Exercise Induced Changes in Salivary and Serum Metabolome in Trained Standardbred, Assessed by 1H-NMR

 , ,
, ,
Abstract
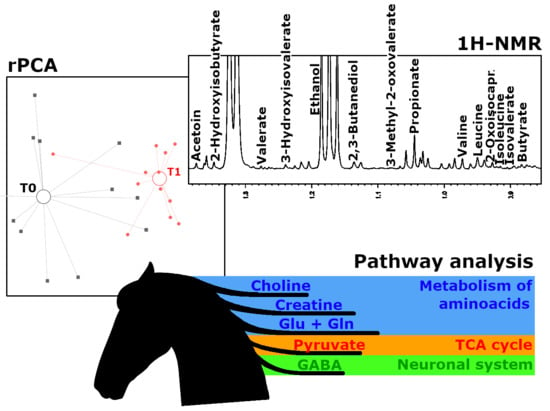
1. Introduction
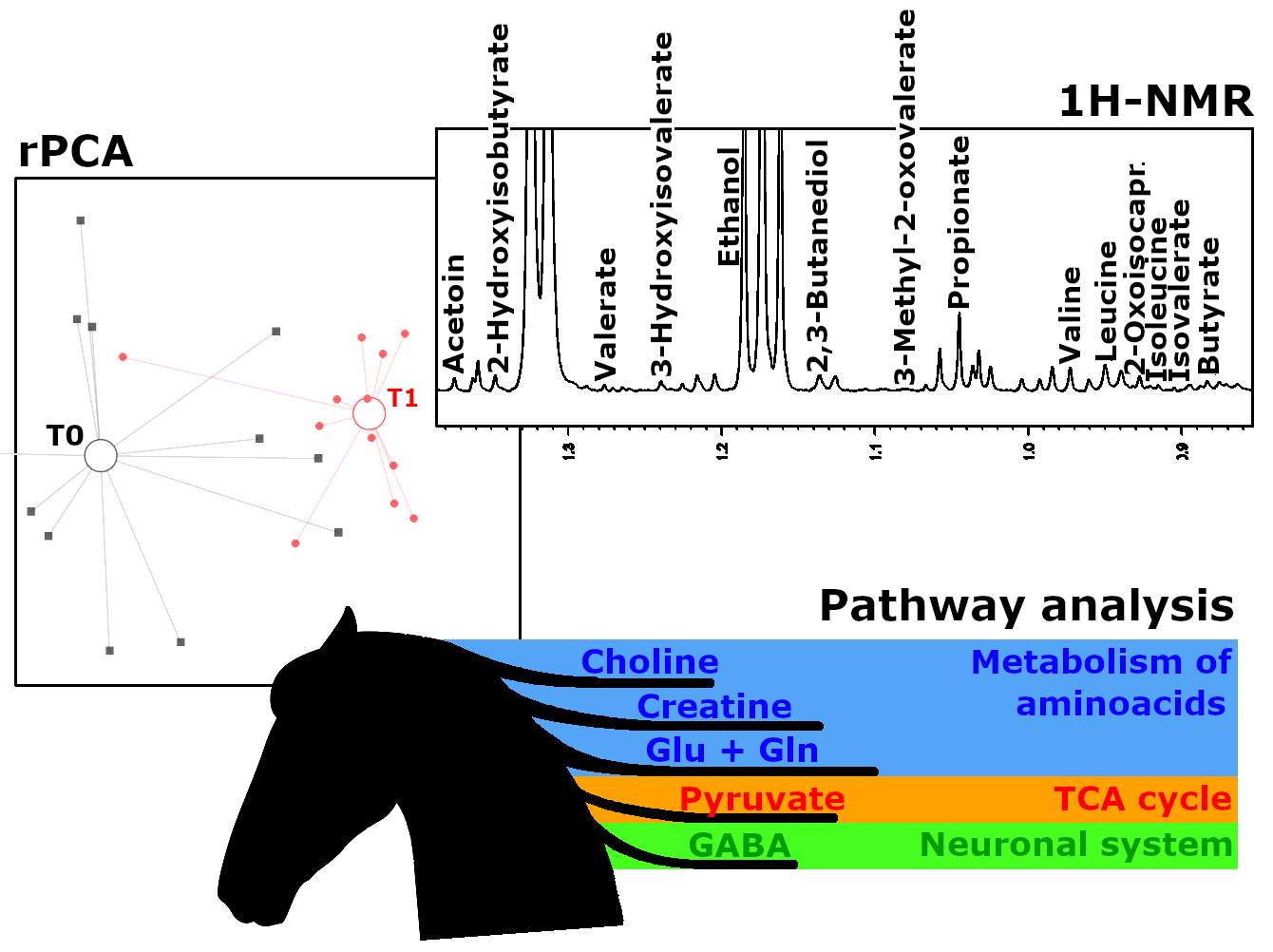
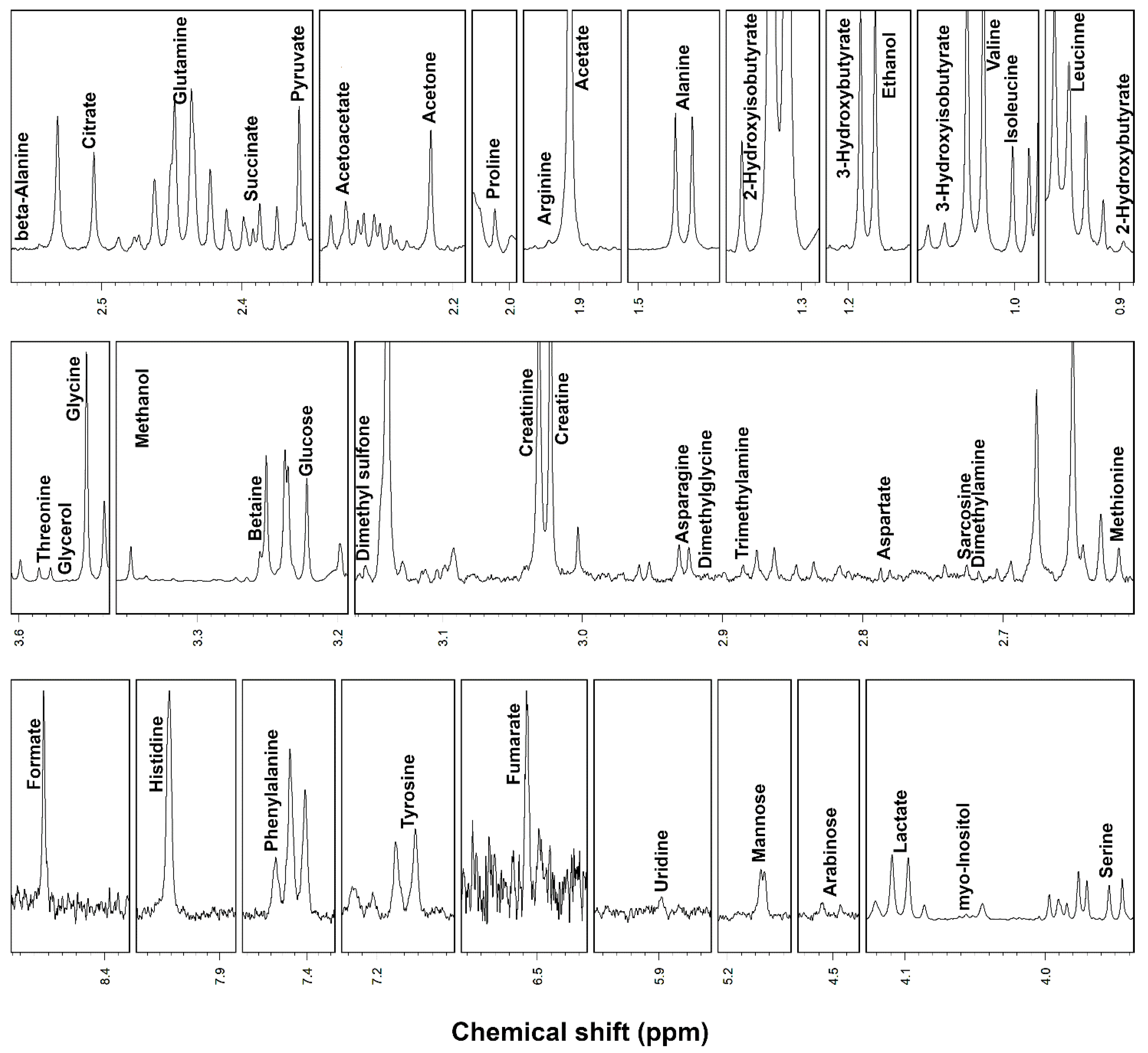
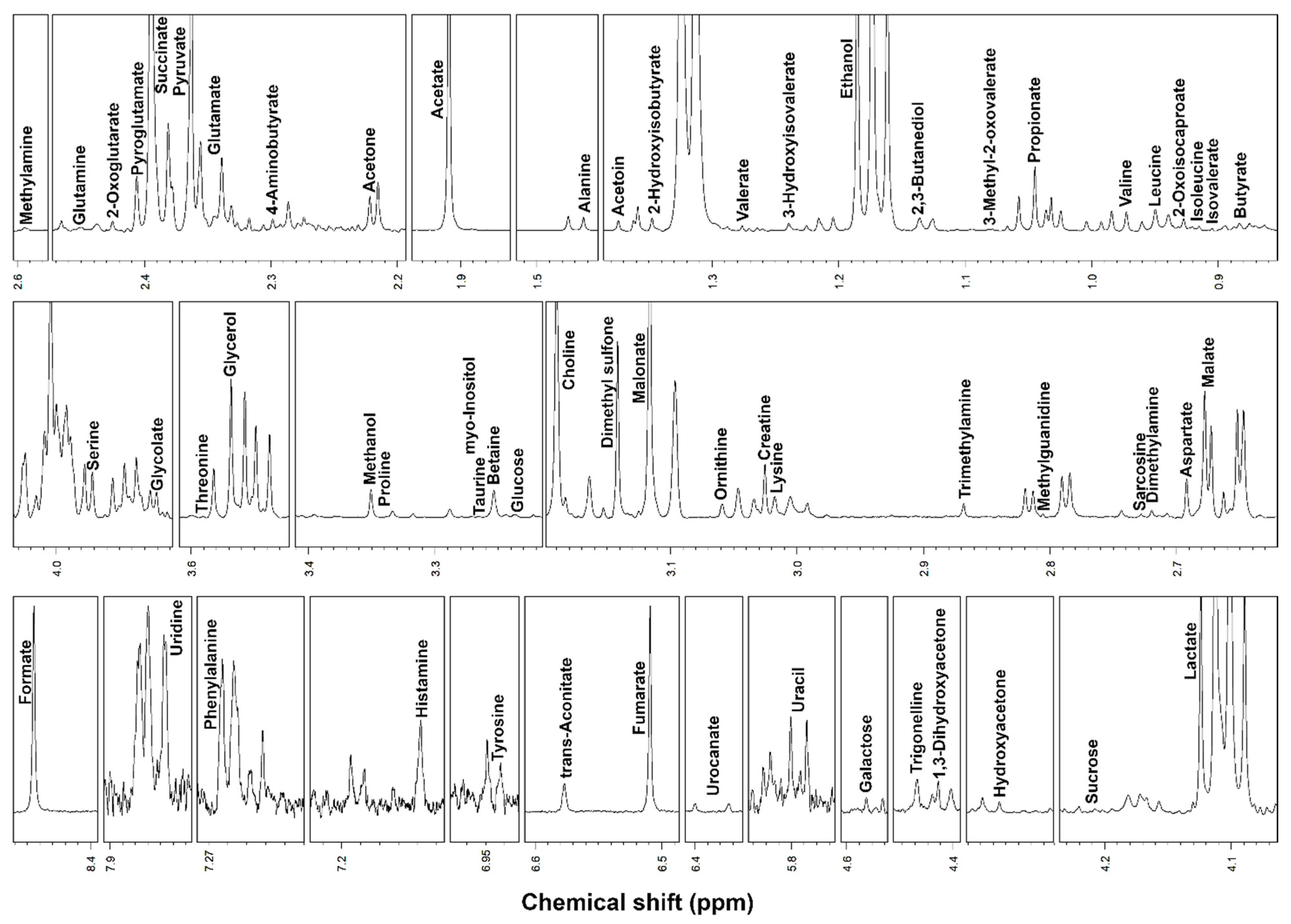
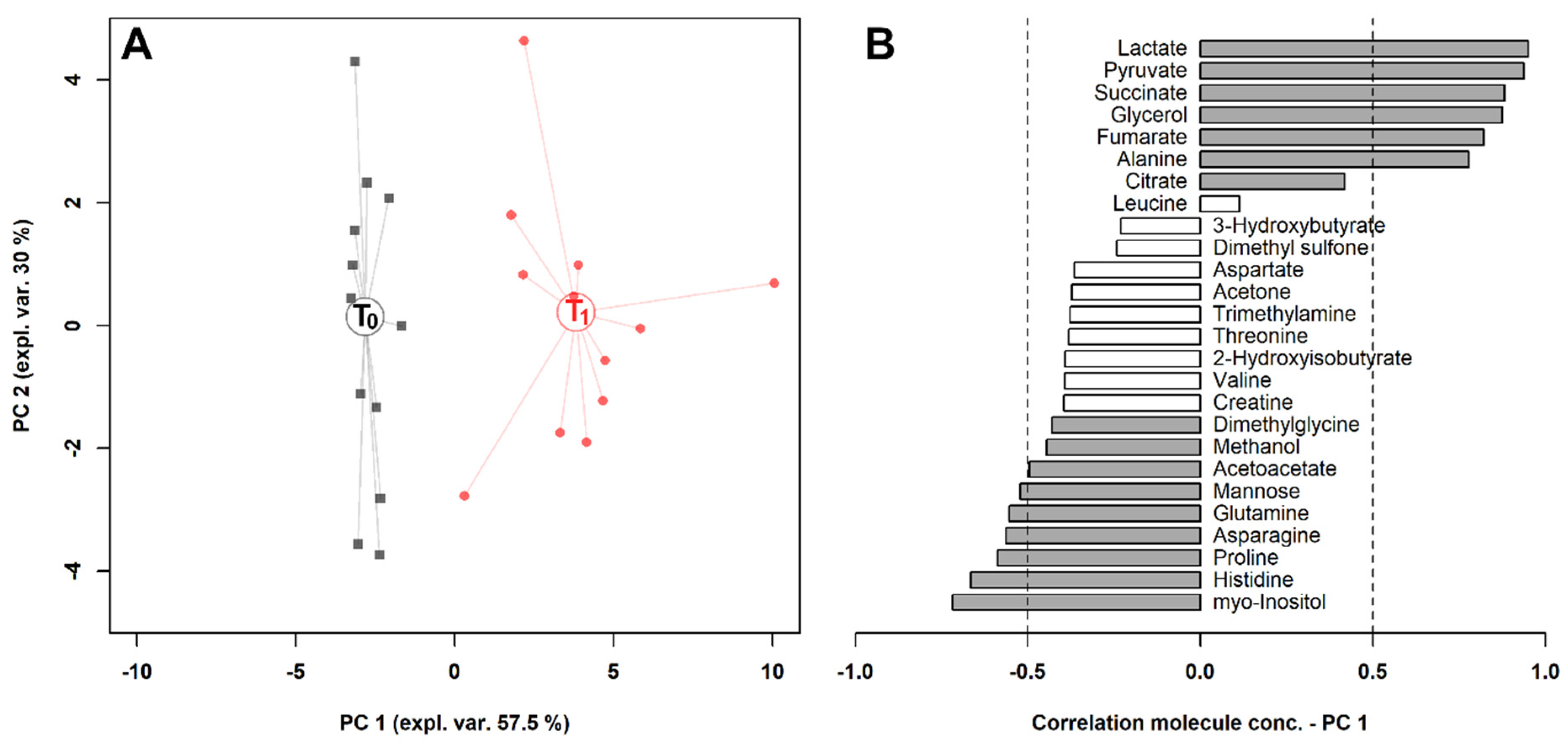
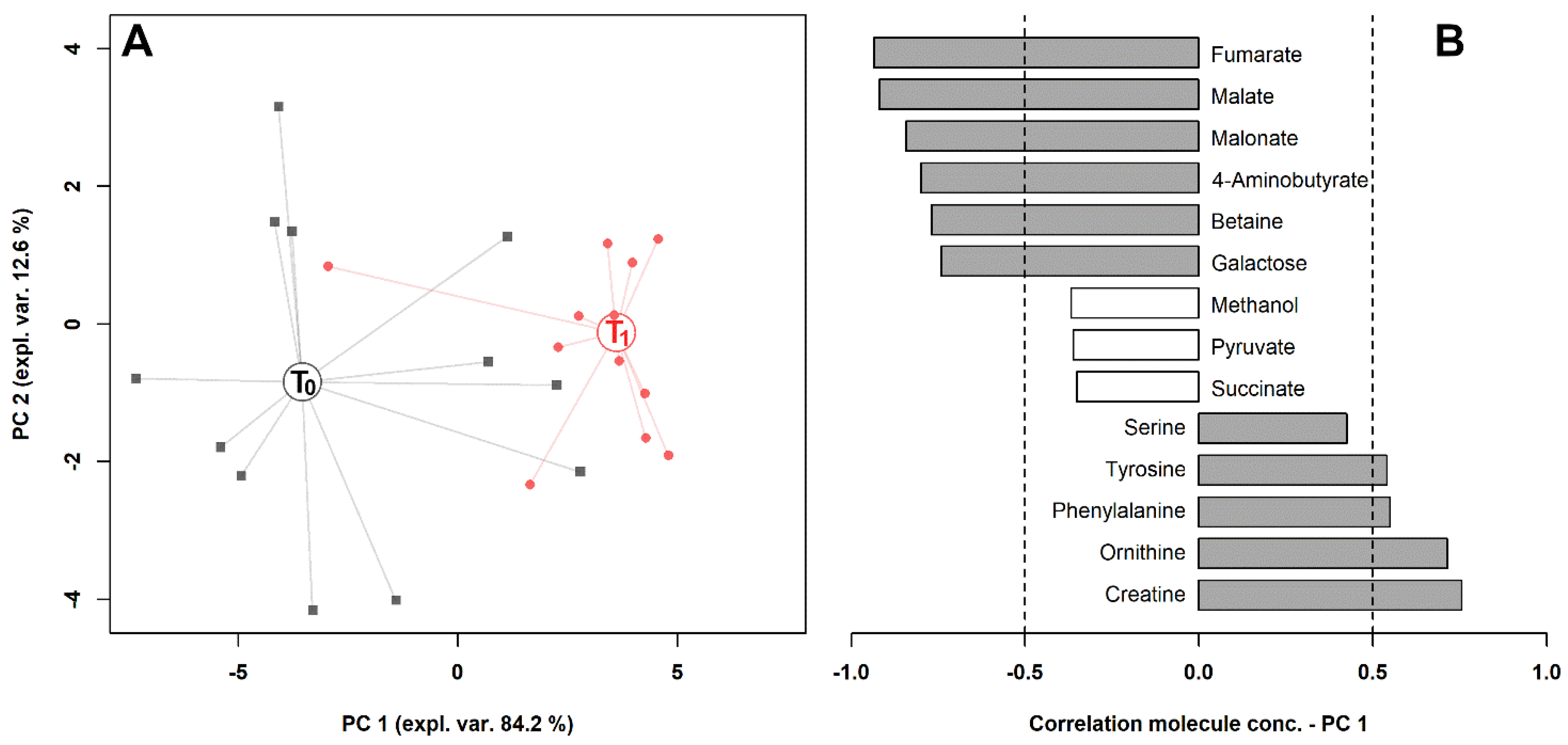
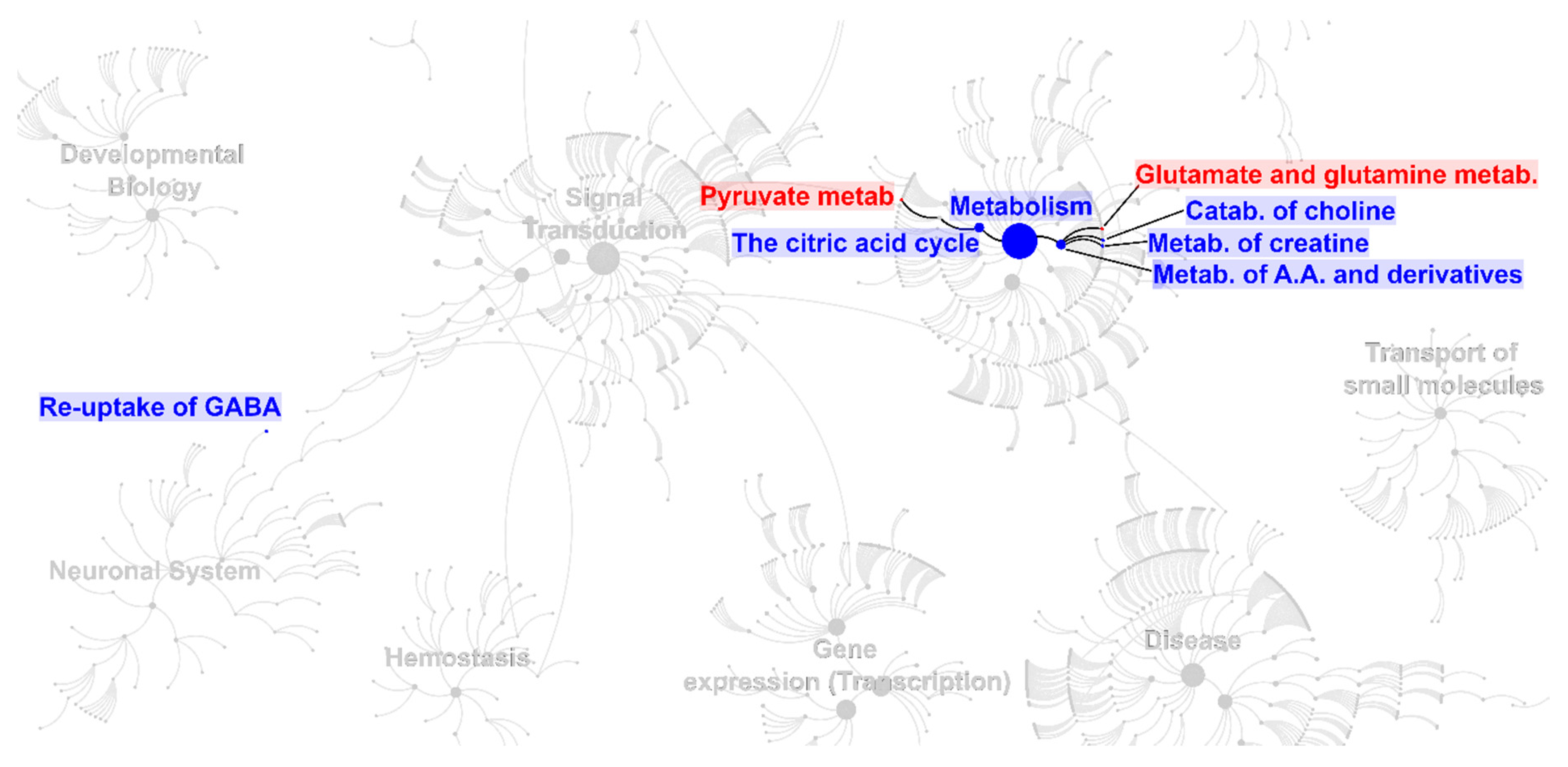
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Procedure
4.3. Laboratory Analysis.
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dunn, W.B.; Broadhurst, D.I.; Atherton, H.J.; Goodacre, R.; Griffin, J.L. Systems level studies of mammalian metabolomes: The roles of mass spectrometry and nuclear magnetic resonance spectroscopy. Chem. Soc. Rev. 2011, 40, 387–426. [Google Scholar] [CrossRef] [PubMed]
- Gorostiaga, E.M.; Navarro-Amezqueta, I.; Calbet, J.A.; Hellsten, Y.; Cusso, R.; Guerrero, M.; Granados, C.; González-Izal, M.; Ibañez, J.; Izquierdo, M. Energy metabolism during repeated sets of leg press exercise leading to failure or not. PLoS ONE 2012, 7, e40621. [Google Scholar] [CrossRef]
- Schoenfeld, B.J. Potential mechanisms for a role of metabolic stress in hypertrophic adaptations to resistance training. Sports Med. 2013, 43, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Tesch, P.A.; Colliander, E.B.; Kaiser, P. Muscle metabolism during intense, heavy-resistance exercise. Eur. J. Appl. Physiol. Occup. Physiol. 1986, 55, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Villanueva, A.; Edge, J.; Suriano, R.; Hamer, P.; Bishop, D. The recovery of repeated-sprint exercise is associated with PCr resynthesis, while muscle pH and EMG amplitude remain depressed. PLoS ONE 2012, 7, e51977. [Google Scholar] [CrossRef] [PubMed]
- Psychogios, N.; Hau, D.D.; Peng, J.; Guo, A.C.; Mandal, R.; Bouatra, S.; Young, N. The human serum metabolome. PLoS ONE 2011, 6, 16957. [Google Scholar] [CrossRef]
- Ryan, D.; Robards, K. Metabolomics: The greatest omics of them all? Anal. Chem. 2006, 78, 7954–7958. [Google Scholar] [CrossRef]
- Berton, R.; Conceição, M.S.; Libardi, C.A.; Canevarolo, R.R.; Gáspari, A.F.; Chacon-Mikahil, M.P.; Zeri, A.C.; Cavaglieri, C.R. Metabolic time-course response after resistance exercise: A metabolomics approach. J. Sports Sci. 2017, 35, 1211–1218. [Google Scholar] [CrossRef]
- Contreras-Aguilar, M.D.; Escribano, D.; Martínez-Subiela, S.; Martín-Cuervo, M.; Lamy, E.; Tecles, F.; Cerón, J.J. Changes in saliva analytes in equine acute abdominal disease: A sialochemistry approach. BMC Vet. Res. 2019, 15, 187. [Google Scholar] [CrossRef]
- Ghizoni, J.S.; Nichele, R.; de Oliveira, M.T.; Pamato, S.; Pereira, J.R. The utilization of saliva as an early diagnostic tool for oral cancer: microRNA as a biomarker. Clin. Transl. Oncol. 2020, 22, 804–812. [Google Scholar] [CrossRef]
- Greabu, M.; Battino, M.; Mohora, M.; Totan, A.; Didilescu, A.; Spinu, T. Saliva—A diagnostic window to the body, both in health and in disease. J. Med. Life 2009, 2, 124–132. [Google Scholar] [PubMed]
- Tecles, F.; Escribano, D.; Martínez-Miró, S.; Hernández, F.; Contreras, M.D.; Cerón, J.J. Cholinesterase in porcine saliva: Analytical characterization and behavior after experimental stress. Res. Vet. Sci. 2016, 106, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Tecles, F.; Contreras-Aguilar, M.D.; Martínez-Miró, S.; Tvarijonaviciute, A.; Martínez-Subiela, S.; Escribano, D.; Cerón, J.J. Total esterase measurement in saliva of pigs: Validation of an automated assay, characterization and changes in stress and disease conditions. Res. Vet. Sci. 2017, 114, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Tecles, F.; Rubio, C.P.; Contreras-Aguilar, M.D.; Lopez-Arjona, M.; Martinez-Miro, S.; Martinez-Subiela, S.; Cerón, J.J. Adenosine deaminase activity in pig saliva: Analytical validation of two spectrophotometric assays. J. Vet. Diagn. Investig. 2018, 30, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Aguilar, M.D.; Escribano, D.; Quiles, A.; López-Arjona, M.; Cerón, J.J.; Martínez-Subiela, S.; Hevia, M.L.; Tecles, F. Evaluation of new biomarkers of stress in saliva of sheep. Animal 2019, 13, 1278–1286. [Google Scholar] [CrossRef]
- Contreras-Aguilar, M.D.; Escribano, D.; Martín-Cuervo, M.; Tecles, F.; Cerón, J.J. Salivary alpha-amylase activity and cortisol in horses with acute abdominal disease: A pilot study. BMC Vet. Res. 2018, 14, 156. [Google Scholar] [CrossRef]
- Barranco, T.; Tvarijonaviciute, A.; Tecles, F.; Carrillo, J.M.; Sanchez-Resalt, C.; Jimenez-Reyes, P.; Rubio, M.; García-Balletbó, M.; Cerón, J.; Ramon Cugat, R. Changes in creatine kinase, lactate dehydrogenase and aspartate aminotransferase in saliva samples after an intense exercise: A pilot study. J. Sports Med. Phys. 2018, 58, 910–916. [Google Scholar]
- Jacobsen, S.; Top Adler, D.M.; Bundgaard, L.; Sørensen, M.A.; Andersen, P.H.; Bendixen, E. The use of liquid chromatography tandem mass spectrometry to detect proteins in saliva from horses with and without systemic inflammation. Vet. J. 2014, 202, 483–488. [Google Scholar]
- Bilancio, G.; Cavallo, P.; Lombardi, C.; Guarino, E.; Cozza, V.; Giordano, F.; Palladino, G.; Cirillo, M. Salivary levels of phosphorus and urea as indices of their plasma levels in nephropathic patients. J. Clin. Lab. Anal. 2018, 32, 1–6. [Google Scholar] [CrossRef]
- de Sousa-Pereira, P.; Cova, M.; Abrantes, J.; Ferreira, R.; Trindade, F.; Barros, A.; Gomes, P.; Colaço, B.; Amado, F.; Esteves, P.J.; et al. Cross-species comparison of mammalian saliva using an LC-MALDI based proteomic approach. Proteomics 2015, 15, 1598–1607. [Google Scholar] [CrossRef]
- Pitti, E.; Petrella, G.; Di Marino, S.; Summa, V.; Perrone, M.; D’Ottavio, S.; Bernardini, A.; Cicero, D.O. Salivary Metabolome and Soccer Match: Challenges for Understanding Exercise induced Changes. Metabolites 2019, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Navas de Solis, C. Cardiovascular Response to Exercise and Training, Exercise Testing in Horses. Vet. Clin. N. Am. Equine Pract. 2019, 35, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Bazzano, M.; Giudice, E.; Rizzo, M.; Congiu, F.; Zumbo, A.; Arfuso, F.; Di Pietro, S.; Bruschetta, D.; Piccione, G. Application of a combined global positioning and heart rate monitoring system in jumper horses during an official competition—A preliminary study. Acta Vet. Hung. 2016, 64, 189–200. [Google Scholar] [PubMed]
- Munsters, C.C.; van Iwaarden, A.; van Weeren, R.; Sloet van Oldruitenborgh-Oosterbaan, M.M. Exercise testing in Warmblood sport horses under field conditions. Vet. J. 2014, 202, 11–19. [Google Scholar] [PubMed]
- Luck, M.M.; Le Moyec, L.; Barrey, E.; Triba, M.N.; Bouchemal, N.; Savarin, P.; Robert, C. Energetics of endurance exercise in young horses determined by nuclear magnetic resonance metabolomics. Front. Physiol. 2015, 6, 198. [Google Scholar]
- Mach, N.; Ramayo-Caldas, Y.; Clark, A.; Moroldo, M.; Robert, C.; Barrey, E.; Lòpez, J.M.; Le Moyec, L. Understanding the response to endurance exercise using a systems biology approach: Combining blood metabolomics, transcriptomics and miRNomics in horses. BMC Genom. 2017, 18, 187. [Google Scholar] [CrossRef]
- Laghi, L.; Picone, G.; Capozzi, F. Nuclear magnetic resonance for foodomics beyond food anaysis. Trend Anal. Chem. 2014, 59, 93–102. [Google Scholar]
- Wishart, D.S.; Jewison, T.; Guo, A.C.; Wilson, M.; Knox, C.; Liu, Y.; Scalbert, A. HMDB 3.0—The human metabolome database in 2013. Nucleic Acids Res. 2013, 41, D801–D807. [Google Scholar] [CrossRef]
- Nieman, D.C.; Shanely, R.A.; Gillitt, N.D.; Pappan, K.L.; Lila, M.A. Serum metabolic signatures induced by a three-day intensified exercise period persist after 14 h of recovery in runners. J. Proteome Res. 2013, 12, 4577–4584. [Google Scholar]
- Lewis, G.D.; Farrell, L.; Wood, M.J.; Martinovic, M.; Arany, Z.; Rowe, G.C.; Souza, A.; Cheng, S.; McCabe, E.L.; Yang, E.; et al. Metabolic signatures of exercise in human plasma. Sci. Transl. Med. 2010, 2, 33–37. [Google Scholar]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 7th ed.; International ed.; W.H. Freeman & Company: New York, NY, USA, USA; 2016; p. 612. [Google Scholar]
- Gorostiaga, E.M.; Navarro-Amezqueta, I.; Calbet, J.A.; Sanchez-Medina, L.; Cusso, R.; Guerrero, M.; Izquierdo, M. Blood ammonia and lactate as markers of muscle metabolites during leg press exercise. J. Strength Cond. Res. 2014, 28, 2775–2785. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.A. Carbohydrate Metabolism: Major Metabolic Pathways and their Control. In Textbook of Biochemistry with Clinical Correlation, 4th ed.; Devlin, T.M., Ed.; Wiley-Liss: New York, NY, USA, 1997; pp. 268–317. [Google Scholar]
- Krog-Mikkelsen, I.; Hels, O.; Tetens, I.; Holst, J.J.; Andersen, J.R.; Bukhave, K. The effects of L-arabinose on intestinal sucrase activity: Dose-response studies in vitro and in humans. Am. J. Clin. Nutr. 2011, 94, 472–478. [Google Scholar] [CrossRef]
- Décombaz, J.; Jentjens, R.; Ith, M.; Scheurer, E.; Buehler, T.; Jeukendrup, A.; Boesch, C. Fructose and galactose enhance postexercise human liver glycogen synthesis. Med. Sci. Sports Exerc. 2011, 43, 1964–1971. [Google Scholar] [PubMed]
- Felig, P.W. Amino acid metabolism in exercising man. J. Clin. Investig. 1971, 50, 2703–2714. [Google Scholar] [CrossRef]
- Coomes, M.W. Amino Acid Metabolism. In Textbook of Biochemistry with Clinical Correlation, 4th ed.; Devlin, T.M., Ed.; Wiley-Liss: New York, NY, USA, 1997; pp. 446–483. [Google Scholar]
- Sutton, E.E.; Coill, M.R.; Deuster, P.A. Ingestion of tyrosine: Effects on endurance, muscle strength, and anaerobic performance. Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Pundir, C.S.; Deswal, R.; Kumar, P. Quantitative analysis of sarcosine with special emphasis on biosensors: A review. Biomarkers 2019, 24, 415–422. [Google Scholar]
- Bazzano, M.; Laghi, L.; Zhu, C.; Magi, G.E.; Serri, E.; Spaterna, A.; Tesei, B.; Laus, F. Metabolomics of tracheal wash samples and exhaled breath condensates in healthy horses and horses affected by equine asthma. J. Breath Res. 2018, 12, 046015. [Google Scholar] [CrossRef]
- Olson, M.S. Bioenergetics and Oxidative Metabolism. In Textbook of Biochemistry with Clinical Correlation, 4th ed.; Devlin, T.M., Ed.; Wiley-Liss: New York, NY, USA, 1997; pp. 218–261. [Google Scholar]
- Dame, Z.T.; Aziat, F.; Mandal, R.; Krishnamurthy, R.; Bouatra, S.; Borzouie, S.; Guo, A.C.; Tanvir Sajed, T.; Deng, L.; Lin, H.; et al. The human saliva metabolome. Metabolomics 2015, 11, 1864–1883. [Google Scholar] [CrossRef]
- Duarte, I.F.; Goodfellow, B.J.; Barros, A.; Jones, J.G.; Barosa, C.; Diogo, L.; Garcia, P.; Gil, A.M. Metabolic characterisation of plasma in juveniles with glycogen storage disease type 1a (GSD1a) by high-resolution (1)H NMR spectroscopy. NMR Biomed. 2007, 20, 401–412. [Google Scholar] [CrossRef]
- Laborde, C.M.; Mourino-Alvarez, L.; Posada-Ayala, M.; Alvarez-Llamas, G.; Serranillos-Reus, M.G.; Moreu, J.; Vivanco, F.; Padial, L.R.; Barderas, M.G. Plasma metabolomics reveals a potential panel of biomarkers for early diagnosis in acute coronary syndrome. Metabolomics 2014, 10, 414–424. [Google Scholar] [CrossRef]
- Trushina, E.; Dutta, T.; Persson, X.M.; Mielke, M.M.; Petersen, R.C. Identification of altered metabolic pathways in plasma and CSF in mild cognitive impairment and Alzheimer’s disease using metabolomics. PLoS ONE 2013, 8, e63644. [Google Scholar] [CrossRef] [PubMed]
- Koolman, J.; Roehm, K.H. Color. Atlas of Biochemistry, 2nd ed.; Thieme: Stuttgart, Germany, 2005; p. 352. [Google Scholar]
- Marukawa, H.; Shimomura, T.; Takahashi, K. Salivary substance P, 5-hydroxytryptamine, and gamma-aminobutyric acid levels in migraine and tension-type headache. Headache 1996, 36, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Jezewska, E.; Scinska, A.; Kukwa, W.; Sobolewska, A.; Turzynska, D.; Samochowiec, J.; Bienkowski, P. Gamma-aminobutyric acid concentrations in benign parotid tumours and unstimulated parotid saliva. J. Laryngol. Otol. 2011, 125, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Sawaki, K.; Ouchi, K.; Sato, T.; Kawaguchi, M. Existence of gamma-aminobutyric acid and its biosynthetic and metabolic enzymes in rat salivary glands. Jpn. J. Pharmacol. 1995, 67, 359–363. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Okubo, M.; Kawaguchi, M. Rat submandibular gland perfusion method for clarifying inhibitory regulation of GABAA receptor. J. Pharmacol. Sci. 2013, 122, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Bizzarri, M.; Fuso, A.; Dinicola, S.; Cucina, A.; Bevilacqua, A. Pharmacodynamics and pharmacokinetics of inositol(s) in health and disease. Expert Opin. Drug Metab. Toxicol. 2016, 12, 1181–1196. [Google Scholar] [CrossRef]
- Gilmore, A.P.; Burridge, K. Regulation of vinculin binding to talin and actin by phosphatidylinositol-4-5-bisphosphate. Nature 1996, 381, 531–535. [Google Scholar] [CrossRef]
- Hallman, M.; Spragg, R.; Harrell, J.H.; Moser, K.M.; Gluck, L. Evidence of lung surfactant abnormality in respiratory failure. Study of bronchoalveolar lavage phospholipids, surface activity, phospholipase activity, and plasma myoinositol. J. Clin. Investig. 1982, 70, 673–683. [Google Scholar] [CrossRef]
- Benesch, R.; Benesch, R.E. The effect of organic phosphates from the human erythrocyte on the allosteric properties of hemoglobin. Biochem. Biophys. Res. Commun. 1967, 26, 162–167. [Google Scholar] [CrossRef]
- Lam, G.; Zhao, S.; Sandhu, J.; Yi, R.; Loganathan, D.; Morrissey, B. Detection of myo-inositol tris pyrophosphate (ITPP) in equine following an administration of ITPP. Drug Test. Anal. 2014, 6, 268–276. [Google Scholar]
- Bazzano, M.; Laghi, L.; Zhu, C.; Magi, G.E.; Tesei, B.; Laus, F. Respiratory metabolites in bronchoalveolar lavage fluid (BALF) and exhaled breath condensate (EBC) can differentiate horses affected by severe equine asthma from healthy horses. BMC Vet. Res. 2020. under review. [Google Scholar] [CrossRef] [PubMed]
- Carroll, C.L.; Huntington, P.J. Body condition scoring and weight estimation of horses. Equine Vet. J. 1988, 20, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Faillace, V.; Laus, F.; Bazzano, M.; Laghi, L. Characterization of trotter horses urine metabolome by means of proton nuclear magnetic resonance spectroscopy. Metabolomics 2018, 14, 106. [Google Scholar] [CrossRef] [PubMed]
- Kneen, M.A.; Annegarn, H.J. Algorithm for fitting XRF, SEM and PIXE X-ray spectra backgrounds. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 1996, 109–110, 209–213. [Google Scholar] [CrossRef]
- Box, G.E.P.; Cox, D.R. An Analysis of Transformations. J. R. Stat. Soc. Ser. B 2018, 26, 211–243. [Google Scholar] [CrossRef]
- Hubert, M.; Rousseeuw, P.J.; Vanden Branden, K. ROBPCA: A new approach to robust principal component analysis. Technometrics 2005, 47, 64–79. [Google Scholar] [CrossRef]
- Croft, D.; Mundo, A.F.; Haw, R.; Milacic, M.; Weiser, J.; Wu, G.; Jassal, B. The Reactome pathway knowledgebase. Nucleic Acids Res. 2014, 42, D472–D477. [Google Scholar] [CrossRef]
- Johnson, N.L.; Kotz, S.; Kemp, A.W. Univariate Discrete Distributions, 2nd ed.; Wiley: New York, NY, USA, 2005. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | T0 | T1 | p | Trend * |
|---|---|---|---|---|
| 3-Hydroxybutyrate | 2.09 ± 1.57 | 1.52 ± 1.33 | 4.58 × 10−2 | ↓ |
| 2-Hydroxyisobutyrate | 1.64 ± 0.96 | 0.95 ± 1.19 | 6.19 × 10−5 | ↓ |
| Acetoacetate | 3.73 ± 0.73 | 3.21 ± 0.58 | 3.20 × 10−4 | ↓ |
| Acetone | 3.31 ± 1.42 | 2.56 ± 0.85 | 1.45 × 10−2 | ↓ |
| Alanine | 66.34 ± 10.37 | 104.87 ± 11.85 | 4.87 × 10−8 | ↑ |
| Asparagine | 10.36 ± 1.93 | 7.44 ± 1.82 | 7.92 × 10−5 | ↓ |
| Aspartate | 2.17 ± 1.17 | 1.53 ± 0.61 | 2.23 × 10−2 | ↓ |
| Citrate | 11.24 ± 5.88 | 16.40 ± 7.51 | 1.79 × 10−2 | ↑ |
| Creatine | 22.76 ± 7.67 | 17.94 ± 5.91 | 2.95 × 10−4 | ↓ |
| Dimethyl sulfone | 14.74 ± 8.56 | 11.09 ± 5.96 | 4.32 × 10−3 | ↓ |
| Dimethylglycine | 0.36 ± 0.13 | 0.28 ± 0.09 | 3.91 × 10−3 | ↓ |
| Fumarate | 0.84 ± 0.21 | 1.50 ± 0.73 | 8.82 × 10−3 | ↑ |
| Glutamine | 71.51 ± 11.48 | 58.14 ± 6.78 | 6.18 × 10−5 | ↓ |
| Glycerol | 4.03 ± 3.01 | 45.51 ± 22.46 | 1.84 × 10−5 | ↑ |
| Histidine | 29.91 ± 4.34 | 23.46 ± 3.35 | 1.52 × 10−3 | ↓ |
| Lactate | 141.26 ± 50.20 | 1143.45 ± 582.63 | 1.03 × 10−4 | ↑ |
| Leucine | 35.68 ± 6.37 | 39.22 ± 6.87 | 9.62 × 10−3 | ↑ |
| Mannose | 11.20 ± 2.71 | 7.85 ± 2.09 | 1.33 × 10−5 | ↓ |
| Methanol | 16.98 ± 10.21 | 10.26 ± 8.25 | 1.49 × 10−3 | ↓ |
| myo-Inositol | 10.35 ± 1.68 | 7.37 ± 2.06 | 8.38 × 10−4 | ↓ |
| Proline | 10.57 ± 2.45 | 8.02 ± 1.54 | 4.19 × 10−4 | ↓ |
| Pyruvate | 8.72 ± 2.37 | 26.61 ± 7.59 | 4.38 × 10−6 | ↑ |
| Succinate | 1.12 ± 0.21 | 4.24 ± 1.79 | 6.11 × 10−5 | ↑ |
| Threonine | 37.30 ± 5.45 | 33.26 ± 6.98 | 4.93 × 10−3 | ↓ |
| Trimethylamine | 0.16 ± 0.05 | 0.14 ± 0.05 | 2.73 × 10−3 | ↓ |
| Valine | 54.09 ± 6.79 | 50.40 ± 9.04 | 1.85 × 10−2 | ↓ |
| Molecule | T0 | T1 | p | Trend * |
|---|---|---|---|---|
| 4-Aminobutyrate | 30.09 ± 15.39 | 10.87 ± 16.55 | 4.35 × 10−3 | ↓ |
| Betaine | 143.58 ± 86.06 | 67.19 ± 52.51 | 1.23 × 10−2 | ↓ |
| Creatine | 36.68 ± 19.11 | 62.11 ± 26.59 | 1.28 × 10−2 | ↑ |
| Fumarate | 94.70 ± 58.46 | 23.20 ± 29.32 | 8.35 × 10−4 | ↓ |
| Galactose | 21.68 ± 10.18 | 12.55 ± 12.66 | 1.98 × 10−2 | ↓ |
| Malate | 550.72 ± 395.87 | 138.69 ± 166.50 | 2.11 × 10−3 | ↓ |
| Malonate | 193.93 ± 107.17 | 46.84 ± 35.06 | 5.10 × 10−4 | ↓ |
| Methanol | 131.26 ± 73.70 | 73.05 ± 50.01 | 5.07 × 10−3 | ↓ |
| Ornithine | 22.36 ± 12.53 | 46.91 ± 21.66 | 3.24 × 10−3 | ↑ |
| Phenylalanine | 28.22 ± 11.81 | 53.82 ± 23.36 | 6.10 × 10−3 | ↑ |
| Pyruvate | 70.77 ± 32.43 | 49.34 ± 24.69 | 7.41 × 10−2 | ↓ |
| Sarcosine | 2.65 ± 1.64 | 4.48 ± 2.94 | 2.48 × 10−2 | ↑ |
| Succinate | 210.91 ± 78.35 | 120.47 ± 80.66 | 3.59 × 10−2 | ↓ |
| Tyrosine | 33.32 ± 18.13 | 66.07 ± 46.48 | 3.29 × 10−2 | ↑ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazzano, M.; Laghi, L.; Zhu, C.; Lotito, E.; Sgariglia, S.; Tesei, B.; Laus, F. Exercise Induced Changes in Salivary and Serum Metabolome in Trained Standardbred, Assessed by 1H-NMR. Metabolites 2020, 10, 298. https://doi.org/10.3390/metabo10070298
Bazzano M, Laghi L, Zhu C, Lotito E, Sgariglia S, Tesei B, Laus F. Exercise Induced Changes in Salivary and Serum Metabolome in Trained Standardbred, Assessed by 1H-NMR. Metabolites. 2020; 10(7):298. https://doi.org/10.3390/metabo10070298
Chicago/Turabian StyleBazzano, Marilena, Luca Laghi, Chenglin Zhu, Enrica Lotito, Stefano Sgariglia, Beniamino Tesei, and Fulvio Laus. 2020. "Exercise Induced Changes in Salivary and Serum Metabolome in Trained Standardbred, Assessed by 1H-NMR" Metabolites 10, no. 7: 298. https://doi.org/10.3390/metabo10070298
APA StyleBazzano, M., Laghi, L., Zhu, C., Lotito, E., Sgariglia, S., Tesei, B., & Laus, F. (2020). Exercise Induced Changes in Salivary and Serum Metabolome in Trained Standardbred, Assessed by 1H-NMR. Metabolites, 10(7), 298. https://doi.org/10.3390/metabo10070298