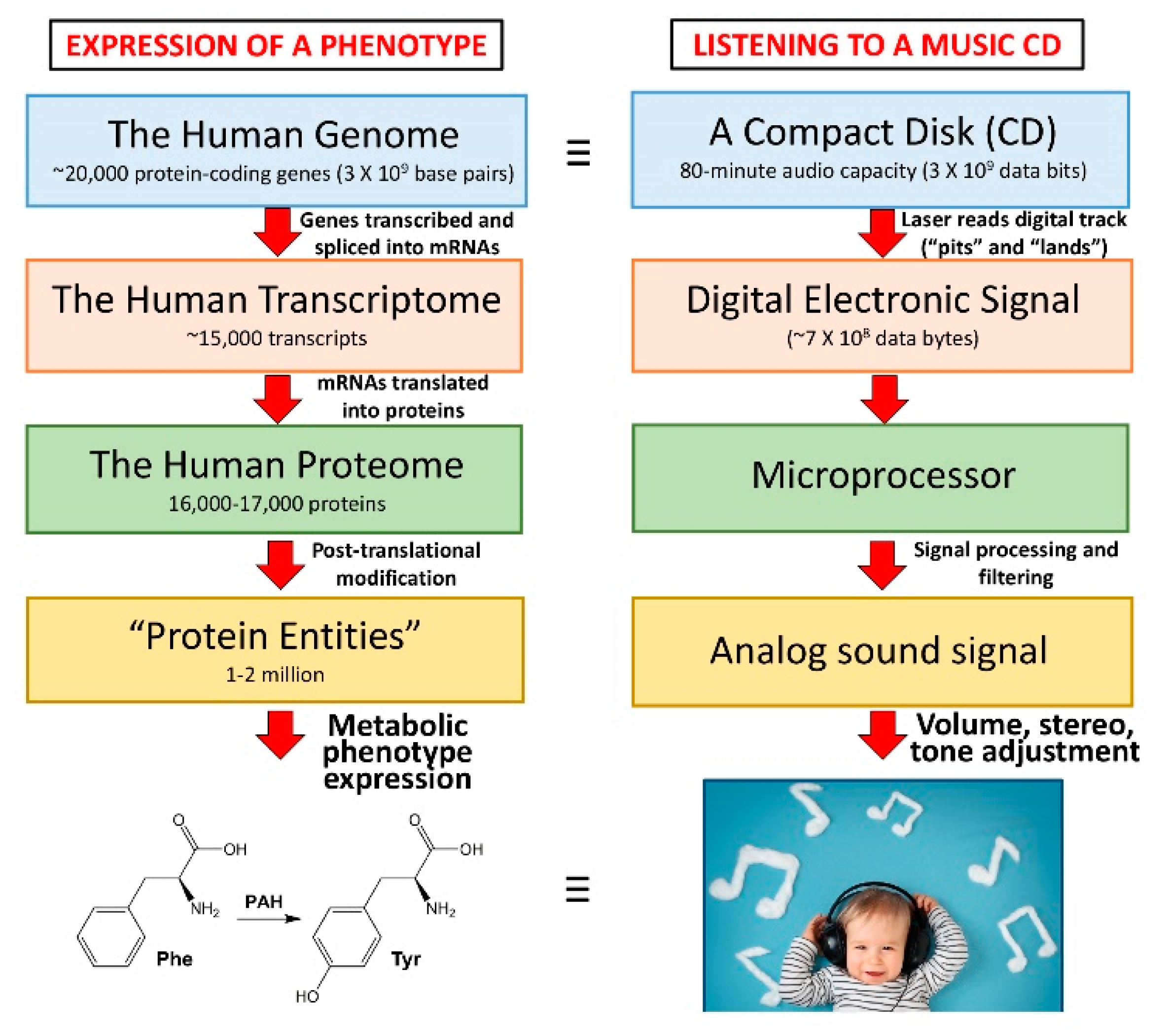
Metabolomic and Lipidomic Biomarkers for Premalignant Liver Disease Diagnosis and Therapy


Abstract
1. The Need for Biomarkers of Premalignant Liver Disease
2. Hepatic Metabolism
3. Metabolomics—The What, the How, and the Why
4. Biomarkers—The Good, the Bad, and the Ugly
5. Biomarkers of Premalignant Liver Disease
5.1. Alcoholic Liver Disease (ALD)
5.2. Cholestasis
5.3. Fibrosis and Cirrhosis
5.4. NAFL and NASH
Overall Summary
Author Contributions
Funding
Conflicts of Interest
References
- Massarweh, N.N.; El-Serag, H.B. Epidemiology of Hepatocellular Carcinoma and Intrahepatic Cholangiocarcinoma. Cancer Control 2017, 24, 1073274817729245. [Google Scholar] [CrossRef]
- Severi, T.; van Malenstein, H.; Verslype, C.; van Pelt, J.F. Tumor initiation and progression in hepatocellular carcinoma: Risk factors, classification, and therapeutic targets. Acta Pharmacol. Sin. 2010, 31, 1409–1420. [Google Scholar] [CrossRef]
- Altekruse, S.F.; Devesa, S.S.; Dickie, L.A.; McGlynn, K.A.; Kleiner, D.E. Histological classification of liver and intrahepatic bile duct cancers in SEER registries. J. Registry Manag. 2011, 38, 201–205. [Google Scholar]
- Baecker, A.; Liu, X.; La Vecchia, C.; Zhang, Z.F. Worldwide incidence of hepatocellular carcinoma cases attributable to major risk factors. Eur. J. Cancer Prev. 2018, 27, 205–212. [Google Scholar] [CrossRef]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Ben-Moshe, S.; Itzkovitz, S. Spatial heterogeneity in the mammalian liver. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 395–410. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Goossens, N.; Guo, J.; Tsai, M.C.; Chou, H.I.; Altunkaynak, C.; Sangiovanni, A.; Iavarone, M.; Colombo, M.; Kobayashi, M.; et al. A hepatic stellate cell gene expression signature associated with outcomes in hepatitis C cirrhosis and hepatocellular carcinoma after curative resection. Gut 2016, 65, 1754–1764. [Google Scholar] [CrossRef]
- Verhulst, S.; Roskams, T.; Sancho-Bru, P.; van Grunsven, L.A. Meta-Analysis of Human and Mouse Biliary Epithelial Cell Gene Profiles. Cells 2019, 8, 1117. [Google Scholar] [CrossRef]
- Aguilar-Bravo, B.; Sancho-Bru, P. Laser capture microdissection: Techniques and applications in liver diseases. Hepatol. Int. 2019, 13, 138–147. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. ‘Metabonomics’: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef]
- Idle, J.R.; Gonzalez, F.J. Metabolomics. Cell Metab. 2007, 6, 348–351. [Google Scholar] [CrossRef]
- Beyoglu, D.; Zhou, Y.; Chen, C.; Idle, J.R. Mass isotopomer-guided decluttering of metabolomic data to visualize endogenous biomarkers of drug toxicity. Biochem. Pharmacol. 2018, 156, 491–500. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Lindon, J.C. Systems biology: Metabonomics. Nature 2008, 455, 1054–1056. [Google Scholar] [CrossRef]
- Gika, H.; Virgiliou, C.; Theodoridis, G.; Plumb, R.S.; Wilson, I.D. Untargeted LC/MS-based metabolic phenotyping (metabonomics/metabolomics): The state of the art. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1117, 136–147. [Google Scholar] [CrossRef]
- Roy, C.; Tremblay, P.Y.; Bienvenu, J.F.; Ayotte, P. Quantitative analysis of amino acids and acylcarnitines combined with untargeted metabolomics using ultra-high performance liquid chromatography and quadrupole time-of-flight mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1027, 40–49. [Google Scholar] [CrossRef]
- Sinclair, K.; Dudley, E. Metabolomics and Biomarker Discovery. Adv. Exp. Med. Biol. 2019, 1140, 613–633. [Google Scholar] [CrossRef]
- Bartel, J.; Krumsiek, J.; Theis, F.J. Statistical methods for the analysis of high-throughput metabolomics data. Comput. Struct. Biotechnol. J. 2013, 4, e201301009. [Google Scholar] [CrossRef]
- Considine, E.C.; Thomas, G.; Boulesteix, A.L.; Khashan, A.S.; Kenny, L.C. Critical review of reporting of the data analysis step in metabolomics. Metabolomics 2017, 14, 7. [Google Scholar] [CrossRef]
- Rosato, A.; Tenori, L.; Cascante, M.; De Atauri Carulla, P.R.; Martins Dos Santos, V.A.P.; Saccenti, E. From correlation to causation: Analysis of metabolomics data using systems biology approaches. Metabolomics 2018, 14, 37. [Google Scholar] [CrossRef]
- Wishart, D.S.; Lewis, M.J.; Morrissey, J.A.; Flegel, M.D.; Jeroncic, K.; Xiong, Y.; Cheng, D.; Eisner, R.; Gautam, B.; Tzur, D.; et al. The human cerebrospinal fluid metabolome. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2008, 871, 164–173. [Google Scholar] [CrossRef]
- Psychogios, N.; Hau, D.D.; Peng, J.; Guo, A.C.; Mandal, R.; Bouatra, S.; Sinelnikov, I.; Krishnamurthy, R.; Eisner, R.; Gautam, B.; et al. The human serum metabolome. PLoS ONE 2011, 6, e16957. [Google Scholar] [CrossRef] [PubMed]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Bjorndahl, T.C.; Krishnamurthy, R.; Saleem, F.; Liu, P.; et al. The human urine metabolome. PLoS ONE 2013, 8, e73076. [Google Scholar] [CrossRef] [PubMed]
- Karu, N.; Deng, L.; Slae, M.; Guo, A.C.; Sajed, T.; Huynh, H.; Wine, E.; Wishart, D.S. A review on human fecal metabolomics: Methods, applications and the human fecal metabolome database. Anal. Chim. Acta 2018, 1030, 1–24. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vazquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Scalbert, A.; Brennan, L.; Manach, C.; Andres-Lacueva, C.; Dragsted, L.O.; Draper, J.; Rappaport, S.M.; van der Hooft, J.J.; Wishart, D.S. The food metabolome: A window over dietary exposure. Am. J. Clin. Nutr. 2014, 99, 1286–1308. [Google Scholar] [CrossRef]
- Lydic, T.A.; Goo, Y.H. Lipidomics unveils the complexity of the lipidome in metabolic diseases. Clin. Transl. Med. 2018, 7, 4. [Google Scholar] [CrossRef]
- Garcia-Ortega, L.F.; Martinez, O. How Many Genes Are Expressed in a Transcriptome? Estimation and Results for RNA-Seq. PLoS ONE 2015, 10, e0130262. [Google Scholar] [CrossRef]
- Wilhelm, M.; Schlegl, J.; Hahne, H.; Gholami, A.M.; Lieberenz, M.; Savitski, M.M.; Ziegler, E.; Butzmann, L.; Gessulat, S.; Marx, H.; et al. Mass-spectrometry-based draft of the human proteome. Nature 2014, 509, 582–587. [Google Scholar] [CrossRef]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Aslebagh, R.; Wormwood, K.L.; Channaveerappa, D.; Wetie, A.G.N.; Woods, A.G.; Darie, C.C. Identification of Posttranslational Modifications (PTMs) of Proteins by Mass Spectrometry. Adv. Exp. Med. Biol. 2019, 1140, 199–224. [Google Scholar] [CrossRef]
- Menetski, J.P.; Hoffmann, S.C.; Cush, S.S.; Kamphaus, T.N.; Austin, C.P.; Herrling, P.L.; Wagner, J.A. The Foundation for the National Institutes of Health Biomarkers Consortium: Past Accomplishments and New Strategic Direction. Clin. Pharmacol. Ther. 2019, 105, 829–843. [Google Scholar] [CrossRef] [PubMed]
- Booth, J. A short history of blood pressure measurement. Proc. R. Soc. Med. 1977, 70, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Dobson, M. Nature of the urine in diabetes. Med. Obs. Inqu. 1776, 5, 298–310. [Google Scholar]
- Garrod, A.E. Inborn Errors of Metabolism; Henry Frowde and Hodder & Stoughton: London, UK, 1909. [Google Scholar]
- Garrod, A.E. The incidence of alkaptonuria: A study in chemical individuality. Lancet 1902, ii, 1616–1620. [Google Scholar] [CrossRef]
- Piro, A.; Tagarelli, G.; Lagonia, P.; Quattrone, A.; Tagarelli, A. Archibald Edward Garrod and alcaptonuria: “Inborn errors of metabolism” revisited. Genet. Med. 2010, 12, 475–476. [Google Scholar] [CrossRef]
- Perlman, R.L.; Govindaraju, D.R. Archibald E. Garrod: The father of precision medicine. Genet. Med. 2016, 18, 1088–1089. [Google Scholar] [CrossRef]
- Phornphutkul, C.; Introne, W.J.; Perry, M.B.; Bernardini, I.; Murphey, M.D.; Fitzpatrick, D.L.; Anderson, P.D.; Huizing, M.; Anikster, Y.; Gerber, L.H.; et al. Natural history of alkaptonuria. N. Engl. J. Med. 2002, 347, 2111–2121. [Google Scholar] [CrossRef]
- Ranganath, L.R.; Khedr, M.; Milan, A.M.; Davison, A.S.; Hughes, A.T.; Usher, J.L.; Taylor, S.; Loftus, N.; Daroszewska, A.; West, E.; et al. Nitisinone arrests ochronosis and decreases rate of progression of Alkaptonuria: Evaluation of the effect of nitisinone in the United Kingdom National Alkaptonuria Centre. Mol. Genet. Metab. 2018, 125, 127–134. [Google Scholar] [CrossRef]
- Adinolfi, M. Embryonic antigens. In Encyclopedia of Immunology, 2nd ed.; Delves, P.J., Ed.; Academic Press: Cambridge, MA, USA, 1998; pp. 798–802. [Google Scholar]
- Abelev, G.I.; Perova, S.D.; Khramkova, N.I.; Postnikova, Z.A.; Irlin, I.S. Production of embryonal alpha-globulin by transplantable mouse hepatomas. Transplantation 1963, 1, 174–180. [Google Scholar] [CrossRef]
- Chan, S.L.; Chan, A.W.H.; Yu, S.C.H. Alpha-Fetoprotein as a Biomarker in Hepatocellular Carcinoma: Focus on Its Role in Composition of Tumor Staging Systems and Monitoring of Treatment Response. In Biomarkers in Disease: Methods, Discoveries and Applications; Patel, V.B., Preedy, V.R., Eds.; Springer Science+Business Media: Dordrecht, The Netherlands, 2017; pp. 623–635. [Google Scholar] [CrossRef]
- Wu, M.; Liu, H.; Liu, Z.; Liu, C.; Zhang, A.; Li, N. Analysis of serum alpha-fetoprotein (AFP) and AFP-L3 levels by protein microarray. J. Int. Med. Res. 2018, 46, 4297–4305. [Google Scholar] [CrossRef]
- Derosa, G.; Maffioli, P. Traditional markers in liver disease. In Biomarkers in Disease: Methods, Discoveries and Applications; Patel, V.B., Preedy, V.R., Eds.; Springer Science+Business Media: Dordrecht, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Lou, J.; Zhang, L.; Lv, S.; Zhang, C.; Jiang, S. Biomarkers for Hepatocellular Carcinoma. Biomark. Cancer 2017, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bruha, R. Osteopontin as a biomarker in liver disease. In Biomarkers in Liver Disease: Methods, Discoveries and Applications; Patel, V.B., Preedy, V.R., Eds.; Springer Science+Business Media: Dordrecht, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Crandall, D.I.; Halikis, D.N. Homogentisic acid oxidase. I. Distribution in animal tissue and relation to tyrosine metabolism in rat kidney. J. Biol. Chem. 1954, 208, 629–638. [Google Scholar]
- Bernardini, G.; Laschi, M.; Geminiani, M.; Braconi, D.; Vannuccini, E.; Lupetti, P.; Manetti, F.; Millucci, L.; Santucci, A. Homogentisate 1,2 dioxygenase is expressed in brain: Implications in alkaptonuria. J. Inherit. Metab. Dis. 2015, 38, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, P.; Seebauer, C.T.; Schnabl, B. Alcoholic liver disease: The gut microbiome and liver cross talk. Alcohol. Clin. Exp. Res. 2015, 39, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Osna, N.A.; Donohue, T.M., Jr.; Kharbanda, K.K. Alcoholic Liver Disease: Pathogenesis and Current Management. Alcohol. Res. 2017, 38, 147–161. [Google Scholar] [PubMed]
- Lieber, C.S.; Teschke, R.; Hasumura, Y.; Decarli, L.M. Differences in hepatic and metabolic changes after acute and chronic alcohol consumption. Fed. Proc. 1975, 34, 2060–2074. [Google Scholar]
- You, M.; Arteel, G.E. Effect of ethanol on lipid metabolism. J. Hepatol. 2019, 70, 237–248. [Google Scholar] [CrossRef]
- Lieber, C.S.; Jones, D.P.; Decarli, L.M. Effects of Prolonged Ethanol Intake: Production of Fatty Liver Despite Adequate Diets. J. Clin. Investig. 1965, 44, 1009–1021. [Google Scholar] [CrossRef]
- Lieber, C.S.; DeCarli, L.M. Liquid diet technique of ethanol administration: 1989 Update. Alcohol Alcohol. 1989, 24, 197–211. [Google Scholar]
- Guo, F.; Zheng, K.; Benede-Ubieto, R.; Cubero, F.J.; Nevzorova, Y.A. The Lieber-DeCarli Diet-A Flagship Model for Experimental Alcoholic Liver Disease. Alcohol Clin. Exp. Res. 2018, 42, 1828–1840. [Google Scholar] [CrossRef]
- Kim, S.J.; Jung, Y.S.; Kwon, D.Y.; Kim, Y.C. Alleviation of acute ethanol-induced liver injury and impaired metabolomics of S-containing substances by betaine supplementation. Biochem. Biophys. Res. Commun. 2008, 368, 893–898. [Google Scholar] [CrossRef] [PubMed]
- Zivkovic, A.M.; Bruce German, J.; Esfandiari, F.; Halsted, C.H. Quantitative lipid metabolomic changes in alcoholic micropigs with fatty liver disease. Alcohol Clin. Exp. Res. 2009, 33, 751–758. [Google Scholar] [CrossRef]
- Manna, S.K.; Patterson, A.D.; Yang, Q.; Krausz, K.W.; Li, H.; Idle, J.R.; Fornace, A.J., Jr.; Gonzalez, F.J. Identification of noninvasive biomarkers for alcohol-induced liver disease using urinary metabolomics and the Ppara-null mouse. J. Proteome Res. 2010, 9, 4176–4188. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.K.; Patterson, A.D.; Yang, Q.; Krausz, K.W.; Idle, J.R.; Fornace, A.J.; Gonzalez, F.J. UPLC-MS-based urine metabolomics reveals indole-3-lactic acid and phenyllactic acid as conserved biomarkers for alcohol-induced liver disease in the Ppara-null mouse model. J. Proteome Res. 2011, 10, 4120–4133. [Google Scholar] [CrossRef]
- Fernando, H.; Bhopale, K.K.; Kondraganti, S.; Kaphalia, B.S.; Shakeel Ansari, G.A. Lipidomic changes in rat liver after long-term exposure to ethanol. Toxicol. Appl. Pharmacol. 2011, 255, 127–137. [Google Scholar] [CrossRef]
- Suciu, A.M.; Crisan, D.A.; Procopet, B.D.; Radu, C.I.; Socaciu, C.; Tantau, M.V.; Stefanescu, H.O.; Grigorescu, M. What’s in Metabolomics for Alcoholic Liver Disease? J. Gastrointestin. Liver Dis. 2018, 27, 51–58. [Google Scholar] [CrossRef]
- Shi, X.; Yao, D.; Chen, C. Identification of N-acetyltaurine as a novel metabolite of ethanol through metabolomics-guided biochemical analysis. J. Biol. Chem. 2012, 287, 6336–6349. [Google Scholar] [CrossRef]
- Johnson, C.H.; Patterson, A.D.; Krausz, K.W.; Lanz, C.; Kang, D.W.; Luecke, H.; Gonzalez, F.J.; Idle, J.R. Radiation metabolomics. 4. UPLC-ESI-QTOFMS-Based metabolomics for urinary biomarker discovery in gamma-irradiated rats. Radiat. Res. 2011, 175, 473–484. [Google Scholar] [CrossRef]
- Johnson, C.H.; Patterson, A.D.; Krausz, K.W.; Kalinich, J.F.; Tyburski, J.B.; Kang, D.W.; Luecke, H.; Gonzalez, F.J.; Blakely, W.F.; Idle, J.R. Radiation metabolomics. 5. Identification of urinary biomarkers of ionizing radiation exposure in nonhuman primates by mass spectrometry-based metabolomics. Radiat. Res. 2012, 178, 328–340. [Google Scholar] [CrossRef]
- Luginbuhl, M.; Rutjens, S.; Konig, S.; Furrer, J.; Weinmann, W. N-Acetyltaurine as a novel urinary ethanol marker in a drinking study. Anal. Bioanal. Chem. 2016, 408, 7529–7536. [Google Scholar] [CrossRef]
- Luginbuhl, M.; Konig, S.; Schurch, S.; Weinmann, W. Evaluation of N-acetyltaurine as an ethanol marker in human blood. Alcohol 2017, 65, 11–18. [Google Scholar] [CrossRef]
- Xie, G.; Zhong, W.; Li, H.; Li, Q.; Qiu, Y.; Zheng, X.; Chen, H.; Zhao, X.; Zhang, S.; Zhou, Z.; et al. Alteration of bile acid metabolism in the rat induced by chronic ethanol consumption. FASEB J. 2013, 27, 3583–3593. [Google Scholar] [CrossRef]
- Xie, G.; Zhong, W.; Zheng, X.; Li, Q.; Qiu, Y.; Li, H.; Chen, H.; Zhou, Z.; Jia, W. Chronic ethanol consumption alters mammalian gastrointestinal content metabolites. J. Proteome Res. 2013, 12, 3297–3306. [Google Scholar] [CrossRef]
- Chen, P.; Torralba, M.; Tan, J.; Embree, M.; Zengler, K.; Starkel, P.; van Pijkeren, J.P.; DePew, J.; Loomba, R.; Ho, S.B.; et al. Supplementation of saturated long-chain fatty acids maintains intestinal eubiosis and reduces ethanol-induced liver injury in mice. Gastroenterology 2015, 148, 203–214. [Google Scholar] [CrossRef]
- Fang, H.; Zhang, A.H.; Sun, H.; Yu, J.B.; Wang, L.; Wang, X.J. High-throughput metabolomics screen coupled with multivariate statistical analysis identifies therapeutic targets in alcoholic liver disease rats using liquid chromatography-mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1109, 112–120. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, A.; Qiu, S.; Sun, H.; Guan, Y.; Wang, X. High-throughput metabolomics approach reveals new mechanistic insights for drug response of phenotypes of geniposide towards alcohol-induced liver injury by using liquid chromatography coupled to high resolution mass spectrometry. Mol. Biosyst. 2017, 13, 73–82. [Google Scholar] [CrossRef]
- Deda, O.; Virgiliou, C.; Orfanidis, A.; Gika, H.G. Study of Fecal and Urinary Metabolite Perturbations Induced by Chronic Ethanol Treatment in Mice by UHPLC-MS/MS Targeted Profiling. Metabolites 2019, 9, 232. [Google Scholar] [CrossRef]
- He, L.; Li, F.; Yin, X.; Bohman, P.; Kim, S.; McClain, C.J.; Feng, W.; Zhang, X. Profiling of Polar Metabolites in Mouse Feces Using Four Analytical Platforms to Study the Effects Of Cathelicidin-Related Antimicrobial Peptide in Alcoholic Liver Disease. J. Proteome Res. 2019, 18, 2875–2884. [Google Scholar] [CrossRef]
- Gao, B.; Lang, S.; Duan, Y.; Wang, Y.; Shawcross, D.L.; Louvet, A.; Mathurin, P.; Ho, S.B.; Starkel, P.; Schnabl, B. Serum and Fecal Oxylipins in Patients with Alcohol-Related Liver Disease. Dig. Dis. Sci. 2019, 64, 1878–1892. [Google Scholar] [CrossRef]
- Michelena, J.; Alonso, C.; Martinez-Arranz, I.; Altamirano, J.; Mayo, R.; Sancho-Bru, P.; Bataller, R.; Gines, P.; Castro, A.; Caballeria, J. Metabolomics Discloses a New Non-invasive Method for the Diagnosis and Prognosis of Patients with Alcoholic Hepatitis. Ann. Hepatol. 2019, 18, 144–154. [Google Scholar] [CrossRef]
- Keitel, V.; Droge, C.; Haussinger, D. Targeting FXR in Cholestasis. Handb. Exp. Pharmacol. 2019, 256, 299–324. [Google Scholar] [CrossRef]
- Ishihara, K.; Katsutani, N.; Asai, N.; Inomata, A.; Uemura, Y.; Suganuma, A.; Sawada, K.; Yokoi, T.; Aoki, T. Identification of urinary biomarkers useful for distinguishing a difference in mechanism of toxicity in rat model of cholestasis. Basic Clin. Pharmacol. Toxicol. 2009, 105, 156–166. [Google Scholar] [CrossRef]
- Cho, J.Y.; Matsubara, T.; Kang, D.W.; Ahn, S.H.; Krausz, K.W.; Idle, J.R.; Luecke, H.; Gonzalez, F.J. Urinary metabolomics in Fxr-null mice reveals activated adaptive metabolic pathways upon bile acid challenge. J. Lipid Res. 2010, 51, 1063–1074. [Google Scholar] [CrossRef]
- Passmore, I.J.; Letertre, M.P.M.; Preston, M.D.; Bianconi, I.; Harrison, M.A.; Nasher, F.; Kaur, H.; Hong, H.A.; Baines, S.D.; Cutting, S.M.; et al. Para-cresol production by Clostridium difficile affects microbial diversity and membrane integrity of Gram-negative bacteria. PLoS Pathog 2018, 14, e1007191. [Google Scholar] [CrossRef]
- Matsubara, T.; Tanaka, N.; Sato, M.; Kang, D.W.; Krausz, K.W.; Flanders, K.C.; Ikeda, K.; Luecke, H.; Wakefield, L.M.; Gonzalez, F.J. TGF-beta-SMAD3 signaling mediates hepatic bile acid and phospholipid metabolism following lithocholic acid-induced liver injury. J. Lipid Res. 2012, 53, 2698–2707. [Google Scholar] [CrossRef]
- Aoki, M.; Konya, Y.; Takagaki, T.; Umemura, K.; Sogame, Y.; Katsumata, T.; Komuro, S. Metabolomic investigation of cholestasis in a rat model using ultra-performance liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2011, 25, 1847–1852. [Google Scholar] [CrossRef]
- Yamazaki, M.; Miyake, M.; Sato, H.; Masutomi, N.; Tsutsui, N.; Adam, K.P.; Alexander, D.C.; Lawton, K.A.; Milburn, M.V.; Ryals, J.A.; et al. Perturbation of bile acid homeostasis is an early pathogenesis event of drug induced liver injury in rats. Toxicol. Appl. Pharmacol. 2013, 268, 79–89. [Google Scholar] [CrossRef]
- Chen, Z.; Zhu, Y.; Zhao, Y.; Ma, X.; Niu, M.; Wang, J.; Su, H.; Wang, R.; Li, J.; Liu, L.; et al. Serum Metabolomic Profiling in a Rat Model Reveals Protective Function of Paeoniflorin Against ANIT Induced Cholestasis. Phytother. Res. 2016, 30, 654–662. [Google Scholar] [CrossRef]
- Ma, X.; Chi, Y.H.; Niu, M.; Zhu, Y.; Zhao, Y.L.; Chen, Z.; Wang, J.B.; Zhang, C.E.; Li, J.Y.; Wang, L.F.; et al. Metabolomics Coupled with Multivariate Data and Pathway Analysis on Potential Biomarkers in Cholestasis and Intervention Effect of Paeonia lactiflora Pall. Front. Pharmacol. 2016, 7, 14. [Google Scholar] [CrossRef]
- Zhang, C.E.; Niu, M.; Li, R.Y.; Feng, W.W.; Ma, X.; Dong, Q.; Ma, Z.J.; Li, G.Q.; Meng, Y.K.; Wang, Y.; et al. Untargeted Metabolomics Reveals Dose-Response Characteristics for Effect of Rhubarb in a Rat Model of Cholestasis. Front. Pharmacol. 2016, 7, 85. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, A.H.; Zou, D.X.; Sun, W.J.; Wu, X.H.; Wang, X.J. Metabolomics coupled with pattern recognition and pathway analysis on potential biomarkers in liver injury and hepatoprotective effects of yinchenhao. Appl. Biochem. Biotechnol. 2014, 173, 857–869. [Google Scholar] [CrossRef]
- Li, Y.F.; Wu, J.S.; Li, Y.Y.; Dai, Y.; Zheng, M.; Zeng, J.K.; Wang, G.F.; Wang, T.M.; Li, W.K.; Zhang, X.Y.; et al. Chicken bile powder protects against alpha-naphthylisothiocyanate-induced cholestatic liver injury in mice. Oncotarget 2017, 8, 97137–97152. [Google Scholar] [CrossRef][Green Version]
- Wu, J.S.; Li, Y.F.; Li, Y.Y.; Dai, Y.; Li, W.K.; Zheng, M.; Shi, Z.C.; Shi, R.; Wang, T.M.; Ma, B.L.; et al. Huangqi Decoction Alleviates Alpha-Naphthylisothiocyanate Induced Intrahepatic Cholestasis by Reversing Disordered Bile Acid and Glutathione Homeostasis in Mice. Front. Pharmacol. 2017, 8, 938. [Google Scholar] [CrossRef]
- Han, H.; Xu, L.; Xiong, K.; Zhang, T.; Wang, Z. Exploration of Hepatoprotective Effect of Gentiopicroside on Alpha-Naphthylisothiocyanate-Induced Cholestatic Liver Injury in Rats by Comprehensive Proteomic and Metabolomic Signatures. Cell Physiol. Biochem. 2018, 49, 1304–1319. [Google Scholar] [CrossRef]
- Zhu, G.; Feng, F. UPLC-MS-based metabonomic analysis of intervention effects of Da-Huang-Xiao-Shi decoction on ANIT-induced cholestasis. J. Ethnopharmacol. 2019, 238, 111860. [Google Scholar] [CrossRef]
- Ma, X.; Idle, J.R.; Krausz, K.W.; Tan, D.X.; Ceraulo, L.; Gonzalez, F.J. Urinary metabolites and antioxidant products of exogenous melatonin in the mouse. J. Pineal Res. 2006, 40, 343–349. [Google Scholar] [CrossRef]
- Ma, X.; Chen, C.; Krausz, K.W.; Idle, J.R.; Gonzalez, F.J. A metabolomic perspective of melatonin metabolism in the mouse. Endocrinology 2008, 149, 1869–1879. [Google Scholar] [CrossRef]
- Yu, H.; Li, Y.; Xu, Z.; Wang, D.; Shi, S.; Deng, H.; Zeng, B.; Zheng, Z.; Sun, L.; Deng, X.; et al. Identification of potential biomarkers in cholestasis and the therapeutic effect of melatonin by metabolomics, multivariate data and pathway analyses. Int J. Mol. Med. 2018, 42, 2515–2526. [Google Scholar] [CrossRef]
- Lin, S.; Wang, T.Y.; Xu, H.R.; Zhang, X.N.; Wang, Q.; Liu, R.; Li, Q.; Bi, K.S. A systemic combined nontargeted and targeted LC-MS based metabolomic strategy of plasma and liver on pathology exploration of alpha-naphthylisothiocyanate induced cholestatic liver injury in mice. J. Pharm. Biomed. Anal. 2019, 171, 180–192. [Google Scholar] [CrossRef]
- Dai, M.; Hua, H.; Lin, H.; Xu, G.; Hu, X.; Li, F.; Gonzalez, F.J.; Liu, A.; Yang, J. Targeted Metabolomics Reveals a Protective Role for Basal PPARalpha in Cholestasis Induced by alpha-Naphthylisothiocyanate. J. Proteome Res. 2018, 17, 1500–1508. [Google Scholar] [CrossRef]
- Wang, B.L.; Zhang, C.W.; Wang, L.; Tang, K.L.; Tanaka, N.; Gonzalez, F.J.; Xu, Y.; Fang, Z.Z. Lipidomics reveal aryl hydrocarbon receptor (Ahr)-regulated lipid metabolic pathway in alpha-naphthyl isothiocyanate (ANIT)-induced intrahepatic cholestasis. Xenobiotica 2019, 49, 591–601. [Google Scholar] [CrossRef]
- Fu, K.; Wang, C.; Gao, Y.; Fan, S.; Zhang, H.; Sun, J.; Jiang, Y.; Liu, C.; Guan, L.; Liu, J.; et al. Metabolomics and Lipidomics Reveal the Effect of Hepatic Vps33b Deficiency on Bile Acids and Lipids Metabolism. Front. Pharmacol. 2019, 10, 276. [Google Scholar] [CrossRef]
- Long, Y.; Dong, X.; Yuan, Y.; Huang, J.; Song, J.; Sun, Y.; Lu, Z.; Yang, L.; Yu, W. Metabolomics changes in a rat model of obstructive jaundice: Mapping to metabolism of amino acids, carbohydrates and lipids as well as oxidative stress. J. Clin. Biochem. Nutr. 2015, 57, 50–59. [Google Scholar] [CrossRef]
- Wei, D.D.; Wang, J.S.; Duan, J.A.; Kong, L.Y. Metabolomic Assessment of Acute Cholestatic Injuries Induced by Thioacetamide and by Bile Duct Ligation, and the Protective Effects of Huang-Lian-Jie-Du-Decoction. Front. Pharmacol. 2018, 9, 458. [Google Scholar] [CrossRef]
- Yang, R.; Zhao, Q.; Hu, D.D.; Xiao, X.R.; Huang, J.F.; Li, F. Metabolomic analysis of cholestatic liver damage in mice. Food Chem. Toxicol. 2018, 120, 253–260. [Google Scholar] [CrossRef]
- Lian, J.S.; Liu, W.; Hao, S.R.; Chen, D.Y.; Wang, Y.Y.; Yang, J.L.; Jia, H.Y.; Huang, J.R. A serum metabolomic analysis for diagnosis and biomarker discovery of primary biliary cirrhosis and autoimmune hepatitis. Hepatobiliary Pancreat Dis. Int. 2015, 14, 413–421. [Google Scholar] [CrossRef]
- Trottier, J.; Bialek, A.; Caron, P.; Straka, R.J.; Heathcote, J.; Milkiewicz, P.; Barbier, O. Metabolomic profiling of 17 bile acids in serum from patients with primary biliary cirrhosis and primary sclerosing cholangitis: A pilot study. Dig. Liver Dis. 2012, 44, 303–310. [Google Scholar] [CrossRef]
- Bell, L.N.; Wulff, J.; Comerford, M.; Vuppalanchi, R.; Chalasani, N. Serum metabolic signatures of primary biliary cirrhosis and primary sclerosing cholangitis. Liver Int. 2015, 35, 263–274. [Google Scholar] [CrossRef]
- Tang, Y.M.; Wang, J.P.; Bao, W.M.; Yang, J.H.; Ma, L.K.; Yang, J.; Chen, H.; Xu, Y.; Yang, L.H.; Li, W.; et al. Urine and serum metabolomic profiling reveals that bile acids and carnitine may be potential biomarkers of primary biliary cirrhosis. Int. J. Mol. Med. 2015, 36, 377–385. [Google Scholar] [CrossRef]
- Vignoli, A.; Orlandini, B.; Tenori, L.; Biagini, M.R.; Milani, S.; Renzi, D.; Luchinat, C.; Calabro, A.S. Metabolic Signature of Primary Biliary Cholangitis and Its Comparison with Celiac Disease. J. Proteome Res. 2019, 18, 1228–1236. [Google Scholar] [CrossRef]
- Ozkan, S.; Ceylan, Y.; Ozkan, O.V.; Yildirim, S. Review of a challenging clinical issue: Intrahepatic cholestasis of pregnancy. World J. Gastroenterol. 2015, 21, 7134–7141. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, X.; Pan, F.; Cui, Y.; Yang, T.; Deng, L.; Shao, Y.; Ding, M. Urinary metabolomic analysis of intrahepatic cholestasis of pregnancy based on high performance liquid chromatography/mass spectrometry. Clin. Chim. Acta. 2017, 471, 292–297. [Google Scholar] [CrossRef]
- Cui, Y.; Xu, B.; Zhang, X.; He, Y.; Shao, Y.; Ding, M. Diagnostic and therapeutic profiles of serum bile acids in women with intrahepatic cholestasis of pregnancy-a pseudo-targeted metabolomics study. Clin. Chim. Acta. 2018, 483, 135–141. [Google Scholar] [CrossRef]
- Wang, P.; Zhong, H.; Song, Y.; Yuan, P.; Li, Y.; Lin, S.; Zhang, X.; Li, J.; Che, L.; Feng, B.; et al. Targeted metabolomics analysis of maternal-placental-fetal metabolism in pregnant swine reveals links in fetal bile acid homeostasis and sulfation capacity. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G8–G16. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, X.; Xu, B.; Cui, Y.; He, Y.; Yang, T.; Shao, Y.; Ding, M. The urinary bile acid profiling analysis of asymptomatic hypercholanemia of pregnancy: A pseudo-targeted metabolomics study. Clin. Chim. Acta 2019, 497, 67–75. [Google Scholar] [CrossRef]
- De Seymour, J.V.; Tu, S.; He, X.; Zhang, H.; Han, T.L.; Baker, P.N.; Sulek, K. Metabolomic profiling of maternal hair suggests rapid development of intrahepatic cholestasis of pregnancy. Metabolomics 2018, 14, 79. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Chen, J.; Feng, C.; He, Y.; Shao, Y.; Ding, M. Targeted metabolomics of sulfated bile acids in urine for the diagnosis and grading of intrahepatic cholestasis of pregnancy. Genes Dis. 2018, 5, 358–366. [Google Scholar] [CrossRef]
- Li, W.W.; Yang, Y.; Dai, Q.G.; Lin, L.L.; Xie, T.; He, L.L.; Tao, J.L.; Shan, J.J.; Wang, S.C. Non-invasive urinary metabolomic profiles discriminate biliary atresia from infantile hepatitis syndrome. Metabolomics 2018, 14, 90. [Google Scholar] [CrossRef]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef]
- Goodman, Z.D. Grading and staging systems for inflammation and fibrosis in chronic liver diseases. J. Hepatol. 2007, 47, 598–607. [Google Scholar] [CrossRef]
- D’Amico, G.; Garcia-Tsao, G.; Pagliaro, L. Natural history and prognostic indicators of survival in cirrhosis: A systematic review of 118 studies. J. Hepatol. 2006, 44, 217–231. [Google Scholar] [CrossRef]
- Fleming, K.M.; Aithal, G.P.; Card, T.R.; West, J. All-cause mortality in people with cirrhosis compared with the general population: A population-based cohort study. Liver Int. 2012, 32, 79–84. [Google Scholar] [CrossRef]
- Chang, M.L.; Yang, S.S. Metabolic Signature of Hepatic Fibrosis: From Individual Pathways to Systems Biology. Cells 2019, 8, 1423. [Google Scholar] [CrossRef]
- Tokushige, K.; Hashimoto, E.; Kodama, K.; Tobari, M.; Matsushita, N.; Kogiso, T.; Taniai, M.; Torii, N.; Shiratori, K.; Nishizaki, Y.; et al. Serum metabolomic profile and potential biomarkers for severity of fibrosis in nonalcoholic fatty liver disease. J. Gastroenterol. 2013, 48, 1392–1400. [Google Scholar] [CrossRef]
- Fishman, J.; Schneider, J.; Hershcope, R.J.; Bradlow, H.L. Increased estrogen-16 alpha-hydroxylase activity in women with breast and endometrial cancer. J. Steroid Biochem. 1984, 20, 1077–1081. [Google Scholar] [CrossRef]
- Batista, A.D.; Barros, C.J.P.; Costa, T.; de Godoy, M.M.G.; Silva, R.D.; Santos, J.C.; de Melo Lira, M.M.; Juca, N.T.; Lopes, E.P.A.; Silva, R.O. Proton nuclear magnetic resonance-based metabonomic models for non-invasive diagnosis of liver fibrosis in chronic hepatitis C: Optimizing the classification of intermediate fibrosis. World J. Hepatol. 2018, 10, 105–115. [Google Scholar] [CrossRef]
- Abellona, U.M.R.; Taylor-Robinson, S.D. Comments on Gabbani, et al. Metabolomic analysis with (1)H NMR for non-invasive diagnosis of hepatic fibrosis degree in patients with chronic hepatitis C. Dig. Liver Dis. 2018, 50, 209–210. [Google Scholar] [CrossRef]
- Wu, F.; Zheng, H.; Yang, Z.T.; Cheng, B.; Wu, J.X.; Liu, X.W.; Tang, C.L.; Lu, S.Y.; Chen, Z.N.; Song, F.M.; et al. Urinary metabonomics study of the hepatoprotective effects of total alkaloids from Corydalis saxicola Bunting on carbon tetrachloride-induced chronic hepatotoxicity in rats using (1)H NMR analysis. J. Pharm. Biomed. Anal. 2017, 140, 199–209. [Google Scholar] [CrossRef]
- Liu, X.W.; Tang, C.L.; Zheng, H.; Wu, J.X.; Wu, F.; Mo, Y.Y.; Liu, X.; Zhu, H.J.; Yin, C.L.; Cheng, B.; et al. Investigation of the hepatoprotective effect of Corydalis saxicola Bunting on carbon tetrachloride-induced liver fibrosis in rats by (1)H-NMR-based metabonomics and network pharmacology approaches. J. Pharm. Biomed. Anal. 2018, 159, 252–261. [Google Scholar] [CrossRef]
- Feng, X.; Li, M.H.; Xia, J.; Deng Ba, D.J.; Ruan, L.Y.; Xing, Y.X.; Chen, C.; Wang, J.S.; Zhong, G.J. Tibetan Medical Formula Shi-Wei-Gan-Ning-Pill Protects Against Carbon Tetrachloride-Induced Liver Fibrosis—An NMR-Based Metabolic Profiling. Front. Pharmacol. 2018, 9, 965. [Google Scholar] [CrossRef]
- Li, M.H.; Feng, X.; Deng Ba, D.J.; Chen, C.; Ruan, L.Y.; Xing, Y.X.; Chen, L.Y.; Zhong, G.J.; Wang, J.S. Hepatoprotection of Herpetospermum caudigerum Wall. against CCl4-induced liver fibrosis on rats. J. Ethnopharmacol. 2019, 229, 1–14. [Google Scholar] [CrossRef]
- Yin, P.; Wan, D.; Zhao, C.; Chen, J.; Zhao, X.; Wang, W.; Lu, X.; Yang, S.; Gu, J.; Xu, G. A metabonomic study of hepatitis B-induced liver cirrhosis and hepatocellular carcinoma by using RP-LC and HILIC coupled with mass spectrometry. Mol. Biosyst. 2009, 5, 868–876. [Google Scholar] [CrossRef]
- Xue, R.; Dong, L.; Wu, H.; Liu, T.; Wang, J.; Shen, X. Gas chromatography/mass spectrometry screening of serum metabolomic biomarkers in hepatitis B virus infected cirrhosis patients. Clin. Chem. Lab. Med. 2009, 47, 305–310. [Google Scholar] [CrossRef]
- Waldhier, M.C.; Almstetter, M.F.; Nurnberger, N.; Gruber, M.A.; Dettmer, K.; Oefner, P.J. Improved enantiomer resolution and quantification of free D-amino acids in serum and urine by comprehensive two-dimensional gas chromatography-time-of-flight mass spectrometry. J. Chromatogr. A 2011, 1218, 4537–4544. [Google Scholar] [CrossRef]
- Bastings, J.; van Eijk, H.M.; Olde Damink, S.W.; Rensen, S.S. d-amino Acids in Health and Disease: A Focus on Cancer. Nutrients 2019, 11, 2205. [Google Scholar] [CrossRef]
- Huang, H.J.; Zhang, A.Y.; Cao, H.C.; Lu, H.F.; Wang, B.H.; Xie, Q.; Xu, W.; Li, L.J. Metabolomic analyses of faeces reveals malabsorption in cirrhotic patients. Dig. Liver Dis. 2013, 45, 677–682. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Z.; Huang, M.; Sun, X.; Liu, B.; Guo, Q.; Chang, Q.; Duan, Z. Taurocholic acid is an active promoting factor, not just a biomarker of progression of liver cirrhosis: Evidence from a human metabolomic study and in vitro experiments. BMC Gastroenterol. 2018, 18, 112. [Google Scholar] [CrossRef]
- Patterson, A.D.; Maurhofer, O.; Beyoglu, D.; Lanz, C.; Krausz, K.W.; Pabst, T.; Gonzalez, F.J.; Dufour, J.F.; Idle, J.R. Aberrant lipid metabolism in hepatocellular carcinoma revealed by plasma metabolomics and lipid profiling. Cancer Res. 2011, 71, 6590–6600. [Google Scholar] [CrossRef]
- Osman, D.; Ali, O.; Obada, M.; El-Mezayen, H.; El-Said, H. Chromatographic determination of some biomarkers of liver cirrhosis and hepatocellular carcinoma in Egyptian patients. Biomed. Chromatogr. 2017, 31. [Google Scholar] [CrossRef]
- Wang, B.; Chen, D.; Chen, Y.; Hu, Z.; Cao, M.; Xie, Q.; Chen, Y.; Xu, J.; Zheng, S.; Li, L. Metabonomic profiles discriminate hepatocellular carcinoma from liver cirrhosis by ultraperformance liquid chromatography-mass spectrometry. J. Proteome Res. 2012, 11, 1217–1227. [Google Scholar] [CrossRef]
- Koller, A.; Aldwin, L.; Natelson, S. Hepatic synthesis of canavaninosuccinate from ureidohomoserine and aspartate, and its conversion to guanidinosuccinate. Clin. Chem. 1975, 21, 1777–1782. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Q.; Cao, G.; Chen, H.; Liu, D.; Su, W.; Yu, X.Y.; Vaziri, N.D.; Liu, X.H.; Bai, X.; Zhang, L.; et al. Gene and protein expressions and metabolomics exhibit activated redox signaling and wnt/beta-catenin pathway are associated with metabolite dysfunction in patients with chronic kidney disease. Redox Biol. 2017, 12, 505–521. [Google Scholar] [CrossRef] [PubMed]
- Mindikoglu, A.L.; Opekun, A.R.; Putluri, N.; Devaraj, S.; Sheikh-Hamad, D.; Vierling, J.M.; Goss, J.A.; Rana, A.; Sood, G.K.; Jalal, P.K.; et al. Unique metabolomic signature associated with hepatorenal dysfunction and mortality in cirrhosis. Transl. Res. 2018, 195, 25–47. [Google Scholar] [CrossRef]
- Liu, Y.; Hong, Z.; Tan, G.; Dong, X.; Yang, G.; Zhao, L.; Chen, X.; Zhu, Z.; Lou, Z.; Qian, B.; et al. NMR and LC/MS-based global metabolomics to identify serum biomarkers differentiating hepatocellular carcinoma from liver cirrhosis. Int J. Cancer 2014, 135, 658–668. [Google Scholar] [CrossRef]
- McPhail, M.J.W.; Shawcross, D.L.; Lewis, M.R.; Coltart, I.; Want, E.J.; Antoniades, C.G.; Veselkov, K.; Triantafyllou, E.; Patel, V.; Pop, O.; et al. Multivariate metabotyping of plasma predicts survival in patients with decompensated cirrhosis. J. Hepatol. 2016, 64, 1058–1067. [Google Scholar] [CrossRef]
- Dai, W.; Yin, P.; Chen, P.; Kong, H.; Luo, P.; Xu, Z.; Lu, X.; Xu, G. Study of urinary steroid hormone disorders: Difference between hepatocellular carcinoma in early stage and cirrhosis. Anal. Bioanal. Chem. 2014, 406, 4325–4335. [Google Scholar] [CrossRef]
- Gao, R.; Cheng, J.; Fan, C.; Shi, X.; Cao, Y.; Sun, B.; Ding, H.; Hu, C.; Dong, F.; Yan, X. Serum Metabolomics to Identify the Liver Disease-Specific Biomarkers for the Progression of Hepatitis to Hepatocellular Carcinoma. Sci. Rep. 2015, 5, 18175. [Google Scholar] [CrossRef]
- Hou, Q.; Duan, Z.J. Metabonomic window into hepatitis B virus-related hepatic diseases. World J. Hepatol. 2016, 8, 1–8. [Google Scholar] [CrossRef]
- Lu, Y.; Fang, J.; Zou, L.; Cui, L.; Liang, X.; Lim, S.G.; Dan, Y.Y.; Ong, C.N. Omega-6-derived oxylipin changes in serum of patients with hepatitis B virus-related liver diseases. Metabolomics 2018, 14, 26. [Google Scholar] [CrossRef]
- Cabral, M.; Martin-Venegas, R.; Moreno, J.J. Differential cell growth/apoptosis behavior of 13-hydroxyoctadecadienoic acid enantiomers in a colorectal cancer cell line. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G664–G671. [Google Scholar] [CrossRef]
- Zhao, C.Q.; Chen, L.; Cai, H.; Yao, W.L.; Zhou, Q.; Zhu, H.M.; Gao, Y.; Liu, P.; Gou, X.J.; Zhang, H. Classification of Gan Dan Shi Re Pattern and Gan Shen Yin Xu Pattern in Patients with Hepatitis B Cirrhosis Using Metabonomics. Evid. Based Complement. Altern. Med. 2018, 2018, 2697468. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Xie, G.; Wen, J.; Wang, J.; Pan, W.; Zhou, Y.; Xiao, Y.; Wang, Y.; Jia, W.; Cai, W. Histamine is correlated with liver fibrosis in biliary atresia. Dig. Liver Dis. 2016, 48, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, B.; Montoliu, C.; MacIntyre, D.A.; Serra, M.A.; Wassel, A.; Jover, M.; Romero-Gomez, M.; Rodrigo, J.M.; Pineda-Lucena, A.; Felipo, V. Serum metabolic signature of minimal hepatic encephalopathy by (1)H-nuclear magnetic resonance. J. Proteome Res. 2010, 9, 5180–5187. [Google Scholar] [CrossRef]
- Martinez-Granados, B.; Morales, J.M.; Rodrigo, J.M.; Del Olmo, J.; Serra, M.A.; Ferrandez, A.; Celda, B.; Monleon, D. Metabolic profile of chronic liver disease by NMR spectroscopy of human biopsies. Int. J. Mol. Med. 2011, 27, 111–117. [Google Scholar] [CrossRef]
- Schofield, Z.; Reed, M.A.; Newsome, P.N.; Adams, D.H.; Gunther, U.L.; Lalor, P.F. Changes in human hepatic metabolism in steatosis and cirrhosis. World J. Gastroenterol. 2017, 23, 2685–2695. [Google Scholar] [CrossRef]
- Gao, H.; Lu, Q.; Liu, X.; Cong, H.; Zhao, L.; Wang, H.; Lin, D. Application of 1H NMR-based metabonomics in the study of metabolic profiling of human hepatocellular carcinoma and liver cirrhosis. Cancer Sci. 2009, 100, 782–785. [Google Scholar] [CrossRef]
- Amathieu, R.; Nahon, P.; Triba, M.; Bouchemal, N.; Trinchet, J.C.; Beaugrand, M.; Dhonneur, G.; Le Moyec, L. Metabolomic approach by 1H NMR spectroscopy of serum for the assessment of chronic liver failure in patients with cirrhosis. J. Proteome Res. 2011, 10, 3239–3245. [Google Scholar] [CrossRef]
- Qi, S.W.; Tu, Z.G.; Peng, W.J.; Wang, L.X.; Ou-Yang, X.; Cai, A.J.; Dai, Y. (1)H NMR-based serum metabolic profiling in compensated and decompensated cirrhosis. World J. Gastroenterol. 2012, 18, 285–290. [Google Scholar] [CrossRef]
- Corbin, I.R.; Ryner, L.N.; Singh, H.; Minuk, G.Y. Quantitative hepatic phosphorus-31 magnetic resonance spectroscopy in compensated and decompensated cirrhosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G379–G384. [Google Scholar] [CrossRef]
- Traussnigg, S.; Kienbacher, C.; Gajdosik, M.; Valkovic, L.; Halilbasic, E.; Stift, J.; Rechling, C.; Hofer, H.; Steindl-Munda, P.; Ferenci, P.; et al. Ultra-high-field magnetic resonance spectroscopy in non-alcoholic fatty liver disease: Novel mechanistic and diagnostic insights of energy metabolism in non-alcoholic steatohepatitis and advanced fibrosis. Liver Int. 2017, 37, 1544–1553. [Google Scholar] [CrossRef]
- Iebba, V.; Guerrieri, F.; Di Gregorio, V.; Levrero, M.; Gagliardi, A.; Santangelo, F.; Sobolev, A.P.; Circi, S.; Giannelli, V.; Mannina, L.; et al. Combining amplicon sequencing and metabolomics in cirrhotic patients highlights distinctive microbiota features involved in bacterial translocation, systemic inflammation and hepatic encephalopathy. Sci. Rep. 2018, 8, 8210. [Google Scholar] [CrossRef]
- Liu, Y.; Li, J.; Jin, Y.; Zhao, L.; Zhao, F.; Feng, J.; Li, A.; Wei, Y. Splenectomy Leads to Amelioration of Altered Gut Microbiota and Metabolome in Liver Cirrhosis Patients. Front. Microbiol. 2018, 9, 963. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Ling, Z.; Chen, D.; Liu, Y.; Yang, F.; Li, L. Disorganized Gut Microbiome Contributed to Liver Cirrhosis Progression: A Meta-Omics-Based Study. Front. Microbiol. 2018, 9, 3166. [Google Scholar] [CrossRef]
- Cox, I.J.; Idilman, R.; Fagan, A.; Turan, D.; Ajayi, L.; Le Guennec, A.D.; Taylor-Robinson, S.D.; Karakaya, F.; Gavis, E.; Andrew Atkinson, R.; et al. Metabolomics and microbial composition increase insight into the impact of dietary differences in cirrhosis. Liver Int. 2019. [Google Scholar] [CrossRef]
- Amathieu, R.; Triba, M.N.; Nahon, P.; Bouchemal, N.; Kamoun, W.; Haouache, H.; Trinchet, J.C.; Savarin, P.; Le Moyec, L.; Dhonneur, G. Serum 1H-NMR metabolomic fingerprints of acute-on-chronic liver failure in intensive care unit patients with alcoholic cirrhosis. PLoS ONE 2014, 9, e89230. [Google Scholar] [CrossRef]
- Dabos, K.J.; Parkinson, J.A.; Sadler, I.H.; Plevris, J.N.; Hayes, P.C. (1)H nuclear magnetic resonance spectroscopy-based metabonomic study in patients with cirrhosis and hepatic encephalopathy. World J. Hepatol. 2015, 7, 1701–1707. [Google Scholar] [CrossRef]
- Weiss, N.; Barbier Saint Hilaire, P.; Colsch, B.; Isnard, F.; Attala, S.; Schaefer, A.; Amador, M.D.; Rudler, M.; Lamari, F.; Sedel, F.; et al. Cerebrospinal fluid metabolomics highlights dysregulation of energy metabolism in overt hepatic encephalopathy. J. Hepatol. 2016, 65, 1120–1130. [Google Scholar] [CrossRef]
- Embade, N.; Marino, Z.; Diercks, T.; Cano, A.; Lens, S.; Cabrera, D.; Navasa, M.; Falcon-Perez, J.M.; Caballeria, J.; Castro, A.; et al. Metabolic Characterization of Advanced Liver Fibrosis in HCV Patients as Studied by Serum 1H-NMR Spectroscopy. PLoS ONE 2016, 11, e0155094. [Google Scholar] [CrossRef]
- Gabbani, T.; Marsico, M.; Bernini, P.; Lorefice, E.; Grappone, C.; Biagini, M.R.; Milani, S.; Annese, V. Metabolomic analysis with (1)H-NMR for non-invasive diagnosis of hepatic fibrosis degree in patients with chronic hepatitis C. Dig. Liver Dis. 2017, 49, 1338–1344. [Google Scholar] [CrossRef]
- Wei, D.D.; Wang, J.S.; Wang, P.R.; Li, M.H.; Yang, M.H.; Kong, L.Y. Toxic effects of chronic low-dose exposure of thioacetamide on rats based on NMR metabolic profiling. J. Pharm. Biomed. Anal. 2014, 98, 334–338. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, X.; Hu, P.; Zhou, W.; Zhang, M.; Liu, J.; Wang, Y.; Liu, P.; Luo, G. Serum Metabolomic Characterization of Liver Fibrosis in Rats and Anti-Fibrotic Effects of Yin-Chen-Hao-Tang. Molecules 2016, 21, 126. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.N.; Zhang, G.B.; Lu, Y.Y.; Chen, Q.L.; Yang, L.; Wang, Z.T.; Liu, P.; Su, S.B. Huangqi decoction alleviates dimethylnitrosamine-induced liver fibrosis: An analysis of bile acids metabolic mechanism. J. Ethnopharmacol. 2016, 189, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; He, J.Q.; Chen, D.Y.; Pan, Q.L.; Yang, J.F.; Cao, H.C.; Li, L.J. Dynamic changes of key metabolites during liver fibrosis in rats. World J. Gastroenterol. 2019, 25, 941–954. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, Y.; Li, N.; Xing, F.; Zhao, J.; Yang, T.; Liu, C.; Feng, N. An herbal-compound-based combination therapy that relieves cirrhotic ascites by affecting the L-arginine/nitric oxide pathway: A metabolomics-based systematic study. J. Ethnopharmacol. 2019, 241, 112034. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, Z.; Li, H.; Li, L.; Li, J.; Yu, C. Metabolic Profile Changes of CCl(4)-Liver Fibrosis and Inhibitory Effects of Jiaqi Ganxian Granule. Molecules 2016, 21, 698. [Google Scholar] [CrossRef]
- Chang, H.; Meng, H.Y.; Wang, Y.; Teng, Z.; Liu, S.M. [Inhibitory effect of Scutellariae Radix on hepatic fibrosis based on urinary metabonomic]. Zhongguo Zhong Yao Za Zhi 2018, 43, 2140–2146. [Google Scholar] [CrossRef]
- Zheng, M.; Li, Y.Y.; Wang, G.F.; Jin, J.Y.; Wang, Y.H.; Wang, T.M.; Yang, L.; Liu, S.Y.; Wu, J.S.; Wang, Z.T.; et al. Protective effect of cultured bear bile powder against dimethylnitrosamine-induced hepatic fibrosis in rats. Biomed. Pharmacother. 2019, 112, 108701. [Google Scholar] [CrossRef]
- EASL; EASD; EASO. EASL-EASD-EASO Clinical Practice Guidelines for the management of non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 1388–1402. [Google Scholar] [CrossRef]
- Puri, P.; Daita, K.; Joyce, A.; Mirshahi, F.; Santhekadur, P.K.; Cazanave, S.; Luketic, V.A.; Siddiqui, M.S.; Boyett, S.; Min, H.K.; et al. The presence and severity of nonalcoholic steatohepatitis is associated with specific changes in circulating bile acids. Hepatology 2018, 67, 534–548. [Google Scholar] [CrossRef]
- Fedchuk, L.; Nascimbeni, F.; Pais, R.; Charlotte, F.; Housset, C.; Ratziu, V.; Group, L.S. Performance and limitations of steatosis biomarkers in patients with nonalcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2014, 40, 1209–1222. [Google Scholar] [CrossRef]
- Mato, J.M.; Alonso, C.; Noureddin, M.; Lu, S.C. Biomarkers and subtypes of deranged lipid metabolism in non-alcoholic fatty liver disease. World J. Gastroenterol. 2019, 25, 3009–3020. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef]
- Puri, P.; Wiest, M.M.; Cheung, O.; Mirshahi, F.; Sargeant, C.; Min, H.K.; Contos, M.J.; Sterling, R.K.; Fuchs, M.; Zhou, H.; et al. The plasma lipidomic signature of nonalcoholic steatohepatitis. Hepatology 2009, 50, 1827–1838. [Google Scholar] [CrossRef] [PubMed]
- Barr, J.; Vazquez-Chantada, M.; Alonso, C.; Perez-Cormenzana, M.; Mayo, R.; Galan, A.; Caballeria, J.; Martin-Duce, A.; Tran, A.; Wagner, C.; et al. Liquid chromatography-mass spectrometry-based parallel metabolic profiling of human and mouse model serum reveals putative biomarkers associated with the progression of nonalcoholic fatty liver disease. J. Proteome Res. 2010, 9, 4501–4512. [Google Scholar] [CrossRef] [PubMed]
- Kalhan, S.C.; Guo, L.; Edmison, J.; Dasarathy, S.; McCullough, A.J.; Hanson, R.W.; Milburn, M. Plasma metabolomic profile in nonalcoholic fatty liver disease. Metabolism 2011, 60, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, L.; Yan, X.; Liu, Q.; Yu, C.; Wei, H.; Li, Y.; Zhang, X.; He, F.; Jiang, Y. A proton nuclear magnetic resonance metabonomics approach for biomarker discovery in nonalcoholic fatty liver disease. J. Proteome Res. 2011, 10, 2797–2806. [Google Scholar] [CrossRef]
- Lehmann, R.; Franken, H.; Dammeier, S.; Rosenbaum, L.; Kantartzis, K.; Peter, A.; Zell, A.; Adam, P.; Li, J.; Xu, G.; et al. Circulating lysophosphatidylcholines are markers of a metabolically benign nonalcoholic fatty liver. Diabetes Care 2013, 36, 2331–2338. [Google Scholar] [CrossRef]
- Ferslew, B.C.; Xie, G.; Johnston, C.K.; Su, M.; Stewart, P.W.; Jia, W.; Brouwer, K.L.; Barritt, A.S.t. Altered Bile Acid Metabolome in Patients with Nonalcoholic Steatohepatitis. Dig. Dis. Sci. 2015, 60, 3318–3328. [Google Scholar] [CrossRef]
- Lake, A.D.; Novak, P.; Shipkova, P.; Aranibar, N.; Robertson, D.G.; Reily, M.D.; Lehman-McKeeman, L.D.; Vaillancourt, R.R.; Cherrington, N.J. Branched chain amino acid metabolism profiles in progressive human nonalcoholic fatty liver disease. Amino. Acids 2015, 47, 603–615. [Google Scholar] [CrossRef]
- Loomba, R.; Quehenberger, O.; Armando, A.; Dennis, E.A. Polyunsaturated fatty acid metabolites as novel lipidomic biomarkers for noninvasive diagnosis of nonalcoholic steatohepatitis. J. Lipid Res. 2015, 56, 185–192. [Google Scholar] [CrossRef]
- Mannisto, V.T.; Simonen, M.; Hyysalo, J.; Soininen, P.; Kangas, A.J.; Kaminska, D.; Matte, A.K.; Venesmaa, S.; Kakela, P.; Karja, V.; et al. Ketone body production is differentially altered in steatosis and non-alcoholic steatohepatitis in obese humans. Liver Int. 2015, 35, 1853–1861. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Gallego, E.; Guirro, M.; Riera-Borrull, M.; Hernandez-Aguilera, A.; Marine-Casado, R.; Fernandez-Arroyo, S.; Beltran-Debon, R.; Sabench, F.; Hernandez, M.; del Castillo, D.; et al. Mapping of the circulating metabolome reveals alpha-ketoglutarate as a predictor of morbid obesity-associated non-alcoholic fatty liver disease. Int. J. Obes. 2015, 39, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Oresic, M.; Leivonen, M.; Gopalacharyulu, P.; Hyysalo, J.; Arola, J.; Verrijken, A.; Francque, S.; Van Gaal, L.; Hyotylainen, T.; et al. Noninvasive Detection of Nonalcoholic Steatohepatitis Using Clinical Markers and Circulating Levels of Lipids and Metabolites. Clin. Gastroenterol. Hepatol. 2016, 14, 1463–1472. [Google Scholar] [CrossRef] [PubMed]
- Chiappini, F.; Coilly, A.; Kadar, H.; Gual, P.; Tran, A.; Desterke, C.; Samuel, D.; Duclos-Vallee, J.C.; Touboul, D.; Bertrand-Michel, J.; et al. Metabolism dysregulation induces a specific lipid signature of nonalcoholic steatohepatitis in patients. Sci. Rep. 2017, 7, 46658. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Zhan, Z.Y.; Cao, H.Y.; Wu, C.; Bian, Y.Q.; Li, J.Y.; Cheng, G.H.; Liu, P.; Sun, M.Y. Urinary metabolomics analysis identifies key biomarkers of different stages of nonalcoholic fatty liver disease. World J. Gastroenterol. 2017, 23, 2771–2784. [Google Scholar] [CrossRef]
- O’Sullivan, J.F.; Morningstar, J.E.; Yang, Q.; Zheng, B.; Gao, Y.; Jeanfavre, S.; Scott, J.; Fernandez, C.; Zheng, H.; O’Connor, S.; et al. Dimethylguanidino valeric acid is a marker of liver fat and predicts diabetes. J. Clin. Investig. 2017, 127, 4394–4402. [Google Scholar] [CrossRef]
- Qi, S.; Xu, D.; Li, Q.; Xie, N.; Xia, J.; Huo, Q.; Li, P.; Chen, Q.; Huang, S. Metabonomics screening of serum identifies pyroglutamate as a diagnostic biomarker for nonalcoholic steatohepatitis. Clin. Chim. Acta 2017, 473, 89–95. [Google Scholar] [CrossRef]
- Keogh, A.; Senkardes, S.; Idle, J.R.; Kucukguzel, S.G.; Beyoglu, D. A Novel Anti-Hepatitis C Virus and Antiproliferative Agent Alters Metabolic Networks in HepG2 and Hep3B Cells. Metabolites 2017, 7, 23. [Google Scholar] [CrossRef]
- Golla, S.; Golla, J.P.; Krausz, K.W.; Manna, S.K.; Simillion, C.; Beyoglu, D.; Idle, J.R.; Gonzalez, F.J. Metabolomic Analysis of Mice Exposed to Gamma Radiation Reveals a Systemic Understanding of Total-Body Exposure. Radiat Res. 2017, 187, 612–629. [Google Scholar] [CrossRef]
- Wang, M.; Keogh, A.; Treves, S.; Idle, J.R.; Beyoglu, D. The metabolomic profile of gamma-irradiated human hepatoma and muscle cells reveals metabolic changes consistent with the Warburg effect. PeerJ 2016, 4, e1624. [Google Scholar] [CrossRef]
- Yang, R.X.; Hu, C.X.; Sun, W.L.; Pan, Q.; Shen, F.; Yang, Z.; Su, Q.; Xu, G.W.; Fan, J.G. Serum Monounsaturated Triacylglycerol Predicts Steatohepatitis in Patients with Non-alcoholic Fatty Liver Disease and Chronic Hepatitis B. Sci. Rep. 2017, 7, 10517. [Google Scholar] [CrossRef] [PubMed]
- Hoyles, L.; Fernandez-Real, J.M.; Federici, M.; Serino, M.; Abbott, J.; Charpentier, J.; Heymes, C.; Luque, J.L.; Anthony, E.; Barton, R.H.; et al. Molecular phenomics and metagenomics of hepatic steatosis in non-diabetic obese women. Nat. Med. 2018, 24, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Mayo, R.; Crespo, J.; Martinez-Arranz, I.; Banales, J.M.; Arias, M.; Minchole, I.; Aller de la Fuente, R.; Jimenez-Aguero, R.; Alonso, C.; de Luis, D.A.; et al. Metabolomic-based noninvasive serum test to diagnose nonalcoholic steatohepatitis: Results from discovery and validation cohorts. Hepatol. Commun. 2018, 2, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Perakakis, N.; Polyzos, S.A.; Yazdani, A.; Sala-Vila, A.; Kountouras, J.; Anastasilakis, A.D.; Mantzoros, C.S. Non-invasive diagnosis of non-alcoholic steatohepatitis and fibrosis with the use of omics and supervised learning: A proof of concept study. Metabolism 2019, 101, 154005. [Google Scholar] [CrossRef] [PubMed]
- Pietzner, M.; Budde, K.; Homuth, G.; Kastenmuller, G.; Henning, A.K.; Artati, A.; Krumsiek, J.; Volzke, H.; Adamski, J.; Lerch, M.M.; et al. Hepatic Steatosis Is Associated With Adverse Molecular Signatures in Subjects Without Diabetes. J. Clin. Endocrinol. Metab. 2018, 103, 3856–3868. [Google Scholar] [CrossRef] [PubMed]
- Romero-Ibarguengoitia, M.E.; Vadillo-Ortega, F.; Caballero, A.E.; Ibarra-Gonzalez, I.; Herrera-Rosas, A.; Serratos-Canales, M.F.; Leon-Hernandez, M.; Gonzalez-Chavez, A.; Mummidi, S.; Duggirala, R.; et al. Family history and obesity in youth, their effect on acylcarnitine/aminoacids metabolomics and non-alcoholic fatty liver disease (NAFLD). Structural equation modeling approach. PLoS ONE 2018, 13, e0193138. [Google Scholar] [CrossRef]
- Boone, S.; Mook-Kanamori, D.; Rosendaal, F.; den Heijer, M.; Lamb, H.; de Roos, A.; le Cessie, S.; Willems van Dijk, K.; de Mutsert, R. Metabolomics: A search for biomarkers of visceral fat and liver fat content. Metabolomics 2019, 15, 139. [Google Scholar] [CrossRef]
- Tan, Y.; Liu, X.; Zhou, K.; He, X.; Lu, C.; He, B.; Niu, X.; Xiao, C.; Xu, G.; Bian, Z.; et al. The Potential Biomarkers to Identify the Development of Steatosis in Hyperuricemia. PLoS ONE 2016, 11, e0149043. [Google Scholar] [CrossRef]
- Von Schonfels, W.; Patsenker, E.; Fahrner, R.; Itzel, T.; Hinrichsen, H.; Brosch, M.; Erhart, W.; Gruodyte, A.; Vollnberg, B.; Richter, K.; et al. Metabolomic tissue signature in human non-alcoholic fatty liver disease identifies protective candidate metabolites. Liver Int. 2015, 35, 207–214. [Google Scholar] [CrossRef]
- Alonso, C.; Fernandez-Ramos, D.; Varela-Rey, M.; Martinez-Arranz, I.; Navasa, N.; Van Liempd, S.M.; Lavin Trueba, J.L.; Mayo, R.; Ilisso, C.P.; de Juan, V.G.; et al. Metabolomic Identification of Subtypes of Nonalcoholic Steatohepatitis. Gastroenterology 2017, 152, 1449–1461. [Google Scholar] [CrossRef]
- Chen, Y.; Li, C.; Liu, L.; Guo, F.; Li, S.; Huang, L.; Sun, C.; Feng, R. Serum metabonomics of NAFLD plus T2DM based on liquid chromatography-mass spectrometry. Clin. Biochem. 2016, 49, 962–966. [Google Scholar] [CrossRef] [PubMed]
- Semmo, N.; Weber, T.; Idle, J.R.; Beyoglu, D. Metabolomics reveals that aldose reductase activity due to AKR1B10 is upregulated in hepatitis C virus infection. J. Viral Hepat. 2015, 22, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.R.; Li, X.; Chan, P.L.; Zhuang, H.; Jia, J.D.; Wang, X.; Lo, Y.R.; Walsh, N. Prevalence of hepatitis C virus infection among key populations in China: A systematic review. Int. J. Infect. Dis. 2019, 80, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Alkhouri, N.; Cikach, F.; Eng, K.; Moses, J.; Patel, N.; Yan, C.; Hanouneh, I.; Grove, D.; Lopez, R.; Dweik, R. Analysis of breath volatile organic compounds as a noninvasive tool to diagnose nonalcoholic fatty liver disease in children. Eur. J. Gastroenterol. Hepatol. 2014, 26, 82–87. [Google Scholar] [CrossRef]
- Reid, D.T.; McDonald, B.; Khalid, T.; Vo, T.; Schenck, L.P.; Surette, M.G.; Beck, P.L.; Reimer, R.A.; Probert, C.S.; Rioux, K.P.; et al. Unique microbial-derived volatile organic compounds in portal venous circulation in murine non-alcoholic fatty liver disease. Biochim. Biophys. Acta 2016, 1862, 1337–1344. [Google Scholar] [CrossRef]
- Pastore, A.; Alisi, A.; di Giovamberardino, G.; Crudele, A.; Ceccarelli, S.; Panera, N.; Dionisi-Vici, C.; Nobili, V. Plasma levels of homocysteine and cysteine increased in pediatric NAFLD and strongly correlated with severity of liver damage. Int. J. Mol. Sci. 2014, 15, 21202–21214. [Google Scholar] [CrossRef]
- Jin, R.; Banton, S.; Tran, V.T.; Konomi, J.V.; Li, S.; Jones, D.P.; Vos, M.B. Amino Acid Metabolism is Altered in Adolescents with Nonalcoholic Fatty Liver Disease-An Untargeted, High Resolution Metabolomics Study. J. Pediatr. 2016, 172, 14–19. [Google Scholar] [CrossRef]
- Troisi, J.; Pierri, L.; Landolfi, A.; Marciano, F.; Bisogno, A.; Belmonte, F.; Palladino, C.; Guercio Nuzio, S.; Campiglia, P.; Vajro, P. Urinary Metabolomics in Pediatric Obesity and NAFLD Identifies Metabolic Pathways/Metabolites Related to Dietary Habits and Gut-Liver Axis Perturbations. Nutrients 2017, 9, 485. [Google Scholar] [CrossRef]
- Goffredo, M.; Santoro, N.; Trico, D.; Giannini, C.; D’Adamo, E.; Zhao, H.; Peng, G.; Yu, X.; Lam, T.T.; Pierpont, B.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature Characterizes Obese Adolescents with Non-Alcoholic Fatty Liver Disease. Nutrients 2017, 9, 642. [Google Scholar] [CrossRef]
- Troisi, J.; Belmonte, F.; Bisogno, A.; Pierri, L.; Colucci, A.; Scala, G.; Cavallo, P.; Mandato, C.; Di Nuzzi, A.; Di Michele, L.; et al. Metabolomic Salivary Signature of Pediatric Obesity Related Liver Disease and Metabolic Syndrome. Nutrients 2019, 11, 274. [Google Scholar] [CrossRef]
- Hartley, A.; Santos Ferreira, D.L.; Anderson, E.L.; Lawlor, D.A. Metabolic profiling of adolescent non-alcoholic fatty liver disease. Wellcome Open Res. 2018, 3, 166. [Google Scholar] [CrossRef]
- Khusial, R.D.; Cioffi, C.E.; Caltharp, S.A.; Krasinskas, A.M.; Alazraki, A.; Knight-Scott, J.; Cleeton, R.; Castillo-Leon, E.; Jones, D.P.; Pierpont, B.; et al. Development of a Plasma Screening Panel for Pediatric Nonalcoholic Fatty Liver Disease Using Metabolomics. Hepatol. Commun. 2019, 3, 1311–1321. [Google Scholar] [CrossRef]
- Tanaka, N.; Matsubara, T.; Krausz, K.W.; Patterson, A.D.; Gonzalez, F.J. Disruption of phospholipid and bile acid homeostasis in mice with nonalcoholic steatohepatitis. Hepatology 2012, 56, 118–129. [Google Scholar] [CrossRef]
- Tanaka, N.; Takahashi, S.; Fang, Z.Z.; Matsubara, T.; Krausz, K.W.; Qu, A.; Gonzalez, F.J. Role of white adipose lipolysis in the development of NASH induced by methionine- and choline-deficient diet. Biochim. Biophys. Acta 2014, 1841, 1596–1607. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.; Stevens, A.P.; Klein, M.S.; Hellerbrand, C.; Dettmer, K.; Gronwald, W.; Oefner, P.J.; Reinders, J. Early changes in the liver-soluble proteome from mice fed a nonalcoholic steatohepatitis inducing diet. Proteomics 2012, 12, 1437–1451. [Google Scholar] [CrossRef] [PubMed]
- Depner, C.M.; Traber, M.G.; Bobe, G.; Kensicki, E.; Bohren, K.M.; Milne, G.; Jump, D.B. A metabolomic analysis of omega-3 fatty acid-mediated attenuation of western diet-induced nonalcoholic steatohepatitis in LDLR-/- mice. PLoS ONE 2013, 8, e83756. [Google Scholar] [CrossRef]
- Lake, A.D.; Novak, P.; Shipkova, P.; Aranibar, N.; Robertson, D.; Reily, M.D.; Lu, Z.; Lehman-McKeeman, L.D.; Cherrington, N.J. Decreased hepatotoxic bile acid composition and altered synthesis in progressive human nonalcoholic fatty liver disease. Toxicol. Appl. Pharmacol. 2013, 268, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.; Huo, T.I.; Cheng, H.Y.; Tsai, J.C.; Liao, J.W.; Lee, M.S.; Qin, X.M.; Hsieh, M.T.; Pao, L.H.; Peng, W.H. Gallic acid ameliorated impaired glucose and lipid homeostasis in high fat diet-induced NAFLD mice. PLoS ONE 2014, 9, e96969. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.S.; Chen, W.C.; Kuo, T.C.; Ho, C.T.; Kuo, C.H.; Tseng, Y.J.; Lu, K.H.; Lin, S.H.; Panyod, S.; Sheen, L.Y. Mass-Spectrometry-Based Serum Metabolomics of a C57BL/6J Mouse Model of High-Fat-Diet-Induced Non-alcoholic Fatty Liver Disease Development. J. Agric. Food Chem. 2015, 63, 7873–7884. [Google Scholar] [CrossRef]
- Tu, L.N.; Showalter, M.R.; Cajka, T.; Fan, S.; Pillai, V.V.; Fiehn, O.; Selvaraj, V. Metabolomic characteristics of cholesterol-induced non-obese nonalcoholic fatty liver disease in mice. Sci. Rep. 2017, 7, 6120. [Google Scholar] [CrossRef]
- Chen, C.J.; Liao, W.L.; Chang, C.T.; Lin, Y.N.; Tsai, F.J. Identification of Urinary Metabolite Biomarkers of Type 2 Diabetes Nephropathy Using an Untargeted Metabolomic Approach. J. Proteome Res. 2018, 17, 3997–4007. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.; Sultan, M.; Kashofer, K.; Ralser, M.; Amstislavskiy, V.; Starmann, J.; Osprian, I.; Grimm, C.; Hache, H.; Yaspo, M.L.; et al. Comparative analysis and modeling of the severity of steatohepatitis in DDC-treated mouse strains. PLoS ONE 2014, 9, e111006. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.J.; Pacana, T. A Lipidomic Readout of Disease Progression in A Diet-Induced Mouse Model of Nonalcoholic Fatty Liver Disease. Trans. Am. Clin. Climatol. Assoc. 2015, 126, 271–288. [Google Scholar] [PubMed]
- Patterson, R.E.; Kalavalapalli, S.; Williams, C.M.; Nautiyal, M.; Mathew, J.T.; Martinez, J.; Reinhard, M.K.; McDougall, D.J.; Rocca, J.R.; Yost, R.A.; et al. Lipotoxicity in steatohepatitis occurs despite an increase in tricarboxylic acid cycle activity. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E484–E494. [Google Scholar] [CrossRef]
- Qian, M.; Hu, H.; Zhao, D.; Wang, S.; Pan, C.; Duan, X.; Gao, Y.; Liu, J.; Zhang, Y.; Yang, S.; et al. Coordinated changes of gut microbiome and lipidome differentiates nonalcoholic steatohepatitis (NASH) from isolated steatosis. Liver Int. 2019. [Google Scholar] [CrossRef]
- Kim, K.E.; Jung, Y.; Min, S.; Nam, M.; Heo, R.W.; Jeon, B.T.; Song, D.H.; Yi, C.O.; Jeong, E.A.; Kim, H.; et al. Caloric restriction of db/db mice reverts hepatic steatosis and body weight with divergent hepatic metabolism. Sci. Rep. 2016, 6, 30111. [Google Scholar] [CrossRef]
- Gogiashvili, M.; Edlund, K.; Gianmoena, K.; Marchan, R.; Brik, A.; Andersson, J.T.; Lambert, J.; Madjar, K.; Hellwig, B.; Rahnenfuhrer, J.; et al. Metabolic profiling of ob/ob mouse fatty liver using HR-MAS (1)H-NMR combined with gene expression analysis reveals alterations in betaine metabolism and the transsulfuration pathway. Anal. Bioanal. Chem. 2017, 409, 1591–1606. [Google Scholar] [CrossRef]
- Li, J.; Liu, Z.; Guo, M.; Xu, K.; Jiang, M.; Lu, A.; Gao, X. Metabolomics profiling to investigate the pharmacologic mechanisms of berberine for the treatment of high-fat diet-induced nonalcoholic steatohepatitis. Evid. Based Complement. Altern. Med. 2015, 2015, 897914. [Google Scholar] [CrossRef]
- Wang, Y.; Niu, M.; Jia, G.L.; Li, R.S.; Zhang, Y.M.; Zhang, C.E.; Meng, Y.K.; Cui, H.R.; Ma, Z.J.; Li, D.H.; et al. Untargeted Metabolomics Reveals Intervention Effects of Total Turmeric Extract in a Rat Model of Nonalcoholic Fatty Liver Disease. Evid. Based Complement. Altern. Med. 2016, 2016, 8495953. [Google Scholar] [CrossRef]
- Van Pelt, C.K.; Huang, M.C.; Tschanz, C.L.; Brenna, J.T. An octaene fatty acid, 4,7,10,13,16,19,22,25-octacosaoctaenoic acid (28:8n-3), found in marine oils. J. Lipid Res. 1999, 40, 1501–1505. [Google Scholar]
- Lu, Y.; Chen, Y.; Wu, Y.; Hao, H.; Liang, W.; Liu, J.; Huang, R. Marine unsaturated fatty acids: Structures, bioactivities, biosynthesis and benefits. RSC Adv. 2019, 9, 35312–35327. [Google Scholar] [CrossRef]
- Han, J.; Dzierlenga, A.L.; Lu, Z.; Billheimer, D.D.; Torabzadeh, E.; Lake, A.D.; Li, H.; Novak, P.; Shipkova, P.; Aranibar, N.; et al. Metabolomic profiling distinction of human nonalcoholic fatty liver disease progression from a common rat model. Obesity (Silver Spring) 2017, 25, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Ming, Y.N.; Zhang, J.Y.; Chen, X.Y.; Zeng, M.D.; Mao, Y.M. Gene-metabolite network analysis in different nonalcoholic fatty liver disease phenotypes. Exp. Mol. Med. 2017, 49, e283. [Google Scholar] [CrossRef] [PubMed]
- Maciejewska, D.; Palma, J.; Dec, K.; Skonieczna-Zydecka, K.; Gutowska, I.; Szczuko, M.; Jakubczyk, K.; Stachowska, E. Is the Fatty Acids Profile in Blood a Good Predictor of Liver Changes? Correlation of Fatty Acids Profile with Fatty Acids Content in the Liver. Diagnostics 2019, 9, 197. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhang, J.; Li, J.; Lei, X.; Xu, D.; Wang, Y.; Li, C.; Li, X.; Mao, Y. Turnover of bile acids in liver, serum and caecal content by high-fat diet feeding affects hepatic steatosis in rats. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Papandreou, C.; Bullo, M.; Tinahones, F.J.; Martinez-Gonzalez, M.A.; Corella, D.; Fragkiadakis, G.A.; Lopez-Miranda, J.; Estruch, R.; Fito, M.; Salas-Salvado, J. Serum metabolites in non-alcoholic fatty-liver disease development or reversion; a targeted metabolomic approach within the PREDIMED trial. Nutr. Metab. 2017, 14, 58. [Google Scholar] [CrossRef]
- Del Chierico, F.; Nobili, V.; Vernocchi, P.; Russo, A.; De Stefanis, C.; Gnani, D.; Furlanello, C.; Zandona, A.; Paci, P.; Capuani, G.; et al. Gut microbiota profiling of pediatric nonalcoholic fatty liver disease and obese patients unveiled by an integrated meta-omics-based approach. Hepatology 2017, 65, 451–464. [Google Scholar] [CrossRef]
- Gawlik, A.; Shmoish, M.; Hartmann, M.F.; Wudy, S.A.; Olczak, Z.; Gruszczynska, K.; Hochberg, Z. Steroid metabolomic signature of liver disease in nonsyndromic childhood obesity. Endocr. Connect. 2019, 8, 764–771. [Google Scholar] [CrossRef]
- Robertson, D.G. Metabonomics in toxicology: A review. Toxicol. Sci. 2005, 85, 809–822. [Google Scholar] [CrossRef]
- Miao, H.; Zhao, Y.H.; Vaziri, N.D.; Tang, D.D.; Chen, H.; Chen, H.; Khazaeli, M.; Tarbiat-Boldaji, M.; Hatami, L.; Zhao, Y.Y. Lipidomics Biomarkers of Diet-Induced Hyperlipidemia and Its Treatment with Poria cocos. J. Agric. Food Chem. 2016, 64, 969–979. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Alcohol Dose | Pathology | Metabolites Reported | Ref. |
|---|---|---|---|---|
| Rat | 20% or 36% of total calories; 24 days | Hepatomegaly Fatty infiltration | Plasma triglycerides↑ Plasma phospholipids↑ Hepatic triglycerides↑ 8-fold | [53] |
| Rat | 5% alcohol Lieber-DeCarli diet; 2-3 months | Fatty infiltration Mild inflammatory infiltrate; 3 months Mild oxidative stress, 3 months | Liver triglycerides↑ Liver cholesterol↑ Liver phospholipids and lysophospholipids↓ | [60] |
| Rat | 6 g/kg alcohol + high-fat diet | Regional laminar necrosis and edema around central vein. Inflammatory cell infiltrate. | Total of 37 core ALD biomarkers identified. Pathways perturbed included TCA cycle, carbohydrate and amino acid metabolism. | [70] |
| Mouse | 5 g/kg every 12 h X 3 | Serum ALT↑ Hepatic CYP2E1↑ | Malondialdehyde↑ Methionine↑ Hypotaurine↑ Taurine↑ SAM↓ GSH↓ | [56] |
| Mouse 129 Sv WT and Ppara-null | 4% alcohol Lieber-DeCarli diet; 2–6 months | Little change after 1 month | Ethylsulfate↑ Ethyl-β-D-glucuronide↑ 4-hydroxyphenylacetic acid (4HPAA)↑ 4HPAA sulfate↑ in both WT and null. Indole-3-lactic acid↑ in null only. | [58] |
| Mouse 129 Sv and C57BL/6 WT and Ppara-null | 4% alcohol Lieber-DeCarli diet; 1 month | Steatosis in B6 null mice | Indole-3-lactic acid↑ and phenyllactic acid↑ in alcohol-treated Ppara-null mouse, both 129 Sv and C57BL/6 | [59] |
| Mouse WT and Cyp2e1-null | 2.2%, 4.5%, 5.4% Lieber-DeCarli semi-solid diet; 21 days | CYP2E1↑ in WT Microvesicular and macrovesicular steatosis around central vein; WT>null | Hepatic and serum triglycerides↑ in WT only. Urinary N-acetyltaurine, 4HPAA sulfate, ethylsulfate, ethyl-β-D-glucuronide↑ | [62] |
| Mouse | 4.896 g/kg; 7 days | ALT↑ AST↑ Focal hepatic necrosis Inflammatory infiltrate | Serum Malondialdehyde↑ GSH↓ GSSG↑ Methylglyoxal↑ | [71] |
| Mouse | 5% alcohol Lieber-DeCarli diet; 8 weeks | Mild steatohepatitis No fibrosis | Correlation between urinary and fecal metabolites. Many fecal and urinary metabolites altered. Amino acid metabolism perturbed. Indole-3-lactic acid↑ | [72] |
| Mouse Cramp-null and WT | 5% alcohol Lieber-DeCarli diet; 24 days | Not clearly stated | In alcohol-fed WT, fecal taurine, α-aminoisobutyric acid, nicotinic acid, serine, SCFAs↓ In alcohol-fed null mice, only nicotinic acid↑ | [73] |
| Micropig | 40% total calories Folate-depleted diet | Not determined | Hepatic triglycerides↑ SCD pathway↑ ELOVL5 pathway↑ PEMT pathway↓ Phospholipid export↓ | [57] |
| Human | 100–300 g/day; 10 days 118 g/day; 11 days, 141 g/day; 8 days | Fatty infiltration | Plasma triglycerides↑ | [53] |
| Human | 30 ALD patients (mean daily alcohol consumption 109.7 g/day) vs. 10 healthy controls | Cirrhosis (80%) Decompensated cirrhosis (DC; 23%) | N-Lauroylglycine↑ in cirrhosis. Decatrienoic acid associated with disease severity. | [61] |
| Human | 30 Alcohol use disorder (AUD), 13 alcoholic hepatitis (AH) and 16 nonalcoholic controls | ALT↑ (AUD = AH) AST↑ (AH>>AUD) | Seven serum oxylipins and nine fecal oxylipins↑ Results related to inflammation and platelet aggregation. Inflammatory ω-6 PUFA oxylipins counteracted by ω-3 bioactive lipid mediators. | [74] |
| Human | 64 AH patients, 26 DC patients without AH | AST and GGT↑ (AH > DC). Other serum markers and MELD score AH = DC | Metabolomic signature of AH claimed but not disclosed. | [75] |
| Species | Manipulation/Condition | Pathology | Analytical Methodology | Metabolites Reported | Ref. |
|---|---|---|---|---|---|
| Rat | Inhibition of bile secretion vs. bile flow obstruction | Intrahepatic cholestasis vs. extrahepatic cholestasis | 1H NMR | Bile acids↑ Bilirubin↑ vs. Bile acids↑ BCAAs↑ SCFAs↑ | [77] |
| Mouse | Fxr-null vs. WT treated with FXR ligands CA and LCA | Cholestasis | UPLC-ESI-QTOFMS | p-Cresol sulfate and β-D-glucuronide↑ Corticosterone metabolites↑ Cholic acid metabolites↑ | [78] |
| Rat | Eisai hyperbilirubinemic rat | Cholestasis | UPLC-TQMS | Taurine↑ Hypotaurine↑ Unconjugated primary and secondary bile acids↑ | [81] |
| Rat | ANIT Methapyrilene Dimethylnitrosamine | Cholestasis | UPLC-TQMS GC-MS UPLC-QTOFMS | Bile acids↑ Arginine↓ Pantothenate↑ Protoporphyrin IX↑ Palmitoyl carnitine↑ Arachidonic, linoleic and oleic acids↓ | [82,83,84,85] |
| Mouse | Vps33b-depleted mouse | Cholestasis | UPLC-MS | Serum bile acids↑ triglycerides↑ and sphingomyelins↑ | [97] |
| Rat | Bile duct ligation (BDL) | Cholestasis | UPLC-QTOFMS | Phenylalanine↑ Glutamate↑ Tyrosine↑ Kynurenine↑ Lactate↑ LPC(14:0) ↑ Glycine↑ Succinate↑ MDA↑ GSH↑ Valine↓ Isoleucine↓ Citrate↓ Palmitate↓ Taurine↓ LPC(19:0)↓ | [98] |
| Rat | TAA or BDL | Cholestasis | 1H NMR | BDL vs. TAA: 2-Hydroxybutyrate↑ BCAAs↑ Lysine↑ Arginine↑ Glycine↑ Citrate↑ 2-Oxoglutarate↑ Fumarate↑ Hippurate↑ Phenacetylglycine↑ | [99] |
| Mouse | ANIT or DDC or LCA | Cholestasis | UPLC-QTOFMS | Phospholipids↑ Protoporphyrin IX↑ GSH↓ | [100] |
| Human | Primary biliary cholangitis | Cholestasis | UPLC-QTOFMS | Primary bile acids↑ Phospholipids↑ Oleic and Linoleic acids↑ | [101,102,103] |
| Human | Intrahepatic cholestasis of pregnancy (ICP) | Cholestasis | HPLC-QTOFMS | MG(22:5) ↑ LPE(22:5) ↑ L-Homocysteine sulfonic acid↑ Glycocholic acid↑ Chenodeoxycholic acid 3-sulfate↑ | [107] |
| Human | Hypercholanemia of pregnancy (HCP) vs. ICP | Cholestasis | UPLC-QTOFMS | Sulfated bile acid pattern used for differential diagnosis of HCP and ICP | [110] |
| Human | Infantile hepatitis syndrome (IHS) vs. biliary atresia | Cholestasis | GC-MS | N-Acetyl-D-mannosamine and α-Aminoadipic acid used for differential diagnosis | [113] |
| Species | Manipulation/Condition | Pathology | Analytical Methodology | Metabolites Reported | Ref. |
|---|---|---|---|---|---|
| Human | NAFLD | Fibrosis progression | CE-TOFMS LC-TOFMS | F0/F1→F4 Etiocholanolone sulfate↓ Dehydroepiandrosterone sulfate↓ 16α-hydroxy-dehydroepiandrosterone sulfate↑ (all in serum) | [119] |
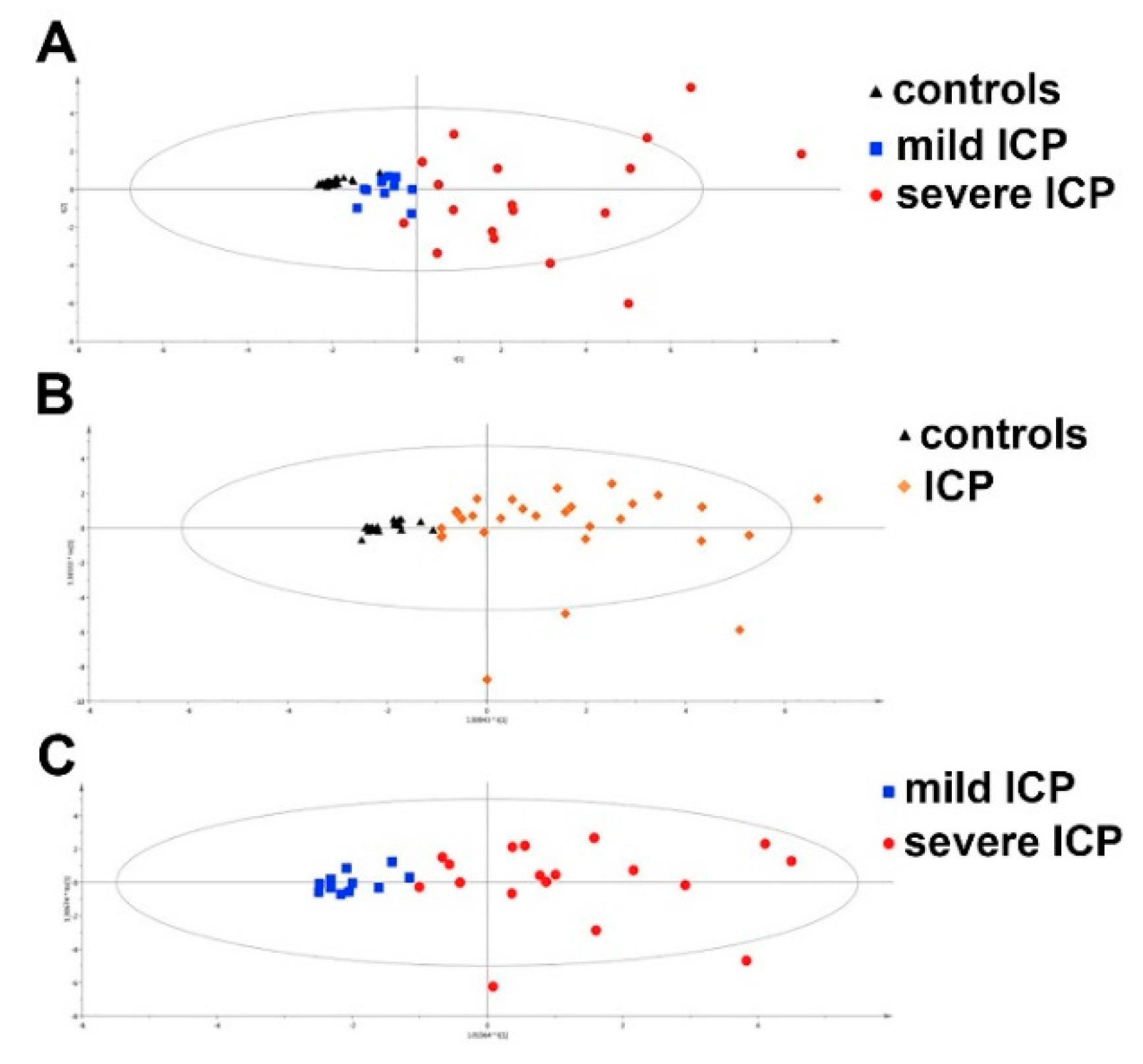
| Human | Fibrosis or Cirrhosis | Significant fibrosis, advanced fibrosis, cirrhosis | 1H NMR | No metabolites reported, only multivariate model used to distinguish pathologies. | [121] |
| Human | Chronic hepatitis B | Cirrhosis | UPLC-QTOFMS | Glycocholic acid↑ Glycochenodeoxycholic acid↑ Taurocholic acid↑ Taurochenodeoxycholic acid↑ (all in serum) | [127] |
| Human | Chronic hepatitis B | Cirrhosis | GC-MS | Acetate↑ Hexanoate↑ Butanoate↑ Glucose↓ Sorbitol↓ (all in serum) | [128] |
| Human | Causes not stated | Cirrhosis | 2D-GC-TOFMS | D-Alanine↑ D-Proline↑ D-Valine↑ D-Leucine↑ D-Threonine↑ (all in serum) | [129] |
| Human | HBV, HCV, alcohol | Cirrhosis | UPLC-QTOFMS | Chenodeoxycholic acid↓ 7-Ketolithocholic acid↓ Urobilin↓ Urobilinogen↓ LPC(16:0)↑ LPC(18:0)↑ LPC(18:1)↑ LPC(18:2)↑ (all in feces) | [131] |
| Human | HBV, alcohol, PBC, cryptogenic cirrhosis | Cirrhosis | UPLC-TQMS | Taurocholic acid↑ Taurochenodeoxycholic acid↑ Tauroursodeoxycholic acid↑ Glycocholic acid↑ Ursodeoxycholic acid↑ Chenodeoxycholic acid↑ Cholic acid↑ Taurolithocholic acid↑ Taurodeoxycholic acid↑ Hyodeoxycholic acid↑ Lithocholic acid↑ Deoxycholic acid↑ (all in serum) | [132] |
| Human | Chronic hepatitis C | Cirrhosis | GC-MS | Proline↑ Serine↑ Glycine↑ Threonine↑ Citrate↑ Xylitol↓ Arabinose↓ Urea↓ (all in urine) | [134] |
| Human | Chronic hepatitis B | Cirrhosis | UPLC-QTOFMS | Phenylalanine↑ Glycochenodeoxycholic acid↑ Oleamide↑ LPC(16:0)↓ PC(16:0/18:2)↓ PC(16:0/22:6)↓ PC(16:0/20:4)↓ PC(18:0/18:2)↓ Canavaninosuccinate↓ (all in serum) | [135] |
| Human | Hepatorenal syndrome | Cirrhosis | UPLC-TQMS | 4-Acetamidobutanoate↑ (in plasma) | [138] |
| Human | Chronic hepatitis B | Cirrhosis | 1H NMR UPLC-QTOFMS | Tyrosine↑ Oxaloacetate↑ Phenylalanine↑ C16-Sphinganine↑ Phytosphingosine↑ Isobutyrate↑ LPC(18:1) ↑ Linoelaidic acid↑ Bilirubin↑ PC(18:4/20:1)↓ PC(14:1/14:1)↓ LPC(16:0)↓ Formate↓ Ascorbate↓ Carnitine↓ α-CEHC↓ (all in serum) | [139] |
| Human | Causes not stated | Decompensated cirrhosis (90-day mortality vs. survivors) | 1H NMR UPLC-QTOFMS | Isoleucine↑ Leucine↑ Lactate↑ Creatinine↑ Urea↑ Tyrosine↑ Histidine↑ Phenylalanine↑ Formate↑ LPC(16:0) ↑ Pyruvate↓ Choline↓ Phosphocholine↓ Glycine↓ Glucose↓ PC(34:2)↓ PC(18:2/18:2)↓ PC(16:0/18:2)↓ PC(18:0/18:2)↓ LPC(18:2)↓ PC(18:2/18:5)↓ PC(22:5/20:4)↓ PI(37:2)↓ PS(41:4)↓ (all in plasma) | [140] |
| Human | Causes not stated | Cirrhosis | UPLC-Orbitrap MS | 16α-Hydroxyestrone↑ 4-Androstenedione↓ 17α-Hydroxyprogesterone↓ 18-Hydroxycorticosterone↓ Cortisol↓ Cortexolone↓ Allotetrahydrocortisol↓ Deoxycorticosterone↓ Epitestosterone↓ Testosterone↓ Dehydroepiandrosterone↓ Etiocholanolone↓ Tetrahydrodeoxycortisol↓ Androsterone↓ 17α-Hydroxypregnenolone↓ Epiandrosterone↓ 11-Oxoetiocholanolone↓ 7β-Hydroxy-dehydroepiandrosterone↓ Androstenetriol↓ Androstenediol↓ Pregnanediol↓ (all in urine) | [141] |
| Human | Chronic hepatitis B | Cirrhosis (vs. HBV) | GC-TOFMS | Serine↑ 5-Oxoproline↑ Phenylalanine↑ Tyrosine↑ Ornithine↑ Citrate↑ Palmitic acid↑ Fructose↓ Glutamate↓ Indole-3-acetic acid↓ arachidonic acid↓ 2-Deoxy-D-glucose↓ (all in serum) | [142] |
| Human | Chronic hepatitis B | Cirrhosis (vs. HBV/HV) | UPLC-TQMS | 9,10-DiHOME↑ 13-HODE↑ TXB2↓ (all in serum) | [144] |
| Human | Chronic hepatitis B | Cirrhosis (GDSR and GSYX patterns, vs. latent pattern (LP)) | GC-TOFMS | GDSR vs. LP: Nonanoate↑ Urea↓ Serine↓ 2-Hydroxybutyrate↓ 2-Hydroxyglutarate↓ Phenylalanine↓ Asparagine↓ Citrulline↓ Tyrosine↓ Arabinose↓ Sorbose↓ Fructose↓ Myristate↓ Palmitolate↓ Palmitate↓ Linolate↓ Tryptamine↓ Glycolate↓ Quinate↓ Petroselinate↓ GSYX vs. LP: 1,5-Anhydrosorbitol↑ Fructose↑ 2-Hydroxybutyrate↓ Serine↓ Threonine↓ 5-Oxoglutarate↓ 2-Hydroxyglutarate↓ Phenylalanine↓ Asparagine↓ Tyrosine↓ Arabinose↓ Arabitol↓ Nonanoate↓ Glycerate↓ Pipecolate↓ Glutarate↓ Quinate↓ α-Tocopherol↓ (all in serum) | [146] |
| Human | Biliary atresia (BA) and neonatal hepatitis syndrome (NHS) | Fibrosis F1 to F4 | UPLC-TQMS | BA/NHS: Histamine↑ Methionine↓ Phenylalanine↓ Serine↓ Threonine↓ Valine↓ Glutamine↓ Sarcosine↓ Lysine↓ F4>F3>F1/F2 in BA: Histamine↑ (all in liver homogenates) | [147] |
| Human | Alcohol | Cirrhosis ± minimal hepatic encephalopathy (MHE) | 1H NMR | MHE+/MHE-: Lactate↑ Glucose↑ TMAO↑ Glycerol↑ LDL↓ VLDL↓ Isoleucine↓ Leucine↓ Valine↓ Alanine↓ Acetoacetate↓ Choline↓ Glycine↓ (all in serum) | [148] |
| Human | Chronic hepatitis C, Chronic hepatitis B, Alcohol, Autoimmunity | Cirrhosis | MAS 1H NMR | Phosphoethanolamine↑ Phosphocholine↑ Glutamate↑ Aspartate↓ α-Glucose↓ β-Glucose↓ (all in liver) | [149] |
| Human | ALD, NASH | Cirrhosis | 1H NMR | ALD Cirrhosis: Isoleucine↑ Valine↑ 1,2-Propanediol↑ Succinate↑ Aspartate↑ Betaine↑ Lactate↑ Glucose↑ Uracil↑ Phenylalanine↑ NASH Cirrhosis: Leucine↑ Isoleucine↑ Valine↑ 1,2-Propanediol↑ Succinate↑ Aspartate↑ Betaine↑ Lactate↑ Phenylalanine↑ Uracil↑ Uridine↓ Inosine↓ (all in liver) | [150] |
| Human | Causes not stated | Cirrhosis | 1H NMR | Acetate↑ Pyruvate↑ Glutamine↑ α-Ketoglutarate↑ Taurine↑ Glycerol↑ Tyrosine↑ 1-Methylhistidine↑ Phenylalanine↑ N-Acetylglycoproteins↑ LDL↓ VLDL↓ Isoleucine↓ Leucine↓ Valine↓ Acetoacetate↓ Choline↓ (all in serum) | [151] |
| Human | Alcohol | Cirrhosis (mild vs. severe liver failure) | 1H NMR | Correlated with severity of liver failure: 3-Hydroxybutyrate↑ Alanine↑ Acetate↑ Choline/Phosphocholine↑ (all in serum) | [152] |
| Human | Chronic hepatitis B | Cirrhosis (compensated vs. decompensated) | 1H NMR | Distinguishing between compensated and decompensated cirrhosis: Succinate, Pyruvate, Phenylalanine, Histidine, Lysine, Glutamine, Acetone, Glutamate, Creatine, Alanine (all in serum) | [153] |
| Human | Causes not stated | Cirrhosis | 1H NMR | Positively correlated with portal blood proinflammatory cytokines IL6, TNFα and IL1β: Trimethylamine Negatively correlated with portal blood proinflammatory cytokines IL6, TNFα and IL1β: Acetate, n-Heptanoate Positively correlated with WBC and platelet counts: Threonine, α-Galactose, β-Glucose (all in feces) | [156] |
| Human | Various liver injuries | Cirrhosis (compensated vs. decompensated) | UPLC-QTOFMS | Lower in LC: N6-Methyladenosine, 1-Methyluric acid, Cinnamic acid, Decenoylcarnitine, Phenacetylglutamine (all in urine) | [158] |
| Human | Various etiologies, incl. HBV, HCV, alcohol, NASH | Cirrhosis US vs. Turkish (TR) population (dietary) | 1H NMR | Lactate (Controls and Decompensated; TR>US), Glucose (Controls and Decompensated; US>TR ) (all in plasma) | [159] |
| Human | Alcohol | Acute-on-chronic liver failure (ACLF) vs. stable compensated or decompensated cirrhosis (CLF) | 1H NMR | ACLF > CLF: 3-Hydroxybutyrate, Lactate, Acetoacetate, Pyruvate, Glutamine, Glutamate, Creatinine, Tyrosine, Phenylalanine (all in serum) | [160] |
| Human | Causes not stated | Stable cirrhosis (C) ± encephalopathy (E) (C±E) and HV | 1H NMR | C±E > HV: Lactate, Pyruvate, Alanine, Threonine, Glycine, Aspartate, Acetoacetate, 3-Hydroxybutyrate, Phenylalanine, Tyrosine, Methionine, Glutamate, Methylamine, Dimethylamine, TMAO, Glycerol C±E < HV: Valine, Glutamine, Histidine, Arginine E > HV: Leucine, Isoleucine C > HV: Myoinositol (all in plasma) | [161] |
| Human | Alcohol, HBV or HCV, NASH | Hepatic encephalopathy (HE), cirrhosis (C), neurological patients without liver disease (NP), HV | UPLC-Orbitrap MS | HE > NP: 13x N-Acetyl metabolites, 5x Glutamate/Glutamine metabolites, 4x Methionine metabolites, 4x Phenylalanine metabolites, 6x Tryptophan metabolites, 6x Fatty acid metabolites, Pyruvate, 5x Amino acid derivatives, 2x Dipeptides, 3x Bile acids, 3x Nucleoside derivatives, Dihydrothymine, 4x Alcohols and polyols, Ribitol/Arabitol, Cortisol, Pyridoxic acid, Phenyl sulfate HE < NP: Alanine, Taurine, Anhydro sorbitol, Levulinic acid (both in CSF and plasma) HE > C: 9x N-Acetyl metabolites, Phenacetylglutamine, 2x Methionine metabolites, 2x Phenylalanine metabolites, 3x Tryptophan metabolites, 4x Fatty acid metabolites, Citrulline, 2x Dipeptides, Taurocholic acid, 3x Nucleosides and derivatives, Anhydro sorbitol, 2x Alcohols, Ribitol/Arabitol, Cortisol, Phenyl sulfate HE < C: Methionine sulfoxide, Levulinic acid (all in plasma) | [162] |
| Human | Chronic hepatitis C | Fibrosis (F4 vs. F0) | 1H NMR | F4 vs. F0: VLDL↑ Citrate↑ Glucose↑ Phenylalanine↑ LDL↓ HDL↓ Choline↓ Acetoacetate↓ Isoleucine/Leucine↓ Creatinine/Creatine↓ Glutamate↓ Glutamine↓ Asparagine↓ Valine↓ Lysine↓ Cysteine↓ Glycerol↓ Arginine↓ Histidine↓ 3-Hydroxybutyrate↓ (all in serum) | [163] |
| Rat | TAA | Fibrosis/Cirrhosis vs. controls | 1H NMR | 3-Hydroxybutyrate↑ Acetoacetate↑ Butyrate↑ Choline↑ Glycine↑ Alanine↑ Leucine↑ Lysine↑ Succinate↑ Valine↑ 2-Oxoglutarate↓ Acetate↓ Adipate↓ Dimethylglycine↓ Lactate↓ Pyruvate↓ TMAO↓ Tyrosine↓ (all in serum) 1-Methylhistidine↑ 3-Hydroxybutyrate↑ Acetate↑ Alanine↑ Butyrate↑ Choline↑ Creatinine↑ Hippurate↑ Isoleucine↑ Pyruvate↑ Succinate↑ Taurine↑ TMAO↑ Tryptophan↑ Valine↑ 2-Hydroxybutyrate↓ 2-Oxoglutarate↓ Acetoacetate↓ Acetone↓ Adipate↓ Citrate↓ Dimethylamine↓ Dimethylglycine↓ Fumarate↓ Methylamine↓ Oxaloacetate↓ Sarcosine↓ Trimethylamine↓ (all in urine) | [165] |
| Rat | Dimethylnitrosamine | Fibrosis | UPLC-QTOFMS | LPC(18:1) ↑ LPC(18:2) ↑ LPC(20:4)↓ FA(22:6)↓ FA(20:4)↓ FA(18:1)↓ FA(18:2)↓ (all in serum) | [166] |
| Rat | Dimethylnitrosamine | Fibrosis | UPLC-QTOFMS | Cholic acid↑ Deoxycholic acid↑ Ursodeoxycholic acid↑ Chenodeoxycholic acid↑ Hyodeoxycholic acid↑ Lithocholic acid↑ Taurocholic acid↑ Taurodeoxycholic acid↑ Tauroursodeoxycholic acid↑ Taurochenodeoxycholic acid↑ Taurohyodeoxycholic acid↑ Taurolithocholic acid↑ Glycocholic acid↑ Glycodeoxycholic acid↑ Glycoursodeoxycholic acid↑ Glycochenodeoxycholic acid↑ (all in serum) | [167] |
| Rat | CCl4 | Fibrosis | UPLC-QTOFMS | Cervonoyl ethanolamide↑ β-Muricholic acid↑ (all in serum) | [168] |
| Rat | CCl4 | Decompensated cirrhosis/ascites | UPLC-Orbitrap MS | Alanine↑ Phenylalanine↑ Tryptophan↑ Tyrosine↑ Nutriacholic acid↑ LPC(16:0)↓ LPC(18:0)↓ LPC(18:2)↓ FA(20:5)↓ Carnitine↓ Creatine↓ Valine↓ Isoleucine↓ Arginine↓ (all in serum) Glutamyltaurine↑ 4,6-Dihydroxyquinoline↑ Phenylalanine↑ TMAO↑ 3-Methyldioxyindole↑ 1,2,3-Trihydroxybenzene↑ Tryptophan↑ Histamine↑ Tyrosine↑ Pantothenic acid↑ 2-Phenylglycine↑ Proline↑ N6,N6,N6-Trimethyllysine↑ Dopamine↑ Phenacetylglycine↓ Creatinine↓ Creatine↓ 4-Acetamidobutanoate↓ Indole↓ Carnitine↓ (all in urine) | [169] |
| Rat | CCl4 | Fibrosis | UPLC-QTOFMS | 12-Ketochenodeoxycholic acid↑ PI(18:0/16:0) ↑ Cervonoyl ethanolamide↑ LPC(18:2)↑ LPC(22:6)↑ PC(18:1/16:0)↑ PC(18:2/16:0)↑ PC(20:4/18:2)↑ PC(22:6/18:1)↑ Creatine↓ Sphinganine↓ Dihydroceramide↓ 8-HETE↓ LPC(18:0)↓ LPC(20:1)↓ LPC(22:0)↓ (all in serum) | [170] |
| Rat | Dimethylnitrosamine | Fibrosis | UPLC-Orbitrap MS | Leucine↓ LPC(16:0)↓ LPC(16:0)↓ LPC(18:0)↓ LPC(20:1)↓ LPC(20:4)↓ LPC(22:6)↓ FA(16:0)↓ FA(18:0)↓ FA(20:4)↓ FA(20:5)↓ FA(22:6)↓ All-trans-retinoic acid↓ Bilirubin↓ (all in serum) | [172] |
| Rat | CCl4 | Fibrosis | 1H NMR | 2-Oxoglutarate↓ Citrate↓ Dimethylamine↓ Creatinine↓ Phenacetylglycine↓ Hippurate↓ Taurine↑ (all in urine) | [123] |
| Rat | CCl4 | Fibrosis | 1H NMR | Glucose↓ Lactate↑ Fumarate↓ NADPH↓ Succinate↑ Acetate↑ 3-Hydroxybutyrate↓ UDP-glucose↑ UDP-galactose↑ (in serum and liver) | [125] |
| Species | Manipulation/Condition | Pathology | Analytical Methodology | Metabolites Reported | Ref. |
|---|---|---|---|---|---|
| Human | Obesity/metabolic syndrome | NAFL and NASH NAFL→NASH | HPLC-TQMS | FA(14:0)↑ FA(16:0)↑ FA(14:1n5)↑ FA(16:1n7)↑ FA(18:2n6)↓ FA(18:3n6)↑ FA(20:3n6)↑ FA(22:6n3)/FA(22:5n3) in PC and PE pools↓ 5-HETE↑ 8-HETE↑ 15-HETE↑ (all in plasma) | [178] |
| Human | Nondiabetic. NAFLD confirmed by liver biopsy | NAFLD vs. HV | UPLC-TQMS GC-MS | Glycocholate↑ Taurocholate↑ Glycochenodeoxycholate↑ Homocysteine↑ Cysteine↑ GSH↓ Glutamylvaline↑ γ-Glutamylleucine↑ γ-Glutamylphenylalanine↑ γ-Glutamyltyrosine↑ Cysteine-glutathione-disulfide↓ Carnitine↑ Propionylcarnitine↑ 2-Methylbutanoylcarnitine↑ Butanoylcarnitine↑ Tyrosine↑ Glutamate↑ Isoleucine↑ Leucine↑ Valine↑ Taurocholate↑ (all in plasma) | [180] |
| Human | Liver samples from normal (17), steatosis (4), NASH (fatty) (14) and NASH (not fatty) (23) | NAFL→NASH | UPLC Orbitrap-MS | Taurocholate↑ Taurodeoxycholate↑ Glycochenodeoxycholate↑ Taurine↑ Cholic acid↓ Glycodeoxycholate↓ (all in liver) Gene expression data consistent with the above (CYP7B1↑) | [222] |
| Human | Dietary intervention study, unrelated healthy surgical liver samples | NAFL (20 insulin-resistant/20 insulin sensitive) vs. control | UPLC-TQMS GC-MS | Insulin-resistant NAFL vs. insulin-sensitive NAFL: Total LPCs↓ LPC(16:0)↓ (all in plasma) | [182] |
| Human | NASH and healthy subjects given high-fat meal to stimulate gall bladder contraction | Fasting and postprandial serum from NASH and healthy subjects | UPLC-TQMS | NASH vs. control (preprandial): Total BAs↑ Glyco-BAs↑ Tauro-BAs↑ NASH vs. control (postprandial): Mainly Total BAs↑ Glyco-BAs↑ (all in serum) | [183] |
| Human | Normal, Steatosis, NASH with steatosis, NASH without steatosis livers | Normal, NAFL, fatty NASH, nonfatty-NASH | UPLC Orbitrap-MS | Control→NAFL: Acetyl carnitine↑ Lauroyl carnitine↑ Butanoyl carnitine↑ Palmitoyl carnitine↑ Hexanoyl carnitine↓ Valine↓ NAFL→NASH: Leucine↑ Isoleucine ↑ Tyrosine↑ Valine↑ Phenylalanine↑ (all in liver) | [184] |
| Human | Biopsy-proven NAFL, biopsy-proven NASH and normal controls with MRI fat fraction <5% | Normal, NAFL and NASH | UPLC-QTRAP MS/MS | NAFL→NASH: PGE2↑ 13,14-dihydro-15-keto-PGD2↑ 11,12-diHETrE↑ 14,15-diHETrE↑ 15-HETE↓ [all AA-derived] (all in plasma) | [185] |
| Human | Obese normal liver, obese NAFL and obese NASH | Normal, NAFL and NASH | 1H NMR | LDL-cholesterol↑ Alanine↑ Histidine↑ Phenylalanine↑ Tyrosine↑ Leucine↑ Free fatty acids↓ Citrate↓ 3-Hydroxybutyrate↓ Acetoacetate↓ (all in serum) | [186] |
| Human | Morbid obesity with and without NAFL | Obesity without NAFL, mild NAFL, moderate NAFL, severe NAFL | UPLC-LITMS GC-TOFMS Metabolon, Inc. | α-Ketoglutarate principal plasma marker with AUROC of 0.743, sensitivity of 80%, specificity of 62.5%. (all in plasma) | [187] |
| Human Mouse | Liver biopsies from patients with normal liver and NAFLD HFD, HFD + nicotinic acid, HFD + hydroquinone, HFD + tert-butylhydroquinone | NASH vs. NAFL vs. control | GC-MS | Nicotinic acid and hydroquinone negatively correlated with steatosis (NAS) score. Nicotinic acid supplementation of HFD prevented fat accumulation and improved serum ALT. (all in liver) | [204] |
| Human | NAFLD, NAFLD + T2DM, control, evaluated by ultrasound | NAFLD, NAFLD + T2DM, control | UPLC-QTOFMS | NAFLD vs. control: Proline↑ Phenylalanine↑ Oleamide↑ Bilirubin↑ Palmitoyl carnitine↑ LPC(20:5)↑ Lyso-PAF C-18↓ NAFLD + T2DM vs. control: Leucine↑ Oleamide↑ LPC(14:0)↑ Bilirubin↑ Tetradecenoyl carnitine↑ Linoleoyl carnitine↑ Tetradecadienoyl carnitine↑ (all in serum) | [206] |
| Human | Hyperuricemia (HU), HU+NAFLD, HU progressed to HU+NAFLD, healthy controls | HU, initial HU+NAFLD, initial HU→outcome HU+NAFLD, healthy controls | UPLC-QTOFMS | HU vs. control: Phosphatidic acid↑ 3,4-Dihydroxyphenylglycol↑ Valine↑ CE(18:0)↑ Uric acid↑ Acetyl carnitine↑ Inosine↓ 5-Hydroxyindoleacetic acid↓ 5-Aminoimidazole ribotide↓ Pyrrolidonecarboxylic acid↓ Glycerophosphocholine↓ HU vs. outcome HU+NAFLD: Phosphatidic acid↑ Inosinic acid↑ Tryptophan↑ Valine↑ Alanine↑ Lactate↑ CE(18:0)↑ Uric acid↑ Trimethylamine↑ Acetyl carnitine↑ 5-Methoxyindoleacetic acid↑ Acetoin↑ Inosine↓ Kynurenine↓ 5-Hydroxyindoleacetic acid↓ Pyrrolidonecarboxylic acid↓ 4-Fumarylacetoacetate↓ Pregnenolone sulfate↓ (all in serum) | [203] |
| Human | Bariatric surgery patients with wedge liver biopsy during surgery classified histologically as non-NASH, non-NAFLD, NAFL and NASH. PNPLA3 I148M variant also determined (more common in NASH). Discovery and validation cohorts used. | non-NASH vs. non-NAFLD vs. NAFL vs. NASH | UPLC-QTOFMS 2D-GC-TOFMS | Strong negative correlation between number of TG double bonds to TG concentrations in NASH relative to non-NASH for both discovery and validation cohorts. A “NASH ClinLipMet score” was developed based upon (i) clinical variables, (ii) PNPLA3 genotype, (iii) lipidomic data and (iv) metabolomic data. This was highest performing combination biomarker with sensitivity of 85.5% and specificity of 72.1% for NASH. (all in liver) | [188] |
| Human | Normal liver, NAFL liver, NASH liver | NASH vs. NAFLD vs. control lipidomics | UPLC-TQMS GC-MS | Thirty-two lipids discriminated NASH with 100% sensitivity and specificity. Accumulated hepatotoxic lipids in NASH included FA(14:0), FA(16:0), FA(16:1n-7), FA(18:1n-7) and FA(18:1n-9). Reduced in NASH: FA(20:4n-6), FA(20:5n-3), FA(22:6n-3), total CER, total SM, total PI, total PS, total PE, total PC. (all in liver) | [189] |
| Human | Nondiabetic NAFL patients with normal liver function, NASH with abnormal liver function, healthy controls. | NASH vs. NAFL vs. control urines. | LC-TQMS | NASH vs. control: Lysine↑ Valine↑ Citrulline↑ Arginine↑ Threonine↑ Tyrosine↑ Leucine↑ Hippurate↑ 3-Indoleacetate↑ 5-Hydroxyindoleacetate↓ 3-Indoleformate↓ Cortisol↓ NASH vs. NAFL: Methyl xanthine↑ Tryptophan↑ 3-Indoleacetate↑ Gluconate↑ Proline↓ (all in urine) | [190] |
| Human | Several large clinical cohorts with CT-defined liver fat plus NASH patients. | NASH vs. controls | UPLC-Q-Orbitrap-MS | Top metabolite correlated with liver fat was 202.1185+, which produced 24 hits in HMDB. Dimethylguanidino valeric acid (DMGV) chosen on basis of GWAS, which found SNPs for AGXT2 that produces DMGV. (all in plasma) | [191] |
| Human | NAFLD criteria met/not met at baseline, after dietary manipulation. | Non-NAFLD, Non-NAFLD→NAFLD, NAFLD→Non-NAFLD | UPLC-QTOFMS | Phospholipid and sphingolipid changes not of great statistical significance. Also lipid groups, not individual lipids, given only. (all in serum) | [241] |
| Human | NAFL and NASH based upon liver biopsy, healthy controls. | NASH vs. NAFL vs. controls | HPLC-Orbitrap-MS | Five metabolites increased control→NAFL→NASH – Uracil, α-Linolenic acid (all-cis-9,12,15-octadecatrienoic acid), Glutamate, Glutamine and 5-Oxoproline, which was chosen as a biomarker with a better AUROC for NASH vs. NAFL, than adiponectin, TNF-α, or IL-8. (all in serum) | [192] |
| Human | NAFL and NASH confirmed by liver biopsy | NASH vs. NAFL | High-field 1H MRS and ultra-high-field 31P MRS (in vivo) | Many MRS alterations correlated with NAFL→NASH, mostly with advanced fibrosis, e.g. phosphoethanolamine/total phosphorus (TP) ratio. ATP/TP↓ in advanced fibrosis and ATP flux↓ in NASH | [155] |
| Human | Chronic hepatitis B (CHB) with biopsy-proven NAFLD and without NAFLD, healthy controls | CHB +NAFLD vs. CHB-NAFLD vs. controls | UPLC-QTOFMS | Most neutral lipids and ceramides were elevated in CHB+NAFLD but decreased in CHB-NAFLD vs. healthy controls. Monounsaturated TGs were a good predictor of NASH, superior to cytokeratin-18 or ALT. | [196] |
| Human | Hepatic steatosis in morbidly obese women | Metagenomic signature of hepatic steatosis | 1H NMR (urine and plasma) UPLC-TQMS (plasma) | Microbiota metabolite produced from phenylalanine, phenylacetic acid (PAA) associated with steatosis. | [197] |
| Human | Biopsy-proven subjects with normal liver (NL), NAFL and NASH | NASH vs. NAFL vs. NL discovery and validation cohorts | UPLC-QTOFMS | Triglycerides are elevated NAFL > NL ≥ NASH. Of the 28 TGs measured, TG(46:0), (48:0), (53:0), (44:1), (48:1), (49:1), (52:1), (53:1), (50:2), (54:5) and (58:2) were always NAFL > NL and NASH < NAFL. (all in serum) | [198] |
| Human | Large study of 769 nondiabetic patients with liver fat content measured by MRI and correlated with metabolite profiles of urine and fasting plasma | 613 plasma and 587 urine samples across a range of liver pathologies (34.7% with steatosis) | UPLC-LITMS 1H NMR | Associations in plasma with LFC: BCAAs↑ Aromatic amino acids↑ Dipeptides↑ Proline↑ Tryptophan↑ Indoleacetate↑ Urate↑ Piperine↑ 7α-Hydroxy-3-oxo-cholestenoate↑ Ether-PCs↓ 3-Phenylpropionate↓ Proline betaine↓ Associations in urine with LFC: BCAA derivatives↑ Lactate↑ Isovalerylglycine↓ Isobutyrylglycine↓ γ-Glutamylthreonine↓ 4-Vinylphenol sulfate↓ Hippurate↓ Cinnamoylglycine↓ | [200] |
| Human | NAFLD determined by hepatic ultrasound | BMI < 25 vs. BMI > 30 vs. BMI > 30 with NAFLD | TQMS for 31 acyl carnitines and 7 amino acids | Family history predicted obesity correlating with amino acids that contributed to an increase in specific acyl carnitines. Excess FFAs related to obesity were associated with NAFLD. | [201] |
| Human | Patients with normal fasting glucose. Visceral adipose tissue (VAT) assessed by MRI. Hepatic TG content (HTGC) determined by proton-MR spectroscopy. | Range of VAT and HTGC | ESI-FIA-MS/MS | Associated with HTGC: LPC(14:0), PC(28:1), PC(30:0), PC(32:1), PC(32:2), PC(34:1), PC(34:3), PC(34:4), PC(36:1), PC(36:2), PC(36:3), PC(36:6), PC(38:3), PC(38:5), PC(40:4), PC(40:5), SM(22:3), Tryptophan, Tyrosine (all in plasma) | [202] |
| Human | NAFL and NASH determined by liver biopsy and healthy controls by ultrasound and liver enzymes | NASH vs. NAFL vs. controls | UPLC-Orbitrap-MS GC | Lipid group trends: DG: NASH > NAFL > healthy PG: NASH ≈ NAFL > healthy PA: NASH ≈ NAFL > healthy AcCa: NASH < NAFL ≈ healthy CE: NASH < NAFL < healthy LPC: NASH < NAFL ≈ healthy SM: NASH < NAFL ≈ healthy FA(16:0): NASH > NAFL > healthy FA(16:1n-7cis): NASH > NAFL > healthy FA(18:1n-9cis): NASH > NAFL > healthy FA(18:2n-6): NASH < NAFL < healthy FA(20:4n-6): NASH < NAFL < healthy | [199] |
| Human | Children, overweight or obese, with or without clinical/radiological signs of NAFLD | NAFLD vs. control | Selective ion flow tube mass spectrometry (SIFT-MS) | Acetaldehyde↑ Acetone↑ Isoprene↑ Pentane↑ Trimethylamine↑ (all in breath) | [209] |
| Human | Children with biopsy-proven NAFLD and matched healthy controls | NAFLD vs. control | HPLC | Homocysteine↑ Cysteine↑ CysGly↑ GSH↓ (all in plasma) | [211] |
| Human | Children with obesity and NAFL confirmed by MRS and matched obese controls | NAFL vs. control | UPLC-Q-Orbitrap-MS | Tyrosine↑ Glutamate↑ Octanoic acid↑ Linoleic acid↓ (all in plasma) | [212] |
| Human | Children with obesity, NAFL, NASH and healthy controls | NASH vs. NAFL vs. control | GC-MS | 1-Butanol↑ (in NAFL) 1-Pentanol↑ (in NAFL) ↓ (in NASH) Phenol↑ (in NAFL) 2-Butanone↑ (in NAFL and NASH) 4-Methyl-2-pentanone↓ (in NAFL)↑ (in NASH) (all in feces) Metagenomics also conducted. Correlations with NAFLD and certain VOCs reported | [242] |
| Human | Children with NAFLD, with or without obesity | Obese − NAFL, obese + NAFL, normal weight healthy controls | GC-MS | NAFL vs. control: Glucose↑ 1-Methylhistidine↑ Pseudouridine↑ Glycolic acid↑ Mannose↓ p-Cresol sulfate↓ Kynurenine↓ Hydroquinone↓ Adipate↓ Phenylacetic acid↓ Small intestine bacterial overgrowth (SIBO): Glycolic acid↑ Mannose↑ Valine↓ p-Cresol sulfate↓ Butanoate↓ Adipate↓ (all in urine) | [213] |
| Human | Children with obesity, with and without NAFLD assessed by MRI | Obese + NAFLD vs. obese − NAFLD | UPLC-QTRAP-MS | Isoleucine↑ Leucine↑ Valine↑ C4-carnitine↑ C5-carnitine↑ C14:1-OH-carnitine↑ Tryptophan↑ Lysine↑ Glutamate↑ PC(32:1)↑ (all in plasma) | [214] |
| Human | Children with obesity, with and without NAFL assessed by ultrasound (US), with and without metabolic syndrome (MetS) and nonobese controls | Obese + NAFL vs. obese − NAFL Obese + MetS vs. obese − MetS | GC-MS | Obese − NAFL vs. controls: Palmitate↑ Myristate↑ Urea↑ N-Acetylgalactosamine↑ Maltose↑ Gluconate↑ Isoleucine↑ Hydroxybutanoate↓ Malate↓ Obese + NAFL vs. controls: Laurate↑ Maltose↑ (all in saliva) | [215] |
| Human | Adolescents with NAFLD assessed by US or ALT/AST | US vs. ALT vs. AST diagnostic methods | Biochemical lipid analysis 1H NMR | Many differences in lipid profiles, amino acids (alanine, glutamine, histidine; BCAAs; aromatic amino acids) and ketone bodies (acetate, acetoacetate, β-hydroxybutyrate) (all in plasma) | [216] |
| Human | Adolescents with obesity and with or without NAFLD confirmed by MRI | NAFLD vs. non-NAFLD | UPLC-Q-Orbitrap-MS | Leucine/Isoleucine↑ Tryptophan↑ Serine↓ Dihydrothymine↓ LPE(20:0)↓ LPC(18:1)↓ (all in plasma) | [217] |
| Human | Children with or without NAFLD confirmed by ultrasound and liver enzymes | NAFLD vs. non-NAFLD | GC-MS | 24-h Urinary steroid profiles: Cortisol (obese controls)↑ Tetrahydrocortisone (NAFLD)↑ Overall data pointed to 5α-reductase↑, 21-hydroxylase↑ and 11β-hydroxysteroid dehydrogenase 1↓ | [243] |
| Human Mouse | Morbidly obese, nondiabetic Gnmt-null vs. WT | NAFL→NASH NASH vs. control | UPLC-QTOFMS | PC(14:0/20:4)↑ LPC(18:1)↑ PC(P-24:0/0:0)↓ PC(P-22:0/0:0)↓ PC(O-20:0/0:0)↓ FA(20:4)↓ Glutamate↓ (all in serum) Results consistent with human studies | [179] |
| Mouse Human | Methionine and choline deficient diet (MCD) HBV-negative, NAFLD confirmed by liver biopsy | NASH vs. NAFL vs. control | 1H NMR | Glucose↑ Lactate↑ Glutamate↑ Taurine↑ TG↑ Total cholesterol↑ LDL cholesterol↑ Glucose↑ Lactate↑ Glutamate↑ Taurine↑ (all in serum) | [181] |
| Mouse Human | Mat1a-KO vs. WT mouse liver and serum metabolome NAFL and NASH discovery and validation cohorts | Mat1a-KO vs. WT Clustering analysis into M-subtype and Non-M-subtype based upon mouse metabolomes | UPLC-QTOFMS | M-subtype NASH biomarkers: Amino acids (5), Fatty acyls (8), Triglycerides (3), Glycerophospholipids (37), Sphingomyelins (1) Non-M-subtype NASH biomarkers: Amino acids (1), Fatty acids (1), Bile acids (1), Triglycerides (3) M-subtype patients: 34% NASH Non-M-subtype patients: 39% NASH | [205] |
| Mouse | MCD | NASH vs. control | UPLC-QTOFMS | Tauro-β-muricholate↑ Taurocholate↑ 12-HETE↑ LPC(16:0)↓ LPC(18:0)↓ LPC(18:1)↓ (all in serum) | [218] |
| Mouse | MCD vs. choline-supplemented MCD (MCS) | Differential effects of methionine and choline deficiency | UPLC-QTOFMS | MCD vs MCS: Oleic acid↑ Linoleic acid↑ Total nonesterified fatty acids↑ (all in serum) | [219] |
| Mouse | NASH-inducing diet (35% lard, 1.25% cholesterol, 0.5% sodium cholate) | NAFLD vs. control | HPLC-TQMS | Glycerol↑ Free cholesterol↑ Esterified cholesterol↑ Putrescine↑ N8-Acetylspermidine↓ Spermine↓ Adenine↓ Adenosine↓ Homocysteine↓ Methylthioadenosine↓ S-Adenosylhomocysteine↓ S-Adenosylmethionine↓ Proteomic findings were consistent with the above (all in liver) | [220] |
| Mouse | Ldlr-null mice fed a Western diet (energy as 17% protein, 43% carbohydrate, 41% fat, 0.2% cholesterol) + olive oil (WD + O) | NAFLD/NASH vs. control | UPLC-LITMS GC-MS Metabolon, Inc. | Saturated fatty acids↑ MUFAs↑ Palmitoyl-sphingomyelin↑ Cholesterol↑ n-6 PUFA↑ 12-HETE↑ C20-22 n-3 PUFA-containing phosphoglycerolipids↓ 18-HEPE↓ 17,18-diHETE↓ S-Lactoyl-glutathione↓ (all in liver) F3-Isoprostanes↓ (in urine) | [221] |
| Mouse | HFD (60% calories from fat) and normal chow (12.7% calories from fat) | NAFLD vs. control | 1H NMR | Glucose↑ Total cholesterol↑ HDL-cholesterol↑ AST↑ ALT↑ Phosphatidylcholine↑ Pyruvate↓ Acetate↓ Lactate↓ Citrate↓ Arginine↓ Ornithine↓ Acetoacetate↓ 3-Hydroxybutyrate↓ Isoleucine↓ Leucine↓ Valine↓ Glutamate↓ Glutamine↓ Tyrosine↓ Phenylalanine↓ Alanine↓ Lysine↓ Glycine↓ Betaine↓ Isobutanoate↓ 1-Methylhistidine↓ (all in serum) Total cholesterol↑ Triglycerides↑ Fatty acids↑ PUFA/MUFA↓ (all in liver) Pyruvate↑ Creatinine↑ Taurine↑ Glycine↑ Formate↑ Butanoate↑ Guanidinoacetate↑ Glucose↑ 1-Methylnicotinamide↑ Nicotinamide N-oxide↑ Acetoacetate↓ Succinate↓ Citrate↓ 2-Oxoglutarate↓ Trimethylamine↓ Trans-aconitate↓ Hippurate↓ Trigonelline↓ Niacinamide↓ Tyrosine↓ 1-Methylhistidine↓ Phenylalanine↓ (all in urine) | [223] |
| Mouse | HFD (60% calories from fat) and normal chow (13.5% calories from fat) | NAFLD vs. control | UPLC-QTOFMS GC-MS | Methylhippurate↑ Glycerol 3-phosphate↑ Mannose↑ Ketoleucine↑ 2-Ketohexanoate↑ Hydroxyphenyllactate↑ Succinate↑ Xylose/Ribose/Arabinose↓ Glucuronate↓ Catechol↓ 4-Coumarate↓ Hippurate↓ Taurocholate↓ Glycochenodeoxycholate↓ Glycocholate↓ Histamine↓ (all in serum) | [224] |
| Mouse | A/J, C57BL/6J and PWD/PhJ strains fed standard diet with 0.1% DDC | DDC-treated vs. control | UPLC-Q-LITMS | Putrescine↑ Arginine↑ Citrulline↑ cAMP↑ 2-Oxoglutarate↑ Asparagine↑ Glutamate↑ (all in liver) | [227] |
| Mouse | HFD-fed mice (42% calories from fat, 43% from carbohydrates, 15% from protein) vs. standard chow (17% from fat, 58% from carbohydrates, 25% from protein) | NAFLD vs. control | UPLC-Q-LITMS | SFA-DAGs↑ MUFA-DAGs↑ PUFA-DAGs↓ SFA-CEs↑ MUFA-CEs↑ PAs↑ PGs↑ SFA-CERs↑ Sphingosine↑ Sphingosine-1-phosphate↑ Dihydrosphingosine↑ Dihydrosphingosine-1-phosphate↑ Galactosylceramide↓ Glucosylceramide↓ Lactosylceramide↑ Globotrioseacylceramide↑ TxB2↑ PGF2α↑ All other eicosanoids↓ Pattern changed from weeks 16-52. (all in liver) | [228] |
| Mouse | db/db leptin receptor-deficient mice with insulin resistance and steatosis subjected to caloric restriction (CR), db/m mice without insulin resistance and steatosis | db/db, pre- and post-caloric restriction, db/m | 1H NMR UPLC-QTOFMS | db/db vs. db/m: Acetone↑ 3-Hydroxybutyrate↑ Lactate↑ Acetate↑ Glutathione↑ Ascorbate↑ Many glycerolipids↑ db/db + CR vs. db/db: 3-Hydroxybutyrate↓ Ascorbate↓ Many glycerolipids↓ RT-PCR findings consistent with metabolomic data (all in liver) | [231] |
| Mouse | Leptin-deficient obese ob/ob mice and nonsteatotic ob/+ heterozygous mice | Intact liver tissues of two mouse lines compared | HR-MAS 1H NMR | Many lipid 1H signals highly statistically significantly elevated in steatotic ob/ob livers compared with nonsteatotic ob/+ livers, as expected. ob/ob livers vs. ob/+ livers: Betaine↑ Phenylalanine↑ Uridine↑ Creatinine↓ Glutamate↓ Glycine↓ Glycolate↓ Trimethylamine N-oxide↓ N,N-Dimethylglycine↓ ADP↓ AMP↓ | [232] |
| Mouse | High-trans-fat high-fructose diet (TFD) for 8 weeks (steatosis) and 24 weeks (NASH) | TFD-fed, normal diet-fed | Fasting hepatic mitochondrial flux by 13C NMR isotopomer analysis. LC-TQMS lipidomics | 8-week (steatosis) vs. 24-week (NASH): Endogenous glucose production↑ TCA cycle flux↑ Anaplerosis↑ Pyruvate cycling↑ Control vs. 8-week (steatosis): Total diacylglycerols↑ Total ceramides↑ C8-acyl carnitine↑ C16-acyl carnitine↑ 8-week (steatosis) vs. 24-week (NASH): DG(16:1/16:1)↑ DG(16:0/18:1)↑ DG(34:2)↑ DG(18:1/18:1)↑ DG(18:1/18:2)↑ DG(18:2/18:2)↑ DG(16:0/20:4)↑ DG(18:0/20:4)↑ DG(18:1/20:4)↑ DG(18:2/20:4)↑ DG(18:2/20:2)↑ DG(16:1/22:6)↑ DG(18:1/22:6)↑ C6-acyl carnitine↑ C8-acyl carnitine↑ C14-acyl carnitine↑ C16-acyl carnitine↑ CER(20:0)↓ CER(22:0)↓ (all in liver) | [229] |
| Mouse | High-fat, high-cholesterol, cholate (HFDCC)-fed mice with NAFLD without obesity | HFDCC vs. control | GC-TOFMS UPLC-QTOFMS | Total cholesterol↑ CE(16:1), (18:1), (18:2), (18:3), (20:1), (20:3), (20:4), (22:5), (22:6)↑ Cholic acid↑ DGs↑ TGs↑ CERs↑ SMs↑ LPCs↑ PC/PE↑ PEs↓ Xylitol↑ Xanthosine↑ Squalene↑ Phenylethylamine↑ Citrate↓ G-1-P↓ Saccharic acid↓ (all in liver) Total cholesterol↑ CE(16:1), (18:1), (18:2), (18:3), (20:1), (20:3), (20:4), (22:5)↑ Cholic acid↑ Deoxycholic acid↑ CERs↑ SMs↑ PEs↑ FFAs↓ Glycerol↓ TGs↓ LPEs↓ (all in plasma) | [225] |
| Rat | HFD-induced NASH | HFD vs. control diet | UPLC-QTOFMS | Glucose↑ Triglycerides↑ LDL-cholesterol↑ SM(36:1)↑ LPC(18:1)↑ LPC(20:2)↑ SM(34:2)↑ PC(34:1)↑ PC(38:4)↑ PC(38:3)↑ LPC(17:1)↑ PC(35:2)↑ FA(20:1)↑ FA(20:3)↑ FA(22:3)↑ Phytomonate↑ LPC(14:0)↑ 13-HpODE↑ PC(37:4)↓ PC(38:4)↓ PC(38:6)↓ SM(34:1)↓ SM(34:2)↓SM(42:3)↓ SM(40:1)↓ PC(40:5)↓ PC(40:6)↓ PC(40:8)↓ Creatine↓ Indoxyl sulfate↓ (all in serum) | [233] |
| Rat | HFD-induced NASH, positive controls (methionine plus choline supplementation), control diet | HFD vs. positive control vs. control diet | UPLC-QTOFMS | HFD vs. control: FA(28:8)↑ CE(12:0)↑ PG(14:0/18:1)↑ Cortisone↓ Antrosta-1,4-diene-3,17-dione↓ All-trans-retinoyl-β-glucuronide↓ LPA(18:2)↓ PE(15:0/22:2)↓ Cortol↓ 21-Hydroxypregnenolone↓ Cortolone↓ Urobilin↓ LPA(18:1)↓ PA(P-20:0/14:0)↓ (all in serum) | [234] |
| Rat Human | Rats fed HFD to lead to steatosis, rats fed MCD diet to lead to NASH, rats fed methionine and choline sufficient diet as controls→liver samples NASH (fatty), NASH (not fatty), steatosis, healthy liver samples | NASH vs. NAFL vs. control (rat and human) | UPLC Orbitrap-MS | Bile acid metabolomics: Significant BA profile differences between rat MCD and human NASH. Amino acid metabolomics: Asparagine↑ Citrulline↑ Lysine↑ comparable between rat MCD and human NASH. Fatty acid, carnitine and LPC metabolomics: Stearoyl carnitine↑ only lipid in both rat MCD and human NASH | [237] |
| Rat | HFD, MCD diet and streptozocin (STZ) in rats. Metabolomics and transcriptomics on serum and liver. | NAFL vs. NASH vs. NAFL + T2DM | UPLC-QTOFMS | Venn diagram for HFD, MCD and HFD+STZ serum: Stearoyl carnitine↑ (9E)-octadecenoyl carnitine↑ docosapentaenoic acid↑ vitamin D2↑ | [238] |
| Rat | HFD/cholesterol diet vs. normal diet | Stage of steatosis, inflammation and fibrosis determined histologically | GC | Correlation between liver and blood cell total fatty acids for control diet: FA(16:1), FA(22:6), FA(18:1n-7), FA(22:5) Correlation between liver and blood cell total fatty acids for HFD/cholesterol diet: FA(22:6), FA(18:1n-7) | [239] |
| Rat | HFD vs. normal diet | NAFLD established by histology and liver enzymes | UPLC-TQMS | BAs in liver: Taurocholate↑ Taurohyodeoxycholate↓ Ursodeoxycholate↓ BAs in caecal contents: Cholate↑ Hyodeoxycholate↓ Muricholate↓ BAs in serum: Taurocholate↑ Hyodeoxycholate↓ Taurohyodeoxycholate↓ | [240] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beyoğlu, D.; Idle, J.R. Metabolomic and Lipidomic Biomarkers for Premalignant Liver Disease Diagnosis and Therapy. Metabolites 2020, 10, 50. https://doi.org/10.3390/metabo10020050
Beyoğlu D, Idle JR. Metabolomic and Lipidomic Biomarkers for Premalignant Liver Disease Diagnosis and Therapy. Metabolites. 2020; 10(2):50. https://doi.org/10.3390/metabo10020050
Chicago/Turabian StyleBeyoğlu, Diren, and Jeffrey R. Idle. 2020. "Metabolomic and Lipidomic Biomarkers for Premalignant Liver Disease Diagnosis and Therapy" Metabolites 10, no. 2: 50. https://doi.org/10.3390/metabo10020050
APA StyleBeyoğlu, D., & Idle, J. R. (2020). Metabolomic and Lipidomic Biomarkers for Premalignant Liver Disease Diagnosis and Therapy. Metabolites, 10(2), 50. https://doi.org/10.3390/metabo10020050