

Independent and Combined Effects of Probiotics and Prebiotics as Supplements or Food-Rich Diets on a Propionic-Acid-Induced Rodent Model of Autism Spectrum Disorder

 , ,
, ,  and
and
Abstract

1. Introduction
2. Results
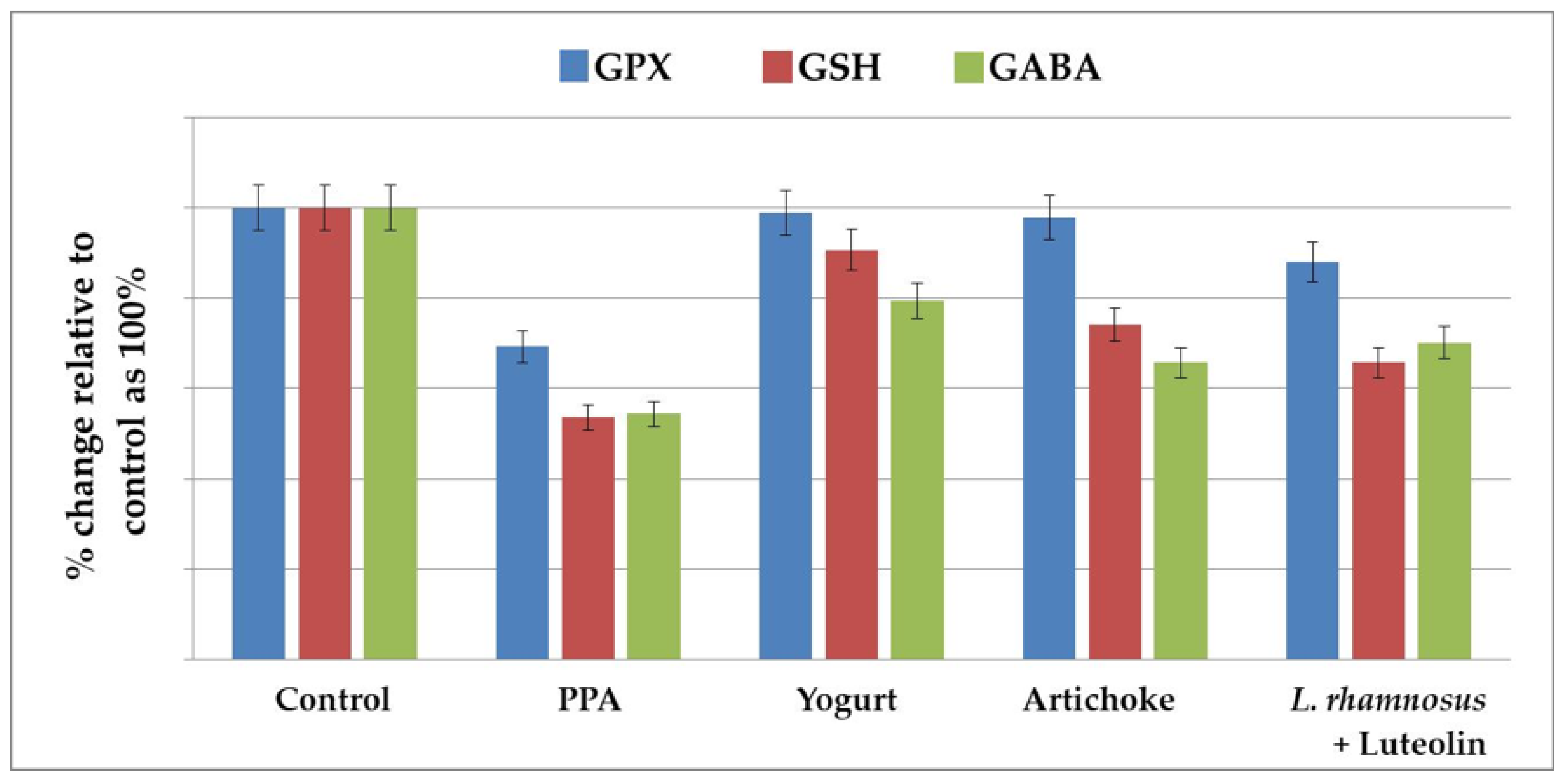
2.1. Effect of the Nutritional Interventions on GPX1 and GSH as Oxidative Stress Markers and the Impaired GABA Neurotransmitter
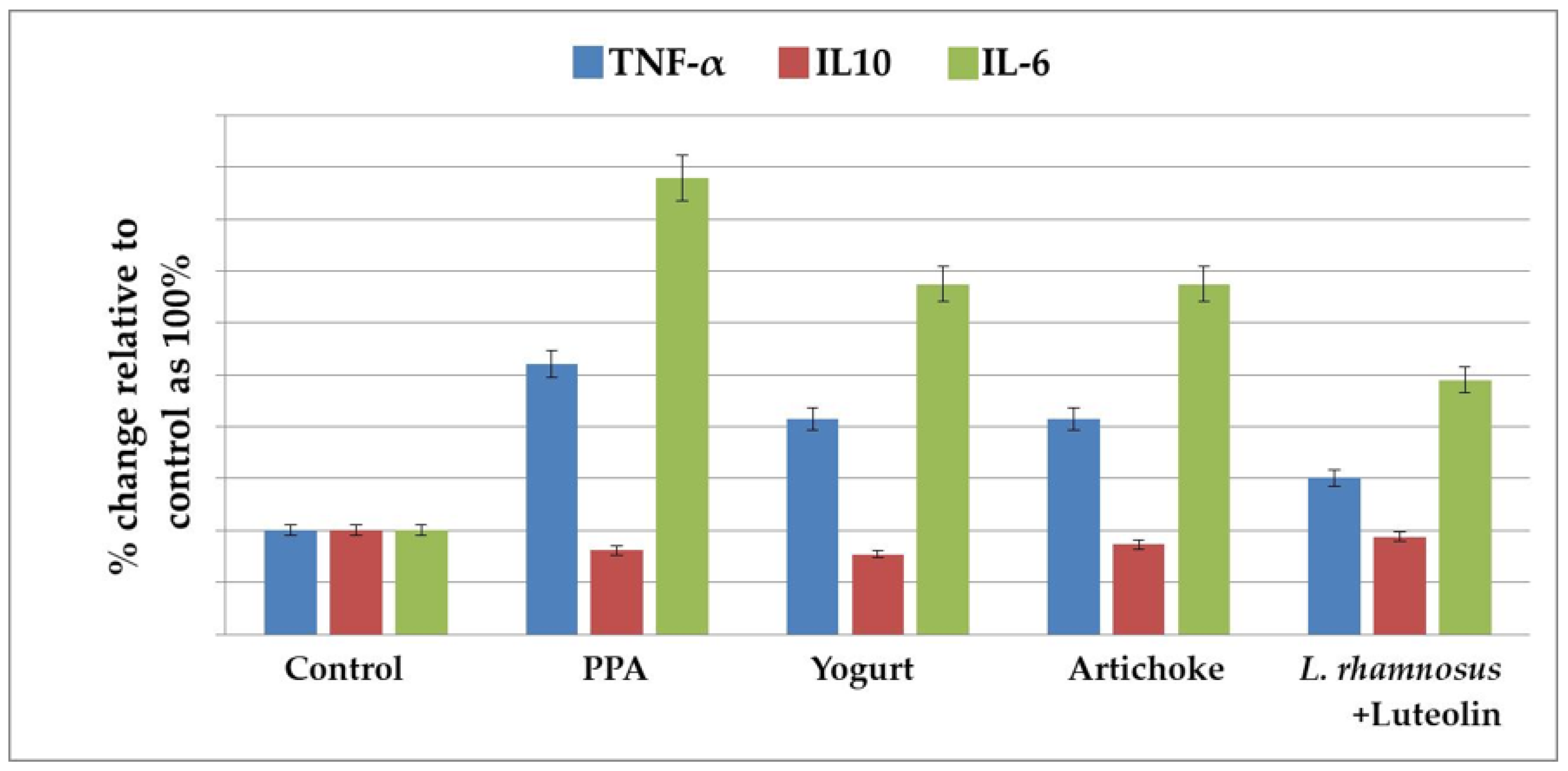
2.2. Effect of the Nutritional Interventions on the Levels of TNF-α, IL-10, and IL-6 as Neuroinflammatory Markers
2.3. Receiver Operating Characteristic Analysis for Evaluating Predictive Values of the Measured Variables in the PPA-Induced Autism Model and Different Nutritionally Treated Groups
3. Discussion
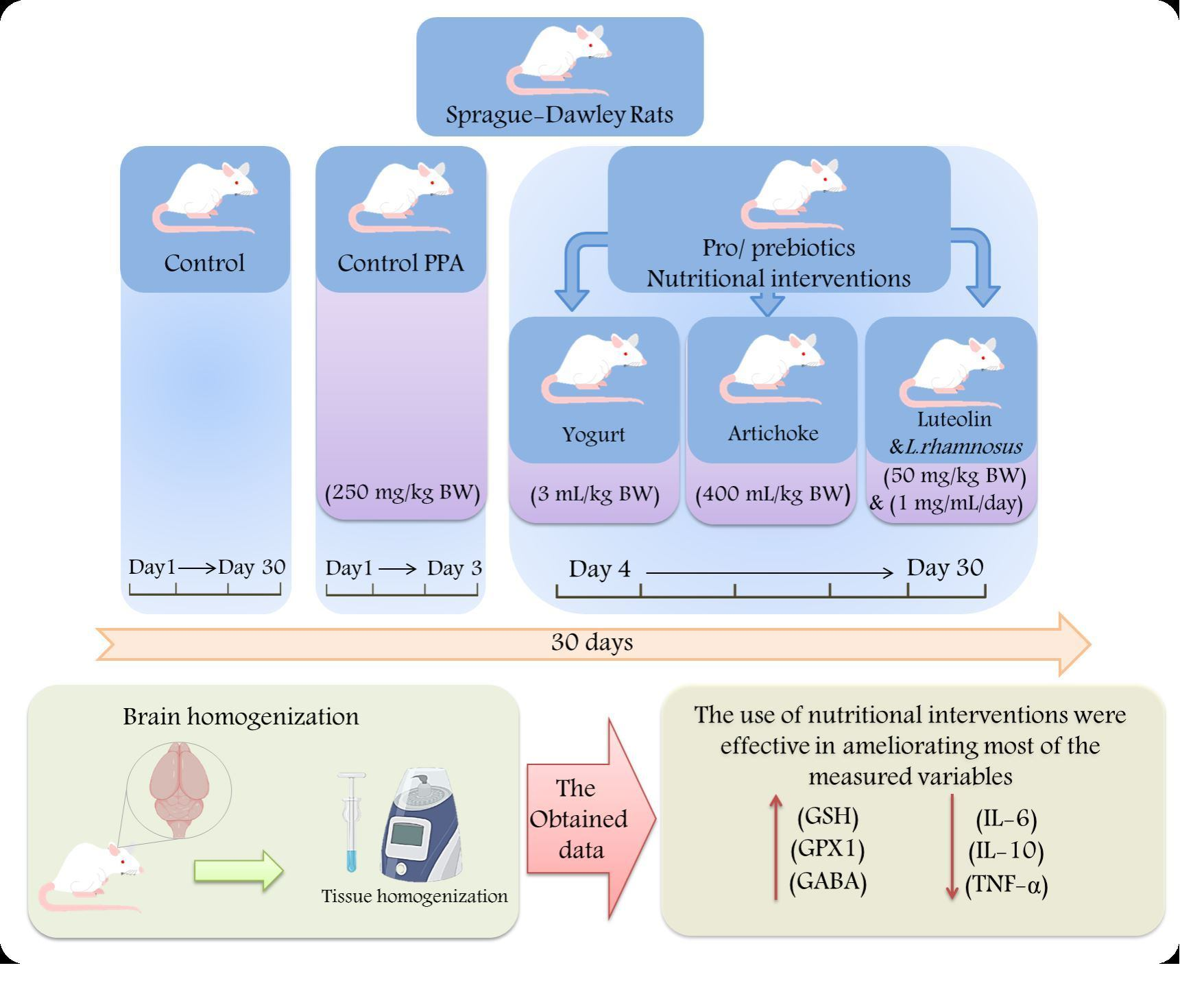
4. Materials and Methods
4.1. Materials
4.2. Preparation of Cynara Scolymus L. (Artichoke) Extract
4.3. Animals
4.4. Preparation of Brain Tissue Homogenates
4.5. Biochemical Analyses
4.5.1. Determination of GSH
4.5.2. Determination of GPX1
4.5.3. Determination of GABA
4.5.4. Determination of IL-6
4.5.5. Determination of IL-10
4.6. Statistical Analyses
5. Conclusions
6. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vargas, D.L.; Nascimbene, C.; Krishnan, C.; Zimmerman, A.W.; Pardo, C.A. Neuroglial Activation and Neuroinflammation in the Brain of Patients With Autism. Ann. Neurol. 2005, 57, 67–81. [Google Scholar] [CrossRef]
- Fung, T.C.; Olson, C.A.; Hsiao, E.Y. Interactions between the Microbiota, Immune and Nervous Systems in Health and Disease. Nat. Neurosci. 2017, 20, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Sherwin, E.; Dinan, T.G.; Cryan, J.F. Recent Developments in Understanding the Role of the Gut Microbiota in Brain Health and Disease. Ann. N. Y. Acad. Sci. 2017, 17, 5–25. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.T.; Chana, G.; Pardo, C.A.; Achim, C.; Semendeferi, K.; Buckwalter, J.; Courchesne, E.; Everall, I.P. Microglial Activation and Increased Microglial Density Observed in the Dorsolateral Prefrontal Cortex in Autism. Biol. Psychiatry 2010, 68, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Vismara, L.A.; Rogers, S.J. Behavioral Treatments in Autism Spectrum Disorder: What Do We Know? Annu. Rev. Clin. Psychol. 2016, 6, 447–468. [Google Scholar] [CrossRef] [PubMed]
- Louis, P. Does the Human Gut Microbiota Contribute to the Etiology of Autism Spectrum Disorders? Dig. Dis. Sci. 2012, 57, 1987–1989. [Google Scholar] [CrossRef]
- Abdellatif, B.; McVeigh, C.; Bendriss, G.; Chaari, A. The Promising Role of Probiotics in Managing the Altered Gut in Autism Spectrum Disorders. Int. J. Mol. Sci. 2020, 21, 4159. [Google Scholar] [CrossRef]
- Adams, J.B.; Audhya, T.; Mcdonough-means, S.; Rubin, R.A.; Quig, D.; Geis, E.; Gehn, E.; Loresto, M.; Mitchell, J.; Atwood, S.; et al. Nutritional and Metabolic Status of Children with Autism vs. Neurotypical Children, and the Association with Autism Severity. Nutr. Metab. 2011, 8, 34. [Google Scholar] [CrossRef]
- De Angelis, M.; Piccolo, M.; Vannini, L.; Siragusa, S.; De Giacomo, A.; Serrazzanetti, I.; Cristofori, F.; Guerzoni, M.E.; Gobbetti, M. Fecal Microbiota and Metabolome of Children with Autism and Pervasive Developmental Disorder Not Otherwise Specified. PLoS ONE 2013, 8, e76993. [Google Scholar] [CrossRef]
- Finegold, S.M.; Downes, J.; Summanen, P.H. Microbiology of Regressive Autism. Anaerobe 2012, 18, 260–262. [Google Scholar] [CrossRef]
- Williams, B.L.; Hornig, M.; Buie, T.; Bauman, M.L.; Cho Paik, M.; Wick, I.; Bennett, A.; Jabado, O.; Hirschberg, D.L.; Lipkin, W.I. Impaired Carbohydrate Digestion and Transport and Mucosal Dysbiosis in the Intestines of Children with Autism and Gastrointestinal Disturbances. PLoS ONE 2011, 6, e24585. [Google Scholar] [CrossRef] [PubMed]
- Navarro, F.; Liu, Y.; Rhoads, J.M. Can Probiotics Benefit Children with Autism Spectrum Disorders? World J. Gastroenterol. 2016, 22, 10093–10102. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.; Roberfroid, M.B. Critical Review Dietary Modulation of the Human Colonie Microbiota: Introducing the Concept of Prebiotics. Am. Inst. Nutr. 1995, 125, 1401–1412. [Google Scholar]
- De Angelis, M.; Francavilla, R.; Piccolo, M.; De Giacomo, A.; De Angelis, M.; Francavilla, R.; Piccolo, M.; De Giacomo, A. Autism Spectrum Disorders and Intestinal Microbiota. Gut Microbes 2015, 6, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, J.; Wu, F.; Zheng, H.; Peng, Q.; Zhou, H. Altered Composition and Function of Intestinal Microbiota in Autism Spectrum Disorders: A Systematic Review. Transl. Psychiatry 2019, 9, 13. [Google Scholar] [CrossRef]
- Kang, D.; Adams, J.B.; Gregory, A.C.; Borody, T.; Chittick, L.; Fasano, A.; Khoruts, A.; Geis, E.; Maldonado, J.; Mcdonough-means, S.; et al. Microbiota Transfer Therapy Alters Gut Ecosystem and Improves Gastrointestinal and Autism Symptoms: An Open-Label Study. Microbiome 2017, 5, 1–16. [Google Scholar] [CrossRef]
- Strati, F.; Cavalieri, D.; Albanese, D.; De Felice, C.; Donati, C.; Hayek, J.; Jousson, O.; Leoncini, S.; Renzi, D.; Calabrò, A.; et al. New Evidences on the Altered Gut Microbiota in Autism Spectrum Disorders. Microbiome 2017, 5, 10. [Google Scholar] [CrossRef]
- Kang, D.; Ilhan, Z.E.; Isern, N.G.; Hoyt, D.W.; Daniel, P.; Shaffer, M.; Lozupone, C.A.; Hahn, J.; Adams, J.B.; Krajmalnik-brown, R. Differences in Fecal Microbial Metabolites and Microbiota of Children with Autism Spectrum Disorders. Anaerobe 2018, 49, 121–131. [Google Scholar] [CrossRef]
- Brandão, T.L.S.; Silva, J.C.L.; Campos, S.É.D.; Francelino, J.O. Supplementation of Prebiotics and Probiotics in Autistic Children: Integrative Review. Res. Soc. Dev. 2022, 2022, e12811124061. [Google Scholar] [CrossRef]
- Pranckutė, R.; Kaunietis, A.; Kuisiene, N.; Čitavičius, D.J. Combining Prebiotics with Probiotic Bacteria Can Enhance Bacterial Growth and Secretion of Bacteriocins. Int. J. Biol. Macromol. 2016, 89, 669–679. [Google Scholar] [CrossRef]
- Costabile, A.; Frohberg, C.; Kolida, S.; Klinder, A.; Gietl, E.; Ba, M.; Gibson, G.R. Bifidogenic Effect of a Very-Long-Chain Inulin Extracted from Globe Artichoke (Cynara Scolymus) in Healthy Human Subjects. Br. J. Nutr. 2010, 104, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.A.; Pereira, C.; Ntatsi, G.; Danalatos, N.; Barros, L.; Ferreira, I.C.F.R. Nutritional Value and Chemical Composition of Greek Artichoke Genotypes. Food Chem. 2017, 267, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.; Kim, M.Y.; Cho, J.Y. Anti-Inflammatory Effects of Luteolin: A Review of in Vitro, in Vivo, and in Silico Studies. J. Ethnopharmacol. 2018, 225, 342–358. [Google Scholar] [CrossRef]
- Kok, C.R.; Hutkins, R. Yogurt and Other Fermented Foods as Sources of Health-Promoting Bacteria. Nutr. Rev. 2018, 76, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Tillisch, K.; Gupta, A.; Mayer, E.A.; Tillisch, K.; Gupta, A. Gut/Brain Axis and the Microbiota. J. Clin. Investig. 2015, 125, 926–938. [Google Scholar] [CrossRef]
- Shultz, S.R.; Macfabe, D.F. Propionic Acid Animal Model of Autism Propionic Acid Animal Model of Autism. Compr. Guide Autism 2014, 1755–1778. [Google Scholar] [CrossRef]
- Choi, J.; Lee, S.; Won, J.; Jin, Y.; Hong, Y.; Hur, T.; Kim, J.; Lee, S.; Hong, Y. Pathophysiological and Neurobehavioral Characteristics of a Propionic Acid-Mediated Autism-like Rat Model. PLoS ONE 2018, 13, e0192925. [Google Scholar]
- Abuaish, S.; Al-Otaibi, N.M.; Aabed, K.; Abujamel, T.S.; Alzahrani, S.A.; Alotaibi, S.M.; Bhat, R.S.; Arzoo, S.; Algahtani, N.; Moubayed, N.M.; et al. The Efficacy of Fecal Transplantation and Bifidobacterium Supplementation in Ameliorating Propionic Acid-Induced Behavioral and Biochemical Autistic Features in Juvenile Male Rats. J. Mol. Neurosci. 2022, 72, 372–381. [Google Scholar] [CrossRef]
- Ali, E.; Elmalahy, H.; Abbas, O.; Abu Almaaty, A. Is propionic acid a suitable model for autism? Alfarama J. Basic Appl. Sci. 2022, 3, 45–63. [Google Scholar] [CrossRef]
- El-Ansary, A.K.; Bacha, A.B.; Kotb, M. Etiology of Autistic Features: The Persisting Neurotoxic Effects of Propionic Acid. J. Neuroinflamm. 2012, 9, 74. [Google Scholar] [CrossRef]
- Müller-stierlin, A.S.; Teasdale, S.; Sabrina, M. Brain, Behavior & Immunity—Health Nutritional Psychiatry in the Treatment of Psychotic Disorders: Current Hypotheses and Research Challenges. Brain Behav. Immun. Health 2020, 19, 100070. [Google Scholar] [CrossRef]
- Kling, D.N.; Marcial, G.E.; Roberson, D.N.; Lorca, G.L.; Gonzalez, C.F. The Synergistic Contribution of Lactobacillus and Dietary Phytophenols in Host Health. In Probiotics and Prebiotics in Human Nutrition and Health; IntechOpen: London, UK, 2016. [Google Scholar]
- Bölte, S. Is Autism Curable? Dev. Med. Child Neurol. 2014, 56, 927–931. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, D.A.; Genuis, S.J.; Frye, R.E. Environmental Toxicants and Autism Spectrum Disorders: A Systematic Review. Transl. Psychiatry 2014, 4, e360-23. [Google Scholar] [CrossRef] [PubMed]
- Pugsley, K.; Scherer, S.W.; Bellgrove, M.A. Environmental Exposures Associated with Elevated Risk for Autism Spectrum Disorder May Augment the Burden of Deleterious de Novo Mutations among Probands. Mol. Psychiatry 2021, 27, 710–730. [Google Scholar] [CrossRef]
- Liu, X.; Lin, J.; Zhang, H.; Khan, N.U.; Zhang, J.; Tang, X. Oxidative Stress in Autism Spectrum Disorder—Current Progress of Mechanisms and Biomarkers. Front. Psychiatry 2022, 13, 813304. [Google Scholar] [CrossRef]
- Castejon, A.M.; Spaw, J.A.; Rozenfeld, I.; Sheinberg, N.; Kabot, S.; Shaw, A.; Hardigan, P.; Faillace, R.; Packer, E.E. Improving Antioxidant Capacity in Children With Autism: A Randomized, Double-Blind Controlled Study with Cysteine-Rich Whey Protein. Front. Psychiatry 2021, 12, 669089. [Google Scholar] [CrossRef]
- Dringen, R.; Gutterer, J.M.; Hirrlinger, J. Glutathione Metabolism in Brain Metabolic Interaction between Astrocytes and Neurons in the Defense against Reactive Oxygen Species. Eur. J. Biochem. 2000, 267, 4912–4916. [Google Scholar] [CrossRef]
- Al Suhaibani, A.; Ben Bacha, A.; Alonazi, M.; Bhat, R.S.; El-Ansary, A. Testing the Combined Effects of Probiotics and Prebiotics against Neurotoxic Effects of Propionic Acid Orally Administered to Rat Pups. Food Sci. Nutr. 2021, 9, 4440–4451. [Google Scholar] [CrossRef]
- Macfabe, D.F.; Cain, N.E.; Boon, F.; Ossenkopp, K.; Cain, D.P. Effects of the Enteric Bacterial Metabolic Product Propionic Acid on Object-Directed Behavior, Social Behavior, Cognition, and Neuroinflammation in Adolescent Rats: Relevance to Autism Spectrum Disorder. Behav. Brain Res. 2011, 217, 47–54. [Google Scholar] [CrossRef]
- Gjorgievski, N.; Tomovska, J.; Dimitrovska, G.; Makarijoski, B.; Shariati, M.A. Determination of The Antioxidant Activity in Yogurt. J. Hyg. Eng. Des. 2014, 8, 88–91. [Google Scholar]
- Ben Salem, M.; Affes, H.; Athmouni, K.; Ksouda, K.; Dhouibi, R.; Sahnoun, Z.; Hammami, S.; Zeghal, K.M. Chemicals Compositions, Antioxidant and Anti-Inflammatory Activity of Cynara Scolymus Leaves Extracts, and Analysis of Major Bioactive Polyphenols by HPLC. Evid.-Based Complement. Altern. Med. 2017, 2017, 4951937. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Lee, J.W.; Lim, S.D. The Probiotic Characteristics and GABA Production of Lactobacillus Plantarum K154 Isolated from Kimchi. Food Sci. Biotechnol. 2014, 23, 1951–1957. [Google Scholar] [CrossRef]
- Kochalska, K.; Oakden, W.; Słowik, T.; Chudzik, A.; Pankowska, A.; Łazorczyk, A.; Kozioł, P.; Andres-Mach, M.; Pietura, R.; Rola, R.; et al. Dietary Supplementation with Lactobacillus Rhamnosus JB-1 Restores Brain Neurochemical Balance and Mitigates the Progression of Mood Disorder in a Rat Model of Chronic Unpredictable Mild Stress. Nutr. Res. 2020, 82, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Bjorklund, G.; Saad, K.; Chirumbolo, S.; Kern, J.K.; Geier, D.A.; Geier, M.R.; Urbina, M.A. Immune Dysfunction and Neuroinflammation in Autism Spectrum Disorder. Acta Neurobiol. Exp. 2016, 76, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Lucchina, L.; Depino, A.M. Altered Peripheral and Central Inflammatory Responses in a Mouse Model of Autism. Autism Res. 2014, 7, 273–289. [Google Scholar] [CrossRef]
- Abdelli, L.S.; Samsam, A.; Naser, S.A. Propionic Acid Induces Gliosis and Neuro-Inflammation through Modulation of PTEN/AKT Pathway in Autism Spectrum Disorder. Sci. Rep. 2019, 9, 8824. [Google Scholar] [CrossRef]
- Baroja, M.L.; Kirjavainen, P.V.; Hekmat, S.; Reid, G. Anti-Inflammatory Effects of Probiotic Yogurt in Inflammatory Bowel Disease Patients. Clin. Exp. Immunol. 2007, 149, 470–479. [Google Scholar] [CrossRef]
- Putt, K.K.; Pei, R.; White, H.M.; Bolling, B.W. Yogurt Inhibits Intestinal Barrier Dysfunction in Caco-2 Cells by Increasing Tight Junctions. Food Funct. 2017, 8, 406–414. [Google Scholar] [CrossRef]
- Chaves, S.; Perdigon, G.; De Moreno De Leblanc, A. Yoghurt Consumption Regulates the Immune Cells Implicated in Acute Intestinal Inflammation and Prevents the Recurrence of the Inflammatory Process in a Mouse Model. J. Food Prot. 2011, 74, 801–811. [Google Scholar] [CrossRef]
- Gobbato, N.; Rachid, M.; Perdigón, G. Anti-Inflammatory Effect of Yoghurt in an Experimental Inflammatory Bowel Disease in Mouse. J. Dairy Res. 2008, 75, 497–504. [Google Scholar] [CrossRef]
- Al-Ayadhi, L.; Zayed, N.; Bhat, R.S.; Moubayed, N.M.S.; Al-Muammar, M.N.; El-Ansary, A. The Use of Biomarkers Associated with Leaky Gut as a Diagnostic Tool for Early Intervention in Autism Spectrum Disorder: A Systematic Review. Gut Pathog. 2021, 13, 54. [Google Scholar] [CrossRef] [PubMed]
- Sabater, C.; Molina-Tijeras, J.A.; Vezza, T.; Corzo, N.; Montilla, A.; Utrilla, P. Intestinal Anti-Inflammatory Effects of Artichoke Pectin and Modified Pectin Fractions in the Dextran Sulfate Sodium Model of Mice Colitis. Artificial Neural Network Modelling of Inflammatory Markers. Food Funct. 2019, 10, 7793–7805. [Google Scholar] [CrossRef] [PubMed]
- Wauquier, F.; Boutin-Wittrant, L.; Viret, A.; Guilhaudis, L.; Oulyadi, H.; Bourafai-Aziez, A.; Charpentier, G.; Rousselot, G.; Cassin, E.; Descamps, S.; et al. Metabolic and Anti-Inflammatory Protective Properties of Human Enriched Serum Following Artichoke Leaf Extract Absorption: Results from an Innovative Ex Vivo Clinical Trial. Nutrients 2021, 13, 2653. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C.J. Genes and Molecules of Lactobacilli Supporting Probiotic Action. Microbiol. Mol. Biol. Rev. 2008, 72, 728–764. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Claes, I.J.J.; Verhoeven, T.L.A.; Vanderleyden, J.; Keersmaecker, S.C.J. De Exopolysaccharides of Lactobacillus Rhamnosus GG Form a Protective Shield against Innate Immune. Microb. Biotechnol. 2010, 4, 368–374. [Google Scholar] [CrossRef]
- Corr, S.C.; Hill, C.; Gahan, C.G.M. Understanding the Mechanisms by Which Probiotics Inhibit Gastrointestinal Pathogens. Adv Food Nutr. Res. 2009, 56, 1–15. [Google Scholar]
- Mack, D.R.; Ahrne, S.; Hyde, L.; Wei, S.; Hollingsworth, M.A. Extracellular MUC3 Mucin Secretion Follows Adherence Of. Gut 2003, 52, 827–834. [Google Scholar] [CrossRef]
- Wells, J.M. Immunomodulatory Mechanisms of Lactobacilli. Microb. Cell Factories 2011, 10, S17. [Google Scholar] [CrossRef]
- Abreu, M.T. Erratum: Toll-like Receptor Signalling in the Intestinal Epithelium: How Bacterial Recognition Shapes Intestinal Function (Nature Reviews Immunology (2010) 10 (131-144)). Nat. Rev. Immunol. 2010, 10, 215. [Google Scholar] [CrossRef]
- Ayyanna, R.; Ankaiah, D.; Arul, V. Anti-Inflammatory and Antioxidant Properties of Probiotic Bacterium Lactobacillus Mucosae AN1 and Lactobacillus Fermentum SNR1 in Wistar Albino Rats. Front. Microbiol. 2018, 14, 3063. [Google Scholar] [CrossRef]
- Oksaharju, A.; Kooistra, T.; Kleemann, R.; Van Duyvenvoorde, W.; Miettinen, M.; Lappalainen, J.; Lindstedt, K.A.; Kovanen, P.T.; Korpela, R.; Kekkonen, R.A. Effects of Probiotic Lactobacillus Rhamnosus GG and Propionibacterium Freudenreichii Ssp. Shermanii JS Supplementation on Intestinal and Systemic Markers of Inflammation in ApoE*3Leiden Mice Consuming a High-Fat Diet. Br. J. Nutr. 2013, 110, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.C.W.; Gorbovskaya, I.; Hahn, M.K.; Müller, D.J. The Gut Microbiome in Schizophrenia and the Potential Benefits of Prebiotic and Probiotic Treatment. Nutrients 2021, 13, 1152. [Google Scholar] [CrossRef] [PubMed]
- Mcguinness, A.J.; Davis, J.A.; Dawson, S.L.; Loughman, A.; Collier, F.; Hely, M.O.; Simpson, C.A.; Green, J. OPEN A Systematic Review of Gut Microbiota Composition in Observational Studies of Major Depressive Disorder, Bipolar Disorder and Schizophrenia. Mol. Psychiatry 2022, 27, 1920–1935. [Google Scholar] [CrossRef] [PubMed]
- Ligezka, A.N.; Sonmez, A.I.; Corral-frias, M.P.; Golebiowski, R.; Lynch, B.; Croarkin, P.E.; Romanowicz, M. A systematic review of microbiome changes and impact of probiotic supplementation in children and adolescents with neuropsychiatric disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 8, 110187. [Google Scholar] [CrossRef]
- Shahrbabaki, M.E.; Sabouri, S.; Sabahi, A.; Barfeh, D.; Divsalar, P. The Efficacy of Probiotics for Treatment of Bipolar Disorder-Type 1: A Randomized, Double-Blind, Placebo Controlled Trial. Iran J. Psychiatry 2020, 15, 10–16. [Google Scholar] [CrossRef]
- Nahm, F.S. Receiver Operating Characteristic Curve: Overview and Practical Use for Clinicians. Korean J. Anesthesiol. 2022, 75, 25–36. [Google Scholar] [CrossRef]
- Issazadeh, K.; Ali Abadi, M.A.; Kazemi Darsanaki, R.; Alikhani, F.; Dadras, H.; Tajehmiri, A. Isolation, Identification and Analysis of Probiotic Properties of Lactobacillus spp. from Traditional Yoghurts in North of Iran. J. Pure Appl. Microbiol. 2013, 7, 2965–2971. [Google Scholar]
- Kusuma, G.D.; Paseephol, T.; Sherkat, F. Prebiotic and Rheological Effects of Jerusalem Artichoke Inulin in Low-Fat Yogurt. Aust. J. Dairy Technol. 2009, 64, 59–163. [Google Scholar]
- Heidarian, E.; Soofiniya, Y. Hypolipidemic and Hypoglycemic Effects of Aerial Part of Cynara Scolymus in Streptozotocin-Induced Diabetic Rats. J. Med. Plants Res. 2011, 5, 2717–2723. [Google Scholar]
- Alghamdi, M.A.; Al-ayadhi, L.; Hassan, W.M.; Bhat, R.S.; Alonazi, M.A.; El-ansary, A. Bee Pollen and Probiotics May Alter Brain Neuropeptide Levels in a Rodent Model of Autism Spectrum Disorders. Metabolites 2022, 12, 562. [Google Scholar] [CrossRef]
- Abu-Elsaad, N.; El-Karef, A. Protection against Nonalcoholic Steatohepatitis through Targeting IL-18 and IL-1alpha by Luteolin. Pharmacol. Rep. 2019, 71, 688–694. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Parameters | Groups | Mean ± S.D. | p- Value |
|---|---|---|---|
| GPX | Control | 108.60 ± 11.05 | 0.001 |
| PPA | 75.19 ± 9.72 a | ||
| Yogurt | 107.33 ± 15.44 b | ||
| Artichoke | 106.21 ± 18.80 b | ||
| L. rhamnosus GG + Luteolin | 95.48 ± 14.47 b | ||
| GSH | Control | 15.18 ± 2.27 | 0.001 |
| PPA | 8.13 ± 1.82 a | ||
| Yogurt | 13.75 ± 2.97 b | ||
| Artichoke | 11.24 ± 2.08 ab | ||
| L. rhamnosus GG + Luteolin | 9.97 ± 1.75 ab | ||
| GABA | Control | 65.91 ± 10.41 | 0.004 |
| PPA | 35.80 ± 5.82 a | ||
| Yogurt | 52.33 ± 18.13 b | ||
| Artichoke | 43.29 ± 12.07 a | ||
| L. rhamnosus GG + Luteolin | 46.22 ± 17.77 a |
| Parameters | Groups | Mean ± S.D. | p-Value |
|---|---|---|---|
| TNF-α | Control | 14.40 ± 3.53 | 0.001 |
| PPA | 37.53 ± 8.89 a | ||
| Yogurt | 29.85 ± 6.84 ab | ||
| Artichoke | 29.86 ± 7.93 ab | ||
| L. rhamnosus GG + Luteolin | 21.69 ± 4.67 ab | ||
| IL10 | Control | 49.74 ± 3.50 | 0.331 |
| PPA | 40.11 ± 6.76 | ||
| Yogurt | 38.41 ± 2.50 a | ||
| Artichoke | 43.07 ± 7.64 | ||
| L. rhamnosus GG + Luteolin | 46.79 ± 5.03 | ||
| IL-6 | Control | 10.86 ± 0.80 | 0.001 |
| PPA | 47.69 ± 9.33 a | ||
| Yogurt | 36.60 ± 8.39 ab | ||
| Artichoke | 36.61 ± 9.73 ab | ||
| L. rhamnosus GG + Luteolin | 26.60 ± 5.73 ab |
| Groups | AUC | Cut-Off Value | Sensitivity % | Specificity % | p- Value | |
|---|---|---|---|---|---|---|
| GPX1 | PPA | 1.000 | 92.785 | 100.0% | 100.0% | 0.004 |
| Yogurt | 0.833 | 95.660 | 66.7% | 100.0% | 0.055 | |
| Artichoke | 0.667 | 99.800 | 50.0% | 83.3% | 0.337 | |
| L. rhamnosus GG + Luteolin | 0.667 | 95.780 | 50.0% | 100.0% | 0.337 | |
| GSH | PPA | 1.000 | 11.905 | 100.0% | 100.0% | 0.004 |
| Yogurt | 0.889 | 12.095 | 66.7% | 100.0% | 0.025 | |
| Artichoke | 1.000 | 12.860 | 100.0% | 100.0% | 0.004 | |
| L. rhamnosus GG + Luteolin | 1.000 | 11.865 | 100.0% | 100.0% | 0.004 | |
| GABA | PPA | 0.889 | 60.800 | 100.0% | 83.3% | 0.025 |
| Yogurt | 0.833 | 60.005 | 83.3% | 83.3% | 0.055 | |
| Artichoke | 0.917 | 62.190 | 100.0% | 83.3% | 0.016 | |
| L. rhamnosus GG + Luteolin | 0.972 | 62.190 | 100.0% | 83.3% | 0.006 |
| Groups | AUC | Cut-Off Value | Sensitivity % | Specificity % | p- Value | |
|---|---|---|---|---|---|---|
| TNF-α | PPA | 1.000 | 26.105 | 100.0% | 100.0% | 0.004 |
| Yogurt | 0.972 | 17.235 | 100.0% | 83.3% | 0.006 | |
| Artichoke | 0.861 | 18.955 | 66.7% | 100.0% | 0.037 | |
| L. rhamnosus GG + Luteolin | 0.861 | 19.090 | 83.3% | 100.0% | 0.037 | |
| IL-10 | PPA | 1.000 | 87.335 | 100.0% | 100.0% | 0.004 |
| Yogurt | 0.944 | 72.205 | 83.3% | 100.0% | 0.010 | |
| Artichoke | 0.917 | 57.470 | 100.0% | 66.7% | 0.016 | |
| L. rhamnosus GG + Luteolin | 0.722 | 58.015 | 83.3% | 66.7% | 0.200 | |
| IL-6 | PPA | 1.000 | 26.610 | 100.0% | 100.0% | 0.004 |
| Yogurt | 1.000 | 17.240 | 100.0% | 100.0% | 0.004 | |
| Artichoke | 1.000 | 14.960 | 100.0% | 100.0% | 0.004 | |
| L. rhamnosus\GG + Luteolin | 1.000 | 14.125 | 100.0% | 100.0% | 0.004 |
| Group | No. of Rats | Pro-/Prebiotic Nutritional Interventions | Dosage |
|---|---|---|---|
| 1 a | 6 | - | - |
| 2 b | 6 | - | (250 mg PPA/kg BW) for 3 days |
| 3 | 6 | Yogurt c | (3 mL/kg BW) |
| 4 | 6 | Artichoke d | (400 mL/kg BW) |
| 5 | 6 | Luteolin and L. rhamnosus GG e | (50 mg/kg BW) and 0.2 mL (1 × 109 CFU) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsubaiei, S.R.M.; Alfawaz, H.A.; Almubarak, A.Y.; Alabdali, N.A.; Ben Bacha, A.; El-Ansary, A. Independent and Combined Effects of Probiotics and Prebiotics as Supplements or Food-Rich Diets on a Propionic-Acid-Induced Rodent Model of Autism Spectrum Disorder. Metabolites 2023, 13, 50. https://doi.org/10.3390/metabo13010050
Alsubaiei SRM, Alfawaz HA, Almubarak AY, Alabdali NA, Ben Bacha A, El-Ansary A. Independent and Combined Effects of Probiotics and Prebiotics as Supplements or Food-Rich Diets on a Propionic-Acid-Induced Rodent Model of Autism Spectrum Disorder. Metabolites. 2023; 13(1):50. https://doi.org/10.3390/metabo13010050
Chicago/Turabian StyleAlsubaiei, Sana Razhan M., Hanan A. Alfawaz, Abdullah Yaseen Almubarak, Nouf Ahmed Alabdali, Abir Ben Bacha, and Afaf El-Ansary. 2023. "Independent and Combined Effects of Probiotics and Prebiotics as Supplements or Food-Rich Diets on a Propionic-Acid-Induced Rodent Model of Autism Spectrum Disorder" Metabolites 13, no. 1: 50. https://doi.org/10.3390/metabo13010050
APA StyleAlsubaiei, S. R. M., Alfawaz, H. A., Almubarak, A. Y., Alabdali, N. A., Ben Bacha, A., & El-Ansary, A. (2023). Independent and Combined Effects of Probiotics and Prebiotics as Supplements or Food-Rich Diets on a Propionic-Acid-Induced Rodent Model of Autism Spectrum Disorder. Metabolites, 13(1), 50. https://doi.org/10.3390/metabo13010050