholic Fatty Liver Disease in High-Fat Diet-Fed Mice. J. Nutr. Biochem. 2021, 87, 108505. [Google Scholar] [CrossRef] [PubMed]
- Zamani-Garmsiri, F.; Ghasempour, G.; Aliabadi, M.; Hashemnia, S.M.R.; Emamgholipour, S.; Meshkani, R. Combination of Metformin and Chlorogenic Acid Attenuates Hepatic Steatosis and Inflammation in High-Fat Diet Fed Mice. IUBMB Life 2021, 73, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Chen, Y.; Huang, X.; Yang, Z.; Zhang, J.; Yu, X.; Fang, J.; Tao, J.; You, K.; Cheng, Z.; et al. Icaritin Ameliorates Hepatic Steatosis via Promoting Fatty Acid β-Oxidation and Insulin Sensitivity. Life Sci. 2021, 268, 119000. [Google Scholar] [CrossRef]
- Sun, X.; Duan, X.; Wang, C.; Liu, Z.; Sun, P.; Huo, X.; Ma, X.; Sun, H.; Liu, K.; Meng, Q. Protective Effects of Glycyrrhizic Acid against Non-Alcoholic Fatty Liver Disease in Mice. Eur. J. Pharmacol. 2017, 806, 75–82. [Google Scholar] [CrossRef]
- Duan, X.; Meng, Q.; Wang, C.; Liu, Z.; Sun, H.; Huo, X.; Sun, P.; Ma, X.; Peng, J.; Liu, K. Effects of Calycosin against High-Fat Diet-Induced Nonalcoholic Fatty Liver Disease in Mice. J. Gastroenterol. Hepatol. 2018, 33, 533–542. [Google Scholar] [CrossRef]
- Yin, X.; Liu, Z.; Wang, J. Tetrahydropalmatine Ameliorates Hepatic Steatosis in Nonalcoholic Fatty Liver Disease by Switching Lipid Metabolism via AMPK-SREBP-1c-Sirt1 Signaling Axis. Phytomedicine 2023, 119, 155005. [Google Scholar] [CrossRef]
- Yang, Y.; Sheng, J.; Sheng, Y.; Wang, J.; Zhou, X.; Li, W.; Kong, Y. Lapachol Treats Non-Alcoholic Fatty Liver Disease by Modulating the M1 Polarization of Kupffer Cells via PKM2. Int. Immunopharmacol. 2023, 120, 110380. [Google Scholar] [CrossRef]
- Shi, X.-Y.; Zheng, X.-M.; Liu, H.-J.; Han, X.; Zhang, L.; Hu, B.; Li, S. Rotundic Acid Improves Nonalcoholic Steatohepatitis in Mice by Regulating Glycolysis and the TLR4/AP1 Signaling Pathway. Lipids Health Dis. 2023, 22, 214. [Google Scholar] [CrossRef] [PubMed]
- Ban, D.; Hua, S.; Zhang, W.; Shen, C.; Miao, X.; Liu, W. Costunolide Reduces Glycolysis-Associated Activation of Hepatic Stellate Cells via Inhibition of Hexokinase-2. Cell Mol. Biol. Lett. 2019, 24, 52. [Google Scholar] [CrossRef]
- Lian, N.; Jin, H.; Zhang, F.; Wu, L.; Shao, J.; Lu, Y.; Zheng, S. Curcumin Inhibits Aerobic Glycolysis in Hepatic Stellate Cells Associated with Activation of Adenosine Monophosphate-Activated Protein Kinase. IUBMB Life 2016, 68, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.-Y.; Choi, M.-S. Eriocitrin Improves Adiposity and Related Metabolic Disorders in High-Fat Diet-Induced Obese Mice. J. Med. Food 2020, 23, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Xiao, L.; Li, M.; Liu, S.; Wang, Y.; Huang, L.; Liu, S.; Jiang, T.; Zhou, L.; Li, Y. Perillartine Protects against Metabolic Associated Fatty Liver in High-Fat Diet-Induced Obese Mice. Food Funct. 2023, 14, 961–977. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.-S.; Choi, J.-Y.; Kwon, E.-Y. Fisetin Alleviates Hepatic and Adipocyte Fibrosis and Insulin Resistance in Diet-Induced Obese Mice. J. Med. Food 2020, 23, 1019–1032. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, R.; Zheng, Y.; Song, M.; Zhang, S.; Sun, Y.; Wei, M.; Fan, X. Diosgenin Ameliorates Non-Alcoholic Fatty Liver Disease by Modulating the Gut Microbiota and Related Lipid/Amino Acid Metabolism in High Fat Diet-Fed Rats. Front. Pharmacol. 2022, 13, 854790. [Google Scholar] [CrossRef]
- Shi, L.; Guo, S.; Zhang, S.; Gao, X.; Liu, A.; Wang, Q.; Zhang, T.; Zhang, Y.; Wen, A. Glycyrrhetinic Acid Attenuates Disturbed Vitamin a Metabolism in Non-Alcoholic Fatty Liver Disease through AKR1B10. Eur. J. Pharmacol. 2020, 883, 173167. [Google Scholar] [CrossRef]
- Nishino, A.; Maoka, T.; Yasui, H. Preventive Effects of β-Cryptoxanthin, a Potent Antioxidant and Provitamin A Carotenoid, on Lifestyle-Related Diseases—A Central Focus on Its Effects on Non-Alcoholic Fatty Liver Disease (NAFLD). Antioxidants 2021, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- Kusu, H.; Yoshida, H.; Kudo, M.; Okuyama, M.; Harada, N.; Tsuji-Naito, K.; Akagawa, M. Tomatidine Reduces Palmitate-Induced Lipid Accumulation by Activating AMPK via Vitamin D Receptor-Mediated Signaling in Human HepG2 Hepatocytes. Mol. Nutr. Food Res. 2019, 63, e1801377. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-J.; Huang, W.-C.; Yu, M.-C.; Chen, Y.-L.; Shen, S.-C.; Yeh, K.-W.; Liou, C.-J. Tomatidine Ameliorates Obesity-Induced Nonalcoholic Fatty Liver Disease in Mice. J. Nutr. Biochem. 2021, 91, 108602. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Song, A.; Ma, M.; Wang, P.; Zhang, X.; Lu, C.; Zhang, J.; Zheng, S.; Jin, H. Curcumol Inhibits Ferritinophagy to Restrain Hepatocyte Senescence through YAP/NCOA4 in Non-Alcoholic Fatty Liver Disease. Cell Prolif. 2021, 54, e13107. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jiang, W.; Feng, Y.; Wu, L.; Jia, Y.; Zhao, R. Betaine Alleviates High-Fat Diet-Induced Disruptionof Hepatic Lipid and Iron Homeostasis in Mice. Int. J. Mol. Sci. 2022, 23, 6263. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yan, J.; Guan, F.; Jin, Z.; Xie, J.; Wang, C.; Liu, M.; Liu, J. Zeaxanthin Prevents Ferroptosis by Promoting Mitochondrial Function and Inhibiting the P53 Pathway in Free Fatty Acid-Induced HepG2 Cells. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2023, 1868, 159287. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.-B.; Chu, X.-L.; Jin, Y.-X.; Jiang, J.-J.; Zhao, X.; Yu, M. Epigallocatechin Gallate Alleviates High-Fat Diet-Induced Hepatic Lipotoxicity by Targeting Mitochondrial ROS-Mediated Ferroptosis. Front. Pharmacol. 2023, 14, 1148814. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, J.; Gao, Q.; Shan, X.; Wang, J.; Lv, Z. Study on the Attenuated Effect of Ginkgolide B on Ferroptosis in High Fat Diet Induced Nonalcoholic Fatty Liver Disease. Toxicology 2020, 445, 152599. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Xie, Z.; Li, E.-W.; Yuan, Y.; Fu, Y.; Wang, P.; Zhang, X.; Qiao, Y.; Xu, J.; Hölscher, C.; et al. Dehydroabietic Acid Improves Nonalcoholic Fatty Liver Disease through Activating the Keap1/Nrf2-ARE Signaling Pathway to Reduce Ferroptosis. J. Nat. Med. 2021, 75, 540–552. [Google Scholar] [CrossRef]
- Jiang, J.-J.; Zhang, G.-F.; Zheng, J.-Y.; Sun, J.-H.; Ding, S.-B. Targeting Mitochondrial ROS-Mediated Ferroptosis by Quercetin Alleviates High-Fat Diet-Induced Hepatic Lipotoxicity. Front. Pharmacol. 2022, 13, 876550. [Google Scholar] [CrossRef]
- Ye, Q.; Jiang, Y.; Wu, D.; Cai, J.; Jiang, Z.; Zhou, Z.; Liu, L.; Ling, Q.; Wang, Q.; Zhao, G. Atractylodin Alleviates Nonalcoholic Fatty Liver Disease by Regulating Nrf2-Mediated Ferroptosis. Heliyon 2023, 9, e18321. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Choi, H.; Chung, J. Icariin Supplementation Suppresses the Markers of Ferroptosis and Attenuates the Progression of Nonalcoholic Steatohepatitis in Mice Fed a Methionine Choline-Deficient Diet. Int. J. Mol. Sci. 2023, 24, 12510. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Xiao, Y.; Zhou, J.; Luo, Z.; Yu, L.; Liao, Q.; Liu, S.; Qi, X.; Zhang, H.; Hou, M.; et al. Arbutin Alleviates Fatty Liver by Inhibiting Ferroptosis via FTO/SLC7A11 Pathway. Redox Biol. 2023, 68, 102963. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Xia, L.; Song, J.; Hu, H.; Zang, N.; Yang, J.; Zou, Y.; Wang, L.; Zheng, X.; He, Q.; et al. Puerarin Ameliorates Metabolic Dysfunction-Associated Fatty Liver Disease by Inhibiting Ferroptosis and Inflammation. Lipids Health Dis. 2023, 22, 202. [Google Scholar] [CrossRef] [PubMed]
- Porcu, C.; Sideri, S.; Martini, M.; Cocomazzi, A.; Galli, A.; Tarantino, G.; Balsano, C. Oleuropein Induces AMPK-Dependent Autophagy in NAFLD Mice, Regardless of the Gender. Int. J. Mol. Sci. 2018, 19, 3948. [Google Scholar] [CrossRef] [PubMed]
- Santini, S.J.; Tarantino, G.; Iezzi, A.; Alisi, A.; Balsano, C. Copper-Catalyzed Dicarbonyl Stress in NAFLD Mice: Protective Effects of Oleuropein Treatment on Liver Damage. Nutr. Metab. 2022, 19, 9. [Google Scholar] [CrossRef] [PubMed]
- Miyata, M.; Matsushita, K.; Shindo, R.; Shimokawa, Y.; Sugiura, Y.; Yamashita, M. Selenoneine Ameliorates Hepatocellular Injury and Hepatic Steatosis in a Mouse Model of NAFLD. Nutrients 2020, 12, 1898. [Google Scholar] [CrossRef] [PubMed]
- Mun, J.; Kim, S.; Yoon, H.-G.; You, Y.; Kim, O.-K.; Choi, K.-C.; Lee, Y.-H.; Lee, J.; Park, J.; Jun, W. Water Extract of Curcuma longa L. Ameliorates Non-Alcoholic Fatty Liver Disease. Nutrients 2019, 11, 2536. [Google Scholar] [CrossRef] [PubMed]
- Le, T.N.H.; Choi, H.-J.; Jun, H.-S. Ethanol Extract of Liriope platyphylla Root Attenuates Non-Alcoholic Fatty Liver Disease in High-Fat Diet-Induced Obese Mice via Regulation of Lipogenesis and Lipid Uptake. Nutrients 2021, 13, 3338. [Google Scholar] [CrossRef]
- Guo, Y.; Sun, Q.; Wang, S.; Zhang, M.; Lei, Y.; Wu, J.; Wang, X.; Hu, W.; Meng, H.; Li, Z.; et al. Corydalis saxicola Bunting Total Alkaloids Improve NAFLD by Suppressing de Novo Lipogenesis through the AMPK-SREBP1 Axis. J. Ethnopharmacol. 2024, 319, 117162. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Maggi, F.; Daglia, M.; Habtemariam, S.; Rastrelli, L.; Nabavi, S.M. Pharmacological Effects of Capparis spinosa L. Phytother. Res. 2016, 30, 1733–1744. [Google Scholar] [CrossRef] [PubMed]
- Akbari, R.; Yaghooti, H.; Jalali, M.T.; Khorsandi, L.S.; Mohammadtaghvaei, N. Capparis spinosa Improves Non-Alcoholic Steatohepatitis through down-Regulating SREBP-1c and a PPARα-Independent Pathway in High-Fat Diet-Fed Rats. BMC Res. Notes 2022, 15, 315. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Luo, Y.; Wang, L.; Zhang, K.; Peng, J.; Fan, G. Therapeutic Effect of Curcumin on Metabolic Diseases: Evidence from Clinical Studies. Int. J. Mol. Sci. 2023, 24, 3323. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Fujita, S.; Yamagishi, A.; Shirai, T.; Maeda, Y.; Suzuki, T.; Kobayashi, K.-I.; Inoue, J.; Yamamoto, Y. Brown Rice Inhibits Development of Nonalcoholic Fatty Liver Disease in Obese Zucker (Fa/Fa) Rats by Increasing Lipid Oxidation Via Activation of Retinoic Acid Synthesis. J. Nutr. 2021, 151, 2705–2713. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Zhang, Y.; Lin, S.; Chen, Y.; Wang, Z.; Feng, H.; Fang, G.; Quan, S. Berberine Ameliorates Metabolic-Associated Fatty Liver Disease Mediated Metabolism Disorder and Redox Homeostasis by Upregulating Clock Genes: Clock and Bmal1 Expressions. Molecules 2023, 28, 1874. [Google Scholar] [CrossRef]
- Zheng, X.; Zhao, M.-G.; Jiang, C.-H.; Sheng, X.-P.; Yang, H.-M.; Liu, Y.; Yao, X.-M.; Zhang, J.; Yin, Z.-Q. Triterpenic Acids-Enriched Fraction from Cyclocarya paliurus Attenuates Insulin Resistance and Hepatic Steatosis via PI3K/Akt/GSK3β Pathway. Phytomedicine 2020, 66, 153130. [Google Scholar] [CrossRef] [PubMed]
- Dangana, E.O.; Omolekulo, T.E.; Areola, E.D.; Olaniyi, K.S.; Soladoye, A.O.; Olatunji, L.A. Sodium Acetate Protects against Nicotine-Induced Excess Hepatic Lipid in Male Rats by Suppressing Xanthine Oxidase Activity. Chem. Biol. Interact. 2020, 316, 108929. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, Z.; Othman, Z.A.; Suleiman, J.B.; Mustaffa, K.M.F.; Jalil, N.A.C.; Ghazali, W.S.W.; Zulkipli, N.N.; Mohamed, M.; Kamaruzaman, K.A. Therapeutic Effects of Heterotrigona itama (Stingless Bee) Bee Bread in Improving Hepatic Lipid Metabolism through theActivation of the Keap1/Nrf2 Signaling Pathway in an Obese Rat Model. Antioxidants 2022, 11, 2190. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, H.; Hu, J.; Zheng, J.; Zhou, J.; Botchlett, R.; Matthews, D.; Zeng, T.; Chen, L.; Xiao, X.; et al. Indole Alleviates Diet-Induced Hepatic Steatosis and Inflammation in a Manner Involving Myeloid Cell 6-Phosphofructo-2-Kinase/Fructose-2,6-Biphosphatase 3. Hepatology 2020, 72, 1191–1203. [Google Scholar] [CrossRef]
- Yang, Z.; Su, H.; Lv, Y.; Tao, H.; Jiang, Y.; Ni, Z.; Peng, L.; Chen, X. Inulin Intervention Attenuates Hepatic Steatosis in Rats via Modulating Gut Microbiota and Maintaining Intestinal Barrier Function. Food Res. Int. 2023, 163, 112309. [Google Scholar] [CrossRef]
- Liao, J.; Cao, Y.; Zhao, J.; Yu, B.; Wang, Y.; Li, W.; Li, H.; Lv, S.; Wen, W.; Cui, H.; et al. Aqueous Extract of Polygala japonica Houtt. Ameliorated Nonalcoholic Steatohepatitis in Mice through Restoring the Gut Microbiota Disorders and Affecting the Metabolites in Feces and Liver. Phytomedicine 2023, 118, 154937. [Google Scholar] [CrossRef] [PubMed]
- Pastorino, G.; Cornara, L.; Soares, S.; Rodrigues, F.; Oliveira, M.B.P.P. Liquorice (Glycyrrhiza glabra): A Phytochemical and Pharmacological Review. Phytother. Res. 2018, 32, 2323–2339. [Google Scholar] [CrossRef]
- Ujianti, I.; Sianipar, I.R.; Prijanti, A.R.; Hasan, I.; Arozal, W.; Jusuf, A.A.; Wibowo, H.; Prihartono, J.; Amani, P.; Santoso, D.I.S. Effect of Roselle Flower Extract (Hibiscus sabdariffa Linn.) on Reducing Steatosis and Steatohepatitis in Vitamin B12 Deficiency Rat Model. Medicina 2023, 59, 1044. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Xiong, H.; Tsao, R.; Shahidi, F.; Wen, X.; Liu, J.; Jiang, L.; Sun, Y. Green Pea (Pisum sativum L.) Hull Polyphenol Extract Alleviates NAFLD through VB6/TLR4/NF-κB and PPAR Pathways. J. Agric. Food Chem. 2023, 71, 16067–16078. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, J.; Zhang, Y.; Li, J.; Sun, W. Black Sesame Seeds Ethanol Extract Ameliorates Hepatic Lipid Accumulation, Oxidative Stress, and Insulin Resistance in Fructose-Induced Nonalcoholic Fatty Liver Disease. J. Agric. Food Chem. 2018, 66, 10458–10469. [Google Scholar] [CrossRef] [PubMed]
- Veeramani, C.; Alsaif, M.A.; Al-Numair, K.S. Lavatera Critica, a Green Leafy Vegetable, Controls High Fat Diet Induced Hepatic Lipid Accumulation and Oxidative Stress through the Regulation of Lipogenesis and Lipolysis Genes. Biomed. Pharmacother. 2017, 96, 1349–1357. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Rasul, A.; Sadiqa, A.; Sarfraz, I.; Hussain, G.; Nageen, B.; Liu, X.; Watanabe, N.; Selamoglu, Z.; Ali, M.; et al. Curcumol: From Plant Roots to Cancer Roots. Int. J. Biol. Sci. 2019, 15, 1600–1609. [Google Scholar] [CrossRef]
- Deng, G.; Liu, C.; Zhao, J.; Wang, M.; Li, Y.; Yang, M.; Ye, H.; Li, J.; Qin, M.; Wu, C.; et al. Exocarpium Citri Grandis Alleviates the Aggravation of NAFLD by Mitigating Lipid Accumulation and Iron Metabolism Disorders. J. Ethnopharmacol. 2023, 313, 116559. [Google Scholar] [CrossRef]
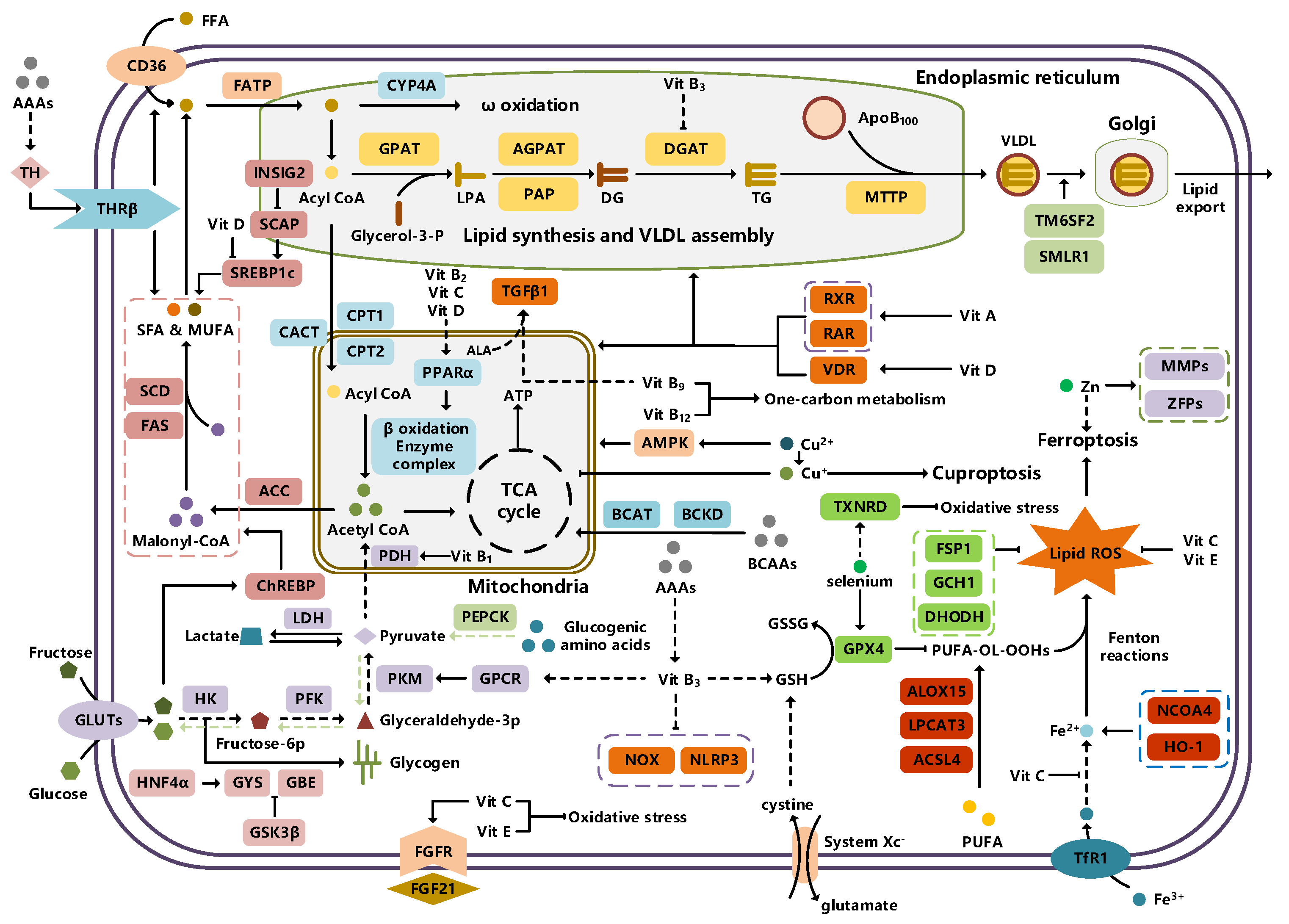

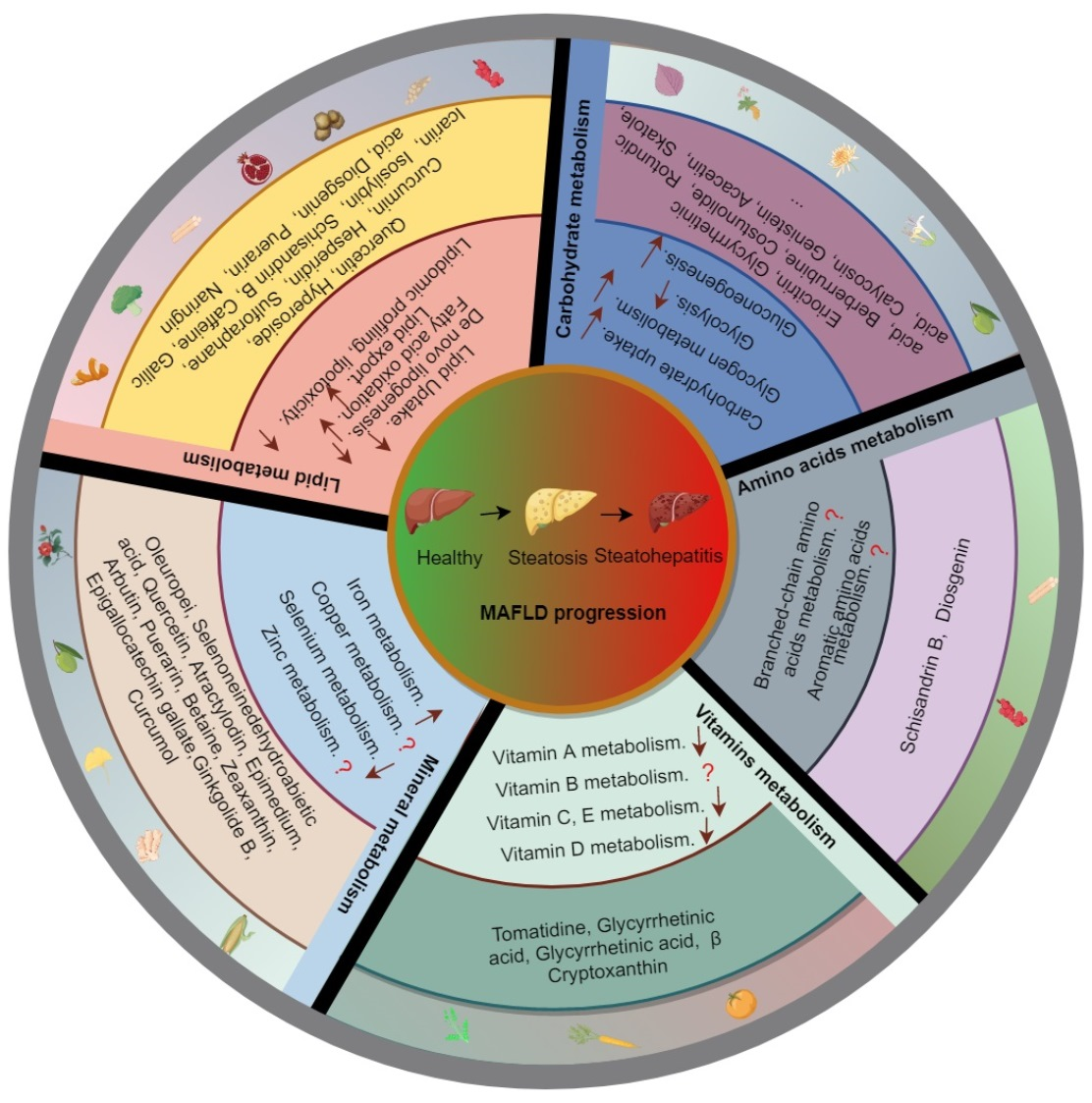{kind=link}
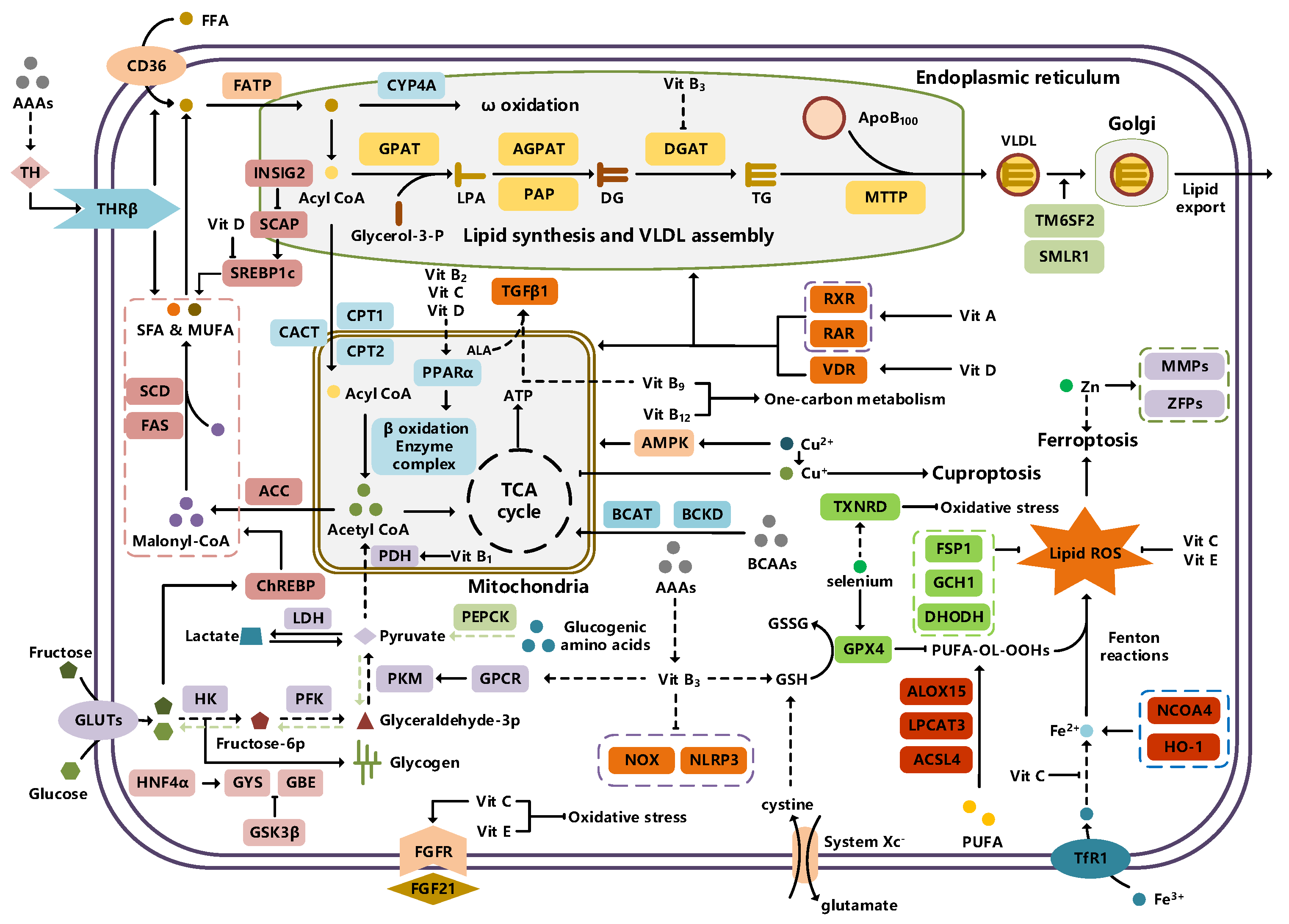{kind=link}
| Compound | Chemical Structure | Potential Mechanism | Potential Targets | References |
|---|---|---|---|---|
| Z-ligustilide |  | Lipid uptake | CD36, FATP5 | [265] |
| n-Butylidenephthalide | 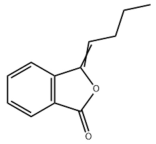 | Lipid uptake | CD36, FATP5 | [265] |
| Puerarin |  | Lipid uptake | CD36, FATP5 | [266] |
| Cerulic acid |  | Lipid uptake | FABP, CD36 | [267] |
| P-coumaric acid | 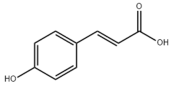 | Lipid uptake | FABP, CD36 | [267] |
| Diosgenin | 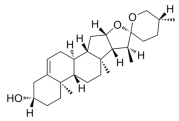 | Lipid uptake | CD36, FATP2, FABP | [268,269] |
| Naringin | 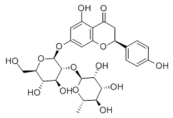 | De novo lipogenesis | ChREBP, SREBP-1c | [270] |
| Diosgenin |  | De novo lipogenesis | AMPK, SREBP-1c | [269,271] |
| Gallic acid |  | De novo lipogenesis | AMPK | [272] |
| Baicalein | 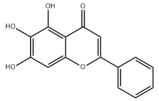 | De novo lipogenesis | AMPK | [273] |
| Quercetin | 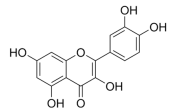 | De novo lipogenesis | ACC, AMPK | [274] |
| Caffeine |  | De novo lipogenesis | AKT, SREBP1c | [275] |
| Chlorogenic acid |  | De novo lipogenesis | XBP1 | [276] |
| Geniposide |  | De novo lipogenesis | XBP1 | [276] |
| Polydatin |  | De novo lipogenesis | XBP1 | [276] |
| Schisandrin B |  | Fatty acid oxidation | AMPK | [277] |
| Isosilybin |  | Fatty acid oxidation | AMPK, PPARα, ACOX1, CPT1α | [278] |
| Icariin |  | Fatty acid oxidation | PPARα, ACOX1, CYP4A3, CPT1α | [279] |
| Ferulicacid |  | Fatty acid oxidation | CPT1α, ACOX1, HMGCS2 | [280] |
| Sulforaphane |  | Fatty acid oxidation | FGF21, FGFR1, p38 MAPK, PPARα, CPT1α | [281] |
| Hesperidin | 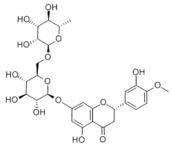 | Fatty acid oxidation | SIRT1, PGC1α | [282] |
| Formononetin | 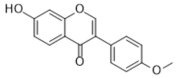 | Fatty acid oxidation | SIRT1, PGC1α | [283] |
| Curcumin |  | Lipid output | ApoB100 | [284] |
| Hyperoside | 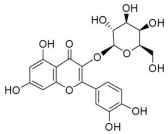 | Lipid output | ApoC3, VLDL | [285] |
| Quercetin | 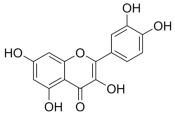 | Lipid output | IRE1, XBP1s, VLDL | [286] |
| Taurine | 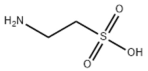 | Lipid output | IRE1, XBP1s, ATF6, CHOP, MTTP, VLDL | [287] |
| Chrysin | 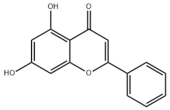 | Lipid output | PKC, HNF4α, VLDL | [288] |
| Berberrubine | 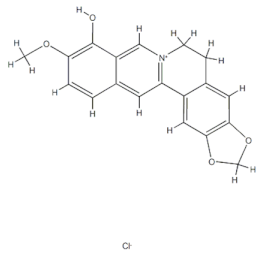 | Carbohydrate intake | GLUT2 | [289] |
| Astragaloside IV |  | Carbohydrate intake | PTP1B, IR, IRS1 | [290] |
| Protocatechuic Acid | 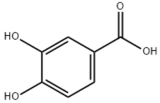 | Carbohydrate intake | AKT, GLUT2 | [291] |
| Epicatechin |  | Carbohydrate intake | AKT, GLUT2 | [291] |
| Coniferaldehyde | 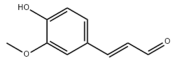 | Carbohydrate intake | AMPK, GLUT2 | [292] |
| Skatole | 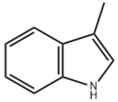 | Carbohydrate intake | PERK, IRE1, etc. | [293] |
| Acacetin | 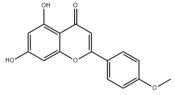 | Glycogen synthesis | AMPK | [294] |
| Bavachin | 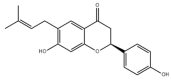 | Glycogen synthesis | AKT, GSK3β | [295] |
| Genistein | 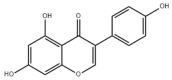 | Glycogen synthesis | GSK3β | [296] |
| Chlorogenic acid | 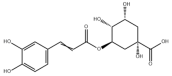 | Glycogen synthesis | GSK3β | [297] |
| Coniferaldehyde |  | Glycogen synthesis | GK | [292] |
| Icaritin | 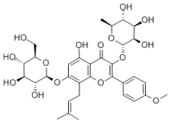 | Glycogen synthesis | AKT, GSK3β | [298] |
| Glycyrrhizic acid | 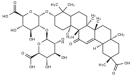 | Glycogen synthesis | GSK3β | [299] |
| Calycosin |  | Glycogen synthesis | GSK3β | [300] |
| Tetrahydropalmatine |  | Glycolysis | AMPK | [301] |
| Lapachol |  | Glycolysis | PKM2 | [302] |
| Rotundic acid |  | Glycolysis | TLR4, AP1 | [303] |
| costunolide |  | Glycolysis | HK2 | [304] |
| Curcumin |  | Glycolysis | AMPK, HK, PFK2 | [305] |
| Glycyrrhizic acid |  | Gluconeogenesis | G6Pase, PEPCK | [299] |
| Berberrubine |  | Gluconeogenesis | G6Pase, PEPCK | [289] |
| Eriocitrin |  | Gluconeogenesis | PEPCK | [306] |
| Perillartine |  | Gluconeogenesis | AKT, RORγ, G6P, PEPCK | [307] |
| Fisetin |  | Gluconeogenesis | G6Pase, PEPCK | [308] |
| Diosgenin |  | Amino acid metabolism | — | [309] |
| Schisandrin B |  | Amino acid metabolism | — | [277] |
| Glycyrrhetinic acid |  | Vitamin A metabolism | AKR1B10 | [310] |
| β cryptoxanthin |  | Vitamin A metabolism | IRS, etc. | [311] |
| Tomatidine |  | Vitamin D metabolism | VDR, AMPK | [312,313] |
| Curcumol |  | Iron metabolism | YAP, NCOA4 | [314] |
| Betaine | 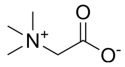 | Iron metabolism | FPN, HAMP | [315] |
| Zeaxanthin | 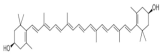 | Iron metabolism | p53, GPX4, SLC7A11, SAT1, ALOX15 | [316] |
| Epigallocatechin gallate | 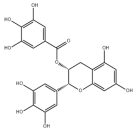 | Iron metabolism | GPX4, COX2, ACSL4 | [317] |
| Ginkgolide B | 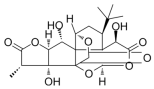 | Iron metabolism | Nrf2, GPX4, HO-1, TFR1, FTH1 | [318] |
| Dehydroabietic acid |  | Iron metabolism | Nrf2, GPX4, HO-1, FSP1 | [319] |
| Quercetin |  | Iron metabolism | GPX4, COX2, ACSL4 | [320] |
| Atractylodin |  | Iron metabolism | Nrf2, GPX4, FTH1, SLC7A11 | [321] |
| Icariin |  | Iron metabolism | Nrf2, SLC7A11, GPX4 | [322] |
| Arbutin |  | Iron metabolism | FTO, SLC7A11 | [323] |
| Puerarin |  | Iron metabolism | SIRT1, Nrf2 | [324] |
| Oleuropein |  | Copper metabolism | AMPK, CTR1, CTR2, ATP7B, COX17, CCS, ATOX1 | [325,326] |
| Selenoneine |  | Selenium metabolism | HMOX1, GSTA1/2, GPX1, Selenoprotein | [327] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Wang, Z.; Qin, H. Examining the Pathogenesis of MAFLD and the Medicinal Properties of Natural Products from a Metabolic Perspective. Metabolites 2024, 14, 218. https://doi.org/10.3390/metabo14040218
Fu Y, Wang Z, Qin H. Examining the Pathogenesis of MAFLD and the Medicinal Properties of Natural Products from a Metabolic Perspective. Metabolites. 2024; 14(4):218. https://doi.org/10.3390/metabo14040218
Chicago/Turabian StyleFu, Yansong, Zhipeng Wang, and Hong Qin. 2024. "Examining the Pathogenesis of MAFLD and the Medicinal Properties of Natural Products from a Metabolic Perspective" Metabolites 14, no. 4: 218. https://doi.org/10.3390/metabo14040218
APA StyleFu, Y., Wang, Z., & Qin, H. (2024). Examining the Pathogenesis of MAFLD and the Medicinal Properties of Natural Products from a Metabolic Perspective. Metabolites, 14(4), 218. https://doi.org/10.3390/metabo14040218