

The Virulence Factor Macrophage Infectivity Potentiator (Mip) Influences Branched-Chain Amino Acid Metabolism and Pathogenicity of Legionella pneumophila

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Bacterial Cultures
2.3. Minimal Inhibitory Concentration (MIC) Assay
2.4. Infection Experiments
2.5. Gas Chromatograph–Mass Spectrometry (GC/MS), Sample Preparation, and Analysis
2.6. Isotopic Tracing
2.7. Statistics
3. Results
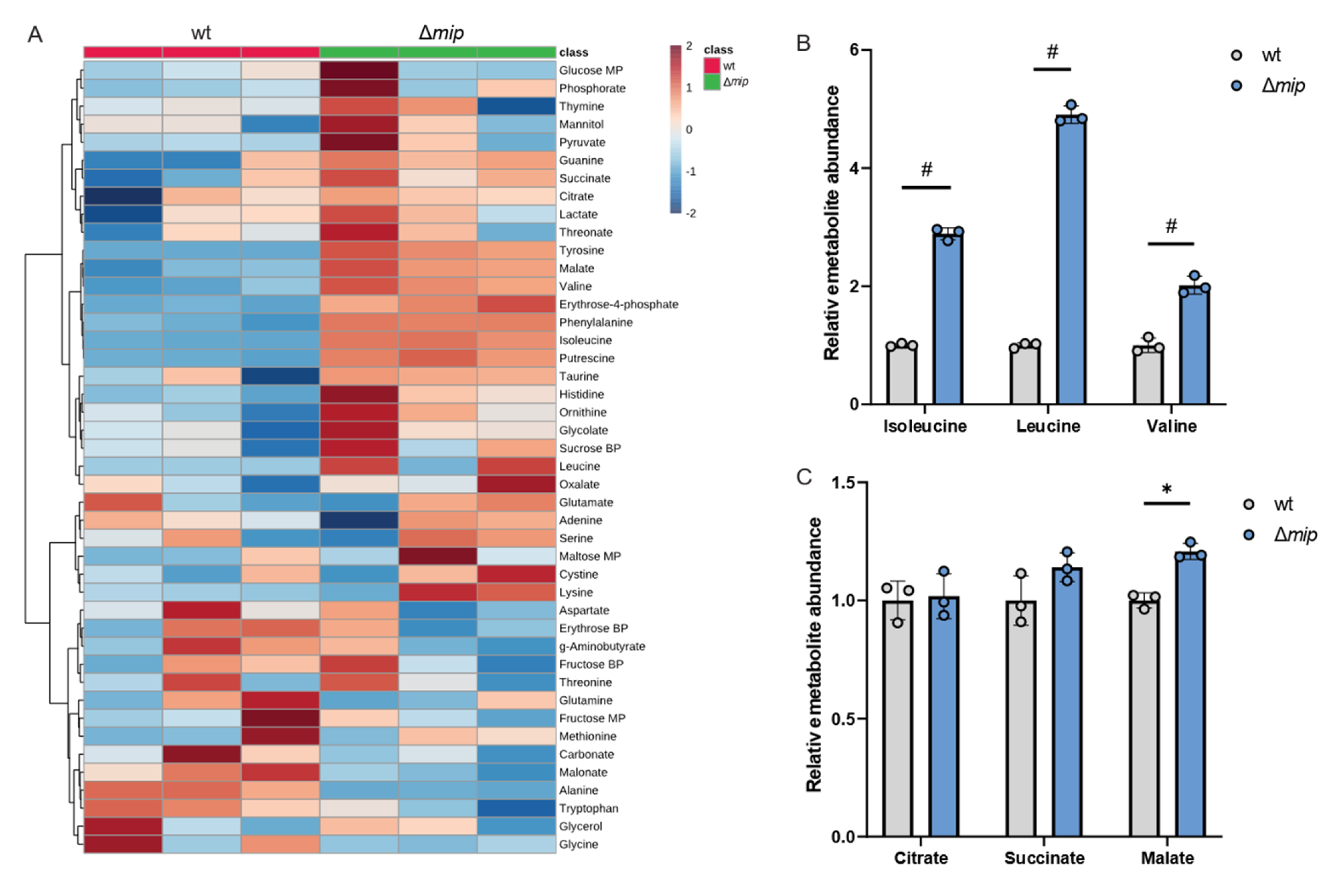
3.1. Impaired PPIase Activity Influences Lp Growth and Virulence
3.2. Metabolic Impact of Genetically Altered Mip Activity in Engineered Lp
3.3. Metabolic Impact of Chemical Inhibition of PPIase in Lp
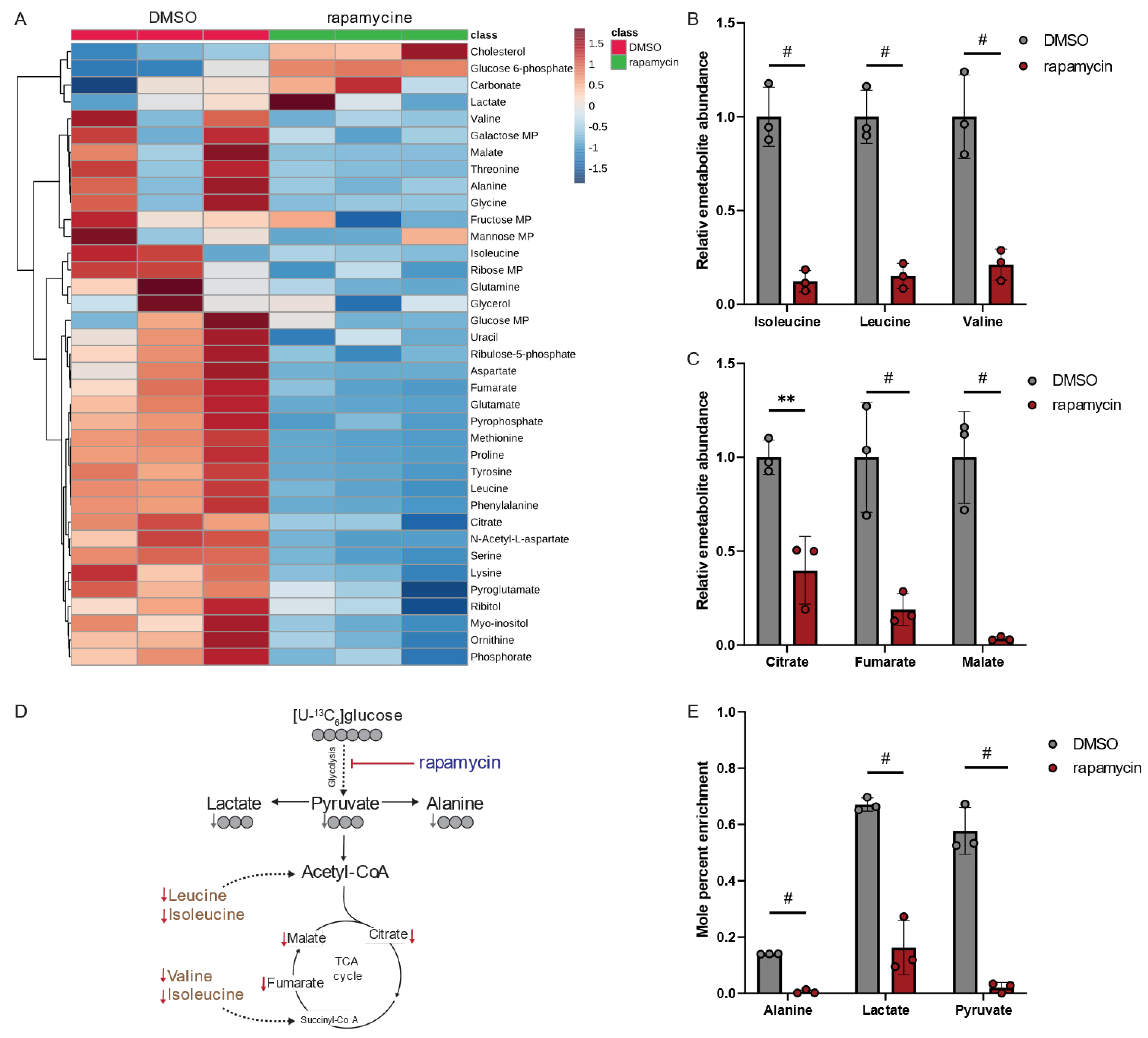
3.4. Metabolic Consequences of Altered PPIase Activity in Human Macrophages
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 12C | 12-carbon |
| 13C | 13-carbon |
| Δmip strain | Mip knockout strain |
| ACES | N-(2-acetamido)-2-aminoethanesulfonic acid |
| BCAAs | Branched-chain amino acids |
| BCYE | Buffered charcoal yeast extract |
| BP | By-product |
| Cfu | Colony forming unit |
| DMSO | Dimethyl sulfoxide |
| ECM | Extracellular matrix |
| FKBP | FK506-binding protein |
| GC/MS | Gas chromatograph coupled to mass spectrometry |
| LC/MS | Liquid chromatograph coupled to mass spectrometry |
| LD | Legionnaires’ disease |
| Lp | Legionella pneumophila |
| Mip | Macrophage infectivity potentiator protein |
| MP | Main product |
| MSTFA | N-methyl-N-(trimethylsilyl)trifluoracetamid |
| MTBSTFA | N-(tert-butyldimethylsilyl)-N-methyltrifluoracetamid |
| MOI | Multiplicity of infection |
| mTOR | Mammalian target of rapamycin |
| PMA | Phorbol 12-myristate 13-acetate |
| PPIase | Peptidyl-prolyl-cis/trans-isomerase |
| Wt | Wild type |
| YEB | Yeast extract beef |
References
- Rasch, J.; Ünal, C.M.; Klages, A.; Karsli, Ü.; Heinsohn, N.; Brouwer, R.M.H.J.; Richter, M.; Dellmann, A.; Steinert, M. Peptidyl-Prolyl-cis/trans-Isomerases Mip and PpiB of Legionella pneumophila Contribute to Surface Translocation, Growth at Suboptimal Temperature, and Infection. Infect. Immun. 2018, 87, e00939-17. [Google Scholar] [CrossRef]
- He, W.; Henne, A.; Lauterbach, M.; Geißmar, E.; Nikolka, F.; Kho, C.; Heinz, A.; Dostert, C.; Grusdat, M.; Cordes, T.; et al. Mesaconate is synthesized from itaconate and exerts immunomodulatory effects in macrophages. Nat. Metab. 2022, 4, 524–533. [Google Scholar] [CrossRef]
- Cordes, T.; Metallo, C.M. Itaconate Alters Succinate and Coenzyme A Metabolism via Inhibition of Mitochondrial Complex II and Methylmalonyl-CoA Mutase. Metabolites 2021, 11, 117. [Google Scholar] [CrossRef]
- de Goede, K.E.; Harber, K.J.; Gorki, F.S.; Verberk, S.G.; Groh, L.A.; Keuning, E.D.; Struys, E.A.; van Weeghel, M.; Haschemi, A.; de Winther, M.P.; et al. d-2-Hydroxyglutarate is an anti-inflammatory immunometabolite that accumulates in macrophages after TLR4 activation. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166427. [Google Scholar] [CrossRef] [PubMed]
- Kaymak, I.; Luda, K.M.; Duimstra, L.R.; Ma, E.H.; Longo, J.; Dahabieh, M.S.; Faubert, B.; Oswald, B.M.; Watson, M.J.; Kitchen-Goosen, S.M.; et al. Carbon source availability drives nutrient utilization in CD8+ T cells. Cell Metab. 2022, 34, 1298–1311.e6. [Google Scholar] [CrossRef]
- Cordes, T.; Kuna, R.S.; McGregor, G.H.; Khare, S.V.; Gengatharan, J.; Muthusamy, T.; Metallo, C.M. 1-Deoxysphingolipid synthesis compromises anchorage-independent growth and plasma membrane endocytosis in cancer cells. J. Lipid Res. 2022, 63, 100281. [Google Scholar] [CrossRef] [PubMed]
- Mondino, S.; Schmidt, S.; Rolando, M.; Escoll, P.; Gomez-Valero, L.; Buchrieser, C. Legionnaires’ Disease: State of the Art Knowledge of Pathogenesis Mechanisms of Legionella. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 439–466. [Google Scholar] [CrossRef]
- Eisenreich, W.; Heuner, K. The life stage-specific pathometabolism of Legionella pneumophila. FEBS Lett. 2016, 590, 3868–3886. [Google Scholar] [CrossRef] [PubMed]
- Gillmaier, N.; Schunder, E.; Kutzner, E.; Tlapák, H.; Rydzewski, K.; Herrmann, V.; Stämmler, M.; Lasch, P.; Eisenreich, W.; Heuner, K. Growth-related Metabolism of the Carbon Storage Poly-3-hydroxybutyrate in Legionella pneumophila. J. Biol. Chem. 2016, 291, 6471–6482. [Google Scholar] [CrossRef]
- Cianciotto, N.P.; Stamos, J.K.; Kamp, D.W. Infectivity of Legionella pneumophila mip Mutant for Alveolar Epithelial Cells. Curr. Microbiol. 1995, 30, 247–250. [Google Scholar] [CrossRef]
- Karagöz, M.S.; Ünal, C.M.; Mayer, B.E.; Müsken, M.; Borrero-de Acuña, J.M.; Steinert, M. Legionella pneumophila PPIase Mip Interacts with the Bacterial Proteins SspB, Lpc2061, and FlaA and Promotes Flagellation. Infect. Immun. 2022, 90, e0027622. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.; Khan, A.S.; Kamphausen, T.; Schmausser, B.; Unal, C.; Lorenz, U.; Fischer, G.; Hacker, J.; Steinert, M. Collagen binding protein Mip enables Legionella pneumophila to transmigrate through a barrier of NCI-H292 lung epithelial cells and extracellular matrix. Cell. Microbiol. 2007, 9, 450–462. [Google Scholar] [CrossRef]
- Cordes, T.; Metallo, C.M. Tracing insights into human metabolism using chemical engineering approaches. Curr. Opin. Chem. Eng. 2016, 14, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Jepras, R.I.; Fitzgeorge, R.B.; Baskerville, A. A Comparison of Virulence of Two Strains of Legionella Pneumophila Based on Experimental Aerosol Infection of Guinea-Pigs. Epidemiol. Infect. 1985, 95, 29–38. [Google Scholar] [CrossRef]
- Wintermeyer, E.; Ludwig, B.; Steinert, M.; Schmidt, B.; Fischer, G.; Hacker, J. Influence of Site Specifically Altered Mip Proteins on Intracellular Survival of Legionella pneumophila in Eukaryotic Cells. Infect. Immun. 1995, 63, 4576–4583. [Google Scholar] [CrossRef] [PubMed]
- Cordes, T.; Metallo, C.M. Quantifying Intermediary Metabolism and Lipogenesis in Cultured Mammalian Cells Using Stable Isotope Tracing and Mass Spectrometry. In High-Throughput Metabolomics: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2019; Volume 1978, pp. 219–241. [Google Scholar] [CrossRef]
- Hiller, K.; Hangebrauk, J.; Jäger, C.; Spura, J.; Schreiber, K.; Schomburg, D. MetaboliteDetector: Comprehensive analysis tool for targeted and nontargeted GC/MS based metabolome analysis. Anal. Chem. 2009, 81, 3429–3439. [Google Scholar] [CrossRef]
- Sündermann, A.; Eggers, L.F.; Schwudke, D. Liquid Extraction: Bligh and Dyer. In Encyclopedia of Lipidomics; Wenk, M.R., Ed.; Springer: Dordrecht, The Netherlands, 2016; pp. 1–4. [Google Scholar] [CrossRef]
- Oliva, G.; Sahr, T.; Buchrieser, C. The Life Cycle of L. pneumophila: Cellular Differentiation Is Linked to Virulence and Metabolism. Front. Cell. Infect. Microbiol. 2018, 8, 3. [Google Scholar] [CrossRef]
- Rasch, J.; Theuerkorn, M.; Ünal, C.; Heinsohn, N.; Tran, S.; Fischer, G.; Weiwad, M.; Steinert, M. Novel Cycloheximide Derivatives Targeting the Moonlighting Protein Mip Exhibit Specific Antimicrobial Activity Against Legionella pneumophila. Front. Bioeng. Biotechnol. 2015, 3, 41. [Google Scholar] [CrossRef]
- Pomplun, S.; Sippel, C.; Hähle, A.; Tay, D.; Shima, K.; Klages, A.; Ünal, C.M.; Rieß, B.; Toh, H.T.; Hansen, G.; et al. Chemogenomic Profiling of Human and Microbial FK506-Binding Proteins. J. Med. Chem. 2018, 61, 3660–3673. [Google Scholar] [CrossRef]
- Meier-Kriesche, H.U.; Li, S.; Gruessner, R.W.; Fung, J.J.; Bustami, R.T.; Barr, M.L.; Leichtman, A.B. Immunosuppression: Evolution in practice and trends, 1994–2004. Am. J. Transplant. 2006, 6 Pt 2, 1111–1131. [Google Scholar] [CrossRef]
- Martel, R.R.; Klicius, J.; Galet, S. Inhibition of the immune response by rapamycin, a new antifungal antibiotic. Can. J. Physiol. Pharmacol. 1977, 55, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Heitman, J.; Movva, N.R.; Hall, M.N. Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 1991, 253, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Heitman, J.; Koller, A.; Kunz, J.; Henriquez, R.; Schmidt, A.; Movva, N.R.; Hall, M.N. The immunosuppressant FK506 inhibits amino acid import in Saccharomyces cerevisiae. Mol. Cell. Biol. 1993, 13, 5010–5019. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef]
- Tang, Y.; Wallace, M.; Sanchez-Gurmaches, J.; Hsiao, W.Y.; Li, H.; Lee, P.L.; Vernia, S.; Metallo, C.M.; Guertin, D.A. Adipose tissue mTORC2 regulates ChREBP-driven de novo lipogenesis and hepatic glucose metabolism. Nat. Commun. 2016, 7, 11365. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Tian, J.; Miao, Z.; Liang, W.; Wang, G. Metabolome and Transcriptome Profiling Reveal Carbon Metabolic Flux Changes in Yarrowia lipolytica Cells to Rapamycin. J. Fungi 2022, 8, 939. [Google Scholar] [CrossRef]
- Abshire, C.F.; Dragoi, A.M.; Roy, C.R.; Ivanov, S.S. MTOR-Driven Metabolic Reprogramming Regulates Legionella pneumophila Intracellular Niche Homeostasis. PLoS Pathog. 2016, 12, e1006088. [Google Scholar] [CrossRef]
- Sancak, Y.; Peterson, T.R.; Shaul, Y.D.; Lindquist, R.A.; Thoreen, C.C.; Bar-Peled, L.; Sabatini, D.M. The Rag GTPases bind raptor and mediate amino acid signaling to mTORC1. Science 2008, 320, 1496–1501. [Google Scholar] [CrossRef]
- Yip, C.K.; Murata, K.; Walz, T.; Sabatini, D.M.; Kang, S.A. Structure of the human mTOR complex I and its implications for rapamycin inhibition. Mol. Cell 2010, 38, 768–774. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikolka, F.; Karagöz, M.S.; Nassef, M.Z.; Hiller, K.; Steinert, M.; Cordes, T. The Virulence Factor Macrophage Infectivity Potentiator (Mip) Influences Branched-Chain Amino Acid Metabolism and Pathogenicity of Legionella pneumophila. Metabolites 2023, 13, 834. https://doi.org/10.3390/metabo13070834
Nikolka F, Karagöz MS, Nassef MZ, Hiller K, Steinert M, Cordes T. The Virulence Factor Macrophage Infectivity Potentiator (Mip) Influences Branched-Chain Amino Acid Metabolism and Pathogenicity of Legionella pneumophila. Metabolites. 2023; 13(7):834. https://doi.org/10.3390/metabo13070834
Chicago/Turabian StyleNikolka, Fabian, Mustafa Safa Karagöz, Mohamed Zakaria Nassef, Karsten Hiller, Michael Steinert, and Thekla Cordes. 2023. "The Virulence Factor Macrophage Infectivity Potentiator (Mip) Influences Branched-Chain Amino Acid Metabolism and Pathogenicity of Legionella pneumophila" Metabolites 13, no. 7: 834. https://doi.org/10.3390/metabo13070834
APA StyleNikolka, F., Karagöz, M. S., Nassef, M. Z., Hiller, K., Steinert, M., & Cordes, T. (2023). The Virulence Factor Macrophage Infectivity Potentiator (Mip) Influences Branched-Chain Amino Acid Metabolism and Pathogenicity of Legionella pneumophila. Metabolites, 13(7), 834. https://doi.org/10.3390/metabo13070834