
Chemical Composition of the Red Sea Green Algae Ulva lactuca: Isolation and In Silico Studies of New Anti-COVID-19 Ceramides

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
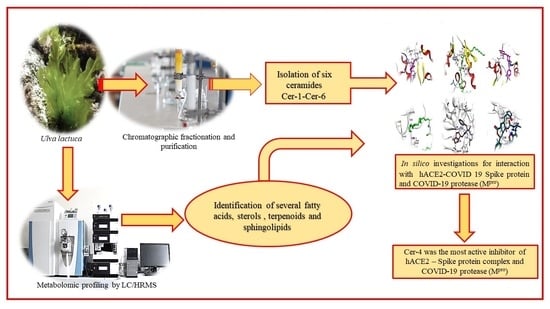
:
1. Introduction
2. Results and Discussion
2.1. Metabolic Profiling
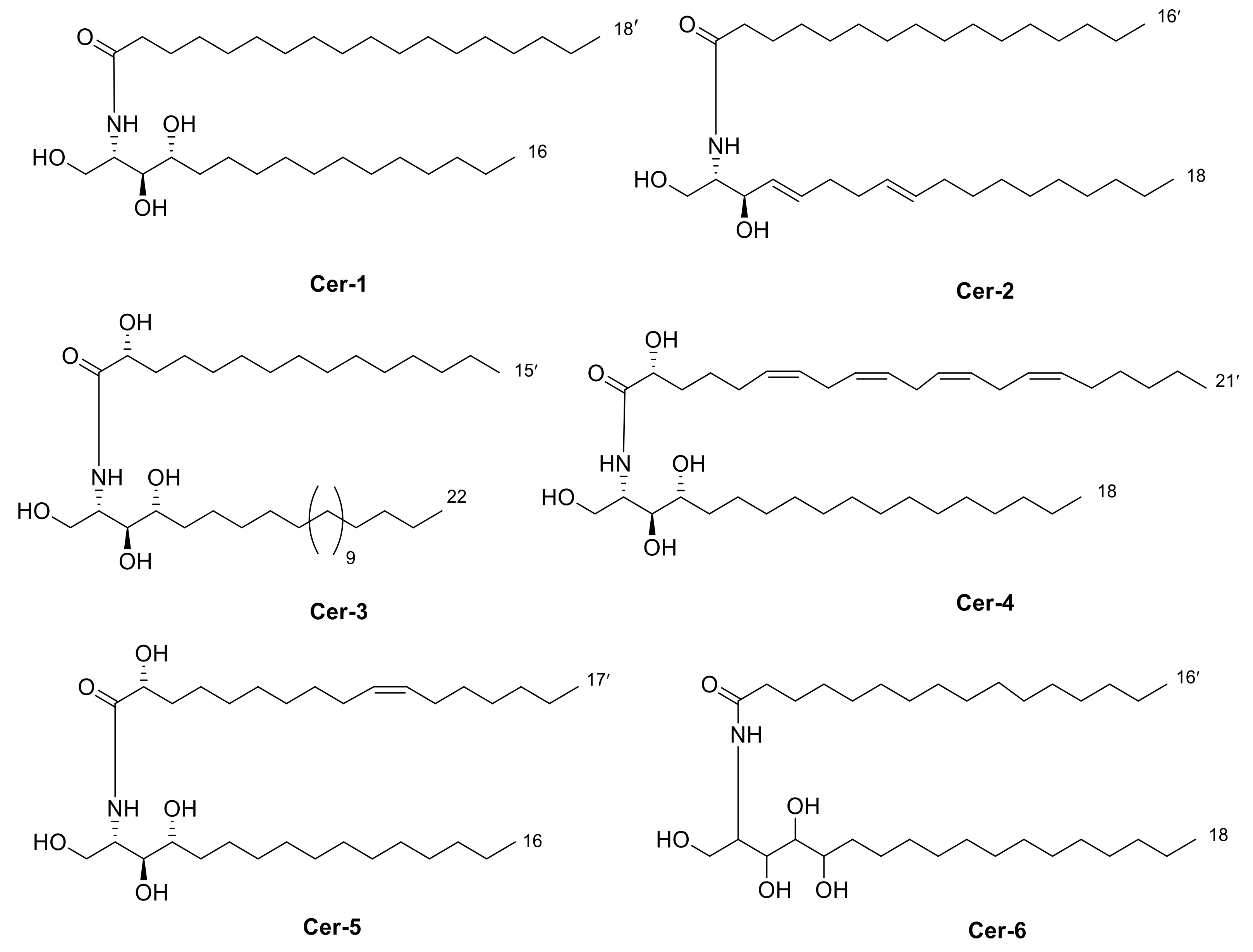
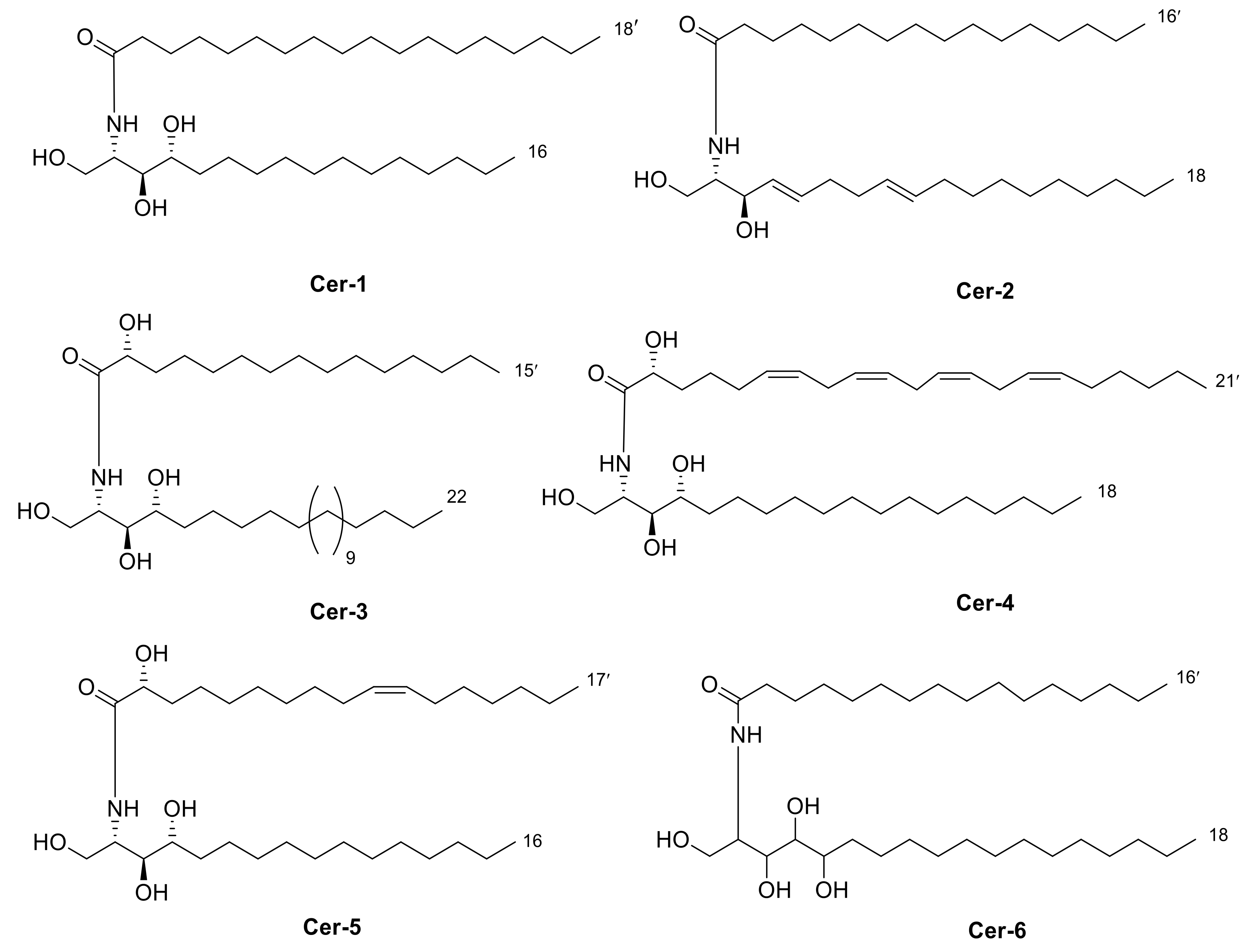
2.2. Isolation of Compounds 1–6
2.3. Identification of Isolated Compounds
2.4. Docking Studies
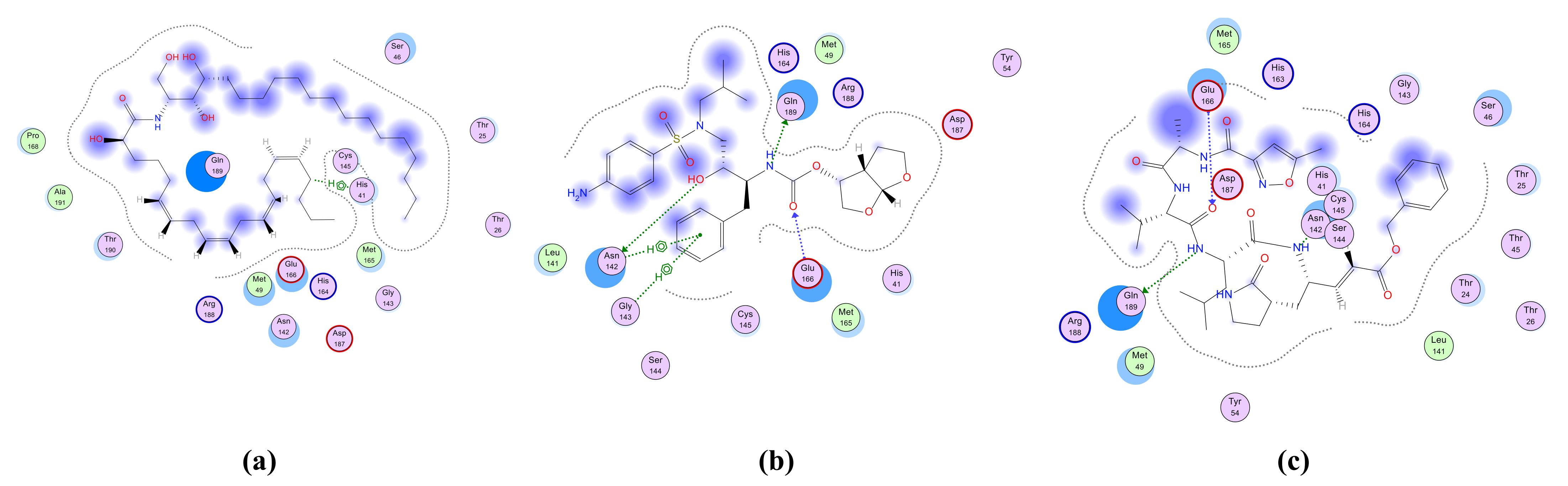
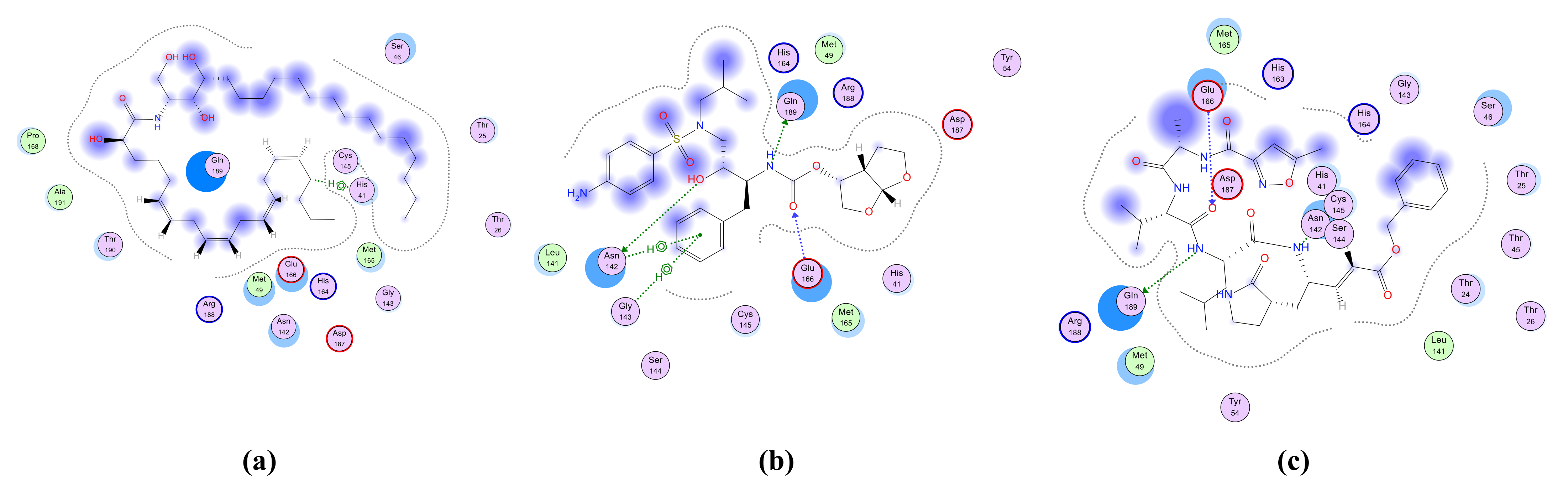
2.4.1. Molecular Docking Simulation of Isolated Ceramides and Detected Metabolites by LC-ESI-HRMS Technique to Site 4 of hACE2–SARS-CoV-2 Spike Protein Complex
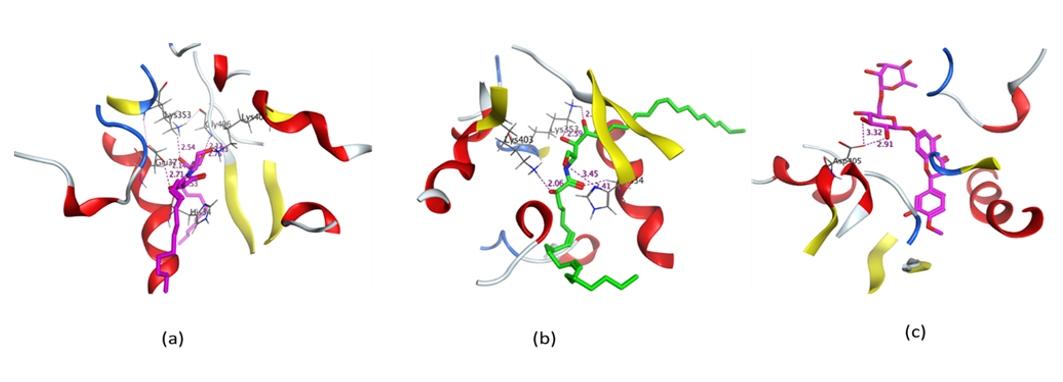
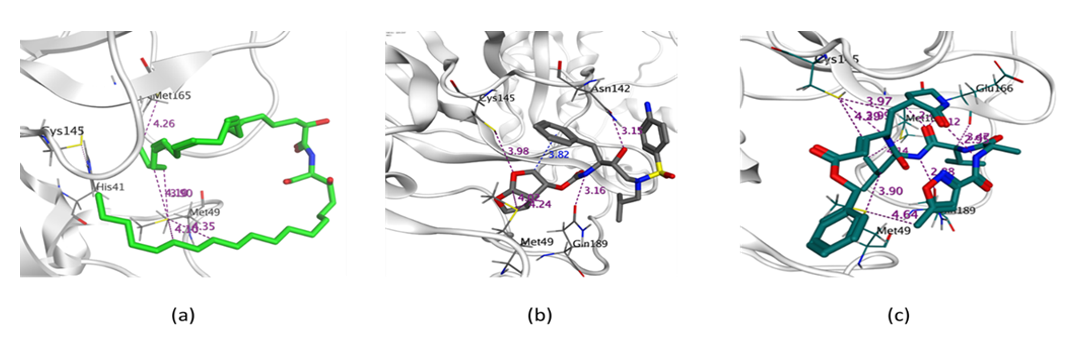
2.4.2. Molecular Docking Simulation of Isolated Ceramides and Detected Metabolites by LC-ESI-HRMS Technique to SARS-CoV-2 Mpro
2.4.3. ADME Studies
3. Materials and Methods
3.1. Plant Material
3.2. General Experimental Procedure
3.3. Metabolic Profiling
3.4. Extraction and Isolation
3.5. Docking Studies
3.5.1. Test Compounds Preparation
3.5.2. Optimization of the Enzymes’ Active Site
3.5.3. Docking of the Test Molecules to the Biggest Site of hACE2 Complex with SARS-CoV-2 Spike Protein and to SARS-CoV-2 Mpro
3.5.4. ADME Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bosch, B.J.; van der Zee, R.; de Haan, C.A.; Rottier, P.J. The coronavirus spike protein is a class I virus fusion protein: Structural and functional characterization of the fusion core complex. J. Virol. 2003, 77, 8801–88011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omrani, A.S.; Saad, M.M.; Baig, K.; Bahloul, A.; Abdul-Matin, M.; Alaidaroos, A.Y.; Almakhlafi, G.A.; Albarrak, M.M.; Memish, Z.A.; Albarrak, A.M. Ribavirin and interferon alfa-2a for severe Middle East respiratory syndrome coronavirus infection: A retrospective cohort study. Lancet Infect. Dis. 2014, 14, 1090–1095. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.Y.; Li, J.L.; Yang, X.L.; Chmura, A.A.; Zhu, G.; Epstein, J.H.; Mazet, J.K.; Hu, B.; Zhang, W.; Peng, C.; et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature 2013, 503, 535–538. [Google Scholar] [CrossRef]
- Han, D.P.; Penn-Nicholson, A.; Cho, M.W. Identification of critical determinants on ACE2 for SARS-CoVentry and development of a potent entry inhibitor. Virology 2006, 350, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Chen, P.; Wang, J.; Feng, J.; Zhou, H.; Li, X.; Zhong, W.; Hao, P. Evolution of the novel coronavirus from the ongoing Wuhan outbreak and modeling of its spike protein for risk of human transmission. Sci. China Life Sci. 2020, 63, 457–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, V.P.; Ignacio, B.L.; Martins, N.T.; Dobler, L.; Enrich-Prast, A. The Ulva spp. Conundrum: What Does the Ecophysiology of Southern Atlantic Specimens Tell Us? J. Mar. Biol. 2019, 2019, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.H.; Jang, J.E.; Kim, J.H.; Byeon, S.Y.; Kim, S.; Choi, S.K.; Kang, Y.H.; Park, S.R.; Lee, H.J. Species composition, diversity, and distribution of the genus Ulva along the coast of Jeju Island, Korea based on molecular phylogenetic analysis. PLoS ONE 2019, 14, e0219958. [Google Scholar] [CrossRef]
- Moustafa, Y.T.; Saeed, S.M. Nutritional evaluation of green macroalgae, Ulva sp. and related water nutrients in the Southern Mediterranean Sea coast, Alexandria shore, Egypt. Egypt. Acad. J. Biol. Sci. 2014, 5, 1–19. [Google Scholar] [CrossRef]
- Silva, M.; Vieira, L.; Almeida, A.P.; Kijjoa, A. The marine macroalgae of the genus Ulva: Chemistry, biological activities and potential applications. Oceanography 2013, 1, 1–6. [Google Scholar] [CrossRef]
- Kidgell, J.; Magnusson, M.; de Nys, R.; Glasson, C. Ulvan: A systematic review of extraction, composition and function. Algal Res. 2019, 39, 101422. [Google Scholar] [CrossRef]
- Abou El Azm, N.; Fleita, D.; Rifaat, D.; Mpingirika, E.Z.; Amleh, A.; El-Sayed, M.M.H. Production of Bioactive Compounds from the Sulfated Polysaccharides Extracts of Ulva lactuca: Post-Extraction Enzymatic Hydrolysis Followed by Ion-Exchange Chromatographic Fractionation. Molecules 2019, 24, 2132. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.; Jeong, S.; Lee, I.-K.; Yun, B.-S.; Lee, J.S.; Ro, S.; Park, J.K. Regulation of p53 Activity by (+)-Epiloliolide Isolated from Ulva lactuca. Mar. Drugs 2021, 19, 450. [Google Scholar] [CrossRef]
- Roleda, M.Y.; Lage, S.; Fonn Aluwini, D.; Rebours, C.; Bente Brurberg, M.; Nitschke, U.; Gentili, F.G. Chemical profiling of the Arctic Sea lettuce Ulva lactuca (Chlorophyta) mass-cultivated on land under controlled conditions for food applications. Food Chem. 2021, 341, 127999. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, K.; Dhara, S.; Mani, E.A. Ulvapyrone, a pyrone-linked benzochromene from sea lettuce Ulva lactuca Linnaeus (family Ulvaceae): Newly described anti-inflammatory agent attenuates arachidonate 5-lipoxygenase. Nat. Prod. Res. 2021, 20, 1–11. [Google Scholar] [CrossRef]
- Awad, N.E. Biologically active steroid from the green alga Ulva lactuca. Phytother. Res. 2000, 14, 641–643. [Google Scholar] [CrossRef]
- El Ashry, E.H.; Atta-ur-Rahman; Choudhary, M.I.; Kandil, S.; El Nemr, A.; Gulzar, T.; Shobier, A. Studies on the constituents of the green alga Ulva lactuca. Chem. Nat. Compd. 2011, 47, 335. [Google Scholar] [CrossRef]
- Sun, Y.; Zhan, Y.-C.; Sha, Y.; Pei, Y.-H. Norisoprenoids from Ulva lactuca. J. Asian Nat. Prod. Res. 2007, 9, 321–325. [Google Scholar] [CrossRef]
- Zaid, S.A.A.L.; Hamed, N.N.E.D.; Abdel-Wahab, K.S.E.D.; Abo El-Magd, E.K.; Salah El-Din, R.A.L. Antiviral activities and phytochemical constituents of Egyptian marine seaweeds (Cystoseiramyrica (SG Gmelin) C. Agardh and Ulva lactuca Linnaeus) aqueous extract. Egypt. J. Hosp. Med. 2016, 64, 422–429. [Google Scholar] [CrossRef]
- Mendes Gda, S.; Soares, A.R.; Martins, F.O.; Albuquerque, M.C.; Costa, S.S.; Yoneshigue-Valentin, Y.; Gestinari, L.M.; Santos, N.; Romanos, M.T. Antiviral activity of the green marine alga Ulva fasciata on the replication of human metapneumovirus. Rev. Inst. Med. Trop. Sao Paulo 2010, 52, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, A.R.; Robaina, M.; Mendes, G.S.; Silva, T.S.; Gestinari, L.; Pamplona, O.S.; Gestinari, L.M.; Yoneshigue-Valentin, Y.; Kaiser, C.; Romanos, M.; et al. Antiviral activity of extracts from Brazilian seaweeds against herpes simplex virus. Rev. Bras. Farmacogn. 2012, 22, 714–723. [Google Scholar] [CrossRef] [Green Version]
- Hudson, J.B.; Kim, J.H.; Lee, M.K.; Hong, Y.L. Antiviral compounds in extracts of Korean seaweeds: Evidence for multiple activities. J. Appl. Phycol. 1998, 10, 427. [Google Scholar] [CrossRef]
- Hudson, J.B.; Kim, J.H.; Lee, M.K.; Hong, Y.K.; DeWreede, R.E. Multiple antiviral activities in extracts of seaweeds from British Columbia. Pharm. Biol. 1999, 37, 300–306. [Google Scholar] [CrossRef]
- Ktari, L. Pharmacological Potential of Ulva Species: A Valuable Resource. J. Anal. Pharm. Res. 2017, 6, 00165. [Google Scholar]
- Deyab, M.; Mofeed, J. Evaluation of Biochemical composition and bioactivities of two Ulva species (Ulva lactuca and Ulva fasciata); a comparative study. Biosci. Res. 2019, 16, 3801–3811. [Google Scholar]
- Elyamany, K.; Ibraheem, I.; Abdel-Raouf, N.; Abdel-Hameed, M. Antimicrobial and antiviral activities against Newcastle disease virus (NDV) from marine algae isolated from Qusier and Marsa-Alam Seashore (Red Sea), Egypt. Afr. J. Biotechnol. 2012, 11, 8332–8340. [Google Scholar]
- Ivanova, V.; Rouseva, R.; Kolarova, M.; Serkedjieva, J.; Rachev, R.; Manolova, N. Isolation of a polysaccharide with antiviral effect from Ulva lactuca. Prep. Biochem. 1994, 24, 83–97. [Google Scholar] [CrossRef]
- Aguilar-Briseño, J.A.; Cruz-Suarez, L.E.; Sassi, J.-F.; Ricque-Marie, D.; Zapata-Benavides, P.; Mendoza-Gamboa, E.; Rodríguez-Padilla, C.; Trejo-Avila, L.M. Sulphated Polysaccharides from Ulva clathrate and Cladosiphon okamuranus Seaweeds both Inhibit Viral Attachment/Entry and Cell-Cell Fusion, in NDV Infection. Mar. Drugs 2015, 13, 697–712. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, X.; Song, L.; Liu, S.; Yu, H.; Wang, X.; Qin, Y.; Li, P. Antiviral activity against avian leucosis virus subgroup J of degraded polysaccharides from Ulva pertusa. BioMed. Res. Int. 2018, 5, 9415965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, K.H.; Liang, J.J.; Huang, W.I.; Lin-Chu, S.Y.; Su, C.Y.; Lee, Y.L.; Jan, J.T.; Lin, Y.L.; Cheng, Y.S.; Wong, C.H. In vivo protection provided by a synthetic new alpha-galactosyl ceramide analog against bacterial and viral infections in murine models. Antimicrob. Agents Chemother. 2010, 54, 4129. [Google Scholar] [CrossRef] [Green Version]
- Soudani, N.; Hage-Sleiman, R.; Karam, W.; Dbaibo, G.; Zaraket, H. Ceramide Suppresses Influenza A Virus Replication In Vitro. J. Virol. 2019, 93, e00053-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, H.; Ikeda, K.; Achiwa, K.; Hoshino, H. Synthesis of sulfated cerebroside analogs having mimicks of ceramide and their anti-human immunodeficiency virus type 1 activities. Chem. Pharm. Bull. 1995, 43, 594–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; Garg, H.S.; Chandra, K. Erythro-sphinga-4,8-dienine-N-palmitate: An Antiviral Agent from the Green Alga Ulva fasciata. Bot. Mar. 1996, 39, 1–6. [Google Scholar] [CrossRef]
- Abdelhameed, R.F.A.; Habib, E.S.; Goda, M.S.; Fahim, J.R.; Hassanean, H.A.; Eltamany, E.E.; Ibrahim, A.K.; AboulMagd, A.M.; Fayez, S.; El-kader, A.M.A.; et al. Thalassosterol, a New Cytotoxic Aromatase Inhibitor Ergosterol Derivative from the Red Sea Seagrass Thalassodendron ciliatum. Mar. Drugs 2020, 18, 354. [Google Scholar] [CrossRef]
- Thormar, H.; Isaacs, C.E.; Kim, K.S.; Brown, H.R. Inactivation of visna virus and other enveloped viruses by free fatty acids and monoglycerides. Ann. N. Y. Acad. Sci. 1994, 724, 465–471. [Google Scholar] [CrossRef]
- Abid Ali Khan, M.M.; Jain, D.C.; Bhakuni, R.S.; Zaim, M.; Thakur, R.S. Occurrence of some antiviral sterols in Artemisia annua. Plant Sci. 1991, 75, 161–165. [Google Scholar] [CrossRef]
- Ingallinella, P.; Bianchi, E.; Ladwa, N.A.; Wang, Y.J.; Hrin, R.; Veneziano, M.; Bonelli, F.; Ketas, T.J.; Moore, J.P.; Miller, M.D.; et al. Addition of a cholesterol group to an HIV-1 peptide fusion inhibitor dramatically increases its antiviral potency. Proc. Natl. Acad. Sci. USA 2009, 106, 5801–5806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lembo, D.; Cagno, V.; Civra, A.; Poli, G. Oxysterols: An emerging class of broad-spectrum antiviral effectors. Mol. Aspects Med. 2016, 49, 23–30. [Google Scholar] [CrossRef]
- Zhou, Y.; Pu, J.; Wu, Y. The Role of Lipid Metabolism in Influenza A Virus Infection. Pathogens 2021, 10, 303. [Google Scholar] [CrossRef]
- Ahmed, S.; Ibrahim, A.; Arafa, A.S. Anti-H5N1 virus metabolites from the Red Sea soft coral, Sinularia candidula. Tetrahedron Lett. 2013, 54, 2377–2381. [Google Scholar] [CrossRef]
- Alamsjah, M.A.; Hirao, S.; Ishibashi, F.; Fujita, Y. Isolation and structure determination of algicidal compounds from Ulva fasciata. Biosci. Biotechnol. Biochem. 2005, 69, 2186–2192. [Google Scholar] [CrossRef] [Green Version]
- van Ginneken, V.J.; Helsper, J.P.; de Visser, W.; van Keulen, H.; Brandenburg, W.A. Polyunsaturated fatty acids in various macroalgal species from north Atlantic and tropical seas. Lipids Health Dis. 2011, 10, 104. [Google Scholar] [CrossRef] [Green Version]
- Venkatesalu, V.; Sundaramoorthy, M.; Chandrasekaran, M.; Senthilkumar, A. Seasonal variation on fatty acid composition of some marine macro algae from Gulf of Mannar Marine Biosphere Reserve, Southeast coast of India. Indian J. Mar. Sci. 2012, 41, 442–450. [Google Scholar]
- Sun, Y.; Wang, H.; Guo, G.L.; Pu, Y.F.; Yan, B.L.; Wang, C.H. Isolation, purification, and identification of antialgal substances in green alga Ulva prolifera for antialgal activity against the common harmful red tide microalgae. Environ. Sci. Pollut. Res. Int. 2016, 23, 1449–1459. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Guo, W.; Wang, G.; Wang, R.; Hou, Y.; Liu, K.; Liu, Y.; Wang, W. Sterols from the green alga Ulva australis. Mar. Drugs 2017, 15, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Li, N.; Jiang, P.; Boo, S.M.; Lee, W.J.; Cui, Y.; Hanzhi, L.; Jin, Z.; Zhengyi, L.; Qin, S. Ulva and Enteromorpha (Ulvaceae, Chlorophyta) from two sides of the Yellow Sea: Analysis of nuclear rDNA ITS and plastid rbcL sequence data. Chin. J. Oceanol. Limnol. 2010, 28, 762–768. [Google Scholar] [CrossRef] [Green Version]
- Muralidhar, P.; Radhika, P.; Krishna, N.; Venkata-Rao, D.; Bheemasankara-Rao, C. Sphingolipids from Marine Organisms: A Review. Nat. Prod. Sci. 2003, 9, 117–142. [Google Scholar]
- Chakraborty, K.; Paulraj, R. Sesquiterpenoids with free-radical-scavenging properties from marine macroalga Ulva fasciata Delile. Food Chem. 2010, 122, 31–41. [Google Scholar] [CrossRef]
- Meyer, M.; Guyot, M. New Sphingosines from the Marine Sponge Grayella cyatophora. J. Nat. Prod. 2002, 65, 1722–1723. [Google Scholar] [CrossRef]
- Inagaki, M.; Ikeda, Y.; Kawatake, S.; Nakamura, K.; Tanaka, M.; Misawa, E.; Yamada, M.; Higuchi, R. Isolation and structure of four new ceramides from the Starfish Luidia maculate. Chem. Pharm. Bull. 2006, 54, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.E.; Heald, S.L.; Dally, R.D.; Janis, R.A. Isolation, identification and synthesis of an endogenous arachidonic amide that inhibits calcium channel antagonist 1,4-dihydropyridine binding. Prostaglandins, Leukot. Essent. Fatty Acids 1993, 48, 429–437. [Google Scholar] [CrossRef]
- Inagaki, M.; Isobe, R.; Kawano, Y.; Miyamoto, T.; Komori, T.; Higuchi, R. Isolation and structure of three new ceramides from the Starfish Acanthaster planci. Eur. J. Org. Chem. 1998, 1998, 129–131. [Google Scholar] [CrossRef]
- Qu, Y.; Zhangb, H.; Liua, J. Isolation and Structure of a New Ceramide from the Basidiomycete Hygrophorus eburnesus. Z. Nat. 2004, 59, 241–244. [Google Scholar]
- Garg, H.S.; Sharma, M.; Bhakuni, D.S.; Pramanik, B.N.; Bose, A.K. An antiviral sphingosine derivative from the green alga Ulva fasciata. Tetrahedron Lett. 1992, 33, 1641–1644. [Google Scholar] [CrossRef]
- Cavasotto, C.N.; Di Filippo, J.I. In silico Drug Repurposing for COVID-19: Targeting SARS-CoV-2 Proteins through Docking and Consensus Ranking. Mol. Inform. 2021, 40, e2000115. [Google Scholar] [CrossRef] [PubMed]
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 11 November 2021).
- Kulkarni, S.A.; Krishnan, S.B.B.; Chandrasekhar, B.; Banerjee, K.; Sohn, H.; Madhavan, T. Characterization of Phytochemicals in Ulva intestinalis L. and Their Action against SARS-CoV-2 Spike Glycoprotein Receptor-Binding Domain. Front. Chem. 2021, 9, 735768. [Google Scholar] [CrossRef]
- Romeo, I.; Mesiti, F.; Lupia, A.; Alcaro, S. Current Updates on Naturally Occurring Compounds Recognizing SARS-CoV-2 Druggable Targets. Molecules 2021, 26, 632. [Google Scholar] [CrossRef]
- Anand, A.V.; Balamuralikrishnan, B.; Kaviya, M.; Bharathi, K.; Parithathvi, A.; Arun, M.; Senthilkumar, N.; Velayuthaprabhu, S.; Saradhadevi, M.; Al-Dhabi, N.A.; et al. Medicinal Plants, Phytochemicals, and Herbs to Combat Viral Pathogens Including SARS-CoV-2. Molecules 2021, 26, 1775. [Google Scholar] [CrossRef]
- Ye, R.; Liu, Z. ACE2 exhibits protective effects against LPS-induced acute lung injury in mice by inhibiting the LPS-TLR4 pathway. Exp. Mol. Pathol. 2019, 113, 104350. [Google Scholar] [CrossRef]
- Wu, C.; Liu, Y.; Yang, Y.; Zhang, P.; Zhong, W.; Wang, Y.; Wang, Q.; Xu, Y.; Li, M.; Li, X.; et al. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharm. Sin. B 2020, 10, 766–788. [Google Scholar] [CrossRef] [PubMed]
- Bellavite, P.; Donzelli, A. Hesperidin and SARS-CoV-2: New Light on the Healthy Function of Citrus Fruits. Antioxidants 2020, 9, 742. [Google Scholar] [CrossRef]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V.; Hayashi, Y.; Jung, S.H. An Overview of Severe Acute Respiratory Syndrome–Coronavirus (SARS-CoV) 3CL Protease Inhibitors: Peptidomimetics and Small Molecule Chemotherapy. J. Med. Chem. 2016, 59, 6595–6628. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yang, M.; Ding, Y.; Liu, Y.; Lou, Z.; Zhou, Z.; Sun, L.; Mo, L.; Ye, S.; Pang, H.; et al. The crystal structures of severe acute respiratory syndrome virus main protease and its complex with an inhibitor. Proc. Natl. Acad. Sci. USA 2003, 100, 13190–13195. [Google Scholar] [CrossRef] [Green Version]
- Anand, K.; Palm, G.J.; Mesters, J.R.; Siddell, S.G.; Ziebuhr, J.; Hilgenfeld, R. Structure of coronavirus main proteinase reveals combination of a chymotrypsin fold with an extra alpha-helical domain. EMBO J. 2002, 21, 3213–3224. [Google Scholar] [CrossRef]
- Hegyi, A.; Ziebuhr, J. Conservation of substrate specificities among coronavirus main proteases. J. Gen. Virol. 2002, 83, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of Mpro from COVID-19 virus and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hay, M.; Thomas, D.W.; Craighead, J.L.; Economides, C.; Rosenthal, J. Clinical development success rates for investigational drugs. Nat. Biotechnol. 2014, 32, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, J.L.; Inglese, J.; Walters, M.A. Mitigating risk in academic preclinical drug discovery. Nat. Rev. Drug Discov. 2015, 14, 279–294. [Google Scholar] [CrossRef]
- Ertl, P.; Rohde, B.; Selzer, P. Fast Calculation of Molecular Polar Surface Area as a Sum of Fragment-Based Contributions and Its Application to the Prediction of Drug Transport Properties. J. Med. Chem. 2000, 43, 3714. [Google Scholar] [CrossRef]
- Daina, A.; Zoete, V. A boiled-egg to predict gastrointestinal absorption and brain penetration of small molecules. Chem. Med. Chem. 2016, 11, 1117–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, I.A.; Hollenberg, J. Marine Algae of California; Stanford University Press: Stanford, CA, USA, 1976; p. 827. [Google Scholar]
- Taylor, F.J.R. The Taxonomy and Relationships of Red Tide Flagellates. In Toxic Dinoflagellates; Anderson, D.M., White, A.W., Baden, D.G., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; pp. 11–26. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polarity Mode | Ret. Time (min) | Precursor m/z | MZmine ID | Detected Mass | Expected Mass | Mass Error (ppm) | Name | Structure | Reported Previously in | Ref. | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. Fatty acids | |||||||||||
| 1 | Positive | 7.14 | 249.1854 | 5976 | 248.1781 | 248.1776 | 2.01 | Hexadeca-4,7,10,13-tetraenoic acid | 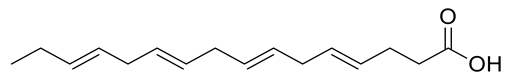 | Ulva fasciata | [41] |
| 2 | Negative | 10.13 | 295.2267 | 667 | 296.2340 | 296.2351 | −3.71 | 11E-Oxo-octadec-12-enoic acid | 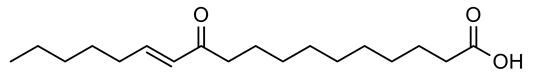 | Ulva fasciata | [41] |
| 3 | Negative | 10.52 | 275.2005 | 3655 | 276.2078 | 276.2089 | −3.98 | 6Z,9Z,12Z,15Z-Octadecatetraenoic acid |  | Ulva fasciata | [41] |
| 4 | Positive | 11.04 | 305.2479 | 565 | 304.2406 | 304.2402 | 1.31 | Arachidonic acid | 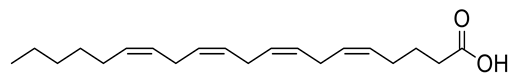 | Ulva lactuca | [42] |
| 5 | Negative | 11.15 | 277.2161 | 2459 | 278.2234 | 278.2246 | −4.31 | Linolenic acid | 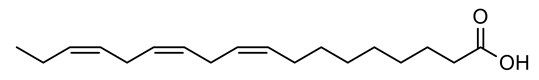 | Ulva fasciata | [41] |
| 6 | Negative | 11.84 | 283.2627 | 2827 | 284.2700 | 284.2715 | −5.28 | Stearic acid |  | Ulva fasciata | [43] |
| 2. Glycerol derivatives/glycerides | |||||||||||
| 7 | Positive | 8.97 | 331.2874 | 145 | 330.2774 | 330.2770 | 1.21 | Glycerol monopalmitate | 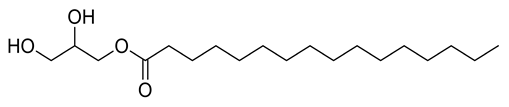 | Ulva prolifera | [44] |
| 8 | Positive | 9.40 | 521.3723 | 8312 | 520.3650 | 520.3611 | 7.49 | 1-O-Octadecanoic acid-3-O-β-D-galactopyranosyl glycerol | 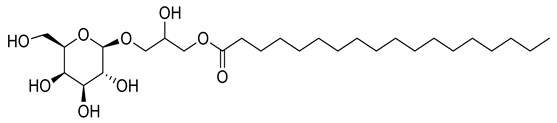 | Ulva prolifera | [44] |
| 9 | Negative | 10.02 | 491.3222 | 2431 | 492.3295 | 492.3298 | −0.61 | 1-O-Palmitoyl-3-O-β-D-galactopyranosyl glycerol |  | Ulva prolifera | [44] |
| 3. Sterols and oxysterols | |||||||||||
| 10 | Positive | 10.41 | 443.3524 | 5136 | 442.3452 | 442.3447 | 1.13 | 5,28-Stigmastadiene-3β,24-diol-7-one | 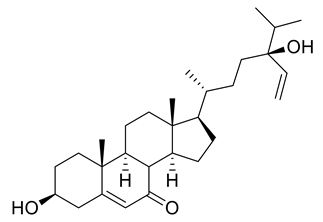 | Ulva australis | [45] |
| 11 | Negative | 13.85 | 395.3160 | 2931 | 396.3353 | 396.3392 | −9.84 | Ergosterol | 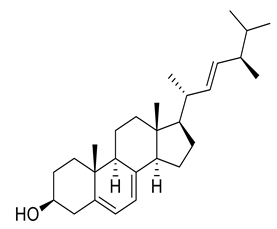 | Ulva pertusa | [46] |
| 12 | Positive | 15.25 | 575.4362 | 6572 | 574.4239 | 574.4233 | 1.04 | 3-O-β-D Glucopyranosyl-clerosterol | 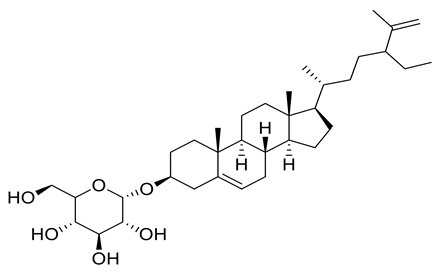 | Ulva lactuca | [16] |
| 13 | Positive | 16.52 | 429.3715 | 5560 | 428.3643 | 428.3654 | −2.57 | 24,28-Epoxy-24-ethylcholesterol |  | Ulva australis | [45] |
| 14 | Positive | 16.52 | 429.3715 | 5560 | 428.3643 | 428.3654 | −2.57 | 5,28-Stigmastadiene-3β,24-diol | 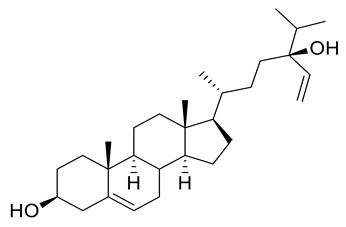 | Ulva australis | [45] |
| 4. Ceramide and sphingoid base | |||||||||||
| 15 | Positive | 12.34 | 328.3198 | 6410 | 327.3125 | 327.3137 | −3.67 | N,N-Dimethyl sphingosine |  | fungi | [42] |
| 16 | Positive | 14.35 | 558.5128 | 7471 | 557.5055 | 557.5019 | 6.46 | N-[pentadecanoate ]-1,3,4,5-tetrahydroxy-2-amino-octadecane | 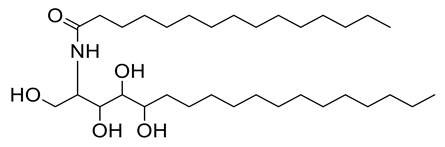 | Ulva fasciata | [47] |
| 5. Terpenoid | |||||||||||
| 17 | Positive | 9.06 | 223.2061 | 5846 | 222.1988 | 222.1984 | 1.80 | 2,5,5-Trimethyl-4-(4′-methyl-3′-pentenyl)-2-cyclohexen-1-ol |  | Ulva fasciata | [48] |
| Cer-1 * | Cer-2 * | Cer-3 * | Cer-4 # | Cer-5 * | Cer-6 * | |
|---|---|---|---|---|---|---|
| 1 | 4.95 (m) | 4.53 (m) | 4.42 (m) | 4.06 (m) | 4.60 (m) | 4.54 (m) |
| 2 | 5.07 (m) | 5.04 (m) | 5.06 (m) | 5.08 (m) | 5.12 (m) | 5.11 (m) |
| 3 | 4.38 (m) | 4.94 (m) | 4.39 (m) | 3.87 (m) | 4.27 (m) | 4.44 (m) |
| 4 | 4.27 (m) | 5.46 (m) | 4.31 (m) | 3.75 (m) | 4.44 (m) | 4.41 (m) |
| 5 | 2.22 (m) | 5.46 (m) | 2.18 (m) | 1.99 (m) | 2.10 (m) | 4.43 (m) |
| 6 | 1.97 (m) | 2.19 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 2.02 (m) |
| 7 | 1.21–1.26 (m) | 1.94 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 8 | 1.21–1.26 (m) | 5.46 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 9 | 1.21–1.26 (m) | 5.46 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 10 | 1.21–1.26 (m) | 1.90 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 11 | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 12 | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 13 | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 14 | 1.67 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 15 | 1.35 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.36 (m) | 1.25–1.30 (m) |
| 16 | 0.84 (t, J = 8) | 1.23–1.29 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 0.85 (t, J = 8) | 1.25–1.30 (m) |
| 17 | - | 1.68 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | - | 1.71 (m) |
| 18 | - | 0.84 (t, J = 8) | 1.10–1.26 (m) | 0.84 (t, J = 8) | - | 0.85 (t, J = 8) |
| 19 | - | - | 1.10–1.26 (m) | - | - | - |
| 20 | - | - | 1.10–1.26 (m) | - | - | - |
| 21 | - | - | 1.70 (m) | - | - | - |
| 22 | - | - | 0.81 (t, J = 4) | - | - | - |
| 1′ | -- | - | - | - | - | - |
| 2′ | 2.45 (t, J = 8) | 4.19 (brt, J = 4) | 4.60 (t, J = 8) | 4.23 (t, J = 8) | 4.97 (t, J = 8) | 2.23 (brt, J = 4) |
| 3′ | 1.84 (m) | 1.98 (m) | 1.95 (m) | 1.95 (m) | 2.10 (m) | 1.71 (m) |
| 4′ | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 5′ | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 6′ | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 5.77 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 7′ | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 5.77 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 8′ | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 9′ | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 5.53 (m) | 2.26 (m) | 1.25–1.30 (m) |
| 10′ | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 5.53 (m) | 5.5 (m) | 1.25–1.30 (m) |
| 11′ | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 1.28–1.36 (m) | 5.5 (m) | 1.25–1.30 (m) |
| 12′ | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 5.45 (m) | 2.26 (m) | 1.25–1.30 (m) |
| 13′ | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.10–1.26 (m) | 5.45 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 14′ | 1.21–1.26 (m) | 1.23–1.29 (m) | 1.70 (m) | 1.28–1.36 (m) | 1.22–1.28 (m) | 1.25–1.30 (m) |
| 15′ | 1.21–1.26 (m) | 1.68 (m) | 0.81 (t, J = 4) | 5.34 (m) | 1.22–1.28 (m) | 1.71 (m) |
| 16′ | 1.67 (m) | 0.84 (t, J = 8) | - | 5.34 (m) | 1.36 (m) | 0.85 (t, J = 8) |
| 17′ | 1.35 (m) | - | - | 1.28–1.36 (m) | 0.85 (t, J = 8) | - |
| 18′ | 0.84 (t, J = 8) | - | -- | 1.28–1.36 (m) | - | - |
| 19′ | - | - | - | 1.28–1.36 (m) | - | - |
| 20′ | - | - | - | 1.74 (m) | - | - |
| 21′ | - | - | - | 0.84 (t, J = 8) | - | - |
| NH | 8.50 (d, J = 8) | 8.57 (d, J = 12) | 8.52 (d, J = 12) | 7.25 (d, J = 12) | 8.58 (d, J = 8) | 8.55 (d, J = 12) |
| Cer-1 * | Cer-2 * | Cer-3 * | Cer-4 # | Cer-5 * | Cer-6 * | |
|---|---|---|---|---|---|---|
| 1 | 62.2 | 62.6 | 61.7 | 61.9 | 61.7 | 61.9 |
| 2 | 53.8 | 54.5 | 52.6 | 54.4 | 52.6 | 52.9 |
| 3 | 76.7 | 75.1 (72) | 76.5 | 77.2 | 76.4 | 72.4 |
| 4 | 73.1 | 131.1 | 72.7 | 74.0 | 72.7 | 73.0 |
| 5 | 33.9 | 132.0 | 33.8 | 32.5 | 33.8 | 76.6 |
| 6 | 29.6–29.9 | 32.9 | 29.2–30.0 | 27.3 | 29.3–29.6 | 34.0 |
| 7 | 29.6–29.9 | 28.0 | 29.2–30.0 | 29.3–29.7 | 29.3–29.6 | 29.5–30.2 |
| 8 | 29.6–29.9 | 131.9 | 29.2–30.0 | 29.3–29.7 | 29.3–29.6 | 29.5–30.2 |
| 9 | 29.6–29.9 | 129.9 | 29.2–30.0 | 29.3–29.7 | 29.3–29.6 | 29.5–30.2 |
| 10 | 29.6–29.9 | 39.9 | 29.2–30.0 | 29.3–29.7 | 29.3–29.6 | 29.5–30.2 |
| 11 | 29.6–29.9 | 29.5–29.9 | 29.2–30.0 | 29.3–29.7 | 29.3–29.6 | 29.5–30.2 |
| 12 | 29.6–29.9 | 29.5–29.9 | 29.2–30.0 | 29.3–29.7 | 29.3–29.6 | 29.5–30.2 |
| 13 | 29.6–29.9 | 29.5–29.9 | 29.2–30.0 | 29.3–29.7 | 29.3–29.6 | 29.5–30.2 |
| 14 | 29.6–29.9 | 29.5–29.9 | 29.2–30.0 | 29.3–29.7 | 29.3–29.6 | 29.5–30.2 |
| 15 | 22.9 | 29.5–29.9 | 29.2–30.0 | 29.3–29.7 | 31.8 | 29.5–30.2 |
| 16 | 14.2 | 31.9 | 29.2–30.0 | 31.6 | 13.9 | 32.0 |
| 17 | - | 22.8 | 29.2–30.0 | 22.6 | - | 22.6 |
| 18 | - | 14.1 | 29.2–30.0 | 14.0 | - | 14.1 |
| 19 | - | - | 29.2–30.0 | - | - | - |
| 20 | - | - | 29.2–30.0 | - | - | - |
| 21 | - | - | 31.8 | - | - | - |
| 22 | - | - | 13.9 | - | - | - |
| 1′ | 173.5 | 175.7 | 175.1 | 175.7 | 175.1 | 175.3 |
| 2′ | 36.9 | 35.6 | 72.1 | 72.5 | 72.1 | 35.6 |
| 3′ | 29.6–29.9 | 25.7 | 35.3 | 34.5 | 35.3 | 26.5 |
| 4′ | 29.6–29.9 | 29.4–29.9 | 29.2–30.0 | 29.3–29.7 | 27.0 | 29.5–30.2 |
| 5′ | 29.6–29.9 | 29.4–29.9 | 29.2–30.0 | 29.3–29.7 | 29.3–29.6 | 29.5–30.2 |
| 6′ | 29.6–29.9 | 29.4–29.9 | 29.2–30.0 | 134.1 | 29.3–29.6 | 29.5–30.2 |
| 7′ | 29.6–29.9 | 29.4–29.9 | 29.2–30.0 | 134.1 | 29.3–29.6 | 29.5–30.2 |
| 8′ | 29.6–29.9 | 29.4–29.9 | 29.2–30.0 | 29.3–29.7 | 22.5 | 29.5–30.2 |
| 9′ | 29.6–29.9 | 29.4–29.9 | 29.2–30.0 | 132.5 | 26.0 | 29.5–30.2 |
| 10′ | 29.6–29.9 | 29.4–29.9 | 29.2–30.0 | 132.5 | 132.7 | 29.5–30.2 |
| 11′ | 29.6–29.9 | 29.4–29.9 | 29.2–30.0 | 29.3–29.7 | 132.7 | 29.5–30.2 |
| 12′ | 29.6–29.9 | 29.4–29.9 | 29.2–30.0 | 131.3 | 26.0 | 29.5–30.2 |
| 13′ | 29.6–29.9 | 29.4–29.9 | 29.2–30.0 | 131.3 | 29.3–29.6 | 29.5–30.2 |
| 14′ | 29.6–29.9 | 31.9 | 31.8 | 29.3–29.7 | 29.3–29.6 | 32.0 |
| 15′ | 29.6–29.9 | 22.8 | 13.9 | 129.0 | 29.3–29.6 | 22.6 |
| 16′ | 29.6–29.9 | 14.1 | - | 126.7 | 31.8 | 14.1 |
| 17′ | 22.9 | - | - | 29.3–29.7 | 13.9 | - |
| 18′ | 14.2 | - | -- | 29.3–29.7 | - | - |
| 19′ | - | - | - | 31.6 | - | - |
| 20′ | - | - | - | 22.6 | - | - |
| 21′ | - | - | - | 14.0 | - | - |
| Compound | dG Kcal/mole | Receptor | |
|---|---|---|---|
| Amino Acid/Type of Bonding/Distance (Å)/Binding Energy (Kcal/mole) | |||
| hACE2 | Spike Protein | ||
| Cer-1 | −8.2488 | GLU 37/H-donor/3.03/−3.2 ARG 393/H-acceptor/3.37/−0.6 | - |
| Cer-2 | −8.0390 | GLU 37/H-donor/2.85/−7.0 | - |
| Cer-3 | −8.3111 | GLU 37/H-donor/2.88/−2.4 | - |
| Cer-4 | −9.6464 | HIS 34/H-donor/3.45/−1.0 HIS 34/H-acceptor/3.38/−0.7 | LYS 403/H-acceptor/2.06/−0.5 |
| Cer-5 | −8.7077 | GLU 37/H-donor/2.71/−3.1 HIS 34/H-pi/3.64/−1.4 | GLY 496/H-acceptor/3.04/−1.7 LYS 403/H-acceptor/2.98/−4.9 |
| Cer-6 | −8.6033 | GLU 37/H-donor/2.85/−2.8 GLU 37/H-donor/2.74/−2.8 ARG 393/H-acceptor/2.88/−4.1 HIS 34/H-pi/3.76/−1.4 | - |
| 1 | −6.0580 | - | - |
| 2 | −5.9953 | LYS 353/H-acceptor/3.23/−1.8 | - |
| 3 | −5.9901 | GLU 37/H-donor/2.94/−6.4 | - |
| 4 | −5.5072 | - | - |
| 5 | −6.0686 | HIS 34/H-donor/3.50/−0.6 | - |
| 6 | −6.2414 | - | - |
| 7 | −6.5090 | - | ASP 406/H-donor/2.90/−3.7 LYS 403/H-acceptor/3.05/−2.3 |
| 8 | −8.4482 | GLU 37/H-donor/2.80/−3.6 HIS 34/H-pi/3.91/−1.3 | LYS 403/H-acceptor/3.05/−7.5 ARG 408/H-acceptor/2.99/−2.8 LYS 403/H-acceptor/3.10/−4.7 |
| 9 | −7.7408 | GLU 37/H-donor/2.90/−2.2 | - |
| 10 | −5.8042 | HIS 34/H-pi/3.72/−1.2 | - |
| 11 | −4.9095 | - | - |
| 12 | −6.5362 | - | SER 494/H-donor/2.89/−2.5 LYS 403/H-acceptor/2.93/−3.7 GLY 496/H-acceptor/3.38/−0.9 |
| 13 | −5.8862 | HIS 34/H-pi/3.7 3/−1.2 | LYS 403/H-acceptor/2.92/−7.0 |
| 14 | −5.4205 | HIS 34/H-pi/3.76/−0.7 | - |
| 15 | −6.1762 | - | - |
| 16 | −8.6033 | GLU 37/H-donor/2.85/−2.8 GLU 37/H-donor/2.74/−2.8 ARG 393/H-acceptor/2.88/−4.1 HIS 34/H-pi/3.76/−1.4 | - |
| 17 | −4.1495 | - | LYS 403/H-acceptor/2.80/−4.2 |
| Hesperidin | −6.0117 | - | ASP 405/H-donor/2.91/−1.8 |
| Compound | dG Kcal/mole | Receptor |
|---|---|---|
| Amino Acid/Type of Bonding/Distance (Å)/Binding Energy (Kcal/mole) | ||
| Cer-1 | −7.4765 | HIS 41/H-pi/4.08/−0.7 |
| Cer-2 | −7.1107 | - |
| Cer-3 | −7.5529 | THR 190/H-donor/2.77/−1.7 GLU 166/H-donor/2.85/−2.4 MET 165/H-donor/3.57/−1.6 |
| Cer-4 | −7.9390 | HIS 41/H-pi/4.38/−0.6 |
| Cer-5 | −7.4910 | - |
| Cer-6 | −6.7077 | GLY 143/H-acceptor/3.09/−0.7 HIS 41/H-pi/4.74/−0.6 |
| 1 | −5.9111 | HIS 163/H-acceptor/2.96/−7.3 |
| 2 | −6.2654 | HIS 41/H-pi/4.07/−1.1 |
| 3 | −6.0563 | GLU 166/H-donor/3.05/−2.9 HIS 163/H-acceptor/3.06/−5.1 |
| 4 | −6.3289 | - |
| 5 | −6.3581 | - |
| 6 | −5.8555 | - |
| 7 | −6.0696 | THR 190/H-donor/2.96/−1.2 GLU 166/H-acceptor/3.10/−1.5 HIS 41/H-pi/3.74/−0.9 |
| 8 | −6.7570 | GLN 189/H-donor/3.31/−0.8 GLU 166/H-donor/3.00/−0.7 ASN 142/H-donor/3.37/−0.8 GLU 166/H-acceptor/2.70/−2.3 HIS 163/H-acceptor/3.02/−2.6 |
| 9 | −7.0953 | HIS 164/H-donor/3.19/−0.7 CYS 145/H-donor/3.81/−1.1 MET 165/H-donor/3.80/−0.8 MET 165/H-donor/3.57/−1.2 HIS 163/H-acceptor/3.31/−2.9 |
| 10 | −5.1913 | GLY 143/H-acceptor/2.94/−2.2 |
| 11 | −5.9792 | - |
| 12 | −6.9449 | THR 190/H-donor/2.95/−1.7 |
| 13 | −5.5319 | - |
| 14 | −5.1384 | GLY 143/H-acceptor/2.97/−0.4 |
| 15 | −6.7512 | PHE 140/H-donor/3.07/−0.8 CYS 145/H-donor/3.78/−1.2 HIS 163/H-acceptor/3.11/−0.7 |
| 16 | −6.7077 | GLY 143/H-acceptor/3.09/−0.7 HIS 41/H-pi/4.74/−0.6 |
| 17 | −5.4313 | - |
| Darunavir | −6.4501 | GLN 189/H-donor/3.16/−2.2 ASN 142/H-donor/3.15/−0.9 GLU 166/H-acceptor/3.01/−4.0 ASN 142/pi-H/4.01/−1.6 GLY 143/pi-H/4.18/−0.6 |
| N3 | −8.4847 | GLN 189/H-donor/2.88/−3.4 CYS 145/H-donor/3.99/−1.5 GLU 166/H-acceptor/3.08/−2.6 |
| Compound | TPSA Å2 | Log Po/w (MLOGP) | GI Absorption | BBB Permeant | PAINS Alert |
|---|---|---|---|---|---|
| Cer-1 | 89.79 | 4.98 | Low | No | 0 |
| Cer-2 | 69.56 | 5.61 | Low | No | 0 |
| Cer-3 | 110.02 | 4.71 | Low | No | 0 |
| Cer-4 | 110.02 | 4.67 | Low | No | 0 |
| Cer-5 | 110.02 | 3.88 | Low | No | 0 |
| Cer-6 | 110.02 | 3.99 | Low | No | 0 |
| 1 | 37.30 | 3.81 | High | Yes | 0 |
| 2 | 54.37 | 3.59 | High | Yes | 0 |
| 3 | 37.30 | 4.29 | High | Yes | 0 |
| 4 | 37.30 | 4.75 | High | No | 0 |
| 5 | 37.30 | 4.38 | High | Yes | 0 |
| 6 | 37.30 | 4.67 | High | No | 0 |
| 7 | 66.76 | 3.18 | High | Yes | 0 |
| 8 | 145.91 | 0.89 | Low | No | 0 |
| 9 | 145.91 | 0.48 | Low | No | 0 |
| 10 | 57.53 | 4.72 | High | Yes | 0 |
| 11 | 20.23 | 6.33 | Low | No | 0 |
| 12 | 99.38 | 3.85 | High | No | 0 |
| 13 | 32.76 | 5.80 | Low | No | 0 |
| 14 | 40.46 | 5.7 | Low | No | 0 |
| 15 | 43.70 | 3.44 | High | Yes | 0 |
| 16 | 110.02 | 3.99 | Low | No | 0 |
| 17 | 20.23 | 3.56 | Low | No | 0 |
| Hesperidin | 234.29 | −3.04 | Low | No | 0 |
| Darunavir | 148.80 | 1.18 | Low | No | 0 |
| N3 | 197.83 | 0.38 | Low | No | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eltamany, E.E.; Elhady, S.S.; Goda, M.S.; Aly, O.M.; Habib, E.S.; Ibrahim, A.K.; Hassanean, H.A.; Abdelmohsen, U.R.; Safo, M.K.; Ahmed, S.A. Chemical Composition of the Red Sea Green Algae Ulva lactuca: Isolation and In Silico Studies of New Anti-COVID-19 Ceramides. Metabolites 2021, 11, 816. https://doi.org/10.3390/metabo11120816
Eltamany EE, Elhady SS, Goda MS, Aly OM, Habib ES, Ibrahim AK, Hassanean HA, Abdelmohsen UR, Safo MK, Ahmed SA. Chemical Composition of the Red Sea Green Algae Ulva lactuca: Isolation and In Silico Studies of New Anti-COVID-19 Ceramides. Metabolites. 2021; 11(12):816. https://doi.org/10.3390/metabo11120816
Chicago/Turabian StyleEltamany, Enas E., Sameh S. Elhady, Marwa S. Goda, Omar M. Aly, Eman S. Habib, Amany K. Ibrahim, Hashim A. Hassanean, Usama Ramadan Abdelmohsen, Martin K. Safo, and Safwat A. Ahmed. 2021. "Chemical Composition of the Red Sea Green Algae Ulva lactuca: Isolation and In Silico Studies of New Anti-COVID-19 Ceramides" Metabolites 11, no. 12: 816. https://doi.org/10.3390/metabo11120816
APA StyleEltamany, E. E., Elhady, S. S., Goda, M. S., Aly, O. M., Habib, E. S., Ibrahim, A. K., Hassanean, H. A., Abdelmohsen, U. R., Safo, M. K., & Ahmed, S. A. (2021). Chemical Composition of the Red Sea Green Algae Ulva lactuca: Isolation and In Silico Studies of New Anti-COVID-19 Ceramides. Metabolites, 11(12), 816. https://doi.org/10.3390/metabo11120816