
The Cholesterol Metabolite Cholest-5-en-3-One Alleviates Hyperglycemia and Hyperinsulinemia in Obese (db/db) Mice


Abstract
1. Introduction
2. Results
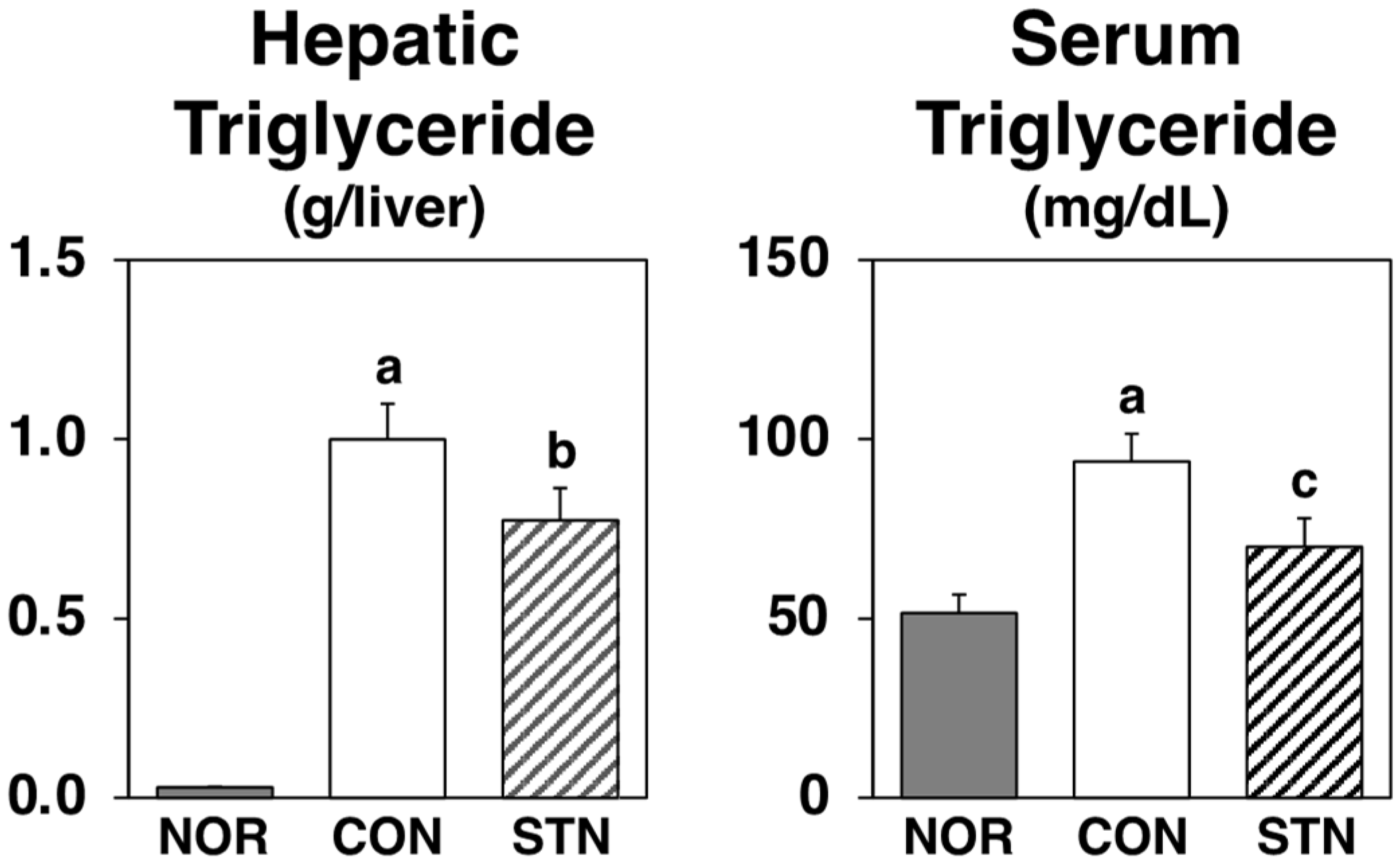
2.1. Effects of Experimental Diets on Growth Parameters, Hepatic Triglyceride, and Serum Triglyceride
2.2. Effects of Experimental Diets on Glucose and Insulin Levels in the Serum
2.3. Effects of Experimental Diets on Adipocytokine Levels in the Serum
2.4. Effects of Experimental Diets on mRNA Levels in Perirenal White Adipose Tissue
2.5. NFκB Reporter Gene Assay In Vitro
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Measurement of Hepatic Triglyceride Levels
4.3. Serum Parameters
4.4. Analysis of mRNA Expression in Perirenal WAT
4.5. In Vitro NFκB Reporter Gene Assay
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Diabetes Federation. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021; Available online: http://www.diabetesatlas.org/ (accessed on 17 December 2021).
- Fisher, M. Diabetes and atherogenesis. Heart 2004, 90, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Varughese, G.I.; Tomson, J.; Lip, G.Y. Type 2 diabetes mellitus: A cardiovascular perspective. Int. J. Clin. Pract. 2005, 59, 798–816. [Google Scholar] [CrossRef] [PubMed]
- Eckel, R.H.; Grundy, A.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2005, 365, 1415–1428. [Google Scholar] [CrossRef]
- Chen, H.; Charlat, O.; Tartaglia, L.A.; Woolf, E.A.; Weng, X.; Ellis, S.J.; Lakey, N.D.; Culpepper, J.; More, K.J.; Breitbart, R.E.; et al. Evidence that the diabetes gene encodes the leptin receptor: Identification of a mutation in the leptin receptor gene in db/db mice. Cell 1996, 84, 491–495. [Google Scholar] [CrossRef]
- Lee, G.H.; Proenca, R.; Montez, J.M.; Carroll, K.M.; Darvishzadah, J.G.; Lee, G.I.; Friedman, J.M. Abnormal splicing of the leptin receptor in diabetic mice. Nature 1996, 379, 632–635. [Google Scholar] [CrossRef]
- Makhmudova, U.; Schulze, P.C.; Lütjohann, D.; Weingärtner, O. Phytosterols and Cardiovascular Disease. Curr. Atheroscler. Rep. 2021, 23, 68. [Google Scholar] [CrossRef]
- Shirouchi, B.; Yanagi, S.; Okawa, C.; Koga, M.; Sato, M. 6-Ketocholestanol suppresses lipid accumulation by decreasing FASN gene expression through SREBP-dependent regulation in HepG2 cells. Cytotechnology 2020, 72, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.J.; Ficorilli, J.V.; Zhang, Y.; Bramlett, K.S.; Beyer, T.P.; Borchert, K.; Dowless, M.S.; Houck, K.A.; Burris, T.P.; Eacho, P.I.; et al. A 15-ketosterol is a liver X receptor ligand that suppresses sterol-responsive element binding protein-2 activity. J. Lipid Res. 2006, 47, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K. Anti-obesity effect of cholest-4-en-3-one, an intestinal catabolite of cholesterol, on mice. J. Nutr. Sci. Vitaminol. 1993, 39, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Shimizu, T.; Nakata, T. The cholesterol metabolite cholest-4-en-3-one and its 3-oxo derivatives suppress body weight gain, body fat accumulation and serum lipid concentration in mice. Bioorg. Med. Chem. Lett. 1998, 8, 2133–2138. [Google Scholar] [CrossRef]
- Ikeda, I.; Konno, R.; Shimizu, T.; Ide, T.; Takahashi, N.; Kawada, T.; Nagao, K.; Inoue, N.; Yanagita, T.; Hamada, T.; et al. Campest-5-en-3-one, an oxidized derivative of campesterol, activates PPARalpha, promotes energy consumption and reduces visceral fat deposition in rats. Biochim. Biophys. Acta 2006, 1760, 800–807. [Google Scholar] [CrossRef]
- Tamaru, S.; Suzuki, Y.; Sakono, M.; Fukuda, N.; Ikeda, I.; Konno, R.; Shimizu, T.; Suzuki, K. Dietary 5-campestenone (campest-5-en-3-one) enhances fatty acid oxidation in perfused rat liver. J. Nutr. Sci. Vitaminol. 2006, 52, 127–133. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Suzuki, K.; Konno, R.; Shimzu, T.; Nagashima, T.; Kimura, A. A fermentation product of phytosterol including campestenone reduces body fat storage and body weight gain in mice. J. Nutr. Sci. Vitaminol. 2007, 53, 63–67. [Google Scholar] [CrossRef]
- Boccellino, M.; D’Angelo, S. Anti-Obesity Effects of Polyphenol Intake: Current Status and Future Possibilities. Int. J. Mol. Sci. 2020, 21, 5642. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhang, W.; Li, X.; Xu, Y.; Cao, J.; Jiang, W. The anti-obesogenic effects of dietary berry fruits: A review. Food Res. Int. 2021, 147, 110539. [Google Scholar] [CrossRef] [PubMed]
- Leitner, D.R.; Frühbeck, G.; Yumuk, V.; Schindler, K.; Micic, D.; Woodward, E.; Toplak, H. Obesity and Type 2 Diabetes: Two Diseases with a Need for Combined Treatment Strategies—EASO Can Lead the Way. Obes. Facts 2017, 10, 483–492. [Google Scholar] [CrossRef]
- Suzuki, K.; Tanaka, M.; Konno, R.; Kaneko, Y. Effects of 5-campestenone (24-methylcholest-5-en-3-one) on the type 2 diabetes mellitus model animal C57BL/KsJ-db/db mice. Horm. Metab. Res. 2002, 34, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Konno, R.; Kaneko, Y.; Suzuki, K.; Matsui, Y. Effect of 5-Campestenone (24-methylcholest-5-en-3-one) on Zucker diabetic fatty rats as a type 2 diabetes mellitus model. Horm. Metab. Res. 2005, 37, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Cao, H. Adipocytokines in obesity and metabolic disease. J. Endocrinol. 2014, 220, T47–T59. [Google Scholar] [CrossRef]
- Chang, E.; Park, C.Y.; Park, S.W. Role of thiazolidinediones, insulin sensitizers, in non-alcoholic fatty liver disease. J. Diabetes Investig. 2013, 4, 517–524. [Google Scholar] [CrossRef]
- Lumeng, C.N.; Saltiel, A.R. Inflammatory links between obesity and metabolic disease. J. Clin. Investig. 2011, 121, 2111–2117. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Tateya, S.; Tamori, Y.; Kotani, K.; Hiasa, K.; Kitazawa, R.; Kitazawa, S.; Miyachi, H.; Maeda, S.; Egashira, K.; et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J. Clin. Investig. 2006, 116, 1494–1505. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Jenkins, J.R.; Trayhurn, P. Expression and secretion of inflammation-related adipokines by human adipocytes differentiated in culture: Integrated response to TNF-alpha. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E731–E740. [Google Scholar] [CrossRef]
- Juste, C.; Gérard, P. Cholesterol-to-Coprostanol Conversion by the Gut Microbiota: What We Know, Suspect, and Ignore. Microorganisms 2021, 9, 1881. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Karin, M. Missing pieces in the NF-κB puzzle. Cell 2002, 109, S81–S96. [Google Scholar] [CrossRef]
- Kim, J.K.; Kim, Y.J.; Fillmore, J.J.; Chen, Y.; Moore, I.; Lee, J.; Yuan, M.; Li, Z.W.; Karin, M.; Perret, P.; et al. Prevention of fat-induced insulin resistance by salicylate. J. Clin. Investig. 2001, 108, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Konrad, D.; Wueest, S. The gut-adipose-liver axis in the metabolic syndrome. Physiology 2014, 29, 304–313. [Google Scholar] [CrossRef]
- Morais, C.A.; de Rosso, V.V.; Estadella, D.; Pisani, L.P. Anthocyanins as inflammatory modulators and the role of the gut microbiota. J. Nutr. Biochem. 2016, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Kataria, M.A.; Saini, V.; Yadav, A. Role of leptin and adiponectin in insulin resistance. Clin. Chim. Acta 2013, 417, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Erten, M. Visfatin as a Promising Marker of Cardiometabolic Risk. Acta Cardiol. Sin. 2021, 37, 464–472. [Google Scholar]
- Teixeira, G.P.; Faria, R.X. Influence of purinergic signaling on glucose transporters: A possible mechanism against insulin resistance? Eur. J. Pharmacol. 2021, 892, 173743. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Tuo, Y.L.; Lu, Y.; Xu, C.R.; Xiang, M. The Role of CARD9 in Metabolic Diseases. Curr. Med. Sci. 2020, 40, 199–205. [Google Scholar] [CrossRef]
- Haneklaus, M.; O’Neill, L.A. NLRP3 at the interface of metabolism and inflammation. Immunol. Rev. 2015, 265, 53–62. [Google Scholar] [CrossRef]
- Tomita, T. Apoptosis in pancreatic β-islet cells in Type 2 diabetes. Bosn. J. Basic. Med. Sci. 2016, 16, 162–179. [Google Scholar] [CrossRef]
- American Institute of Nutrition. Report of the American Institute of Nutrition ad hoc committee on standards for nutritional studies. J. Nutr. 1977, 107, 1340–1348. [Google Scholar] [CrossRef]
- Parish, E.J.; Luo, C.; Parish, S.; Heidepriem, R.W. Selective oxidation of steroidal homoallylic alcohols using pyridium chlorochromate (PPC). Synth. Commun. 1992, 22, 2839–2847. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Fletcher, M.J. A colorimetric method for estimating serum triglycerides. Clin. Chim. Acta 1968, 22, 393–397. [Google Scholar] [CrossRef]
- Ishiyama, M.; Miyazono, Y.; Sasamoto, K.; Ohkura, Y.; Ueno, K. A highly water-soluble disulfonated tetrazolium salt as a chromogenic indicator for NADH as well as cell viability. Talanta 1997, 44, 1299–1305. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Basal Diet | STN Diet b |
|---|---|---|
| % | ||
| Casein | 20 | 20 |
| Cornstarch | 15 | 15 |
| Cellulose | 5 | 5 |
| Mineral mixture a | 3.5 | 3.5 |
| Vitamine mixture a | 1 | 1 |
| DL-Methionine | 0.3 | 0.3 |
| Choline bitartrate | 0.2 | 0.2 |
| Corn oil | 7 | 7 |
| Cholest-5-en-3-one (5-cholestenone) | - | 0.25 |
| Sucrose | 48 | 47.75 |
| NOR | CON | STN | |
|---|---|---|---|
| Initial body weight (g) | 22.2 ± 0.2 | 26.8 ± 0.2 a | 27.1 ± 0.2 b |
| Final body weight (g) | 23.4 ± 0.3 | 37.0 ± 0.5 a | 38.1 ± 0.4 b |
| Body weight gain (g) | 1.24 ± 0.21 | 10.1 ± 0.4 a | 11.0 ± 0.4 b |
| Food intake (g) | 74.7 ± 0.6 | 103 ± 1 a | 103 ± 2 b |
| Liver weight (g/100 g body weight) | 3.90 ± 0.03 | 7.45 ± 0.48 | 5.98 ± 0.56 b,c |
| White adipose tissue weight (g/100 g body weight) | |||
| Total | 2.63 ± 0.17 | 7.22 ± 0.27 a | 6.95 ± 0.31 b |
| Epididymal | 1.85 ± 0.09 | 4.49 ± 0.12 a | 4.32 ± 0.13 b |
| Perirenal | 0.781 ± 0.082 | 2.73 ± 0.18 a | 2.63 ± 0.27 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagao, K.; Inoue, N.; Suzuki, K.; Shimizu, T.; Yanagita, T. The Cholesterol Metabolite Cholest-5-en-3-One Alleviates Hyperglycemia and Hyperinsulinemia in Obese (db/db) Mice. Metabolites 2022, 12, 26. https://doi.org/10.3390/metabo12010026
Nagao K, Inoue N, Suzuki K, Shimizu T, Yanagita T. The Cholesterol Metabolite Cholest-5-en-3-One Alleviates Hyperglycemia and Hyperinsulinemia in Obese (db/db) Mice. Metabolites. 2022; 12(1):26. https://doi.org/10.3390/metabo12010026
Chicago/Turabian StyleNagao, Koji, Nao Inoue, Kunio Suzuki, Takeshi Shimizu, and Teruyoshi Yanagita. 2022. "The Cholesterol Metabolite Cholest-5-en-3-One Alleviates Hyperglycemia and Hyperinsulinemia in Obese (db/db) Mice" Metabolites 12, no. 1: 26. https://doi.org/10.3390/metabo12010026
APA StyleNagao, K., Inoue, N., Suzuki, K., Shimizu, T., & Yanagita, T. (2022). The Cholesterol Metabolite Cholest-5-en-3-One Alleviates Hyperglycemia and Hyperinsulinemia in Obese (db/db) Mice. Metabolites, 12(1), 26. https://doi.org/10.3390/metabo12010026