
Mycorrhiza-Tree-Herbivore Interactions: Alterations in Poplar Metabolome and Volatilome

 , ,
, ,  and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
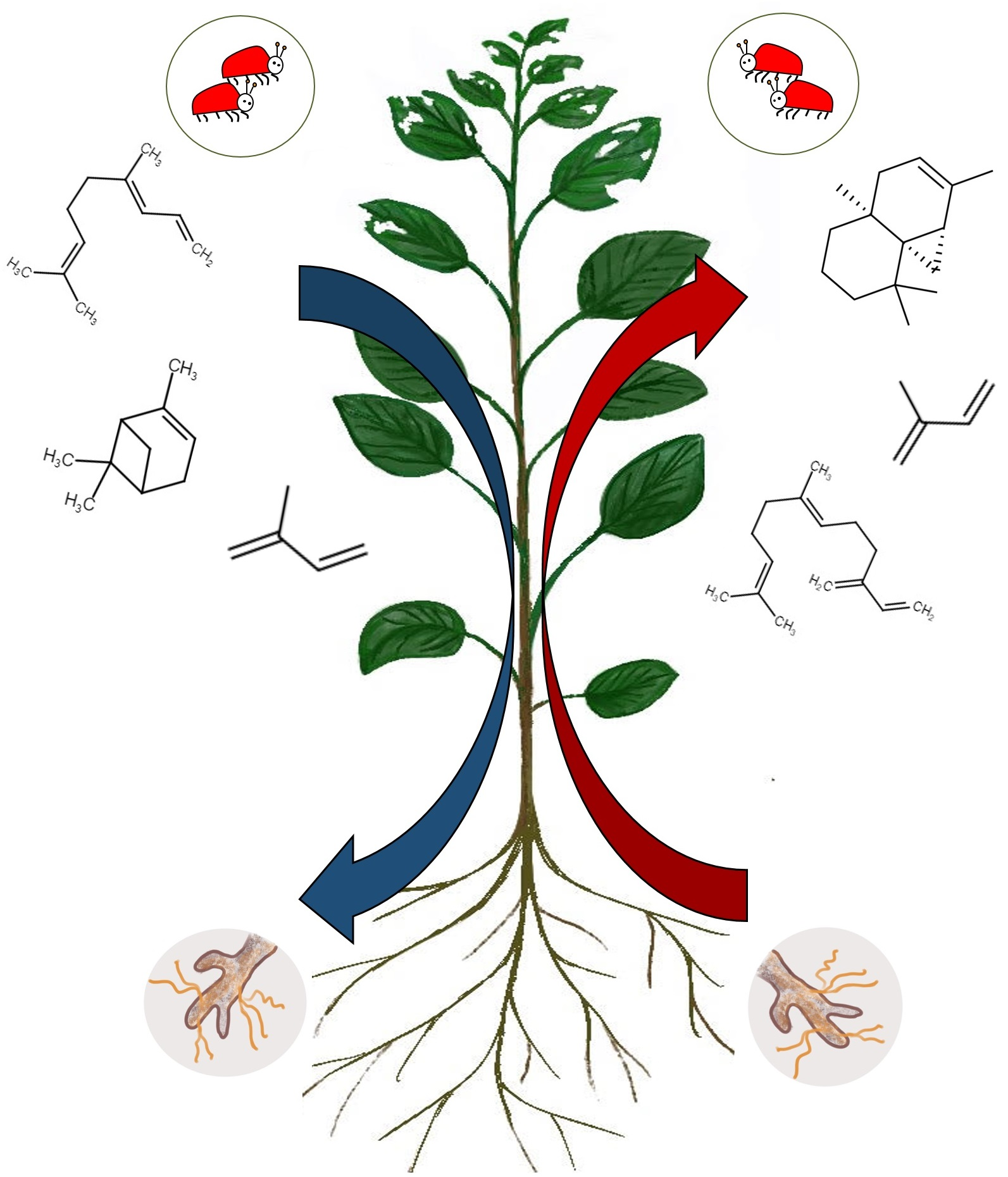
1. Introduction
2. Results
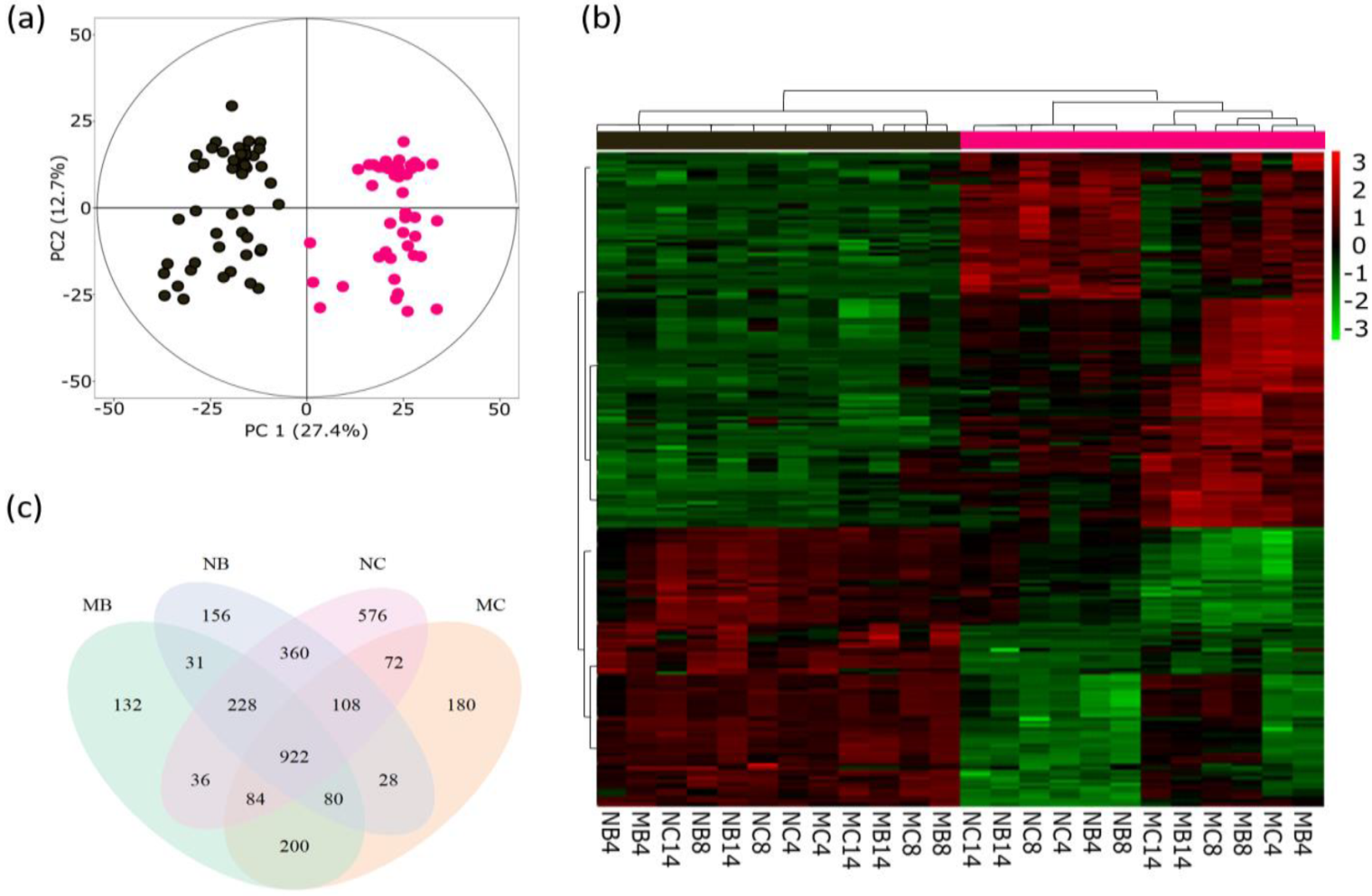
2.1. Mycorrhization and Herbivory Effects on the Poplar Metabolome
2.1.1. Young/Infested and Old/Non-Infested Leaves Show Different Metabolomic Changes
2.1.2. Mycorrhiza and Herbivory Effects on Metabolomes of Different-Aged Leaves
Mycorrhiza and Herbivory Effects on Young/Infested Leaves
Mycorrhiza and Herbivory Effects on Old Leaves
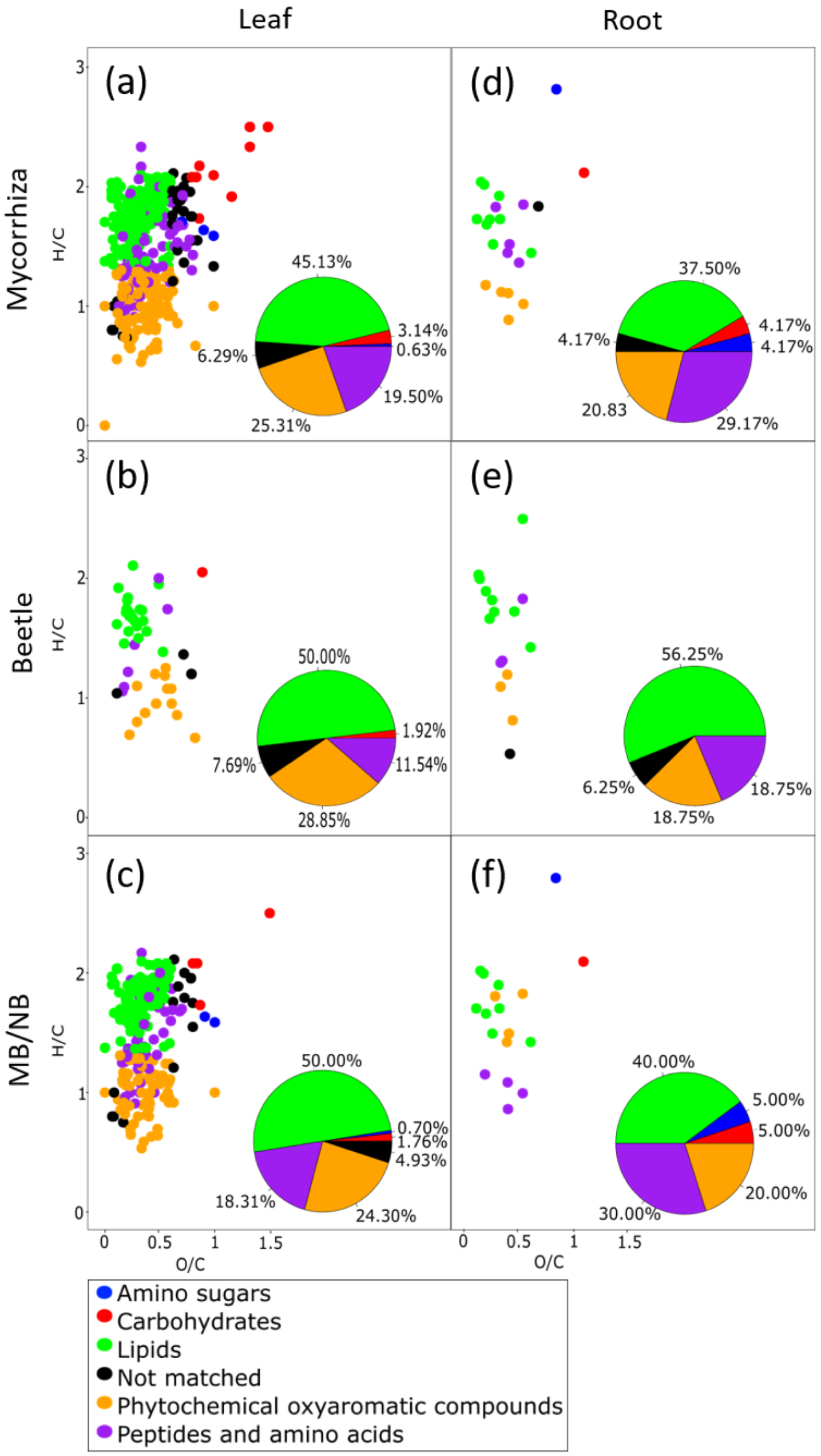
2.1.3. Mycorrhiza and Herbivory Effects on Poplar Root Metabolome
2.2. Mycorrhization Affects the Poplar Defense Response to Beetle Infestation
2.3. The Herbivore-Induced Poplar VOCs Are Released in Different Offsets from Leaves of Mycorrhized and Non-Mycorrhized Trees
3. Discussion
3.1. EMF Affect Differently the Metabolomes of Young and Old Poplar Leaves
3.2. Herbivory Induced Changes in Volatile Metabolome of Infested Leaves
3.3. EMF Alters the Poplar Response to Herbivory
3.4. Conclusions
4. Materials and Methods
4.1. Plant Material and Fungal and Herbivore Treatments
4.2. VOC Collection
4.3. Harvest of the Plant Material
4.4. Metabolite Extraction
4.5. UPLC-qToF-MS Measurements and Data Analysis
4.6. Statistical Analysis and Visualization Tools
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Šimpraga, M.; Ghimire, R.P.; Van Der Straeten, D.; Blande, J.D.; Kasurinen, A.; Sorvari, J.; Holopainen, T.; Adriaenssens, S.; Holopainen, J.K.; Kivimaenpaa, M. Unravelling the functions of biogenic volatiles in boreal and temperate forest ecosystems. Eur. J. For. Res. 2019, 138, 763–787. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahebi, M.; Hanafi, M.M.; van Wijnen, A.J.; Akmar, A.S.N.; Azizi, P.; Idris, A.S.; Taheri, S.; Foroughi, M. Profiling secondary metabolites of plant defence mechanisms and oil palm in response to Ganoderma boninense attack. Int. Biodeter. Biodegr. 2017, 122, 151–164. [Google Scholar] [CrossRef]
- Arimura, G.; Kost, C.; Boland, W. Herbivore-induced, indirect plant defences. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2005, 1734, 91–111. [Google Scholar] [CrossRef]
- Brilli, F.; Ciccioli, P.; Frattoni, M.; Prestininzi, M.; Spanedda, A.F.; Loreto, F. Constitutive and herbivore-induced monoterpenes emitted by Populus x euroamericana leaves are key volatiles that orient Chrysomela populi beetles. Plant Cell Environ. 2009, 32, 542–552. [Google Scholar] [CrossRef]
- Bonfante, P.; Genre, A. Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis. Nat. Commun. 2010, 1. [Google Scholar] [CrossRef] [Green Version]
- Finlay, R.D. Ecological aspects of mycorrhizal symbiosis: With special emphasis on the functional diversity of interactions involving the extraradical mycelium. J. Exp. Bot. 2008, 59, 1115–1126. [Google Scholar] [CrossRef]
- Johansson, J.F.; Paul, L.R.; Finlay, R.D. Microbial interactions in the mycorrhizosphere and their significance for sustainable agriculture. FEMS Microbiol. Ecol. 2004, 48, 1–13. [Google Scholar] [CrossRef]
- Ruyter-Spira, C.; Al-Babili, S.; van der Krol, S.; Bouwmeester, H. The biology of strigolactones. Trends Plant Sci. 2013, 18, 72–83. [Google Scholar] [CrossRef]
- Cope, K.R.; Bascaules, A.; Irving, T.B.; Venkateshwaran, M.; Maeda, J.; Garcia, K.; Rush, T.A.; Ma, C.; Labbe, J.; Jawdy, S.; et al. The ectomycorrhizal fungus Laccaria bicolor produces lipochitooligosaccharides and uses the common symbiosis pathway to colonize Populus roots. Plant Cell 2019, 31, 2386–2410. [Google Scholar] [CrossRef] [Green Version]
- Maillet, F.; Poinsot, V.; Andre, O.; Puech-Pages, V.; Haouy, A.; Gueunier, M.; Cromer, L.; Giraudet, D.; Formey, D.; Niebel, A.; et al. Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhiza. Nature 2011, 469, 58-U1501. [Google Scholar] [CrossRef]
- Ditengou, F.A.; Müller, A.; Rosenkranz, M.; Felten, J.; Lasok, H.; van Doorn, M.M.; Legue, V.; Palme, K.; Schnitzler, J.P.; Polle, A. Volatile signalling by sesquiterpenes from ectomycorrhizal fungi reprogrammes root architecture. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef]
- Luo, Z.B.; Li, K.; Gai, Y.; Gobel, C.; Wildhagen, H.; Jiang, X.N.; Feussner, I.; Rennenberg, H.; Polle, A. The ectomycorrhizal fungus (Paxillus involutus) modulates leaf physiology of poplar towards improved salt tolerance. Environ. Exp. Bot. 2011, 72, 304–311. [Google Scholar] [CrossRef]
- Dreischhoff, S.; Das, I.S.; Jakobi, M.; Kasper, K.; Polle, A. Local responses and systemic induced resistance mediated by ectomycorrhizal fungi. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Kaling, M.; Schmidt, A.; Moritz, F.; Rosenkranz, M.; Witting, M.; Kasper, K.; Janz, D.; Schmitt-Kopplin, P.; Schnitzler, J.P.; Polle, A. Mycorrhiza-triggered transcriptomic and metabolomic networks impinge on herbivore fitness. Plant Physiol. 2018, 176, 2639–2656. [Google Scholar] [CrossRef] [Green Version]
- Gange, A.C.; Gane, D.R.J.; Chen, Y.L.; Gong, M.Q. Dual colonization of Eucalyptus urophylla ST Blake by arbuscular and ectomycorrhizal fungi affects levels of insect herbivore attack. Agric. For. Entomol. 2005, 7, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Rieske, L.K.; Rhoades, C.C.; Miller, S.P. Foliar chemistry and gypsy moth, Lymantria dispar (L.), herbivory on pure American chestnut, Castanea dentata (Fam: Fagaceae), and a disease-resistant hybrid. Environ. Entomol. 2003, 32, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Oddsdottir, E.S.; Eilenberg, J.; Sen, R.; Harding, S.; Halldorsson, G. Early reduction of Otiorhynchus spp. larval root herbivory on Betula pubescens by beneficial soil fungi. Appl. Soil Ecol. 2010, 45, 168–174. [Google Scholar] [CrossRef]
- Song, Y.Y.; Chen, D.M.; Lu, K.; Sun, Z.X.; Zeng, R.S. Enhanced tomato disease resistance primed by arbuscular mycorrhizal fungus. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Adolfsson, L.; Nziengui, H.; Abreu, I.N.; Simura, J.; Beebo, A.; Herdean, A.; Aboalizadeh, J.; Siroka, J.; Moritz, T.; Novak, O.; et al. Enhanced secondary- and hormone metabolism in leaves of arbuscular mycorrhizal Medicago truncatula. Plant Physiol. 2017, 175, 392–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerlach, N.; Schmitz, J.; Polatajko, A.; Schluter, U.; Fahnenstich, H.; Witt, S.; Fernie, A.R.; Uroic, K.; Scholz, U.; Sonnewald, U.; et al. An integrated functional approach to dissect systemic responses in maize to arbuscular mycorrhizal symbiosis. Plant Cell Environ. 2015, 38, 1591–1612. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.J.; Harding, S.A.; Tschaplinski, T.J.; Lindroth, R.L.; Yuan, Y.N. Genome-wide analysis of the structural genes regulating defense phenylpropanoid metabolism in Populus. New Phytol. 2006, 172, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Boeckler, G.A.; Gershenzon, J.; Unsicker, S.B. Phenolic glycosides of the Salicaceae and their role as anti-herbivore defenses. Phytochemistry 2011, 72, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Irmisch, S.; McCormick, A.C.; Boeckler, G.A.; Schmidt, A.; Reichelt, M.; Schneider, B.; Block, K.; Schnitzler, J.P.; Gershenzon, J.; Unsicker, S.B.; et al. Two herbivore-induced cytochrome P450 enzymes CYP79D6 and CYP79D7 catalyze the formation of volatile aldoximes involved in poplar defense. Plant Cell 2013, 25, 4737–4754. [Google Scholar] [CrossRef] [Green Version]
- Irmisch, S.; McCormick, A.C.; Gunther, J.; Schmidt, A.; Boeckler, G.A.; Gershenzon, J.; Unsicker, S.B.; Köllner, T.G. Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalist caterpillar. Plant J. 2014, 80, 1095–1107. [Google Scholar] [CrossRef]
- Papantoniou, D.V.F.; Weinhold, A.; Quijano, T.; Khakimov, B.; Pattison, D.I.; Bak, S.; van Dam, N.M.; Martínez-Medina, A. Cascading effects of root microbial symbiosis on the development and metabolome of the insect herbivore Manduca sexta L. Metabolites 2021, 11, 731. [Google Scholar] [CrossRef]
- Polle, A.; Douglas, C. The molecular physiology of poplars: Paving the way for knowledge-based biomass production. Plant Biol. 2010, 12, 239–241. [Google Scholar] [CrossRef]
- Müller, A.; Kaling, M.; Faubert, P.; Gort, G.; Smid, H.M.; Van Loon, J.J.A.; Dicke, M.; Kanawati, B.; Schmitt-Kopplin, P.; Polle, A.; et al. Isoprene emission by poplar is not important for the feeding behaviour of poplar leaf beetles. BMC Plant Biol. 2015, 15, 165. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.A.; Subhan, N.; Hossain, H.; Hossain, M.; Reza, H.M.; Rahman, M.M.; Ullah, M.O. Hydroxycinnamic acid derivatives: A potential class of natural compounds for the management of lipid metabolism and obesity. Nutr. Metab. 2016, 13. [Google Scholar] [CrossRef] [Green Version]
- Silva, E.C.; Abhayawardhana, P.L.; Lygin, A.V.; Robertson, C.L.; Liu, M.; Liu, Z.; Schneider, R.W. Coumestrol confers partial resistance in soybean plants against Cercospora leaf blight. Phytopathology 2018, 108, 935–947. [Google Scholar] [CrossRef] [Green Version]
- Fellenberg, C.; Corea, O.; Yan, L.H.; Archinuk, F.; Piirtola, E.M.; Gordon, H.; Reichelt, M.; Brandt, W.; Wulff, J.; Ehlting, J.; et al. Discovery of salicyl benzoate UDP-glycosyltransferase, a central enzyme in poplar salicinoid phenolic glycoside biosynthesis. Plant J. 2020, 102, 99–115. [Google Scholar] [CrossRef]
- Marasas, W.F.O. Fumonisins: History, world-wide occurrence and impact. Adv. Exp. Med. Biol. 1996, 392, 1–17. [Google Scholar]
- Goetschi, E.; Angehrn, P.; Gmuender, H.; Hebeisen, P.; Link, H.; Masciadri, R.; Nielsen, J. Cyclothialidine and its congeners—A new class of DNA gyrase inhibitors. Pharmacol Ther. 1993, 60, 367–380. [Google Scholar] [CrossRef]
- Schinkovitz, A.; Gibbons, S.; Stavri, M.; Cocksedge, M.J.; Bucar, F. Ostruthin: An antimycobacterial coumarin from the roots of Peucedanum ostruthium. Planta Med. 2003, 69, 369–371. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, S.; Kuroda, K.; Doi, K.I.; Tanabe, M.; Shibata, T.; Yoshida, T.; Hatano, T. Revised structures of gambiriins A1, A2, B1, and B2, chalcane-flavan dimers from gambir (Uncaria gambir extract). Chem. Pharm. Bull. 2007, 55, 268–272. [Google Scholar] [CrossRef] [Green Version]
- Rivas-Ubach, A.; Liu, Y.; Bianchi, T.S.; Tolic, N.; Jansson, C.; Pasa-Tolic, L. Moving beyond the van Krevelen diagram: A new stoichiometric approach for compound classification in organisms. Anal. Chem. 2018, 90, 6152–6160. [Google Scholar] [CrossRef]
- Song, C.X.; Zhu, F.; Carrion, V.J.; Cordovez, V. Beyond plant microbiome composition: Exploiting microbial functions and plant traits via integrated approaches. Front. Bioeng. Biotech. 2020, 8. [Google Scholar] [CrossRef]
- Lim, G.H.; Singhal, R.; Kachroo, A.; Kachroo, P. Fatty acid- and lipid-mediated signaling in plant defense. Ann. Rev. Phytopathol. 2017, 55, 505–536. [Google Scholar] [CrossRef]
- Langenfeld-Heyser, R.; Gao, J.; Ducic, T.; Tachd, P.; Lu, C.F.; Fritz, E.; Gafur, A.; Polle, A. Paxillus involutus mycorrhiza attenuate NaCl-stress responses in the salt-sensitive hybrid poplar Populus x canescens. Mycorrhiza 2007, 17, 121–131. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, J.; Li, H.; Shi, W.G.; Polle, A.; Lu, M.Z.; Sun, X.M.; Luo, Z.B. Global poplar root and leaf transcriptomes reveal links between growth and stress responses under nitrogen starvation and excess. Tree Physiol. 2015, 35, 1283–1302. [Google Scholar] [CrossRef] [Green Version]
- Morkunas, I.; Ratajczak, L. The role of sugar signaling in plant defense responses against fungal pathogens. Acta Physiol. Plant 2014, 36, 1607–1619. [Google Scholar] [CrossRef] [Green Version]
- Bolouri Moghaddam, M.R.; Van den Ende, W. Sugars and plant innate immunity. J. Exp. Bot. 2012, 63, 3989–3998. [Google Scholar] [CrossRef] [Green Version]
- Gebauer, P.; Korn, M.; Engelsdorf, T.; Sonnewald, U.; Koch, C.; Voll, L.M. Sugar accumulation in leaves of Arabidopsis sweet11/sweet12 double mutants enhances priming of the salicylic acid-mediated defense response. Front. Plant Sci. 2017, 8, 1378. [Google Scholar] [CrossRef] [Green Version]
- Dvorak, P.; Krasylenko, Y.; Zeiner, A.; Samaj, J.; Takac, T. Signaling toward reactive oxygen species-scavenging enzymes in plants. Front. Plant Sci. 2021, 11, 2178. [Google Scholar] [CrossRef]
- Cameron, D.D.; Neal, A.L.; van Wees, S.C.M.; Ton, J. Mycorrhiza-induced resistance: More than the sum of its parts? Trends Plant Sci. 2013, 18, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef]
- Vishwanathan, K.; Zienkiewicz, K.; Liu, Y.; Janz, D.; Feussner, I.; Polle, A.; Haney, C.H. Ectomycorrhizal fungi induce systemic resistance against insects on a nonmycorrhizal plant in a CERK1-dependent manner. New Phytol. 2020, 228, 728–740. [Google Scholar] [CrossRef]
- Keymer, A.; Pimprikar, P.; Wewer, V.; Huber, C.; Brands, M.; Bucerius, S.L.; Delaux, P.M.; Klingl, V.; von Ropenack-Lahaye, E.; Wang, T.L.; et al. Lipid transfer from plants to arbuscular mycorrhiza fungi. eLife 2017, 6. [Google Scholar] [CrossRef]
- Nakada, N.; Shimada, H.; Hirata, T.; Aoki, Y.; Kamiyama, T.; Watanabe, J.; Arisawa, M. Biological characterization of cyclothialidine, a new DNA gyrase inhibitor. Antimicrob. Agents Ch. 1993, 37, 2656–2661. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.B.; Janz, D.; Jiang, X.N.; Gobel, C.; Wildhagen, H.; Tan, Y.P.; Rennenberg, H.; Feussner, I.; Polle, A. Upgrading root physiology for stress tolerance by ectomycorrhizas: Insights from metabolite and transcriptional profiling into reprogramming for stress anticipation. Plant Physiol. 2009, 151, 1902–1917. [Google Scholar] [CrossRef] [Green Version]
- Lackus, N.D.; Muller, A.; Krober, T.D.U.; Reichelt, M.; Schmidt, A.; Nakamura, Y.; Paetz, C.; Luck, K.; Lindroth, R.L.; Constabel, C.P.; et al. The occurrence of sulfated salicinoids in poplar and their formation by sulfotransferase1. Plant Physiol. 2020, 183, 137–151. [Google Scholar] [CrossRef] [Green Version]
- Smiljkovic, M.; Stanisavljevic, D.; Stojkovic, D.; Petrovic, I.; Vicentic, J.M.; Popovic, J.; Grdadolnik, S.G.; Markovic, D.; Sankovic-Babic, S.; Glamoclija, J.; et al. Apigenin-7-O-glucoside versus apigenin: Insight into the modes of anticandidal and cytotoxic actions. EXCLI. J. 2017, 16, 795–807. [Google Scholar] [CrossRef]
- Hamade, K.; Fliniaux, O.; Fontaine, J.X.; Molinie, R.; Nnang, E.O.; Bassard, S.; Guenin, S.; Gutierrez, L.; Laine, E.; Hano, C.; et al. NMR and LC-MS-based metabolomics to study osmotic stress in lignan-deficient flax. Molecules 2021, 26, 767. [Google Scholar] [CrossRef]
- Rosenkranz, M.; Chen, Y.Y.; Zhu, P.Y.; Vlot, A.C. Volatile terpenes—Mediators of plant-to-plant communication. Plant. J. 2021, 108, 617–631. [Google Scholar] [CrossRef]
- McCormick, A.C.; Irmisch, S.; Reinecke, A.; Boeckler, G.A.; Veit, D.; Reichelt, M.; Hansson, B.S.; Gershenzon, J.; Köllner, T.G.; Unsicker, S.B. Herbivore-induced volatile emission in black poplar: Regulation and role in attracting herbivore enemies. Plant Cell Environ. 2014, 37, 1909–1923. [Google Scholar] [CrossRef]
- Šimpraga, M.; Takabayashi, J.; Holopainen, J.K. Language of plants: Where is the word? J. Integr. Plant. Biol. 2016, 58, 343–349. [Google Scholar] [CrossRef] [Green Version]
- Vlot, A.C.; Sales, J.H.; Lenk, M.; Bauer, K.; Brambilla, A.; Sommer, A.; Chen, Y.Y.; Wenig, M.; Nayem, S. Systemic propagation of immunity in plants. New Phytol. 2021, 229, 1234–1250. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Blande, J.D. Where do herbivore-induced plant volatiles go? Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- McCormick, A.C.; Irmisch, S.; Boeckler, G.A.; Gershenzon, J.; Köllner, T.G.; Unsicker, S.B. Herbivore-induced volatile emission from old-growth black poplar trees under field conditions. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Goldhaber-Pasillas, G.D.; Mustafa, N.R.; Verpoorte, R. Jasmonic acid effect on the fatty acid and terpenoid indole alkaloid accumulation in cell suspension cultures of Catharanthus roseus. Molecules 2014, 19, 10242–10260. [Google Scholar] [CrossRef] [Green Version]
- Ullah, C.; Tsai, C.J.; Unsicker, S.B.; Xue, L.J.; Reichelt, M.; Gershenzon, J.; Hammerbacher, A. Salicylic acid activates poplar defense against the biotrophic rust fungus Melampsora larici-populina via increased biosynthesis of catechin and proanthocyanidins. New Phytol. 2019, 221, 960–975. [Google Scholar] [CrossRef] [Green Version]
- Runguphan, W.; Maresh, J.J.; O’Connor, S.E. Silencing of tryptamine biosynthesis for production of nonnatural alkaloids in plant culture. Proc. Natl. Acad. Sci. USA 2009, 106, 13673–13678. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, S.E.; Maresh, J.J. Chemistry and biology of monoterpene indole alkaloid biosynthesis. Nat. Prod. Rep. 2006, 23, 532–547. [Google Scholar] [CrossRef]
- Markovic, D.; Colzi, I.; Taiti, C.; Ray, S.; Scalone, R.; Ali, J.G.; Mancuso, S.; Ninkovic, V. Airborne signals synchronize the defenses of neighboring plants in response to touch. J. Exp. Bot. 2019, 70, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Brilli, F.; Loreto, F.; Baccelli, I. Exploiting plant volatile organic compounds (VOCs) in agriculture to improve sustainable defense strategies and productivity of crops. Front. Plant Sci. 2019, 10, 264. [Google Scholar] [CrossRef]
- Müller, A.; Volmer, K.; Mishra-Knyrim, M.; Polle, A. Growing poplars for research with and without mycorrhizas. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Dool, H.K.P. Generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. 1962, 11, 463–471. [Google Scholar] [CrossRef]
- Eriksson, L.; Byrne, T.; Johansson, E.; Trygg, J.; Vikström, C. Multi-and Megavariate Data Analysis Basic Principles and Applications; Umetrics Academy: Washington, DC, USA, 2013; Volume 1. [Google Scholar]
- van den Berg, R.A.; Hoefsloot, H.C.J.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksson, L.; Trygg, J.; Wold, S. CV-ANOVA for significance testing of PLS and OPLS® models. J. Chemom. 2008, 22, 594–600. [Google Scholar] [CrossRef]
- Cocchi, M.B.A.; Marini, F. Chemometric methods for classification and feature selection. In Comprehensive Analytical Chemistry; Jaumot, J.B.C., Tauler, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate– A practical and powerful approach to multiple testing. J. R. Statist. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Bertic, M.; Schroeder, H.; Kersten, B.; Fladung, M.; Orgel, F.; Buegger, F.; Schnitzler, J.P.; Ghirardo, A. European oak chemical diversity—From ecotypes to herbivore resistance. New Phytol. 2021, 232, 818–834. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sivaprakasam Padmanaban, P.B.; Rosenkranz, M.; Zhu, P.; Kaling, M.; Schmidt, A.; Schmitt-Kopplin, P.; Polle, A.; Schnitzler, J.-P. Mycorrhiza-Tree-Herbivore Interactions: Alterations in Poplar Metabolome and Volatilome. Metabolites 2022, 12, 93. https://doi.org/10.3390/metabo12020093
Sivaprakasam Padmanaban PB, Rosenkranz M, Zhu P, Kaling M, Schmidt A, Schmitt-Kopplin P, Polle A, Schnitzler J-P. Mycorrhiza-Tree-Herbivore Interactions: Alterations in Poplar Metabolome and Volatilome. Metabolites. 2022; 12(2):93. https://doi.org/10.3390/metabo12020093
Chicago/Turabian StyleSivaprakasam Padmanaban, Prasath Balaji, Maaria Rosenkranz, Peiyuan Zhu, Moritz Kaling, Anna Schmidt, Philippe Schmitt-Kopplin, Andrea Polle, and Jörg-Peter Schnitzler. 2022. "Mycorrhiza-Tree-Herbivore Interactions: Alterations in Poplar Metabolome and Volatilome" Metabolites 12, no. 2: 93. https://doi.org/10.3390/metabo12020093
APA StyleSivaprakasam Padmanaban, P. B., Rosenkranz, M., Zhu, P., Kaling, M., Schmidt, A., Schmitt-Kopplin, P., Polle, A., & Schnitzler, J.-P. (2022). Mycorrhiza-Tree-Herbivore Interactions: Alterations in Poplar Metabolome and Volatilome. Metabolites, 12(2), 93. https://doi.org/10.3390/metabo12020093