
In Vitro Sensitive Skin Models: Review of the Standard Methods and Introduction to a New Disruptive Technology


Abstract
:1. Introduction
2. Review of In Vitro 2D/3D Models of Innervated Skin
2.1. Sensory Neuron Culture and Coculture
2.2. Two/Three-Dimensional Models and the Connection between Cell Types
3. Organ-on-Chip Models and Their Use in Dermatology
3.1. Introduction to Microfluidic Devices and Organ-on-Chip Models
3.2. Development of an Innervated Skin-on-a-Chip
3.2.1. Interest and Objectives
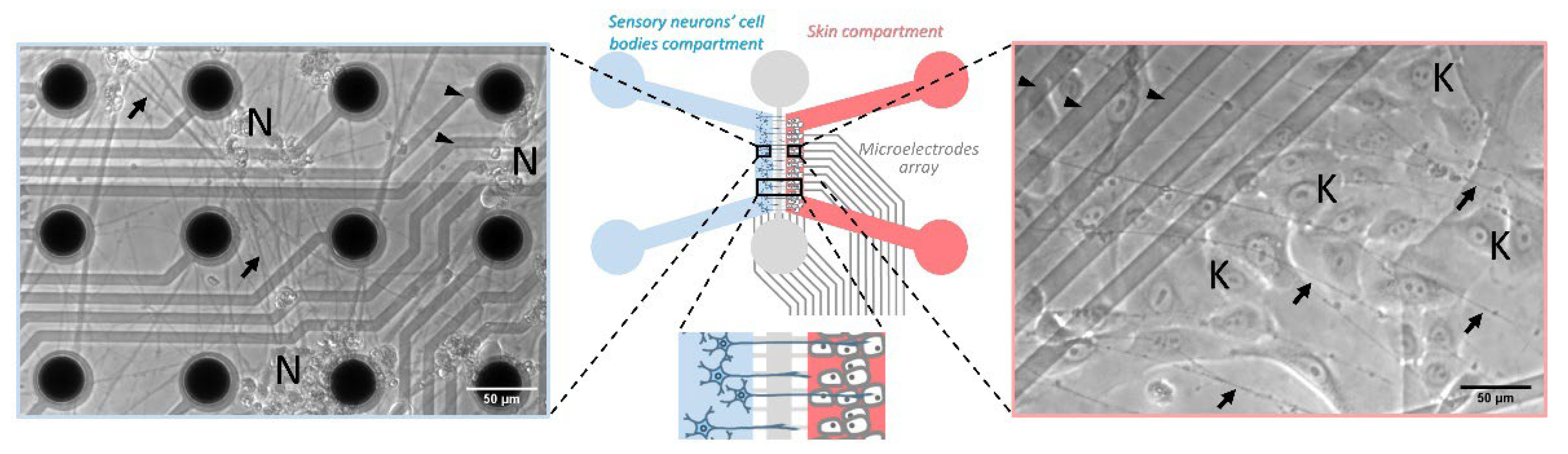
3.2.2. Design of an Innervated Skin-on-Chip
3.2.3. From Standard to Innovative Specific Readouts
4. Future Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laverdet, B.; Danigo, A.; Girard, D.; Magy, L.; Demiot, C.; Desmoulière, A. Skin innervation: Important roles during normal and pathological cutaneous repair. Histol. Histopathol. 2015, 30, 875–892. [Google Scholar] [CrossRef] [PubMed]
- Misery, L.; Brenaut, E.; Le Garrec, R.; Abasq, C.; Genestet, S.; Marcorelles, P.; Zagnoli, F. Neuropathic pruritus. Nat. Rev. Neurol. 2014, 10, 408–416. [Google Scholar] [CrossRef]
- Siemionow, M.; Gharb, B.B.; Rampazzo, A. The Face as a Sensory Organ. Plast. Reconstr. Surg. 2011, 127, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Abraira, V.E.; Ginty, D.D. The Sensory Neurons of Touch. Neuron 2013, 79, 618–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmelz, M.; Schmidt, R.; Bickel, A.; Handwerker, H.O.; Torebjörk, H.E. Specific C-Receptors for Itch in Human Skin. J. Neurosci. 1997, 17, 8003–8008. [Google Scholar] [CrossRef]
- Potenzieri, C.; Undem, B.J. Basic mechanisms of itch. Clin. Exp. Allergy 2012, 42, 8–19. [Google Scholar] [CrossRef] [Green Version]
- Wessberg, J.; Olausson, H.; Fernström, K.W.; Vallbo, A.B. Receptive Field Properties of Unmyelinated Tactile Afferents in the Human Skin. J. Neurophysiol. 2003, 89, 1567–1575. [Google Scholar] [CrossRef]
- Dubin, A.E.; Patapoutian, A. Nociceptors: The sensors of the pain pathway. J. Clin. Investig. 2010, 120, 3760–3772. [Google Scholar] [CrossRef] [Green Version]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Shim, W.-S.; Tak, M.-H.; Lee, M.-H.; Kim, M.; Koo, J.-Y.; Lee, C.-H.; Oh, U. TRPV1 Mediates Histamine-Induced Itching via the Activation of Phospholipase A2 and 12-Lipoxygenase. J. Neurosci. 2007, 27, 2331–2337. [Google Scholar] [CrossRef]
- Misery, L. Sensitive Skins May Be Neuropathic Disorders: Lessons from Studies on Skin and Other Organs. Cosmetics 2021, 8, 14. [Google Scholar] [CrossRef]
- Julius, D. TRP Channels and Pain. Annu. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef] [Green Version]
- Laursen, W.J.; Anderson, E.O.; Hoffstaetter, L.J.; Bagriantsev, S.N.; Gracheva, E.O. Species-specific temperature sensitivity of TRPA1. Temperature 2015, 2, 214–226. [Google Scholar] [CrossRef]
- Maglie, R.; de Araujo, D.S.M.; Antiga, E.; Geppetti, P.; Nassini, R.; de Logu, F. The Role of TRPA1 in Skin Physiology and Pathology. Int. J. Mol. Sci. 2021, 22, 3065. [Google Scholar] [CrossRef]
- Rice, F.; Albrecht, P. Cutaneous Mechanisms of Tactile Perception: Morphological and Chemical Organization of the Innervation to the Skin. Senses A Compr. Ref. 2008, 6, 1–31. [Google Scholar] [CrossRef]
- Le Bars, D.; Adam, F. Nocicepteurs et médiateurs dans la douleur aiguë inflammatoire. In Ann Fr Anesth Reanim; Elsevier Masson: Amsterdam, The Netherlands, 2002; Volume 21, pp. 315–335. [Google Scholar] [CrossRef]
- Misery, L.; Ständer, S.; Szepietowski, J.; Reich, A.; Wallengren, J.C.; Evers, A.W.M.; Takamori, K.; Brenaut, E.; Gall-Ianotto, C.; Fluhr, J.; et al. Definition of Sensitive Skin: An Expert Position Paper from the Special Interest Group on Sensitive Skin of the International Forum for the Study of Itch. Acta Derm. Venereol. 2017, 97, 4–6. [Google Scholar] [CrossRef] [Green Version]
- Buhé, V.; Vié, K.; Guéré, C.; Natalizio, A.; Lhéritier, C.; Gall-Ianotto, C.; Huet, F.; Talagas, M.; Lebonvallet, N.; Marcorelles, P.; et al. Pathophysiological Study of Sensitive Skin. Acta Derm. Venereol. 2016, 96, 314–318. [Google Scholar] [CrossRef] [Green Version]
- Facer, P.; A Casula, M.A.; Smith, G.D.; Benham, C.D.; Chessell, I.P.; Bountra, C.; Sinisi, M.; Birch, R.; Anand, P. Differential expression of the capsaicin receptor TRPV1 and related novel receptors TRPV3, TRPV4 and TRPM8 in normal human tissues and changes in traumatic and diabetic neuropathy. BMC Neurol. 2007, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Koizumib, S.; Fuziwaraa, S.; Dendaa, S.; Inoueb, K.; Dendaa, M. Functional Vanilloid Receptors in Cultured Normal Human Epidermal Keratinocytes. Biochem. Biophys. Res. Commun. 2002, 291, 124–129. [Google Scholar] [CrossRef]
- Fromy, B.; Josset-Lamaugarny, A.; Aimond, G.; Pagnon-Minot, A.; Marics, I.; Tattersall, G.J.; Moqrich, A.; Sigaudo-Roussel, D. Disruption of TRPV3 Impairs Heat-Evoked Vasodilation and Thermoregulation: A Critical Role of CGRP. J. Investig. Dermatol. 2018, 138, 688–696. [Google Scholar] [CrossRef] [Green Version]
- Talagas, M.; Lebonvallet, N.; Leschiera, R.; Sinquin, G.; Elies, P.; Haftek, M.; Pennec, J.-P.; Ressnikoff, D.; La Padula, V.; Le Garrec, R.; et al. Keratinocytes Communicate with Sensory Neurons via Synaptic-like Contacts. Ann. Neurol. 2020, 88, 1205–1219. [Google Scholar] [CrossRef]
- Misery, L. Irritated Skin Is Not Sensitive Skin. JID Innov. 2021, 1, 100031. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.K.; Ai, X.; Hickman, G.E.; MacPhedran, S.E.; Nduaguba, C.O.; Robertson, C.P. The Generation of Neuronal Heterogeneity in a Rat Sensory Ganglion. J. Neurosci. 1997, 17, 2775–2784. [Google Scholar] [CrossRef] [Green Version]
- Hall, A.K. Rodent Sensory Neuron Culture and Analysis. Curr. Protoc. Neurosci. 2006, 36, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-T.; Chen, J.-C. Dorsal Root Ganglia Isolation and Primary Culture to Study Neurotransmitter Release. J. Vis. Exp. JoVE 2018, 140, e57569. [Google Scholar] [CrossRef] [Green Version]
- Cevikbas, F.; Wang, X.; Akiyama, T.; Kempkes, C.; Savinko, T.; Antal, A.; Kukova, G.; Buhl, T.; Ikoma, A.; Buddenkotte, J.; et al. A sensory neuron–expressed IL-31 receptor mediates T helper cell–dependent itch: Involvement of TRPV1 and TRPA1. J. Allergy Clin. Immunol. 2014, 133, 448–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, S.R.; Thé, L.; Batia, L.M.; Beattie, K.; Katibah, G.E.; McClain, S.P.; Pellegrino, M.; Estandian, D.M.; Bautista, D.M. The Epithelial Cell-Derived Atopic Dermatitis Cytokine TSLP Activates Neurons to Induce Itch. Cell 2013, 155, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Château, Y.; Dorange, G.; Clément, J.-F.; Pennec, J.-P.; Gobin, E.; Griscom, L.; Baudrimont, M.; Rougier, N.; Chesné, C.; Misery, L. In Vitro Reconstruction of Neuro-Epidermal Connections. J. Investig. Dermatol. 2007, 127, 979–981. [Google Scholar] [CrossRef] [Green Version]
- Guéniche, A.; Bastien, P.; Ovigne, J.M.; Kermici, M.; Courchay, G.; Chevalier, V.; Breton, L.; Castiel-Higounenc, I. Bifidobacterium longum lysate, a new ingredient for reactive skin. Exp. Dermatol. 2009, 19, e1–e8. [Google Scholar] [CrossRef]
- Muller, Q.; Beaudet, M.-J.; de Serres-Bérard, T.; Bellenfant, S.; Flacher, V.; Berthod, F. Development of an innervated tissue-engineered skin with human sensory neurons and Schwann cells differentiated from iPS cells. Acta Biomater. 2018, 82, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Khammo, N.; Ogilvie, J.; Clothier, R.H. Development of an innervated model of human skin. Altern. Lab. Anim. ATLA 2007, 35, 487–491. [Google Scholar] [CrossRef]
- Roggenkamp, D.; Falkner, S.; Stäb, F.; Petersen, M.; Schmelz, M.; Neufang, G. Atopic Keratinocytes Induce Increased Neurite Outgrowth in a Coculture Model of Porcine Dorsal Root Ganglia Neurons and Human Skin Cells. J. Investig. Dermatol. 2012, 132, 1892–1900. [Google Scholar] [CrossRef] [Green Version]
- Roggenkamp, D.; Köpnick, S.; Stäb, F.; Wenck, H.; Schmelz, M.; Neufang, G. Epidermal Nerve Fibers Modulate Keratinocyte Growth via Neuropeptide Signaling in an Innervated Skin Model. J. Investig. Dermatol. 2013, 133, 1620–1628. [Google Scholar] [CrossRef] [Green Version]
- Blais, M.; Mottier, L.; Germain, M.-A.; Bellenfant, S.; Cadau, S.; Berthod, F. Sensory Neurons Accelerate Skin Reepithelialization via Substance P in an Innervated Tissue-Engineered Wound Healing Model. Tissue Eng. Part A 2014, 20, 2180–2188. [Google Scholar] [CrossRef] [Green Version]
- Cadau, S.; Leoty-Okombi, S.; Pain, S.; Bechetoille, N.; André-Frei, V.; Berthod, F. In vitro glycation of an endothelialized and innervated tissue-engineered skin to screen anti-AGE molecules. Biomaterials 2015, 51, 216–225. [Google Scholar] [CrossRef]
- Martorina, F.; Casale, C.; Urciuolo, F.; Netti, P.A.; Imparato, G. In vitro activation of the neuro-transduction mechanism in sensitive organotypic human skin model. Biomaterials 2017, 113, 217–229. [Google Scholar] [CrossRef]
- Reichert, O.; Fleming, T.; Neufang, G.; Schmelz, M.; Genth, H.; Kaever, V.; Wenck, H.; Stäb, F.; Terstegen, L.; Kolbe, L.; et al. Impaired glyoxalase activity is associated with reduced expression of neurotrophic factors and pro-inflammatory processes in diabetic skin cells. Exp. Dermatol. 2017, 26, 44–50. [Google Scholar] [CrossRef]
- Bize, C.; Lavigne, E.G.; Monneuse, J.-M.; Chrétien, A.; Anchartechahar, V.; Hubaux, R.; Skorski, G.; Salmon, M.; Steinschneider, R. Evaluation of Soothing Activity of Aphloia Theiformis Extract Using an Innovative Model of Human Innerved Reconstructed Epidermis. The IFSCC Congress 2020 Yokohama, Virtual Event, 21–30 October 2020; pp. 1500–1508. [Google Scholar]
- Lebonvallet, N.; Fluhr, J.W.; Le Gall-Ianotto, C.; Leschiera, R.; Talagas, M.; Reux, A.; Bataille, A.; Brun, C.; Oddos, T.; Pennec, J.; et al. A re-innervated in vitro skin model of non-histaminergic itch and skin neurogenic inflammation: PAR2-, TRPV1- and TRPA1-agonist induced functionality. Ski. Health Dis. 2021, 1, e66. [Google Scholar] [CrossRef]
- Gingras, M.; Bergeron, J.; Déry, J.; Durham, H.D.; Berthod, F. In vitro development of a tissue-engineered model of peripheral nerve regeneration to study neurite growth. FASEB J. 2003, 17, 2124–2126. [Google Scholar] [CrossRef] [Green Version]
- Baumbauer, K.M.; DeBerry, J.J.; Adelman, P.C.; Miller, R.H.; Hachisuka, J.; Lee, K.H.; Ross, S.E.; Koerber, H.R.; Davis, B.M.; Albers, K.M. Keratinocytes can modulate and directly initiate nociceptive responses. eLife 2015, 4, e09674. [Google Scholar] [CrossRef]
- Pang, Z.; Sakamoto, T.; Tiwari, V.; Kim, Y.-S.; Yang, F.; Dong, X.; Güler, A.D.; Guan, Y.; Caterina, M.J. Selective keratinocyte stimulation is sufficient to evoke nociception in mice. Pain 2015, 156, 656–665. [Google Scholar] [CrossRef]
- Tsantoulas, C.; Farmer, C.; Machado, P.; Baba, K.; McMahon, S.B.; Raouf, R. Probing Functional Properties of Nociceptive Axons Using a Microfluidic Culture System. PLoS ONE 2013, 8, e80722. [Google Scholar] [CrossRef]
- Valtcheva, M.V.; Copits, B.A.; Davidson, S.; Sheahan, T.D.; Pullen, M.Y.; McCall, J.G.; Dikranian, K.; Gereau, R.W. Surgical extraction of human dorsal root ganglia from organ donors and preparation of primary sensory neuron cultures. Nat. Protoc. 2016, 11, 1877–1888. [Google Scholar] [CrossRef] [Green Version]
- Schutte, S.C.; Kadakia, F.; Davidson, S. Skin-Nerve Co-Culture Systems for Disease Modeling and Drug Discovery. Tissue Eng. Part C: Methods 2021, 27, 89–99. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Marmigere, F.; Ernfors, P. Specification and connectivity of neuronal subtypes in the sensory lineage. Nat. Rev. Neurosci. 2007, 8, 114–127. [Google Scholar] [CrossRef]
- Chambers, S.M.; Qi, Y.; Mica, Y.; Lee, G.; Zhang, X.-J.; Niu, L.; Bilsland, J.; Cao, L.; Stevens, E.; Whiting, P.; et al. Combined small-molecule inhibition accelerates developmental timing and converts human pluripotent stem cells into nociceptors. Nat. Biotechnol. 2012, 30, 715–720. [Google Scholar] [CrossRef] [Green Version]
- Lebonvallet, N.; Boulais, N.; Le Gall, C.; Pereira, U.; Gauché, D.; Gobin, E.; Pers, J.-O.; Jeanmaire, C.; Danoux, L.; Pauly, G.; et al. Effects of the re-innervation of organotypic skin explants on the epidermis. Exp. Dermatol. 2012, 21, 156–158. [Google Scholar] [CrossRef]
- Chéret, J.; Lebonvallet, N.; Buhé, V.; Carre, J.L.; Misery, L.; Le Gall-Ianotto, C. Influence of sensory neuropeptides on human cutaneous wound healing process. J. Dermatol. Sci. 2014, 74, 193–203. [Google Scholar] [CrossRef]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; I Basbaum, A.I.; Julius, D. The Cloned Capsaicin Receptor Integrates Multiple Pain-Producing Stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Zygmunt, P.M.; Petersson, J.; Andersson, D.A.; Chuang, -H.; Sørgård, M.; Di Marzo, V.; Julius, D.; Högestätt, E.D. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 1999, 400, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Thresh, L. Isolation of Capsaicin. Pharm. J. 1846, 6, 941–947. [Google Scholar]
- Guimarães, M.Z.P.; de Vecchi, R.; Vitória, G.; Sochacki, J.K.; Paulsen, B.S.; Lima, I.; Rodrigues da Silva, F.; da Costa, R.F.M.; Castro, N.G.; Breton, L.; et al. Generation of IPSC-Derived Human Peripheral Sensory Neurons Releasing Substance P Elicited by TRPV1 Agonists. Front. Mol. Neurosci. 2018, 11, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulk, M.; Steinhoff, M. Chapter 17—Role of TRP Channels in Skin Diseases. In TRP Channels as Therapeutic Targets; Szallasi, A., Ed.; Academic Press: Boston, MA, USA, 2015; pp. 293–323. ISBN 978-0-12-420024-1. [Google Scholar]
- Armstrong, S.A.; Herr, M.J. Physiology, Nociception; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Taylor, A.M.; Blurton-Jones, M.; Rhee, S.W.; Cribbs, D.H.; Cotman, C.W.; Jeon, N.L. A microfluidic culture platform for CNS axonal injury, regeneration and transport. Nat. Methods 2005, 2, 599–605. [Google Scholar] [CrossRef]
- Li, J.; Chen, J.; Bai, H.; Wang, H.; Hao, S.; Ding, Y.; Peng, B.; Zhang, J.; Li, L.; Huang, W. An Overview of Organs-on-Chips Based on Deep Learning. Research 2022, 2022, 1–20. [Google Scholar] [CrossRef]
- Ponmozhi, J.; Dhinakaran, S.; Varga-Medveczky, Z.; Fónagy, K.; Bors, L.; Iván, K.; Erdő, F. Development of Skin-On-A-Chip Platforms for Different Utilizations: Factors to Be Considered. Micromachines 2021, 12, 294. [Google Scholar] [CrossRef]
- Risueño, I.; Valencia, L.; Jorcano, J.L.; Velasco, D. Skin-on-a-chip models: General overview and future perspectives. APL Bioeng. 2021, 5, 030901. [Google Scholar] [CrossRef]
- Wufuer, M.; Lee, G.; Hur, W.; Jeon, B.; Kim, B.J.; Choi, T.H.; Lee, S. Skin-on-a-chip model simulating inflammation, edema and drug-based treatment. Sci. Rep. 2016, 6, 37471. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Ellett, F.; Thomas, C.N.; Jalali, F.; Anderson, R.R.; Irimia, D.; Raff, A.B. A microscale, full-thickness, human skin on a chip assay simulating neutrophil responses to skin infection and antibiotic treatments. Lab Chip 2019, 19, 3094–3103. [Google Scholar] [CrossRef]
- Wagner, I.; Materne, E.-M.; Brincker, S.; Süßbier, U.; Frädrich, C.; Busek, M.; Sonntag, F.; Sakharov, D.A.; Trushkin, E.V.; Tonevitsky, A.G.; et al. A dynamic multi-organ-chip for long-term cultivation and substance testing proven by 3D human liver and skin tissue co-culture. Lab Chip 2013, 13, 3538–3547. [Google Scholar] [CrossRef] [Green Version]
- Maschmeyer, I.; Hasenberg, T.; Jaenicke, A.; Lindner, M.; Lorenz, A.K.; Zech, J.; Garbe, L.-A.; Sonntag, F.; Hayden, P.; Ayehunie, S.; et al. Chip-based human liver–intestine and liver–skin co-cultures—A first step toward systemic repeated dose substance testing in vitro. Eur. J. Pharm. Biopharm. 2015, 95, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Maschmeyer, I.; Lorenz, A.K.; Schimek, K.; Hasenberg, T.; Ramme, A.P.; Hübner, J.; Lindner, M.; Drewell, C.; Bauer, S.; Thomas, A.; et al. A four-organ-chip for interconnected long-term co-culture of human intestine, liver, skin and kidney equivalents. Lab Chip 2015, 15, 2688–2699. [Google Scholar] [CrossRef] [Green Version]
- Kühnl, J.; Tao, T.P.; Brandmair, K.; Gerlach, S.; Rings, T.; Müller-Vieira, U.; Przibilla, J.; Genies, C.; Jaques-Jamin, C.; Schepky, A.; et al. Characterization of application scenario-dependent pharmacokinetics and pharmacodynamic properties of permethrin and hyperforin in a dynamic skin and liver multi-organ-chip model. Toxicology 2021, 448, 152637. [Google Scholar] [CrossRef]
- Tavares, R.S.N.; Tao, T.P.; Maschmeyer, I.; Maria-Engler, S.S.; Schäfer-Korting, M.; Winter, A.; Zoschke, C.; Lauster, R.; Marx, U.; Gaspar, L.R. Toxicity of topically applied drugs beyond skin irritation: Static skin model vs. Two organs-on-a-chip. Int. J. Pharm. 2020, 589, 119788. [Google Scholar] [CrossRef]
- Ataç, B.; Wagner, I.; Horland, R.; Lauster, R.; Marx, U.; Tonevitsky, A.G.; Azar, R.P.; Lindner, G. Skin and hair on-a-chip: In vitro skin models versus ex vivo tissue maintenance with dynamic perfusion. Lab Chip 2013, 13, 3555–3561. [Google Scholar] [CrossRef] [Green Version]
- Misery, L. Skin, immunity and the nervous system. Br. J. Dermatol. 1997, 137, 843–850. [Google Scholar] [CrossRef]
- Roosterman, D.; Goerge, T.; Schneider, S.W.; Bunnett, N.W.; Steinhoff, M. Neuronal Control of Skin Function: The Skin as a Neuroimmunoendocrine Organ. Physiol. Rev. 2006, 86, 1309–1379. [Google Scholar] [CrossRef]
- Ji, R.-R.; Samad, T.A.; Jin, S.-X.; Schmoll, R.; Woolf, C.J. p38 MAPK Activation by NGF in Primary Sensory Neurons after Inflammation Increases TRPV1 Levels and Maintains Heat Hyperalgesia. Neuron 2002, 36, 57–68. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| 2D/3D Model | Origin of Neurons | Reference | Cell Types of the Model | Stimulus | Response |
|---|---|---|---|---|---|
| 2D model | Animal | [29] | Tri-compartmented culture with (1) rat spinal cords cells; (2) rat DRG; (3) epidermal cells (keratinocytes, Merkel cells and melanocytes) | Hot medium in the epidermal/axonal compartment: - 22 °C (control) - 37 °C (heat stimulation) - 45 °C (painful stimulation) | Electrophysiological recording in the soma compartment by patch clamp showed spikes registered at 37 and 45 °C but not at 22 °C. |
| [44] | Bi-compartmented culture with (1) rat DRG neurons; (2) rat primary epidermal keratinocytes | Systemic application of 100 nM capsaicin (TRPV1 agonist) in the epidermal/axonal compartment | Calcium imaging in the soma compartment showed a similar neuron activation by capsaicin either with or without keratinocytes. | ||
| 3D model | Animal | [37] | Immortal human keratinocyte line of HaCaT cells, HDF, rat DRG neurons | Topical application of 5 µM capsaicin (TRPV1 agonist) | Calcium imaging on neurites showed calcium waves increase by capsaicin. |
| [35] | HEK, HDF, HUVEC or HDMEC, mouse DRG neurons | Systemic application of 0.01 ng/mL capsaicin (TRPV1 agonist) | 4-fold increase of substance P release in culture supernatant with capsaicin treatment. | ||
| [40] | Human skin explant, rat DRG neurons | Topical application of 100 µM SLIGKV (PAR2 agonist), 10 µM capsaicin (TRPV1 agonist) or 3 µM polygodial (TRPA1 agonist) | - Electrophysiological recording by patch-clamp showed that all treatments stimulated reinnervating neurons. - SLIGKV treatment increased TSLP release and decreased VEGF expression. - Capsaicin treatment increased TSLP and CGRP release and decreased the expression of several genes in neurons, including CGRPα, MT5-MMP, and BDNF (involved in skin sensitivity). - Polygodial increased CGRP release and TNF, TSLP, and NGF expression in skin explants and decreased EGF and BDNF expression in neurons. | ||
| Human | [39] | HEK, human iPSC-derived sensory neurons | Systemic application of 10 µM capsaicin (TRPV1 agonist) | Capsaicin treatment increased CGRP release in the culture supernatent. This effect was reduced by applying a soothing agent. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guichard, A.; Remoué, N.; Honegger, T. In Vitro Sensitive Skin Models: Review of the Standard Methods and Introduction to a New Disruptive Technology. Cosmetics 2022, 9, 67. https://doi.org/10.3390/cosmetics9040067
Guichard A, Remoué N, Honegger T. In Vitro Sensitive Skin Models: Review of the Standard Methods and Introduction to a New Disruptive Technology. Cosmetics. 2022; 9(4):67. https://doi.org/10.3390/cosmetics9040067
Chicago/Turabian StyleGuichard, Alexandre, Noëlle Remoué, and Thibault Honegger. 2022. "In Vitro Sensitive Skin Models: Review of the Standard Methods and Introduction to a New Disruptive Technology" Cosmetics 9, no. 4: 67. https://doi.org/10.3390/cosmetics9040067
APA StyleGuichard, A., Remoué, N., & Honegger, T. (2022). In Vitro Sensitive Skin Models: Review of the Standard Methods and Introduction to a New Disruptive Technology. Cosmetics, 9(4), 67. https://doi.org/10.3390/cosmetics9040067