
Effects of Platycodon grandiflorum on Gut Microbiome and Immune System of Immunosuppressed Mouse

 , , , ,
, , , ,  ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
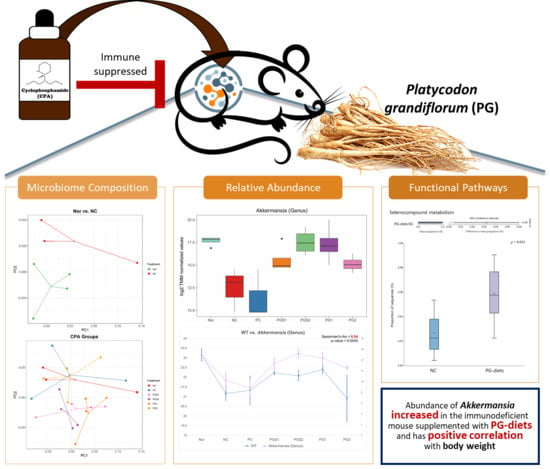
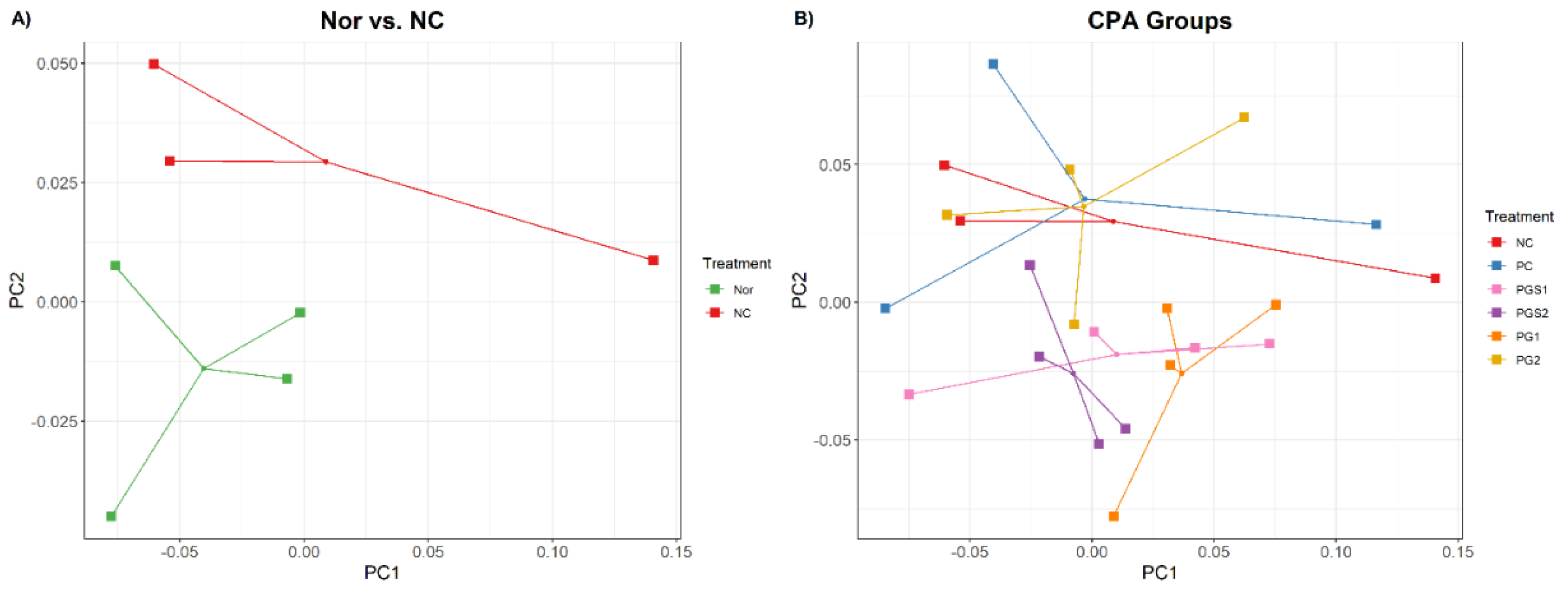
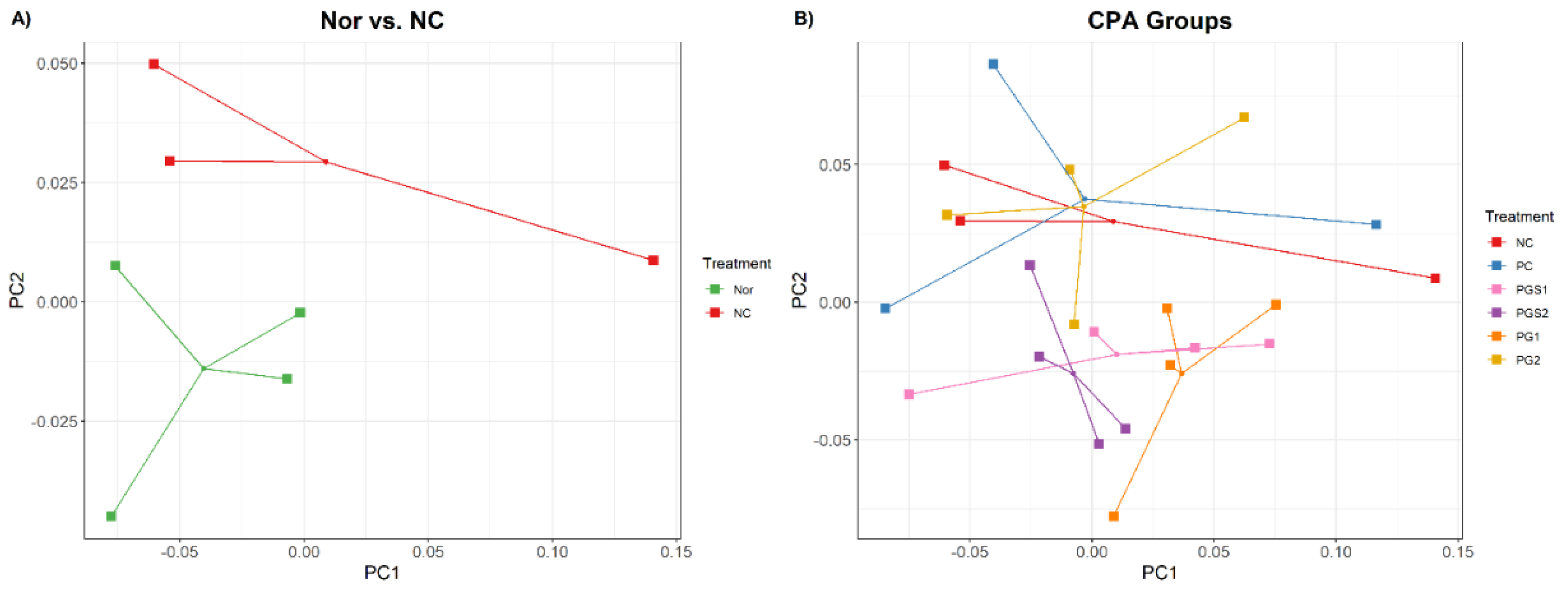
2.1. Effects of Platycodon grandiflorum on Host Immune System and Gut Microbiome Diversity
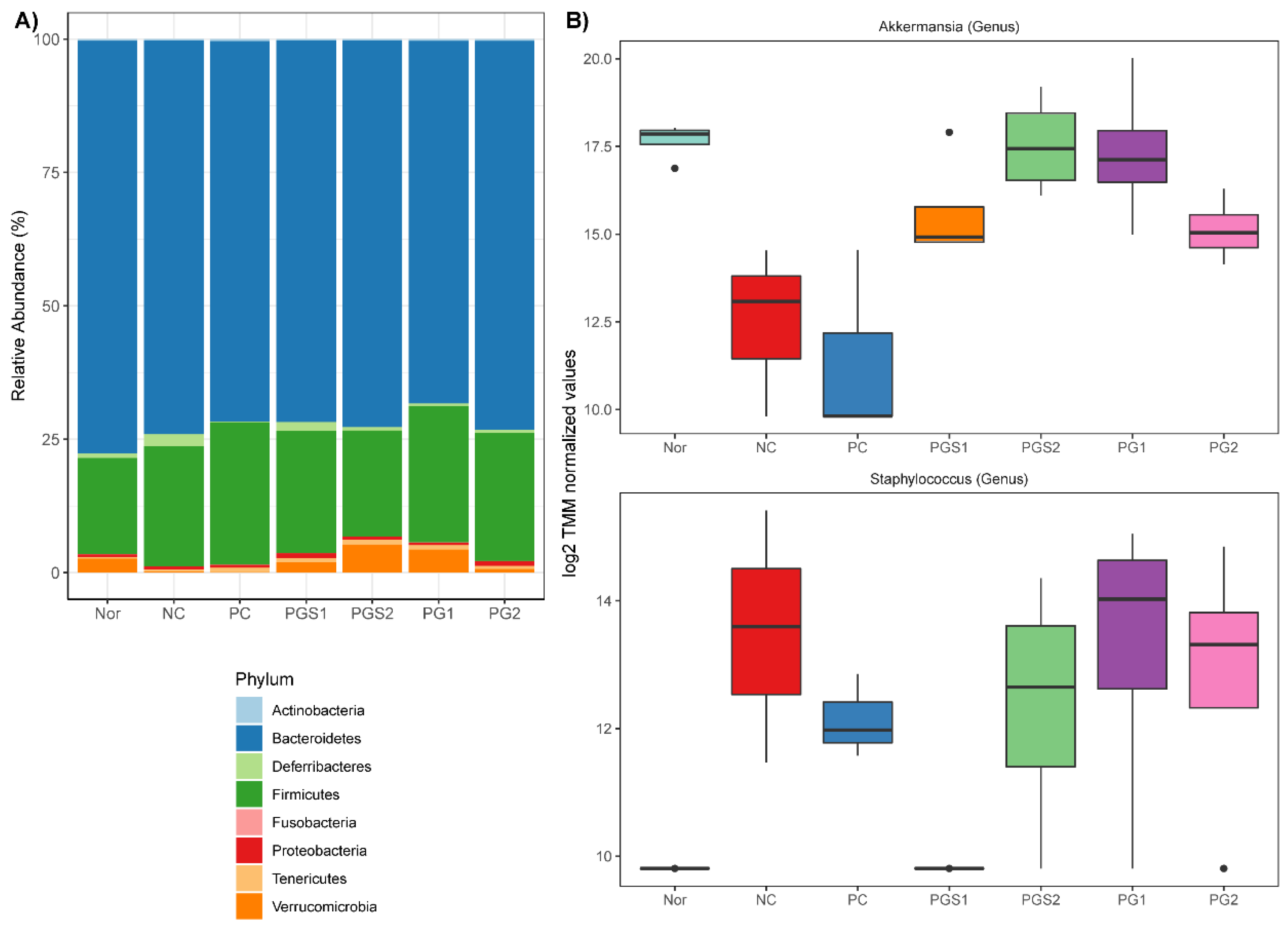
2.2. Change in Microorganism Abundance by Supplement of Platycodon grandiflorum
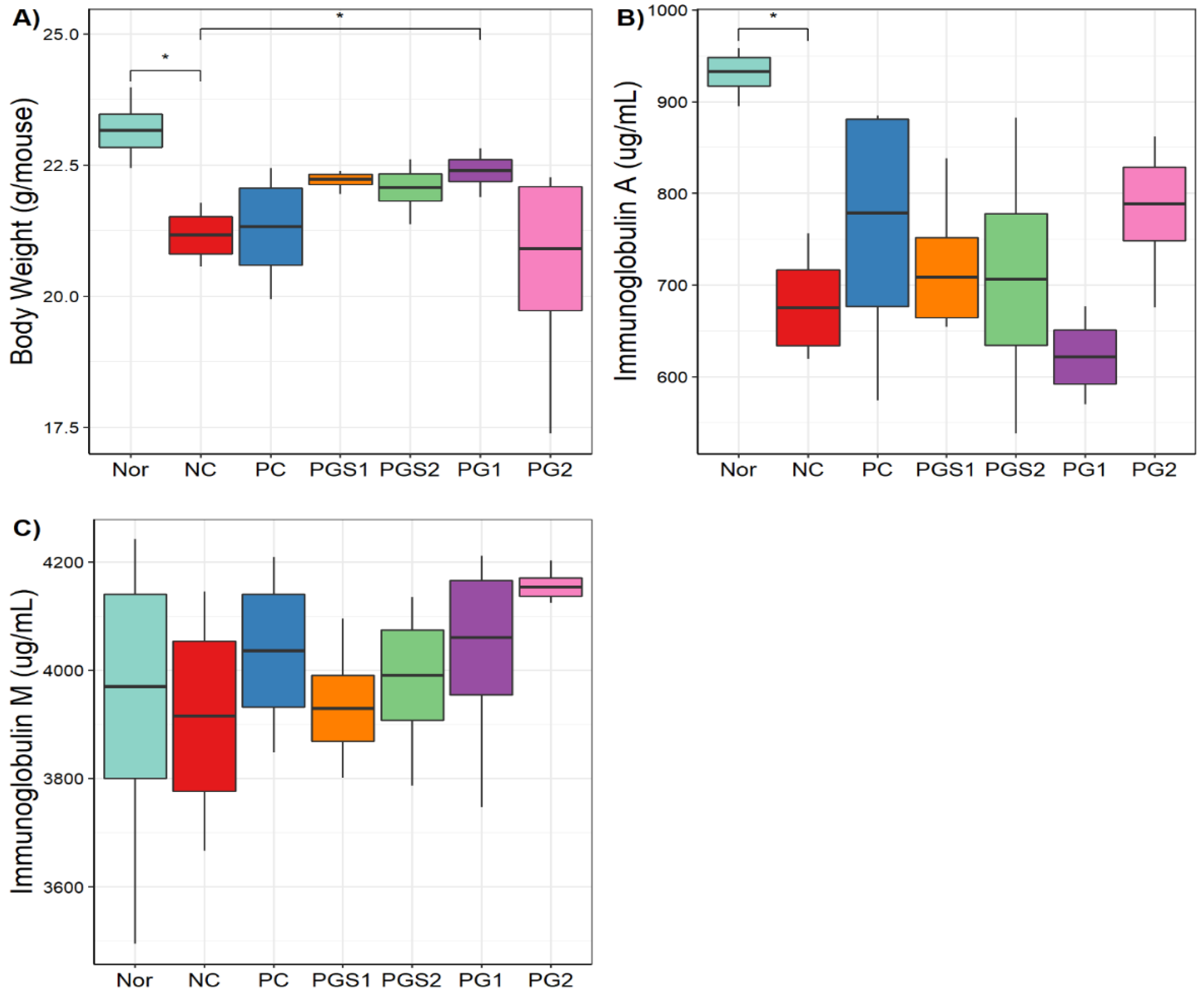
2.3. General Characteristics of Phenotypes in Seven Groups
2.4. Correlation between the Immune-Related Biomarkers and Relative Abundance of Microorganisms
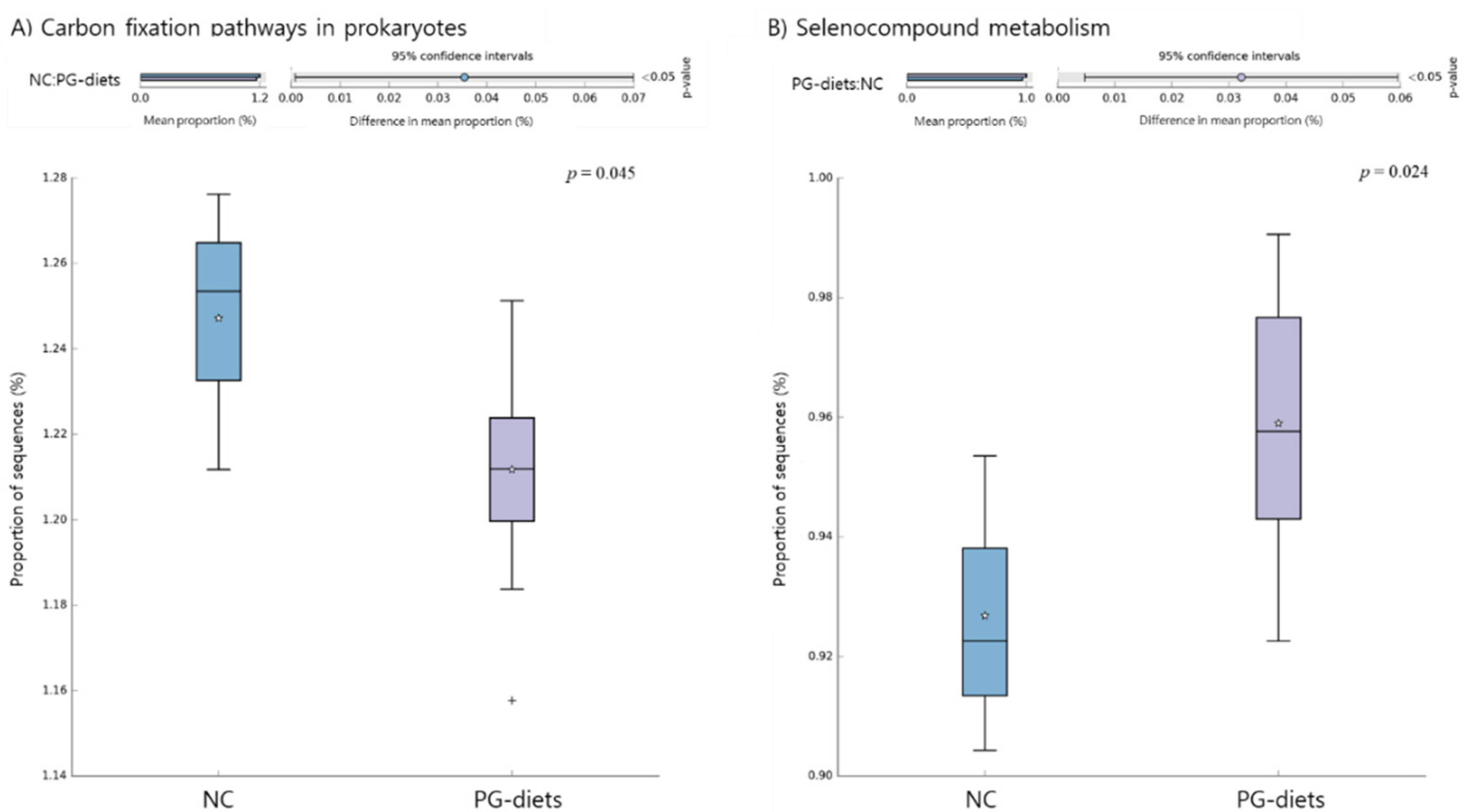
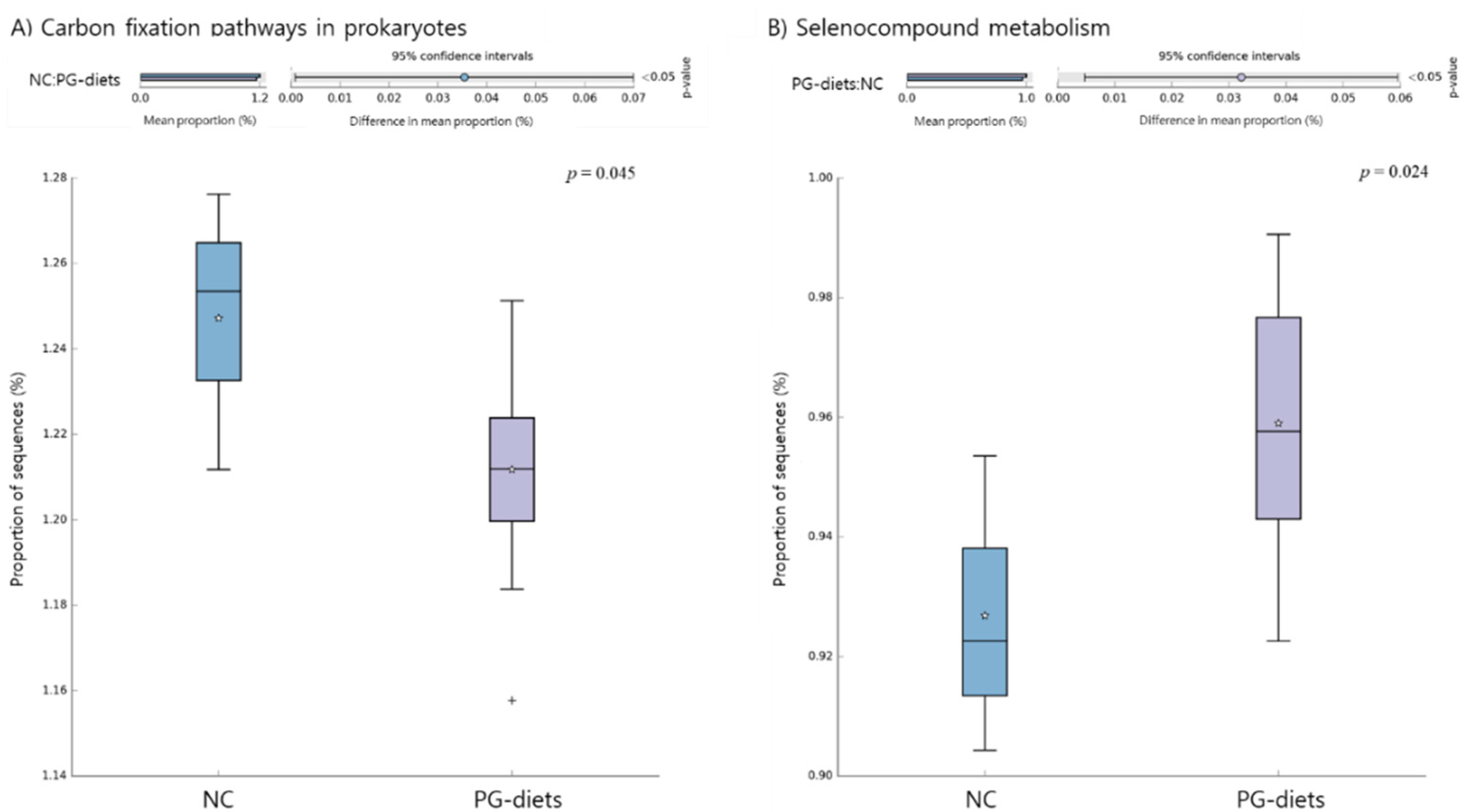
2.5. Functional Pathways of the Microbiome
3. Discussion
4. Materials and Methods
4.1. Preparations of Test Materials
4.2. Animals and Treatments
4.3. Metagenome Sequencing
4.4. Raw Data Processing and Taxonomic Analysis
4.5. Diversity Analysis
4.6. Performance Analysis
4.7. Statistical Analysis
4.8. Functional Profiles of Gut Microbiome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Schley, P.; Field, C. The immune-enhancing effects of dietary fibres and prebiotics. Br. J. Nutr. 2002, 87, S221–S230. [Google Scholar] [CrossRef]
- Lomax, A.R.; Calder, P.C. Prebiotics, immune function, infection and inflammation: A review of the evidence. Br. J. Nutr. 2008, 101, 633–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Tai, W.C.; Hsiao, W.W. Dietary saponins from four popular herbal tea exert prebiotic-like effects on gut microbiota in C57BL/6 mice. J. Funct. Foods 2015, 17, 892–902. [Google Scholar] [CrossRef]
- Lee, J.; Lee, E.; Kim, D.; Lee, J.; Yoo, J.; Koh, B. Studies on absorption, distribution and metabolism of ginseng in humans after oral administration. J. Ethnopharmacol. 2009, 122, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, T.; Ma, C.; Song, W.; Zhang, J.; Yu, Z. Ginsenoside metabolite compound K enhances the efficacy of cisplatin in lung cancer cells. J. Thorac. Dis. 2015, 7, 400. [Google Scholar]
- Kim, K.-A.; Yoo, H.H.; Gu, W.; Yu, D.-H.; Jin, M.J.; Choi, H.-L.; Yuan, K.; Guerin-Deremaux, L.; Kim, D.-H. A prebiotic fiber increases the formation and subsequent absorption of compound K following oral administration of ginseng in rats. J. Ginseng Res. 2015, 39, 183–187. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Brar, M.S.; Leung, F.C.; Hsiao, W.W. Triterpenoid herbal saponins enhance beneficial bacteria, decrease sulfate-reducing bacteria, modulate inflammatory intestinal microenvironment and exert cancer preventive effects in ApcMin/+ mice. Oncotarget 2016, 7, 31226. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-H. Gut microbiota-mediated pharmacokinetics of ginseng saponins. J. Ginseng Res. 2018, 42, 255–263. [Google Scholar] [CrossRef]
- Han, L.-K.; Xu, B.-J.; Kimura, Y.; Zheng, Y.-n.; Okuda, H. Platycodi radix affects lipid metabolism in mice with high fat diet–induced obesity. J. Nutr. 2000, 130, 2760–2764. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.B. Pharmacological studies on Platycodon grandiflorum A. DC. IV. A comparison of experimental pharmacological effects of crude playtcodin with clinical indications of platycodi radix (author’s transl). J. Pharm. Soc. Jpn. 1973, 93, 1188–1194. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.J.; Jeon, S.H.; No, I.R.; Kim, Y.G.; Cho, Y.S. Effect of saponin content and antioxidant activities of Platycodon grandiflorum Radix by cutting length. Korean J. Med. Crop Sci. 2015, 23, 363–369. [Google Scholar] [CrossRef]
- Lee, K.J.; Choi, C.Y.; Chung, Y.C.; Kim, Y.S.; Ryu, S.Y.; Roh, S.H.; Jeong, H.G. Protective effect of saponins derived from roots of Platycodon grandiflorum on tert-butyl hydroperoxide-induced oxidative hepatotoxicity. Toxicol. Lett. 2004, 147, 271–282. [Google Scholar] [CrossRef]
- Kim, J.-W.; Park, S.J.; Lim, J.H.; Yang, J.W.; Shin, J.C.; Lee, S.W.; Suh, J.W.; Hwang, S.B. Triterpenoid saponins isolated from Platycodon grandiflorum inhibit hepatitis C virus replication. Evid. Based Complementary Altern. Med. 2013, 2013, 560417. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.B.; Lee, S.H.; Park, Y.-G.; Choi, J.-H.; Lee, H.K.; Jang, H.H.; Hwang, G.-A.; Choe, J.-S.; Park, S.Y.; Choi, A.J.; et al. Platycodon grandiflorum Extract Ameliorates Cyclophosphamide-Induced Immunosuppression in Mice. J. East Asian Soc. Diet. Life 2019, 29, 303–309. [Google Scholar] [CrossRef]
- Schroeder, H.W., Jr.; Cavacini, L. Structure and function of immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, S41–S52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noh, E.M.; Kim, J.M.; Lee, H.Y.; Song, H.K.; Joung, S.O.; Yang, H.J.; Kim, M.J.; Kim, K.S.; Lee, Y.R. Immuno-enhancement effects of Platycodon grandiflorum extracts in splenocytes and a cyclophosphamide-induced immunosuppressed rat model. BMC Complement. Altern. Med. 2019, 19, 322. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Nie, S.P.; Wang, J.Q.; Liu, X.Z.; Yin, P.F.; Huang, D.F.; Li, W.J.; Gong, D.M.; Xie, M.Y. Chemoprotective effects of Ganoderma atrum polysaccharide in cyclophosphamide-induced mice. Int. J. Biol. Macromol. 2014, 64, 395–401. [Google Scholar] [CrossRef]
- Choi, J.-H.; Lee, E.B.; Park, Y.-G.; Lee, H.K.; Jang, H.H.; Choe, J.; Hwang, K.-A.; Park, S.Y.; Hwang, I.G.; Hong, H.C.; et al. Aged Doraji (Platycodon grandiflorum) Ameliorates Cyclophosphamide-Induced Immunosuppression in Mice. Korean Soc. Pharmacogn. 2019, 50, 219–225. [Google Scholar]
- Zhou, Y.; Chen, X.; Yi, R.; Li, G.; Sun, P.; Qian, Y.; Zhao, X. Immunomodulatory effect of tremella polysaccharides against cyclophosphamide-induced immunosuppression in mice. Molecules 2018, 23, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandvik, A.; Wang, Y.; Morton, H.; Aasen, A.; Wang, J.; Johansen, F.E. Oral and systemic administration of β-glucan protects against lipopolysaccharide-induced shock and organ injury in rats. Clin. Exp. Immunol. 2007, 148, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Jeong, D.Y.; Yang, H.J.; Jeong, S.J.; Kim, M.G.; Yun, C.Y.; Lee, H.Y.; Lee, Y.H.; Shin, D.Y.; Yang, Y.G.; Lee, H.S.; et al. Immunostimulatory effects of blue-berry yeast fermented powder aginst cyclophosphamide-induced immunosuppressed model. J. Physiol Pathnol. Krean Med. 2019, 33, 48–55. [Google Scholar]
- Lee, Y.S.; Lee, G.H.; Kwon, Y.K.; Park, J.H.; Shin, S.W. Immunomodulatory effect of aqueous extracted Zingiberis Rhizoma on cyclophosphamide—Induced immune suppresion. Korean J. Orient. Physiol. Pathol. 2007, 21, 485–490. [Google Scholar]
- Lee, Y.S.; Lee, G.H.; Park, J.H.; Kwon, Y.K.; Shin, S.W. Water extracted Evodiae Fructus Possesses immunomodulatory activities on cyclophosphamide induced immunesuppression. Korean J. Physiol. Pathol. 2007, 21, 1450–1455. [Google Scholar]
- Chung, J.H.; Shin, P.G.; Ryu, J.C.; Jang, D.S.; Cho, S.H. Chemical Compositions of Platycodon grandiflorus (jacquin). A. De Candolle Agric. Chem. Biotechnol. 1997, 40, 148–151. [Google Scholar]
- Shon, M.Y.; Seo, J.K.; Kiom, H.J.; Sung, N.J. Chemical compositions and physiological activities of doraji (Platycodon grandiflorum). J. Korean Soc. Food Sci. Nutr. 2001, 30, 717–720. [Google Scholar]
- Hong, M.W. Statistical analyeses of Platycodi Radix prescriptions. Kor. J. Parmacog. 1974, 5, 61–67. [Google Scholar]
- Kim, E.H.; Gwak, J.Y.; Jung, M.J. Immunomodulatory activity of Platycodon grandiflorum, Codonopsis lanceolata, and Adenophora triphylla extracts in macrophage cells. J. Korean Soc. Food Sci. Nutr. 2018, 47, 1069–1075. [Google Scholar] [CrossRef]
- Tohamy, A.A.; El-Ghor, A.A.; El-Nahas, S.M.; Noshy, M.M. Beta-glucan inhibits the genotoxicity of cyclophosphamide, adriamycin and cisplatin. Mutat. Res. 2003, 541, 45–53. [Google Scholar] [CrossRef]
- Kirmaz, C.; Bayrak, P.; Yilmaz, O.; Yuksel, H. Effects of glucan treatment on the Th1/Th2 balance in patients with allergic rhinitis: A double-blind placebo-controlled study. Eur. Cytokine Netw. 2005, 16, 128–134. [Google Scholar] [PubMed]
- Naito, Y.; Uchiyama, K.; Takagi, T. A next-generation beneficial microbe: Akkermansia muciniphila. J. Clin. Biochem. Nutr. 2018, 63, 33–35. [Google Scholar] [CrossRef] [Green Version]
- Shin, N.R.; Lee, J.C.; Lee, H.Y.; Kim, M.S.; Whon, T.W.; Lee, M.S.; Bae, J.W. An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut 2014, 63, 727–735. [Google Scholar] [CrossRef] [Green Version]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Carbonero, F.; Benefiel, A.C.; Alizadeh-Ghamsari, A.H.; Gaskins, H.R. Microbial pathways in colonic sulfur metabolism and links with health and disease. Front. Physiol. 2012, 3, 448. [Google Scholar] [CrossRef] [Green Version]
- Kasaikina, M.V.; Kravtsova, M.A.; Lee, B.C.; Seravalli, J.; Peterson, D.A.; Walter, J.; Legge, R.; Benson, A.K.; Hatfield, D.L.; Gladyshev, V.N. Dietary selenium affects host selenoproteome expression by influencing the gut microbiota. FASEB J. 2011, 25, 2492–2499. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Chang, Q.; Luan, Y.; Sun, F. Variance adjusted weighted UniFrac: A powerful beta diversity measure for comparing communities based on phylogeny. BMC Bioinform. 2011, 12, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noma, H.; Nagashima, K.; Furukawa, T.A. Permutation inference methods for multivariate meta-analysis. Biometrics 2020, 76, 337–347. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Vegan: Community Ecology Package. 2019. Available online: https://github.com/vegandevs/vegan (accessed on 28 November 2021).
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Series B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbo, M.; Knight, R.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [Green Version]
- Team, R. RStudio: Integrated Development Environment for R; Team, R: Boston, MA, USA, 2020. [Google Scholar]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Louca, S.; Doebeli, M. Efficient comparative phylogenetics on large trees. Bioinformatics 2018, 34, 1053–1055. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.R.; Nagarajan, N.; Pop, M. Statistical methods for detecting differentially abundant features in clinical metagenomic samples. PLoS Comput. Biol. 2009, 5, e1000352. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jhang, S.-Y.; Lee, S.-H.; Lee, E.-B.; Choi, J.-H.; Bang, S.; Jeong, M.; Jang, H.-H.; Kim, H.-J.; Lee, H.-J.; Jeong, H.-C.; et al. Effects of Platycodon grandiflorum on Gut Microbiome and Immune System of Immunosuppressed Mouse. Metabolites 2021, 11, 817. https://doi.org/10.3390/metabo11120817
Jhang S-Y, Lee S-H, Lee E-B, Choi J-H, Bang S, Jeong M, Jang H-H, Kim H-J, Lee H-J, Jeong H-C, et al. Effects of Platycodon grandiflorum on Gut Microbiome and Immune System of Immunosuppressed Mouse. Metabolites. 2021; 11(12):817. https://doi.org/10.3390/metabo11120817
Chicago/Turabian StyleJhang, So-Yun, Sung-Hyen Lee, Eun-Byeol Lee, Ji-Hye Choi, Sohyun Bang, Misun Jeong, Hwan-Hee Jang, Hyun-Ju Kim, Hae-Jeung Lee, Hyun-Cheol Jeong, and et al. 2021. "Effects of Platycodon grandiflorum on Gut Microbiome and Immune System of Immunosuppressed Mouse" Metabolites 11, no. 12: 817. https://doi.org/10.3390/metabo11120817
APA StyleJhang, S.-Y., Lee, S.-H., Lee, E.-B., Choi, J.-H., Bang, S., Jeong, M., Jang, H.-H., Kim, H.-J., Lee, H.-J., Jeong, H.-C., & Lee, S.-J. (2021). Effects of Platycodon grandiflorum on Gut Microbiome and Immune System of Immunosuppressed Mouse. Metabolites, 11(12), 817. https://doi.org/10.3390/metabo11120817