
Comparative Analysis of Various Plant-Growth-Regulator Treatments on Biomass Accumulation, Bioactive Phytochemical Production, and Biological Activity of Solanum virginianum L. Callus Culture Extracts

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Seed Germination
2.3. Callus Culture Establishment
2.4. Extract Preparation
2.5. TPC Determination
2.6. TFC Determination
2.7. High-Performance Liquid Chromatographic Analysis
2.8. Cell-Free Antioxidant Assays
2.8.1. DPPH Assay
2.8.2. FRAP Assay
2.8.3. ABTS Assay
2.9. Antiaging Assay
2.9.1. Inhibition of AGE (Advanced Glycation End Product) Formation
2.9.2. Elastase-Inhibition Assay
2.9.3. Tyrosinase-Inhibition Assay
2.9.4. Collagenase-Inhibition Assay
2.9.5. Hyaluronidase Assay
2.10. Anti-Inflammatory Assays
2.10.1. COX-1- and COX-2-Inhibition Assays
2.10.2. 15-LOX-Inhibition Assay
2.10.3. sPLA2-Inhibition Assay
2.11. Statistical Analysis
3. Results and Discussion
3.1. Callogenesis
3.2. Biomass Accumulation
3.3. Total Phenolic and Total Flavonoid Contents
3.4. HPLC Analysis
3.5. Antioxidant Evaluation
3.6. Antiaging Evaluation
3.7. Anti-Inflammatory Evaluation
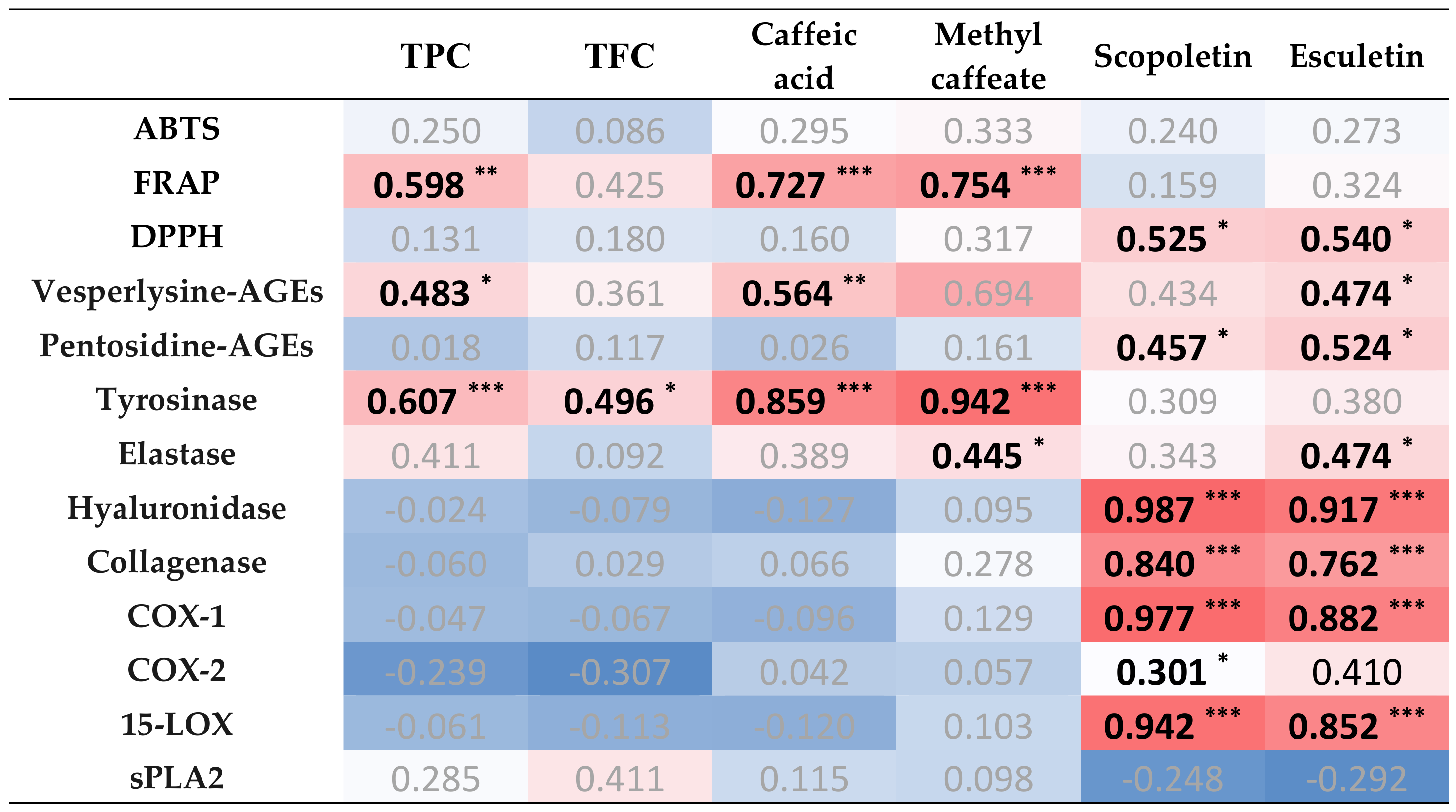
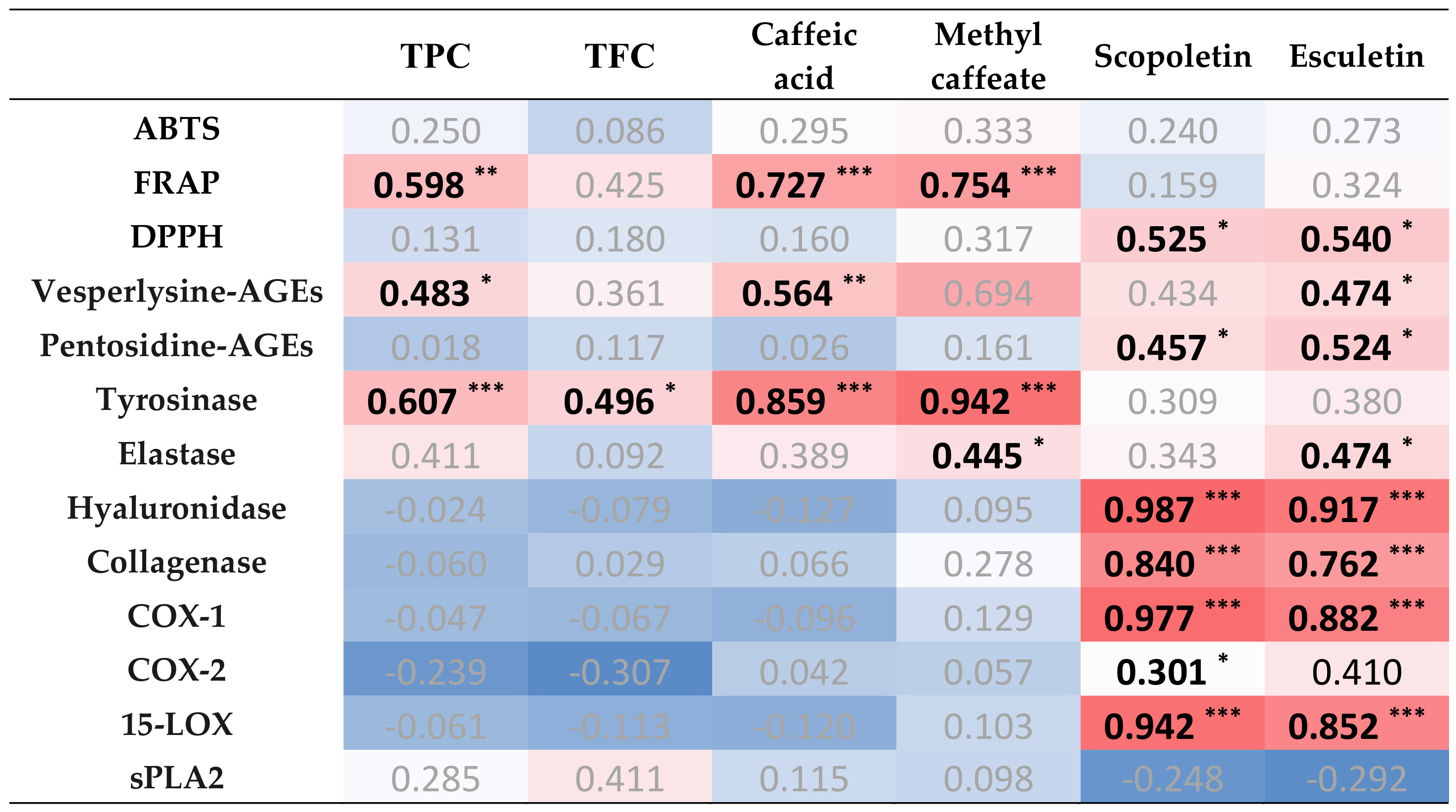
3.8. Correlation Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NAA | Naphthalene acetic acid |
| TDZ | Thidiazuron |
| BAP | Benzylaminopurine |
| MS | Murashige and Skoog |
| HPLC | High-performance liquid chromatography |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl |
| ABTS | Azinobis-3-Ethylbenzthiazoline-6-Sulphonic acid |
| FRAP | Ferric Reducing Antioxidant Potential |
| TPC | Total phenolic content |
| TPP | Total phenolic production |
| TFC | Total flavonoid content |
| TFP | Total flavonoid production |
| PGRs | Plant growth regulators |
| FW | Fresh weight |
| DW | Dry weight |
| PAL | Phenylalanine ammonia-lyase |
| AGE | Advanced glycation end products |
| sPLA2 | Phospholipase A2 |
| COX-1 | Cyclooxygenase-1 |
| COX-2 | Cyclooxygenase-2 |
| 15-LOX | Lipoxygenase |
| ROS | Reactive oxygen species |
| DMSO | Dimethyl sulfoxide |
| OD | Optical density |
References
- Li, F.S.; Weng, J.K. Demystifying traditional herbal medicine with modern approach. Nat. Plants 2017, 3, 17109. [Google Scholar] [CrossRef] [PubMed]
- Ansel, J.L.; Moretti, C.; Raharivelomanana, P.; Hano, C. Cosmetopoeia Foreword. Comptes Rendus. Chim. 2016, 19, 1033–1034. [Google Scholar] [CrossRef]
- Pandey, P.; Garg, A.; Shukla, A. Carpesterol—A novel phytosterol obtained from the plants of the family Solanaceae with evaluation of antineoplastic activity. J. Med. Pharm. Allied Sci. 2016, 1, 1–10. [Google Scholar]
- Pathiratna, L.; Joseph, K.; Perera, M. The effect of some cultural practices on the growth and yield of the medicinal plant Solanum virginianum Schrad. (katuwelbatu). Ceylon J. Sci. (Biol. Sci.) 2004, 32, 75–80. [Google Scholar]
- Rane, M.H.; Sahu, N.K.; Ajgoankar, S.S.; Teli, N.C.; Verma, D.R. A holistic approach on review of Solanum virginianum L. J. Pharm. Pharma Sci. 2014, 3, 1–4. [Google Scholar]
- Rita, P.; Animesh, D.K. An updated overview on Solanum xanthocarpum Schrad and Wendl. Int. J. Res. Ayurveda Pharm. 2011, 2, 730–735. [Google Scholar]
- Reddy, N.; Reddy, R. Solanum xanthocarpum chemical constituents and medicinal properties: A review. Sch. Acad. J. Pharm. 2014, 3, 146–149. [Google Scholar]
- Usman, H.; Ullah, M.A.; Jan, H.; Siddiquah, A.; Drouet, S.; Anjum, S.; Giglioli-Guviarc’h, N.; Hano, C.; Abbasi, B.H. Interactive Effects of wide-spectrum monochromatic lights on phytochemical production, antioxidant and biological activities of Solanum xanthocarpum callus cultures. Molecules 2020, 25, 2201. [Google Scholar] [CrossRef]
- Bourgaud, F.; Hehn, A.; Larbat, R.; Doerper, S.; Gontier, E.; Kellner, S.; Matern, U. Biosynthesis of coumarins in plants: A major pathway still to be unravelled for cytochrome P450 enzymes. Phytochem. Rev. 2006, 5, 293–308. [Google Scholar] [CrossRef]
- Karamat, F.; Olry, A.; Doerper, S.; Vialart, G.; Ullmann, P.; Werck-Reichhart, D.; Bourgaud, F.; Hehn, A. CYP98A22, a phenolic ester 3′-hydroxylase specialized in the synthesis of chlorogenic acid, as a new tool for enhancing the furanocoumarin concentration in Ruta graveolens. BMC Plant Biol. 2012, 12, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, G.R.; Ignacimuthu, S.; Paulraj, M.G.; Sasikumar, P. Antihyperglycemic activity and antidiabetic effect of methyl caffeate isolated from Solanum torvum Swartz. fruit in streptozotocin induced diabetic rats. Eur. J. Pharm. 2011, 670, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Hano, C.; Tungmunnithum, D. Plant Polyphenols, More than Just Simple Natural Antioxidants: Oxidative Stress, Aging and Age-Related Diseases. Medicines 2020, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- Tungmunnithum, D.; Hano, C. Cosmetic Potential of Cajanus cajan (L.) Millsp: Botanical Data, Traditional Uses, Phytochemistry and Biological Activities. Cosmetics 2020, 7, 84. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Kongsawadworakul, P.; Hano, C. A Cosmetic Perspective on the Antioxidant Flavonoids from Nymphaea lotus L. Cosmetics 2021, 8, 12. [Google Scholar] [CrossRef]
- Khan, T.; Frost, S. Floral biodiversity: A question of survival in the Indian Thar Desert. Environmentalist 2001, 21, 231–236. [Google Scholar] [CrossRef]
- Mazid, T.A.; Khan, F. Mohammad, Medicinal plants of rural India: A review of use by Indian folks. Indo Glob. J. Pharm. Sci. 2012, 2, 286–304. [Google Scholar]
- Davies, K.M.; Deroles, S.C. Prospects for the use of plant cell cultures in food biotechnology. Curr. Opin. Biotechnol. 2014, 26, 133–140. [Google Scholar] [CrossRef]
- Matkowski, A. Plant in vitro culture for the production of antioxidants—A review. Biotechnol. Adv. 2008, 26, 548–560. [Google Scholar] [CrossRef]
- Ahmad, N.; Fazal, H.; Abbasi, B.H.; Rashid, M.; Mahmood, T.; Fatima, N. Efficient regeneration and antioxidant potential in regenerated tissues of Piper nigrum L. Plant Cell Tissue Organ Cult. 2010, 102, 129–134. [Google Scholar] [CrossRef]
- Zahir, A.; Abbasi, B.H.; Adil, M.; Anjum, S.; Zia, M. Synergistic effects of drought stress and photoperiods on phenology and secondary metabolism of Silybum marianum. Appl. Biochem. Biotechnol. 2014, 174, 693–707. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Anjum, S.; Hano, C. Differential effects of in vitro cultures of Linum usitatissimum L. (Flax) on biosynthesis, stability, antibacterial and antileishmanial activities of zinc oxide nanoparticles: A mechanistic approach. RSC Adv. 2017, 7, 15931–15943. [Google Scholar] [CrossRef] [Green Version]
- Mozetič, B.; Trebše, P.; Hribar, J. Determination and quantitation of anthocyanins and hydroxycinnamic acids in different cultivars of sweet cherries (Prunus avium L.) from Nova Gorica region (Slovenia). Food Technol. Biotechnol. 2002, 40, 207–212. [Google Scholar]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagliazucchi, D.; Verzelloni, E.; Bertolini, D.; Conte, A. In vitro bio-accessibility and antioxidant activity of grape polyphenols. Food Chem. 2010, 120, 599–606. [Google Scholar] [CrossRef]
- Bose, S.; Munsch, T.; Lanoue, A.; Garros, L.; Tungmunnithum, D.; Messaili, S.; Giglioli-Guivarc’h, N. UPLC-HRMS analysis revealed the differential accumulation of antioxidant and anti-aging lignans and neolignans in in vitro cultures of Linum usitatissimum L. Front. Plant Sci. 2020, 11, 508658. [Google Scholar] [CrossRef] [PubMed]
- Kaewseejan, N.; Siriamornpun, S. Bioactive components and properties of ethanolic extract and its fractions from Gynura procumbens leaves. Ind. Crops Prod. 2015, 74, 271–278. [Google Scholar] [CrossRef]
- Wittenauer, J.; Mäckle, S.; Sußmann, D.; Schweiggert-Weisz, U.; Carle, R. Inhibitory effects of polyphenols from grape pomace extract on collagenase and elastase activity. Fitoterapia 2015, 101, 179–187. [Google Scholar] [CrossRef]
- Chai, W.M.; Huang, Q.; Lin, M.Z.; Ou-Yang, C.; Huang, W.Y.; Wang, Y.X.; Xu, K.L.; Feng, H.L. Condensed tannins from longan bark as inhibitor of tyrosinase: Structure, activity, and mechanism. J. Agric. Food Chem. 2018, 66, 908–917. [Google Scholar] [CrossRef]
- Kolakul, P.; Sripanidkulchai, B. Phytochemicals and anti-aging potentials of the extracts from Lagerstroemia speciosa and Lagerstroemia floribunda. Ind. Crops Prod. 2017, 109, 707–716. [Google Scholar] [CrossRef]
- Shah, M.; Ullah, M.A.; Drouet, S.; Younas, M.; Tungmunnithum, D.; Giglioli-Guivarc’h, N.; Hano, C.; Abbasi, B.H. Interactive Effects of Light and Melatonin on Biosynthesis of Silymarin and Anti-Inflammatory Potential in Callus Cultures of Silybum marianum (L.) Gaertn. Molecules 2019, 24, 1207. [Google Scholar] [CrossRef] [Green Version]
- Murthy, B.; Victor, J.; Singh, R.P.; Fletcher, R.; Saxena, P.K. In vitro regeneration of chickpea (Cicer arietinum L.): Stimulation of direct organogenesis and somatic embryogenesis by thidiazuron. Plant Growth Regul. 1996, 19, 233–240. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Siddiquah, A.; Tungmunnithum, D.; Bose, S.; Younas, M.; Garros, L.; Drouet, S.; Giglioli-Guivarc’h, N.; Hano, C. Isodon rugosus (Wall. ex Benth.) Codd In Vitro Cultures: Establishment, Phytochemical Characterization and In Vitro Antioxidant and Anti-Aging Activities. Int. J. Mol. Sci. 2019, 20, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, M.A.; Tungmunnithum, D.; Garros, L.; Hano, C.; Abbasi, B.H. Monochromatic lights-induced trends in antioxidant and antidiabetic polyphenol accumulation in in vitro callus cultures of Lepidium sativum L. J. Photochem. Photobiol. B 2019, 196, 111505. [Google Scholar] [CrossRef] [PubMed]
- Tariq, U.; Ali, M.; Abbasi, B.H. Morphogenic and biochemical variations under different spectral lights in callus cultures of Artemisia absinthium L. J. Photochem. Photobiol. B 2014, 130, 264–271. [Google Scholar] [CrossRef]
- Nadeem, M.; Abbasi, B.H.; Younas, M.; Ahmad, W.; Zahir, A.; Hano, C. LED-enhanced biosynthesis of biologically active ingredients in callus cultures of Ocimum basilicum. J. Photochem. Photobiol. B 2019, 190, 172–178. [Google Scholar] [CrossRef]
- Szopa, A.; Ekiert, H. Production of biologically active phenolic acids in Aronia melanocarpa (Michx.) Elliott in vitro cultures cultivated on different variants of the Murashige and Skoog medium. Plant Growth Regul. 2014, 72, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Miransari, M. (Ed.) Environmental Stresses in Soybean Production; Academic Press: Cambridge, MA, USA, 2016; pp. 1–311. [Google Scholar]
- Hammerbacher, A.; Paetz, C.; Wright, L.P.; Fischer, T.C.; Bohlmann, J.; Davis, A.J.; Fenning, T.M.; Gershenzon, J.; Schmidt, A. Flavan-3-ols in Norway spruce: Biosynthesis, accumulation, and function in response to attack by the bark beetle-associated fungus Ceratocystis polonica. Plant Physiol. 2014, 164, 2107–2122. [Google Scholar] [CrossRef] [Green Version]
- Pourebad, N.; Motafakkerazad, R.; Kosari-Nasab, M.; Akhtar, N.F.; Movafeghi, A. The influence of TDZ concentrations on in vitro growth and production of secondary metabolites by the shoot and callus culture of Lallemantia iberica. Plant Cell Tissue Organ Cult. 2015, 122, 331–339. [Google Scholar] [CrossRef]
- Khurshid, R.; Khan, T.; Zaeem, A.; Garros, L.; Hano, C.; Abbasi, B.H. Biosynthesis of precious metabolites in callus cultures of Eclipta alba. Plant Cell Tissue Organ Cult. 2018, 135, 287–298. [Google Scholar] [CrossRef]
- Ali, M.; Abbasi, B.H. Thidiazuron-induced changes in biomass parameters, total phenolic content, and antioxidant activity in callus cultures of Artemisia absinthium L. Appl. Biochem. Biotechnol. 2014, 172, 2363–2376. [Google Scholar] [CrossRef]
- Beejmohun, V.; Fliniaux, O.; Hano, C.; Pilard, S.; Grand, E.; Lesur, D.; Cailleu, D.; Lamblin, F.; Lainé, E.; Kovensky, J.; et al. Coniferin dimerisation in lignan biosynthesis in flax cells. Phytochemistry 2007, 68, 2744–2752. [Google Scholar] [CrossRef] [PubMed]
- Garros, L.; Drouet, S.; Corbin, C.; Decourtil, C.; Fidel, T.; Lebas de Lacour, J.; Leclerc, E.A.; Renouard, S.; Tungmunnithum, D.; Doussot, J.; et al. Insight into the Influence of Cultivar Type, Cultivation Year, and Site on the Lignans and Related Phenolic Profiles, and the Health-Promoting Antioxidant Potential of Flax (Linum usitatissimum L.) Seeds. Molecules 2018, 23, 2636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, H.I.; Akladious, S.A. Changes in antioxidants potential, secondary metabolites and plant hormones induced by different fungicides treatment in cotton plants. Pest Biochem. Physiol. 2017, 142, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.; Ullah, S.; Bao, Y.; Wang, B.; Peng, D.; Liu, L. Light-emitting diodes: Whether an efficient source of light for indoor plants. Environ. Sci. Pollut. Res. 2017, 24, 24743–24752. [Google Scholar] [CrossRef]
- Nazir, N.; Tungmunnithum, D.; Bose, S.; Drouet, S.; Garros, L.; Giglioli-Guivarc’h, N.; Abbasi, B.H.; Hano, C. Differential production of phenylpropanoid metabolites in callus cultures of Ocimum basilicum L. with distinct in vitro antioxidant activities and in vivo protective effects against UV stress. J. Agric. Food Chem. 2019, 67, 1847–1859. [Google Scholar] [CrossRef]
- Wong, S.P.; Leong, L.P.; Koh, J.H.W. Antioxidant activities of aqueous extracts of selected plants. Food Chem. 2006, 99, 775–783. [Google Scholar] [CrossRef]
- Sinha, S. Medicinal plants of Manipur, Manipur Association for Science & Society (MASS). In Proceedings of the Manipur Cultural Integration Conference; MCIC Publishers: Imphal, India, 1996. [Google Scholar]
- Ullah, M.A.; Tungmunnithum, D.; Garros, L.; Drouet, S.; Hano, C.; Abbasi, B.H. Effect of Ultraviolet-C Radiation and Melatonin Stress on Biosynthesis of Antioxidant and Antidiabetic Metabolites Produced in In Vitro Callus Cultures of Lepidium sativum L. Int. J. Mol. Sci. 2019, 20, 1787. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Jimenez, A.; García-Molina, F.; Teruel-Puche, J.A.; Saura-Sanmartin, A.; Garcia-Ruiz, P.A.; Ortiz-Lopez, A.; Rodríguez-López, J.N.; Garcia-Canovas, F.; Munoz-Munoz, J. Catalysis and inhibition of tyrosinase in the presence of cinnamic acid and some of its derivatives. Int. J. Biol. Macromol. 2018, 119, 548–554. [Google Scholar] [CrossRef]
- Oshima, N.; Narukawa, Y.; Takeda, T.; Kiuchi, F. Collagenase inhibitors from Viola yedoensis. J. Nat. Med. 2013, 67, 240–245. [Google Scholar] [CrossRef]
- Qing, W.; Zhao, M.; Kou, C.; Lu, M.; Wang, Y. Functionalization of silver nanoparticles with mPEGylated luteolin for selective visual detection of Hg2+ in water sample. RSC Adv. 2018, 8, 28843–28846. [Google Scholar] [CrossRef] [Green Version]
- Younas, M.; Hano, C.; Giglioli-Guivarc’h, N.; Abbasi, B.H. Mechanistic evaluation of phytochemicals in breast cancer remedy: Current understanding and future perspectives. RSC Adv. 2018, 8, 29714–29744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- More, S.K.; Lande, A.A.; Jagdale, P.G.; Adkar, P.P.; Ambavade, S.D. Evaluation of anti-inflammatory activity of Solanum xanthocarpum Schrad and Wendl (Kaṇṭakāri) extract in laboratory animals. Ancient Sci. Life 2013, 32, 222. [Google Scholar]
- Rathee, P.; Chaudhary, H.; Rathee, S.; Rathee, D.; Kumar, V.; Kohli, K. Mechanism of action of flavonoids as anti-inflammatory agents: A review. Inflamm. Allergy-Drug Targets 2009, 8, 229–235. [Google Scholar] [CrossRef]
- Younas, M.; Drouet, S.; Nadeem, M.; Giglioli-Guivarc’h, N.; Hano, C.; Abbasi, B.H. Differential accumulation of silymarin induced by exposure of Silybum marianum L. callus cultures to several spectres of monochromatic lights. J. Photochem. Photobiol. 2018, 184, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.; Ullah, M.A.; Garros, L.; Hano, C.; Abbasi, B.H. Synergistic effects of melatonin and distinct spectral lights for enhanced production of anti-cancerous compounds in callus cultures of Fagonia indica. J. Photochem. Photobiol. 2019, 190, 163–171. [Google Scholar] [CrossRef]
- Drouet, S.; Abbasi, B.H.; Falguières, A.; Ahmad, W.; Ferroud, C.; Doussot, J.; Vanier, J.R.; Lainé, E.; Hano, C. Single Laboratory Validation of a Quantitative Core Shell-Based LC Separation for the Evaluation of Silymarin Variability and Associated Antioxidant Activity of Pakistani Ecotypes of Milk Thistle (Silybum Marianum L.). Molecules 2018, 23, 904. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| PGR Concentration (mg/L) | FW (g/L) | DW (g/L) | Callus-Initiation Days | Callogenic Frequency (%) | Callus Color | Callus Texture |
|---|---|---|---|---|---|---|
| 0.1 TDZ | 169.53 ± 3.63 fgh | 9.99 ± 0.41 c | 7 | 80 % | DG | C |
| 1.0 TDZ | 219.33 ± 5.68 c | 10.32 ± 0.44 c | 7 | 80 % | DG | C |
| 2.5 TDZ | 246.11 ± 5.72 a | 13.03 ± 0.59 ab | 6 | 100 % | DG | C |
| 5.0 TDZ | 251.48 ± 5.19 a | 13.59 ± 0.63 a | 6 | 100 % | DG | C |
| 10.0 TDZ | 244.03 ± 5.88 a | 12.72 ± 0.47 ab | 6 | 80 % | DG | C |
| 0.1 BAP | 26.12 ± 0.62 n | 2.95 ± 0.13 g | 10 | 20 % | YG | C |
| 1.0 BAP | 84.01 ± 1.71 m | 5.21 ± 0.29 f | 9 | 40 % | YG | C |
| 2.5 BAP | 165.26 ± 1.29 g | 9.56 ± 0.26 c | 9 | 60 % | YG | C |
| 5.0 BAP | 167.33 ± 2.73 fgh | 9.88 ± 0.31 c | 9 | 80 % | YG | C |
| 10.0 BAP | 172.79 ± 2.21 f | 9.99 ± 0.37 c | 9 | 60 % | DG | C |
| 0.1 NAA | 109.14 ± 1.88 k | 5.78 ± 0.29 ef | 10 | 40% | LG | F |
| 1.0 NAA | 187.39 ± 2.37 e | 10.00 ± 0.41 c | 9 | 80% | LG | F |
| 2.5 NAA | 199.62 ± 2.91 d | 10.07 ± 0.37 c | 8 | 80% | YG | F |
| 5.0 NAA | 169.48 ± 2.52 g | 9.94 ± 0.33 c | 10 | 60% | YG | F |
| 10.0 NAA | 93.85 ± 2.11 i | 5.80 ± 0.27 ef | 12 | 40% | YG | F |
| 0.1 TDZ + 1.0 NAA | 118.99 ± 2.73 j | 6.60 ± 0.39 e | 8 | 80% | DG | C |
| 1.0 TDZ + 1.0 NAA | 159.48 ± 3.49 h | 8.61 ± 0.44 d | 8 | 80% | DG | C |
| 2.5 TDZ + 1.0 NAA | 231.40 ± 5.44 b | 11.90 ± 0.49 b | 9 | 80% | DG | C |
| 5.0 TDZ + 1.0 NAA | 238.90 ± 5.69 ab | 12.20 ± 0.41 b | 7 | 100% | DG | C |
| 10.0 TDZ + 1.0 NAA | 144.15 ± 2.86 i | 7.85 ± 0.28 d | 9 | 80% | DG | C |
| PGR Concentration (mg/L) | TPC (mg/g DW) | TFC (mg/g DW) | TPP (mg/L) | TFP (mg/L) |
|---|---|---|---|---|
| 0.1 TDZ | 8.98 ± 0.29 b | 0.89 ± 0.13 de | 89.77 ± 2.41 de | 8.89 ± 0.41 f |
| 1.0 TDZ | 8.95 ± 0.26 bc | 0.94 ± 0.11 cd | 92.44 ± 2.22 d | 9.80 ± 0.29 e |
| 2.5 TDZ | 9.61 ± 0.49 ab | 1.16 ± 0.09 bc | 125.27 ± 2.81 b | 15.18 ± 0.52 b |
| 5.0 TDZ | 10.22 ± 0.44 a | 1.48 ± 0.16 ab | 138.90 ± 2.90 a | 20.16 ± 0.48 a |
| 10.0 TDZ | 9.75 ± 0.49 ab | 1.65 ± 0.11 a | 124.13 ± 2.88 b | 21.0 ± 0.61 a |
| 0.1 BAP | 4.90 ± 0.19 g | 0.68 ± 0.09 e | 14.49 ± 0.88 l | 2.02 ± 0.31 k |
| 1.0 BAP | 8.31 ± 0.27 c | 0.87 ± 0.06 d | 43.34 ± 1.78 i | 4.55 ± 0.29 i |
| 2.5 BAP | 9.46 ± 0.21 b | 1.29 ± 0.09 b | 90.45 ± 2.23 de | 12.33 ± 1.19 cd |
| 5.0 BAP | 8.98 ± 0.28 b | 0.74 ± 0.08 e | 88.75 ± 2.19 de | 7.32 ± 0.31 g |
| 10.0 BAP | 8.88 ± 0.35 bc | 0.84 ± 0.08 de | 88.77 ± 2.20 de | 8.43 ± 0.44 fg |
| 0.1 NAA | 4.50 ± 0.23 g | 0.50 ± 0.08 f | 26.0 ± 1.93 k | 2.89 ± 0.16 j |
| 1.0 NAA | 6.57 ± 0.29 f | 0.76 ± 0.12 de | 65.78 ± 2.21 g | 7.62 ± 0.42 hg |
| 2.5 NAA | 7.45 ± 0.27 de | 0.87 ± 0.14 de | 75.04 ± 1.95 f | 8.78 ± 0.37 f |
| 5.0 NAA | 8.50 ± 0.48 bc | 0.84 ± 0.11 de | 84.55 ± 2.20 e | 8.38 ± 0.32 f |
| 10.0 NAA | 5.94 ± 0.49 f | 0.47 ± 0.06 f | 34.46 ± 0.98 j | 2.72 ± 0.21 j |
| 0.1 TDZ + 1.0 NAA | 4.55 ± 0.29 g | 0.69 ± 0.09 e | 30.06 ± 1.18 k | 4.58 ± 0.28 i |
| 1.0 TDZ + 1.0 NAA | 6.83 ± 0.31 ef | 0.63 ± 0.09 ef | 58.83 ± 1.27 h | 5.42 ± 0.41 i |
| 2.5 TDZ + 1.0 NAA | 7.24 ± 0.34 e | 0.84 ± 0.14 de | 86.212.21 e | 10.08 ± 0.66 de |
| 5.0 TDZ + 1.0 NAA | 8.91 ± 0.52 bc | 0.95 ± 0.17 cd | 108.813.32 c | 11.67 ± 0.61 c |
| 10.0 TDZ + 1.0 NAA | 8.25 ± 0.41 cd | 0.97 ± 0.16 cd | 64.78 ± 2.21 g | 7.65 ± 0.49 fgh |
| PGR Concentration (mg/L) | Caffeic Acid (mg/g DW) | Methyl Caffeate (mg/g DW) | Scopoletin (mg/g DW) | Esculetin (mg/g DW) |
|---|---|---|---|---|
| 0.1 TDZ | 0.46 ± 0.020 cd | 14.30 ± 0.66 def | 3.72 ± 0.23 bcd | 0.72 ± 0.039 cd |
| 1.0 TDZ | 0.43 ± 0.017 d | 13.07 ± 0.48 fg | 3.48 ± 0.21 cd | 0.74 ± 0.041 bc |
| 2.5 TDZ | 0.52 ± 0.024 ab | 16.71 ± 0.61 b | 3.32 ± 0.21 cde | 0.79 ± 0.044 ab |
| 5.0 TDZ | 0.56 ± 0.021 a | 18.62 ± 0.60 a | 4.07 ± 0.24 ab | 0.84 ± 0.043 ab |
| 10.0 TDZ | 0.48 ± 0.020 bc | 15.33 ± 0.44 cd | 4.34 ± 0.20 a | 0.87 ± 0.040 a |
| 0.1 BAP | 0.45 ± 0.021 cd | 11.70 ± 0.40 h | 2.32 ± 0.17 f | 0.62 ± 0.032 e |
| 1.0 BAP | 0.54 ± 0.024 a | 15.65 ± 0.43 bc | 2.33 ± 0.19 f | 0.63 ± 0.038 de |
| 2.5 BAP | 0.55 ± 0.026 a | 18.19 ± 0.61 a | 2.98 ± 0.21 e | 0.71 ± 0.033 c |
| 5.0 BAP | 0.49 ± 0.018 bc | 14.95 ± 0.63 de | 2.99 ± 0.22 e | 0.70 ± 0.039 cd |
| 10.0 BAP | 0.48 ± 0.018 bc | 15.05 ± 0.66 de | 3.57 ± 0.20 bcd | 0.77 ± 0.036 bc |
| 0.1 NAA | 0.37 ± 0.020 ef | 9.42 ± 0.39 ij | 3.20 ± 0.20 cde | 0.73 ± 0.030 c |
| 1.0 NAA | 0.43 ± 0.018 de | 12.56 ± 0.37 g | 3.66 ± 0.21 bcd | 0.79 ± 0.032 ab |
| 2.5 NAA | 0.45 ± 0.019 cd | 12.77 ± 0.34 g | 2.86 ± 0.20 e | 0.68 ± 0.031 cde |
| 5.0 NAA | 0.47 ± 0.026 cd | 14.39 ± 0.38 de | 3.25 ± 0.21 de | 0.73 ± 0.040 bc |
| 10.0 NAA | 0.37 ± 0.024 ef | 9.70 ± 0.41 i | 3.33 ± 0.27 de | 0.75 ± 0.038 bc |
| 0.1 TDZ + 1.0 NAA | 0.39 ± 0.024 e | 10.41 ± 0.39 hi | 2.99 ± 0.18 e | 0.70 ± 0.029 cd |
| 1.0 TDZ + 1.0 NAA | 0.33 ± 0.021 f | 8.71 ± 0.33 j | 3.39 ± 0.29 cde | 0.73 ± 0.033 c |
| 2.5 TDZ + 1.0 NAA | 0.40 ± 0.024 e | 11.48 ± 0.37 h | 3.40 ± 0.29 cde | 0.74 ± 0.032 bc |
| 5.0 TDZ + 1.0 NAA | 0.40 ± 0.021 e | 12.04 ± 0.51 gh | 3.94 ± 0.22 bc | 0.81 ± 0.041 ab |
| 10.0 TDZ + 1.0 NAA | 0.46 ± 0.020 d | 14.10 ± 0.50 ef | 3.77 ± 0.22 bc | 0.80 ± 0.040 ab |
| PGR Concentration (mg/L) | ABTS (µM TEAC) | FRAP (µM TEAC) | DPPH (% RSA) |
|---|---|---|---|
| 0.1 TDZ | 212.85 ± 3.81 i | 312.2 ± 4.36 j | 88.1 ± 4.29 bc |
| 1.0 TDZ | 187.34 ± 4.18 k | 267.1 ± 4.24 k | 76.3 ± 3.72 de |
| 2.5 TDZ | 154.82 ± 3.49 l | 461.7 ± 4.98 e | 89.7 ± 3.84 abc |
| 5.0 TDZ | 402.51 ± 5.16 a | 654.6 ± 5.39 a | 92 ± 3.49 ab |
| 10.0 TDZ | 370.49 ± 5.62 c | 510.9 ± 6.16 b | 94.6 ± 2.29 ab |
| 0.1 BAP | 187.38 ± 4.38 j | 378.1 ± 4.62 gh | 72.7 ± 3.94 e |
| 1.0 BAP | 348.61 ± 6.63 d | 469.1 ± 4.29 d | 79.3 ± 4.11 cde |
| 2.5 BAP | 383.2 ± 4.48 b | 653.3 ± 5.62 a | 92.6 ± 3.10 ab |
| 5.0 BAP | 311.7 ± 4.61 e | 489.6 ± 4.70 c | 88.5 ± 4.84 ab |
| 10.0 BAP | 339.3 ± 4.44 d | 519.2 ± 5.81 b | 90.2 ± 4.17 ab |
| 0.1 NAA | 254.7 ± 3.69 h | 378.4 ± 4.88 g | 78.3 ± 3.22 cde |
| 1.0 NAA | 298.9 ± 3.67 f | 454.7 ± 6.17 de | 82.9 ± 3.96 cd |
| 2.5 NAA | 310.3 ± 3.49 e | 411.9 ± 6.29 f | 87.5 ± 3.72 abc |
| 5.0 NAA | 317.1 ± 4.49 e | 468.9 ± 5.16 d | 91.2 ± 4.19 ab |
| 10.0 NAA | 278.9 ± 4.16 g | 382.1 ± 5.44 g | 87.7 ± 4.40 abc |
| 0.1 TDZ + 1.0 NAA | 293.9 ± 4.38 f | 349.7 ± 5.93 i | 89.2 ± 4.27 abc |
| 1.0 TDZ + 1.0 NAA | 311.4 ± 5.81 e | 367.3 ± 5.37 hi | 91.9 ± 4.51 abc |
| 2.5 TDZ + 1.0 NAA | 303.6 ± 5.89 ef | 391.2 ± 4.87 g | 91.1 ± 3.93 abc |
| 5.0 TDZ + 1.0 NAA | 307.5 ± 4.74 ef | 418.4 ± 4.95 f | 86.6 ± 3.72 bcd |
| 10.0 TDZ + 1.0 NAA | 319.9 ± 5.73 e | 449.9 ± 5.81 e | 90.6 ± 4.61 abc |
| PGR Concentration (mg/L) | Vesperlysine AGEs | Pentosidine AGEs | Tyrosinase | Elastase | Hyaluronidase | Collagenase |
|---|---|---|---|---|---|---|
| 0.1 TDZ | 42.87 ± 2.21 cde | 50.78 ± 3.11 c | 25.48 ± 1.84 cd | 1.35 ± 0.18 i | 2.91 ± 0.19 bc | 46.40 ± 2.11 ab |
| 1.0 TDZ | 40.49 ± 1.86 def | 52.77 ± 3.24 abc | 25.65 ± 1.87 cd | 5.36 ± 0.66 bcd | 2.87 ± 0.21 bc | 45.65 ± 2.20 abc |
| 2.5 TDZ | 47.19 ± 1.89 abc | 59.75 ± 3.15 a | 30.71 ± 2.01 ab | 4.78 ± 0.66 cd | 2.62 ± 0.21 cde | 41.80 ± 2.29 bcd |
| 5.0 TDZ | 51.55 ± 2.67 a | 57.12 ± 3.10 ab | 32.87 ± 2.04 a | 4.65 ± 0.43 cd | 3.14 ± 0.31 ab | 49.52 ± 2.69 a |
| 10.0 TDZ | 45.07 ± 1.96 c | 53.84 ± 3.19 abc | 28.42 ± 1.67 bc | 8.52 ± 0.44 a | 3.38 ± 0.19 a | 43.68 ± 2.20 abc |
| 0.1 BAP | 33.47 ± 1.62 h | 36.12 ± 2.89 d | 17.15 ± 1.19 g | 1.78 ± 0.09 h | 1.67 ± 0.16 f | 22.53 ± 2.21 f |
| 1.0 BAP | 40.83 ± 2.09 e | 55.67 ± 2.96 abc | 26.54 ± 1.64 bcd | 4.56 ± 0.23 d | 1.65 ± 0.16 f | 26.35 ± 1.82 f |
| 2.5 BAP | 49.11 ± 2.20 ab | 52.36 ± 2.91 bc | 32.28 ± 1.51 a | 6.12 ± 0.34 b | 2.24 ± 0.19 de | 37.76 ± 1.89 de |
| 5.0 BAP | 41.34 ± 1.94 de | 53.03 ± 3.11 abc | 24.12 ± 1.50 de | 2.56 ± 0.23 fg | 2.27 ± 0.20 de | 36.41 ± 1.86 e |
| 10.0 BAP | 43.46 ± 1.79 cde | 57.88 ± 3.20 ab | 26.47 ± 1.29 cd | 2.89 ± 0.20 ef | 2.81 ± 0.19 bc | 44.74 ± 2.65 abc |
| 0.1 NAA | 26.47 ± 1.54 i | 56.60 ± 3.19 ab | 18.40 ± 1.24 fg | 2.85 ± 0.21 ef | 2.40 ± 0.20 de | 37.26 ± 2.10 de |
| 1.0 NAA | 36.78 ± 1.39 f | 57.26 ± 3.44 ab | 21.28 ± 1.20 ef | 2.74 ± 0.24 fg | 2.79 ± 0.21 bc | 41.38 ± 2.17 bcd |
| 2.5 NAA | 33.94 ± 1.30 h | 49.85 ± 2.79 c | 23.85 ± 1.17 de | 2.95 ± 0.20 ef | 2.16 ± 0.21 e | 36.39 ± 2.20 e |
| 5.0 NAA | 42.80 ± 2.10 cde | 59.64 ± 3.20 ab | 24.87 ± 1.19 de | 1.45 ± 0.19 hi | 2.51 ± 0.20 cde | 38.04 ± 2.12 de |
| 10.0 NAA | 39.63 ± 1.87 def | 50.63 ± 3.19 bc | 16.17 ± 1.04 g | 5.63 ± 0.44 bc | 2.54 ± 0.21 cde | 37.42 ± 2.19 de |
| 0.1 TDZ + 1.0 NAA | 41.32 ± 2.11 de | 54.33 ± 3.22 abc | 18.12 ± 1.0 fg | 0.63 ± 0.07 j | 2.32 ± 0.20 de | 36.95 ± 2.30 de |
| 1.0 TDZ + 1.0 NAA | 39.82 ± 1.93 def | 57.34 ± 3.24 ab | 17.24 ± 1.11 g | 1.36 ± 0.09 hi | 2.79 ± 0.26 bcd | 39.33 ± 2.31 de |
| 2.5 TDZ + 1.0 NAA | 45.61 ± 1.94 bcd | 55.39 ± 3.21 abc | 23.65 ± 2.0 de | 2.45 ± 0.11 g | 2.73 ± 0.23 bcd | 37.42 ± 2.30 de |
| 5.0 TDZ + 1.0 NAA | 42.38 ± 2.11 d | 58.68 ± 3.20 ab | 23.33 ± 2.03 de | 4.12 ± 0.23 d | 3.20 ± 0.26 ab | 40.93 ± 2.44 cde |
| 10.0 TDZ + 1.0 NAA | 45.58 ± 2.21 bc | 52.19 ± 3.10 abc | 24.30 ± 2.04 de | 3.32 ± 0.21 e | 2.96 ± 0.23 abc | 37.21 ± 2.21 de |
| PGR Concentrations (mg/L) | COX-1 | COX-2 | 15-LOX | sPLA2 |
|---|---|---|---|---|
| 0.1 TDZ | 34.52 ± 1.93 abc | 11.36 ± 0.66 bc | 34.63 ± 1.80 ab | 11.34 ± 1.02 ab |
| 1.0 TDZ | 30.28 ± 1.20 d | 10.25 ± 0.61 cd | 32.78 ± 1.67 b | 7.78 ± 0.38 d |
| 2.5 TDZ | 27.56 ± 1.19 e | 15.56 ± 0.71 a | 29.12 ± 1.09 cd | 7.54 ± 0.33 d |
| 5.0 TDZ | 33.12 ± 1.66 bc | 14.90 ± 0.72 a | 38.53 ± 2.29 a | 6.89 ± 0.41 e |
| 10.0 TDZ | 38.06 ± 2.49 a | 12.36 ± 0.40 b | 35.87 ± 2.02 ab | 6.41 ± 0.30 e |
| 0.1 BAP | 15.63 ± 1.38 h | 10.25 ± 0.33 cd | 18.48 ± 1.62 f | 10.36 ± 0.81 abc |
| 1.0 BAP | 17.25 ± 1.41 h | 8.50 ± 0.39 ef | 17.28 ± 1.72 f | 8.41 ± 0.64 cd |
| 2.5 BAP | 25.63 ± 1.27 efg | 7.41 ± 0.31 f | 26.92 ± 1.61 de | 11.10 ± 1.11 ab |
| 5.0 BAP | 23.14 ± 1.21 g | 9.63 ± 0.41 d | 25.47 ± 1.86 de | 5.89 ± 1.01 e |
| 10.0 BAP | 31.20 ± 2.03 bcd | 9.63 ± 0.44 cd | 31.16 ± 2.11 bc | 8.63 ± 0.71 cd |
| 0.1 NAA | 25.31 ± 1.81 ef | 12.54 ± 0.51 b | 26.63 ± 2.03 d | 6.13 ± 0.44 e |
| 1.0 NAA | 30.36 ± 1.96 cde | 14.20 ± 0.62 a | 32.86 ± 1.92 b | 7.45 ± 0.49 de |
| 2.5 NAA | 22.15 ± 1.05 g | 12.36 ± 0.49 b | 23.65 ± 1.49 e | 5.85 ± 0.32 e |
| 5.0 NAA | 26.85 ± 1.23 ef | 14.36 ± 0.51 a | 27.87 ± 1.67 cde | 10 ± 0.40 b |
| 10.0 NAA | 26.95 ± 1.06 e | 14.45 ± 0.62 a | 28.13 ± 1.68 cde | 11.25 ± 0.61 a |
| 0.1 TDZ + 1.0 NAA | 24.15 ± 1.11 fg | 11.74 ± 0.42 b | 25.73 ± 1.37 e | 7.63 ± 0.39 d |
| 1.0 TDZ + 1.0 NAA | 30.29 ± 2.08 cde | 8.52 ± 0.32 ef | 32.92 ± 2.02 b | 8.93 ± 0.46 c |
| 2.5 TDZ + 1.0 NAA | 29.17 ± 2.01 cde | 9.65 ± 0.39 d | 35.56 ± 2.21 ab | 10.14 ± 0.60 ab |
| 5.0 TDZ + 1.0 NAA | 35.70 ± 2.06 ab | 9.34 ± 0.31 de | 35.42 ± 2.08 ab | 8.45 ± 0.48 cd |
| 10.0 TDZ + 1.0 NAA | 32.23 ± 2.11 abcd | 10.64 ± 0.44 cd | 33.87 ± 2.04 ab | 6.76 ± 0.40 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usman, H.; Jan, H.; Zaman, G.; Khanum, M.; Drouet, S.; Garros, L.; Tungmunnithum, D.; Hano, C.; Abbasi, B.H. Comparative Analysis of Various Plant-Growth-Regulator Treatments on Biomass Accumulation, Bioactive Phytochemical Production, and Biological Activity of Solanum virginianum L. Callus Culture Extracts. Cosmetics 2022, 9, 71. https://doi.org/10.3390/cosmetics9040071
Usman H, Jan H, Zaman G, Khanum M, Drouet S, Garros L, Tungmunnithum D, Hano C, Abbasi BH. Comparative Analysis of Various Plant-Growth-Regulator Treatments on Biomass Accumulation, Bioactive Phytochemical Production, and Biological Activity of Solanum virginianum L. Callus Culture Extracts. Cosmetics. 2022; 9(4):71. https://doi.org/10.3390/cosmetics9040071
Chicago/Turabian StyleUsman, Hazrat, Hasnain Jan, Gouhar Zaman, Mehnaz Khanum, Samantha Drouet, Laurine Garros, Duangjai Tungmunnithum, Christophe Hano, and Bilal Haider Abbasi. 2022. "Comparative Analysis of Various Plant-Growth-Regulator Treatments on Biomass Accumulation, Bioactive Phytochemical Production, and Biological Activity of Solanum virginianum L. Callus Culture Extracts" Cosmetics 9, no. 4: 71. https://doi.org/10.3390/cosmetics9040071
APA StyleUsman, H., Jan, H., Zaman, G., Khanum, M., Drouet, S., Garros, L., Tungmunnithum, D., Hano, C., & Abbasi, B. H. (2022). Comparative Analysis of Various Plant-Growth-Regulator Treatments on Biomass Accumulation, Bioactive Phytochemical Production, and Biological Activity of Solanum virginianum L. Callus Culture Extracts. Cosmetics, 9(4), 71. https://doi.org/10.3390/cosmetics9040071