Inonotus obliquus Extract as An Inhibitor of α-MSH-Induced Melanogenesis in B16F10 Mouse Melanoma Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. B16F10 Mouse Melanoma Cell Culture
2.2. Preparation of Inonotus Obliquus Extract
2.3. Cell Viability Assay
2.4. Measurement of Melanin Content
2.5. Tyrosinase Activity Assay
2.6. Expression of Tyrosinase mRNA
2.7. Determination of Tyrosinase Protein
2.8. Mitf Transcriptional Activity
2.9. Statistical Analysis
3. Results
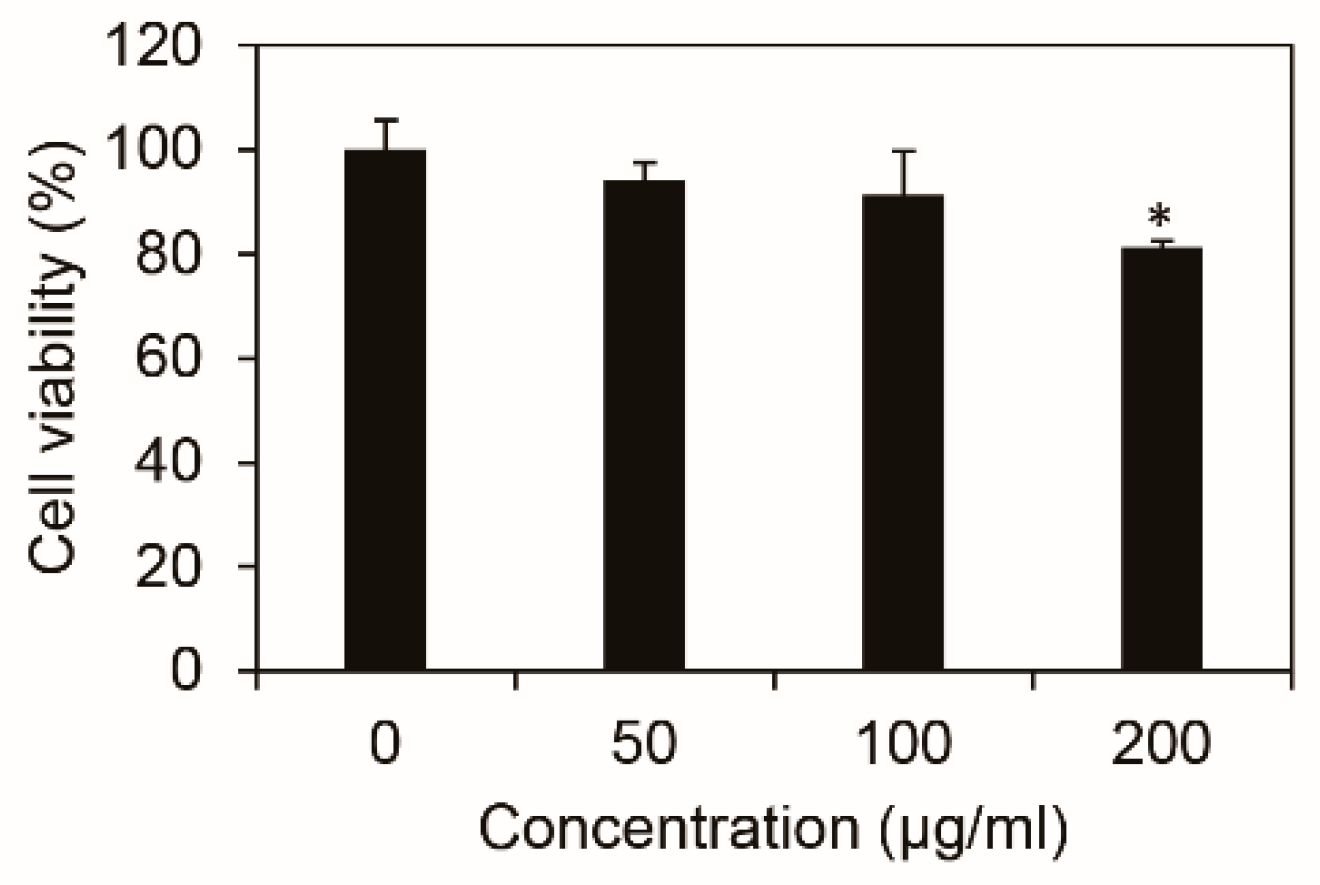
3.1. Cell Viability of Inonotus obliquus Extract in B16F10 Mouse Melanoma Cells
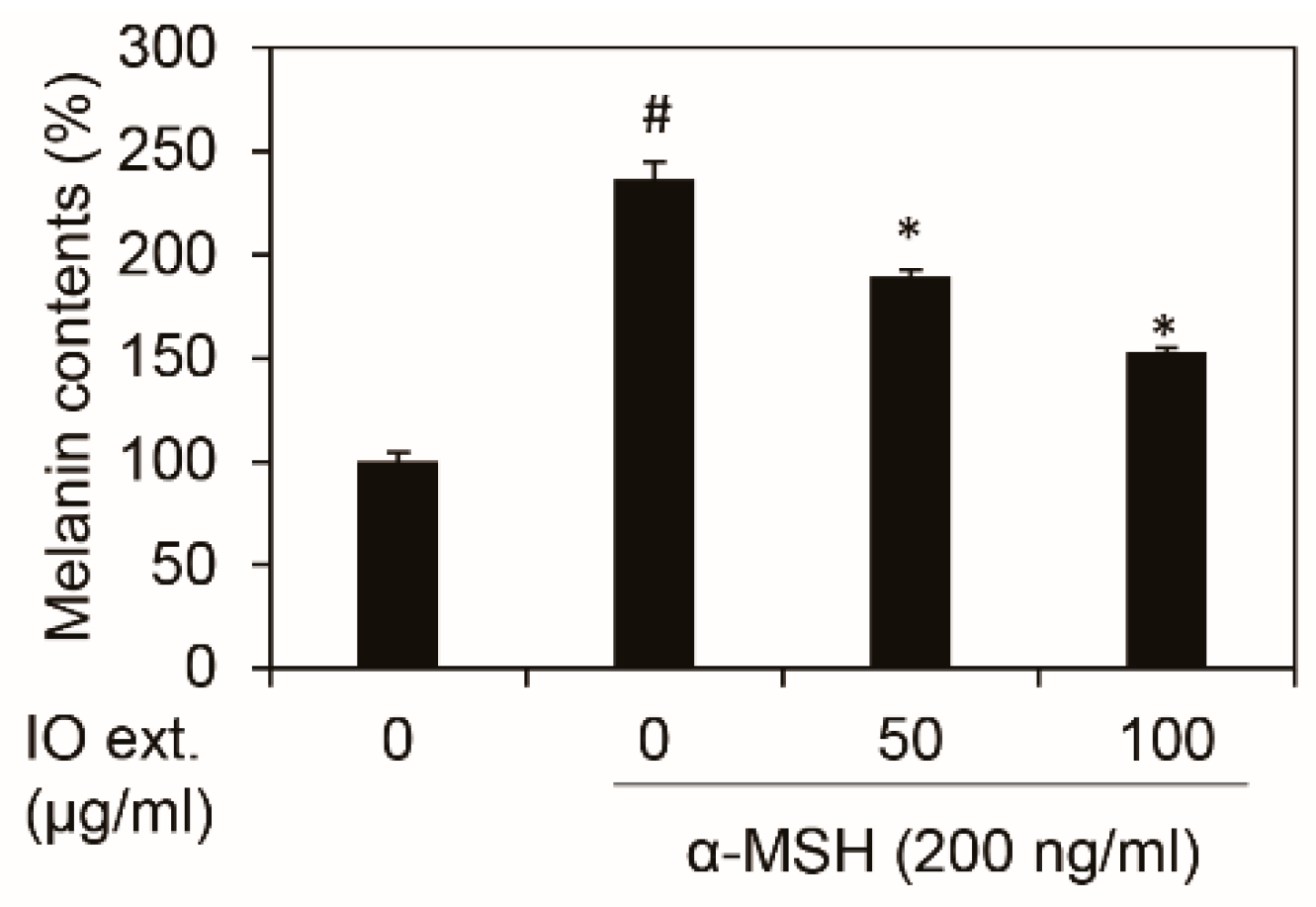
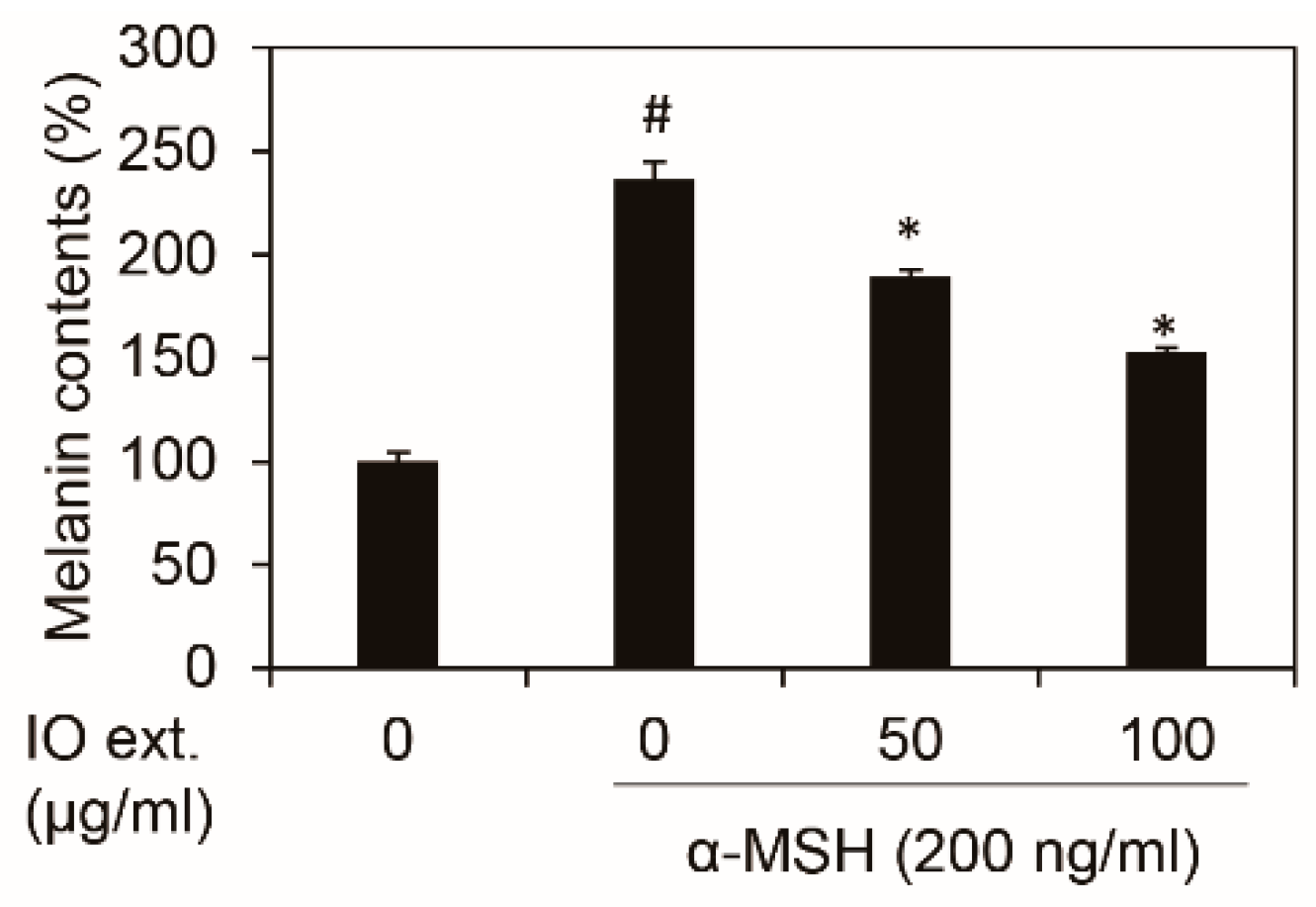
3.2. Inonotus obliquus Extract Decreases Melanin Contents in B16F10 Mouse Melanoma Cells
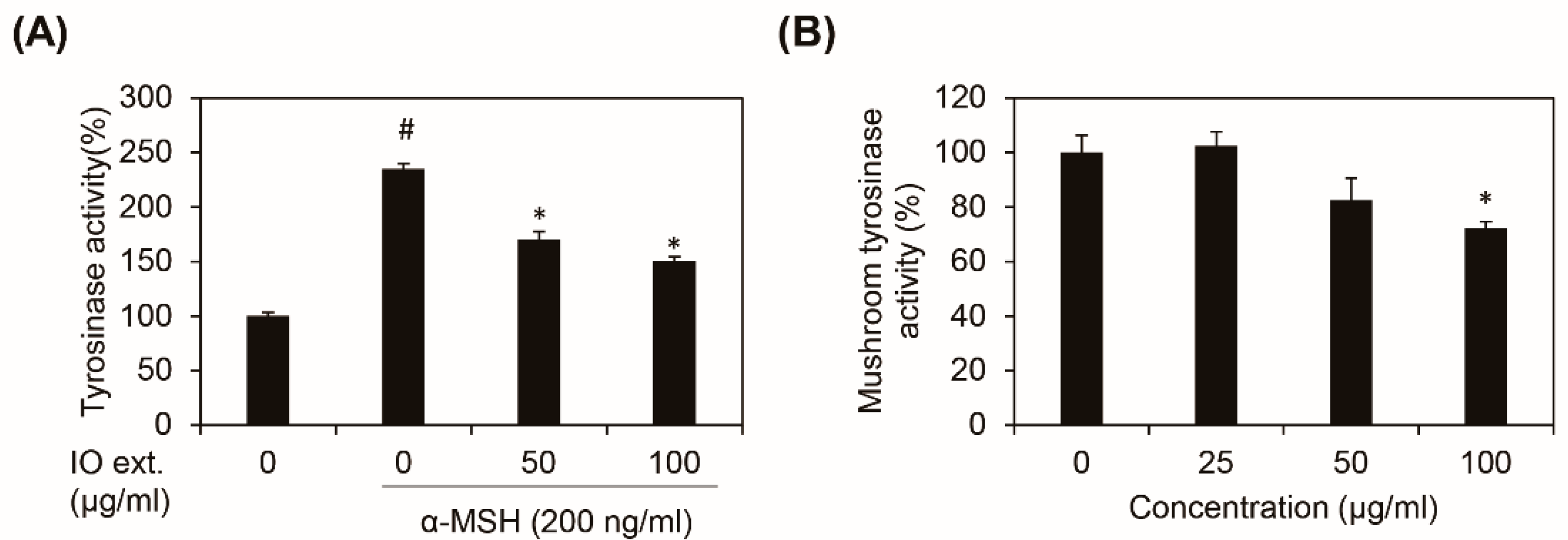
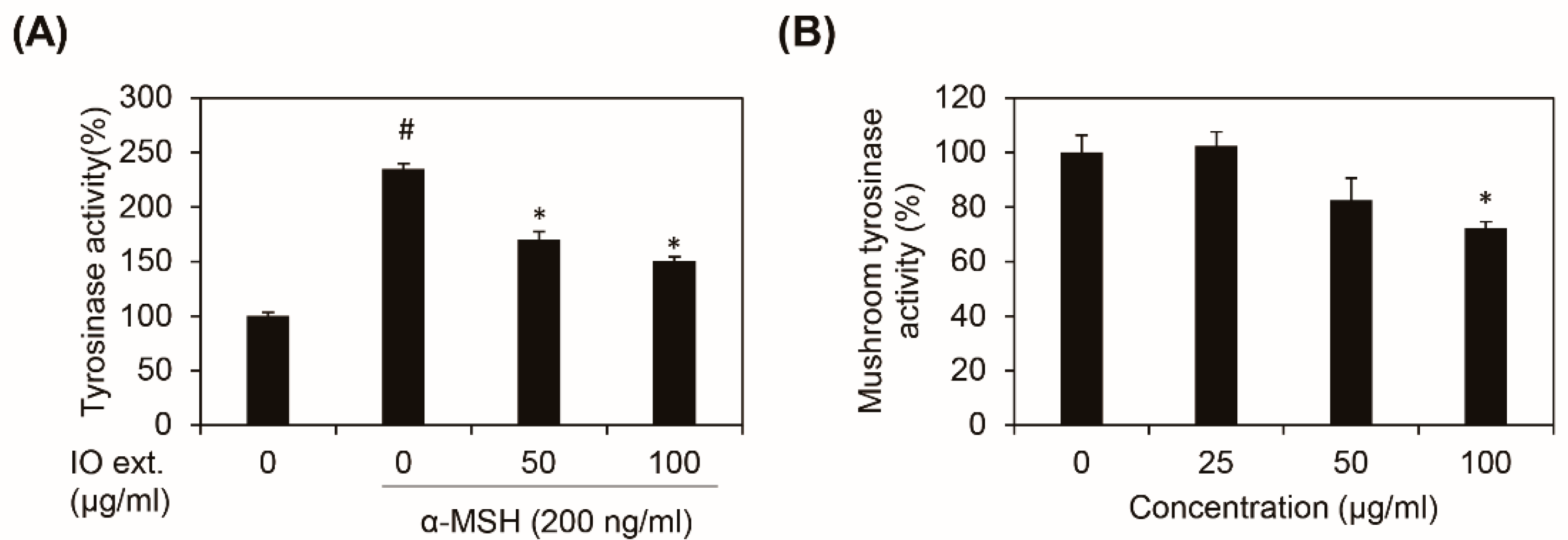
3.3. Inonotus obliquus Extract Directly Inhibits Tyrosinase Activity
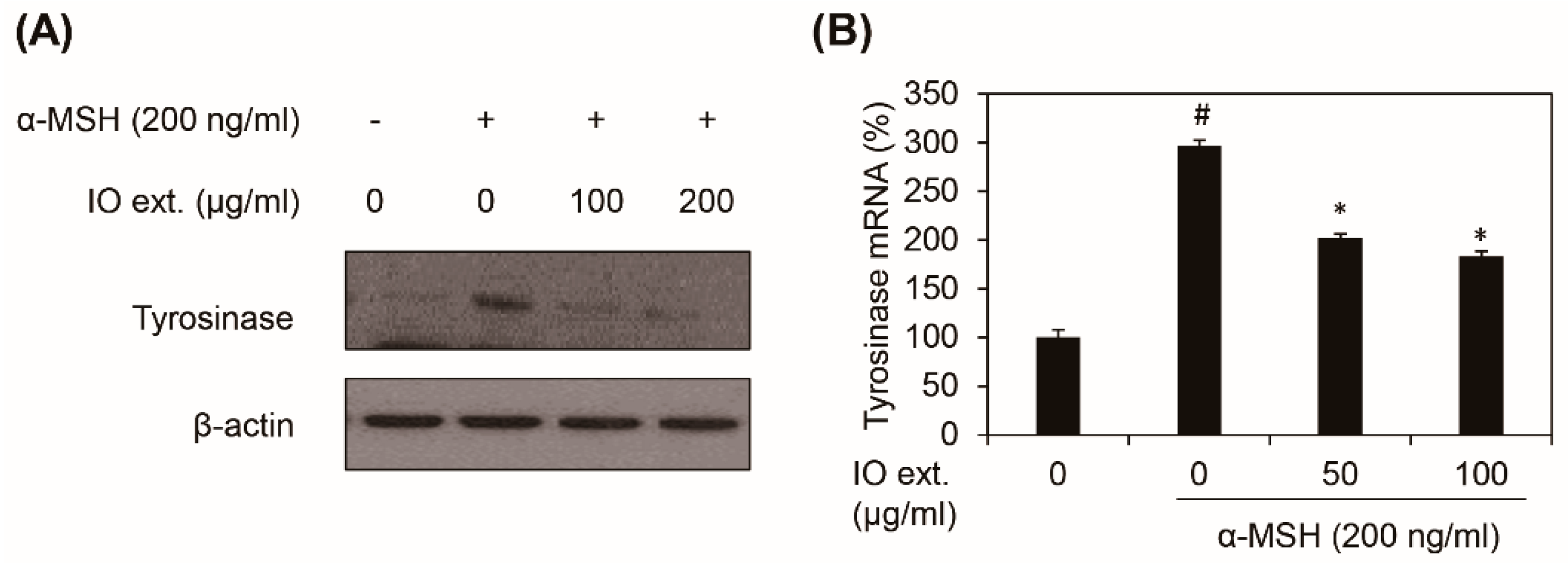
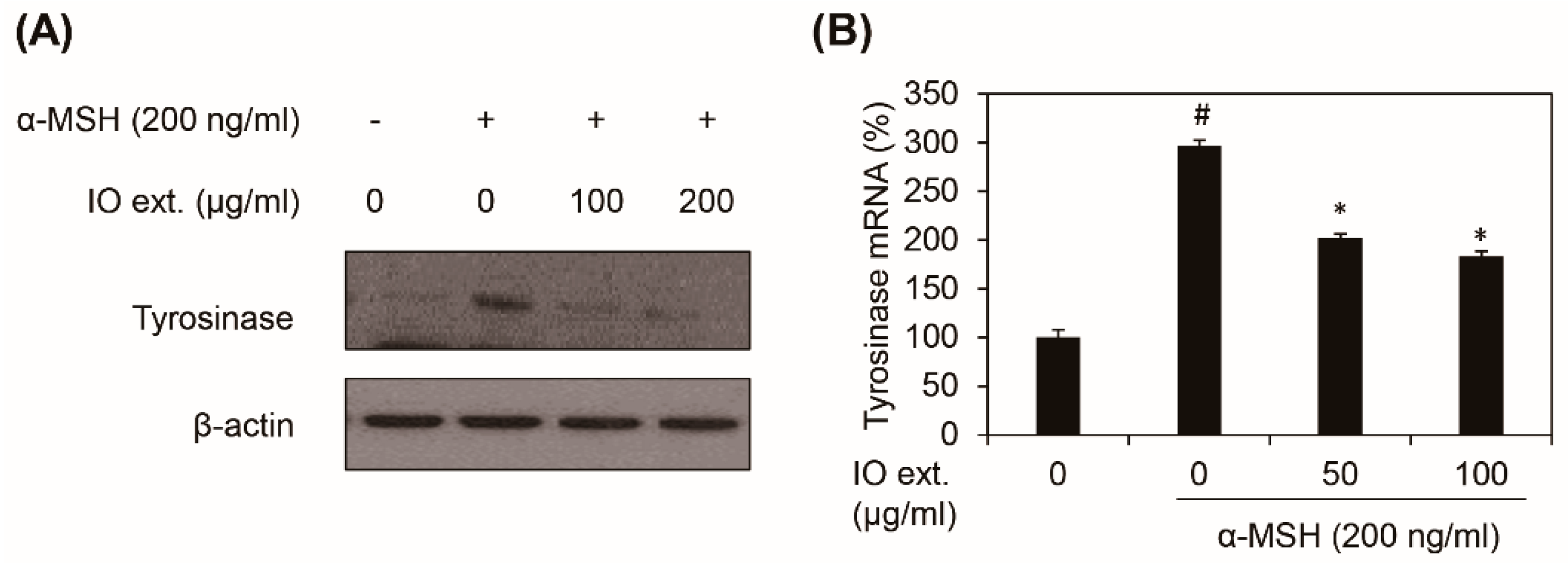
3.4. Inonotus obliquus Extract Downregulates α-MSH-induced Tyrosinase Expression
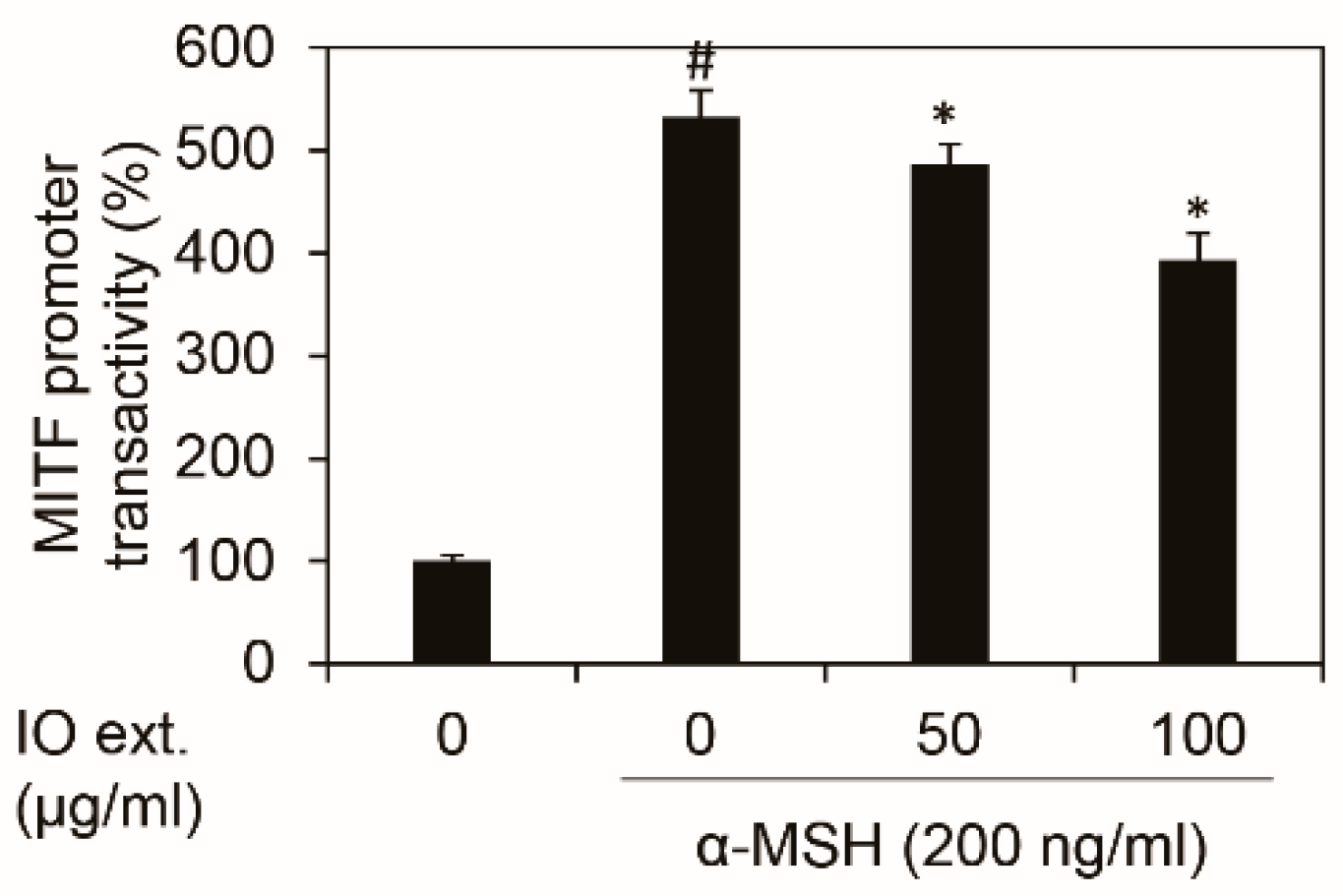
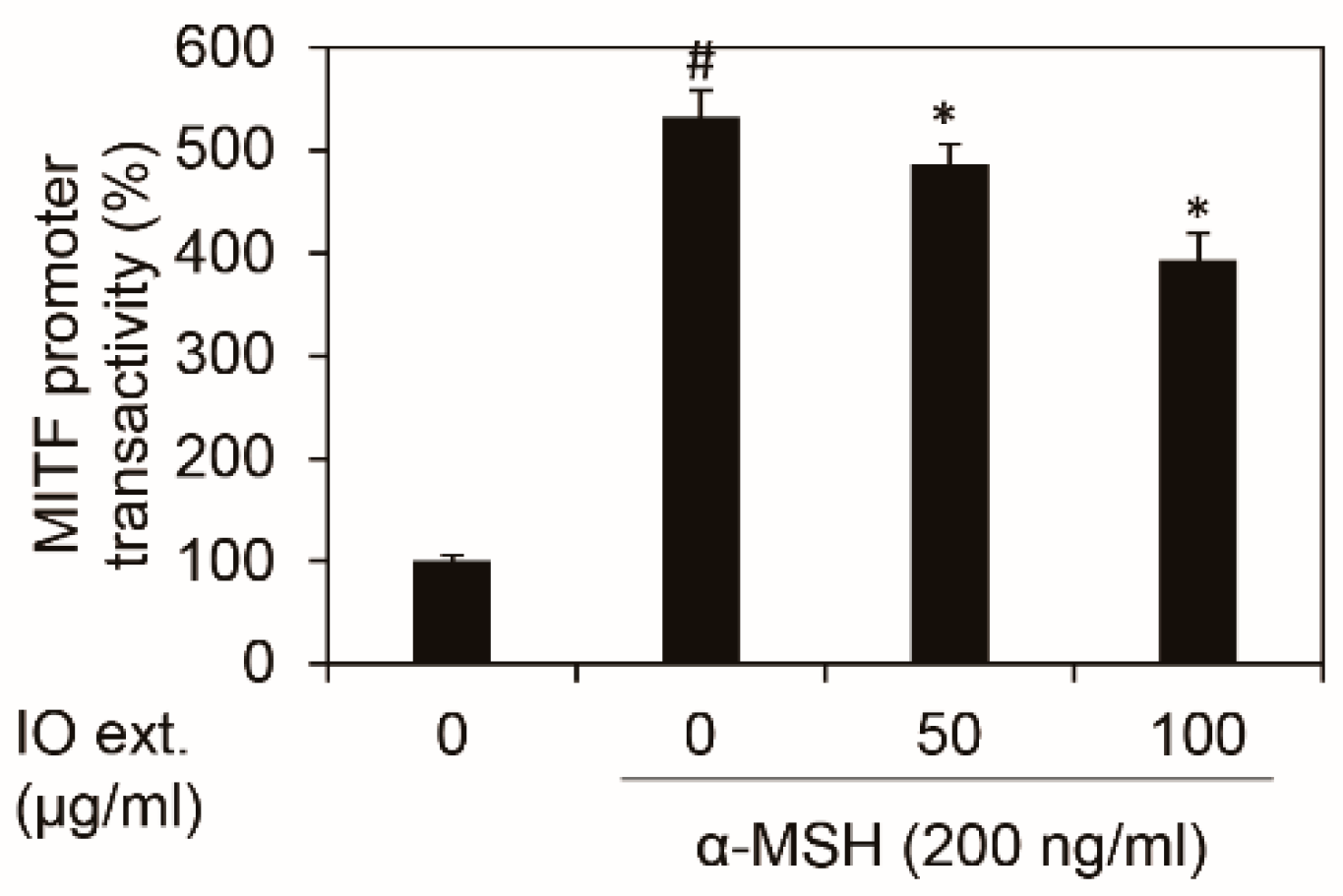
3.5. Inonotus obliquus Extract Downregulates Mitf Transcriptional Activity Stimulated by α-MSH
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seiberg, M. Keratinocyte-melanocyte interactions during melanosome transfer. Pigment. Cell Res. 2001, 14, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin Pigmentation in Mammalian Skin and Its Hormonal Regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef] [PubMed]
- Borovanský, J.; Wiley, I. Melanins and Melanosomes Biosynthesis, Biogenesis, Physiological, and Pathological Functions; John Wiley Distributor c2011: Weinheim/Baden-Wurttemberg, Germany, 2011. [Google Scholar]
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Valverde, P.; Healy, E.; Jackson, I.; Rees, J.L.; Thody, A.J. Variants of the melanocyte-stimulating hormone receptor gene are associated with red hair and fair skin in humans. Nat. Genet. 1995, 11, 328–330. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.S. Natural Melanogenesis Inhibitors Acting Through the Down-Regulation of Tyrosinase Activity. Materials 2012, 5, 1661–1685. [Google Scholar] [CrossRef]
- Li, H.X.; Park, J.U.; Su, X.D.; Kim, K.T.; Kang, J.S.; Kim, Y.R.; Kim, Y.H.; Yang, S.Y. Identification of Anti-Melanogenesis Constituents from Morus alba L. Leaves. Molecules 2018, 23, 2559. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Ji, J.; Park, S.H. Antiwrinkle and antimelanogenesis activity of the ethanol extracts of Lespedeza cuneata G. Don for development of the cosmeceutical ingredients. Food Sci. Nutr. 2018, 6, 1307–1316. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Ko, J.-H.; Kang, H.-K.; Kim, S.; Kang, C.I.; Lee, J.N.; Park, S.-M.; Hyun, C.-G. Antimelanogenic Effects of Polygonum tinctorium Flower Extract from Traditional Jeju Fermentation via Upregulation of Extracellular Signal-Regulated Kinase and Protein Kinase B Activation. Int. J. Mol. Sci. 2018, 19, 2895. [Google Scholar] [CrossRef] [PubMed]
- Shashkina, M.Y.; Shashkin, P.N.; Sergeev, A.V. Chemical and medicobiological properties of chaga (review). Pharm. Chem. J. 2006, 40, 560–568. [Google Scholar] [CrossRef]
- Ichimura, T.; Watanabe, O.; Maruyama, S. Inhibition of HIV-1 protease by water-soluble lignin-like substance from an edible mushroom, Fuscoporia obliqua. Biosci. Biotechnol. Biochem. 1998, 62, 575–577. [Google Scholar] [CrossRef] [PubMed]
- Arata, S.; Watanabe, J.; Maeda, M.; Yamamoto, M.; Matsuhashi, H.; Mochizuki, M.; Kagami, N.; Honda, K.; Inagaki, M. Continuous intake of the Chaga mushroom (Inonotus obliquus) aqueous extract suppresses cancer progression and maintains body temperature in mice. Heliyon 2016, 2, e00111. [Google Scholar] [CrossRef] [PubMed]
- Géry, A.; Dubreule, C.; André, V.; Rioult, J.P.; Bouchart, V.; Heutte, N.; Eldin de Pécoulas, P.; Krivomaz, T.; Garon, D. Chaga (Inonotus obliquus), a Future Potential Medicinal Fungus in Oncology? A Chemical Study and a Comparison of the Cytotoxicity Against Human Lung Adenocarcinoma Cells (A549) and Human Bronchial Epithelial Cells (BEAS-2B). Integr. Cancer Ther. 2018, 17, 832–843. [Google Scholar] [CrossRef] [PubMed]
- Lemieszek, M.K.; Langner, E.; Kaczor, J.; Kandefer-Szerszen, M.; Sanecka, B.; Mazurkiewicz, W.; Rzesky, W. Anticancer effects of fraction isolated from fruiting bodies of Chaga medicinal mushroom, Inonotus obliquus (Pers.:Fr.) Pilát (Aphyllophoromycetideae): In vitro studies. Int. J. Med. Mushrooms 2011, 13, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Zhang, Z.; Lei, Z.; Yang, Y.; Sugiura, N. Comparative studies of antioxidant activity and antiproliferative effect of hot water and ethanol extracts from the mush- room Inonotus obliquus. J. Biosci. Bioeng. 2009, 107, 42–48. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed]
- Videira, I.F.; Moura, D.F.; Magina, S. Mechanisms regulating melanogenesis. Anais Brasileiros de Dermatologia 2013, 88, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Brenner, M.; Hearing, V.J. The regulation of skin pigmentation. J. Biol. Chem. 2007, 282, 27557–27561. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Bahkali, A.H.; Moslem, M.A. Fungi—An unusual source for cosmetics. Fungal Divers. 2010, 43, 1–9. [Google Scholar] [CrossRef]
- Slominski, A.; Moellmann, G.; Kuklinska, E. L-Tyrosine, L-DOPA, and tyrosinase as positive regulators of the subcellular apparatus of melanogenesis in bomirski Ab amelanotic melanoma cells. Pigment. Cell Res. 1989, 2, 109–116. [Google Scholar] [CrossRef] [PubMed]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, E.J.; Cha, H.J. Inonotus obliquus Extract as An Inhibitor of α-MSH-Induced Melanogenesis in B16F10 Mouse Melanoma Cells. Cosmetics 2019, 6, 9. https://doi.org/10.3390/cosmetics6010009
Lee EJ, Cha HJ. Inonotus obliquus Extract as An Inhibitor of α-MSH-Induced Melanogenesis in B16F10 Mouse Melanoma Cells. Cosmetics. 2019; 6(1):9. https://doi.org/10.3390/cosmetics6010009
Chicago/Turabian StyleLee, Eun Ji, and Hwa Jun Cha. 2019. "Inonotus obliquus Extract as An Inhibitor of α-MSH-Induced Melanogenesis in B16F10 Mouse Melanoma Cells" Cosmetics 6, no. 1: 9. https://doi.org/10.3390/cosmetics6010009
APA StyleLee, E. J., & Cha, H. J. (2019). Inonotus obliquus Extract as An Inhibitor of α-MSH-Induced Melanogenesis in B16F10 Mouse Melanoma Cells. Cosmetics, 6(1), 9. https://doi.org/10.3390/cosmetics6010009