

Exploring Skin Longevity Pathways: Rosa hybrid Extract-Mediated AMP-Activated Protein Kinase Activation, Antioxidant, and Autophagic Mechanisms in Human Keratinocytes

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Extraction from Rosa hybrid Flowers
2.2. Volatile Molecules Analysis by GC-MS
2.3. Non-Volatile Molecules Analysis by UHPLC-MS
2.4. Primary Cultured Keratinocytes
2.5. Protein Extraction and Western Blot
2.6. Detection, Visualization, and Quantification of Biomarkers
2.7. Histogram Illustration and Statistical Analysis
3. Results
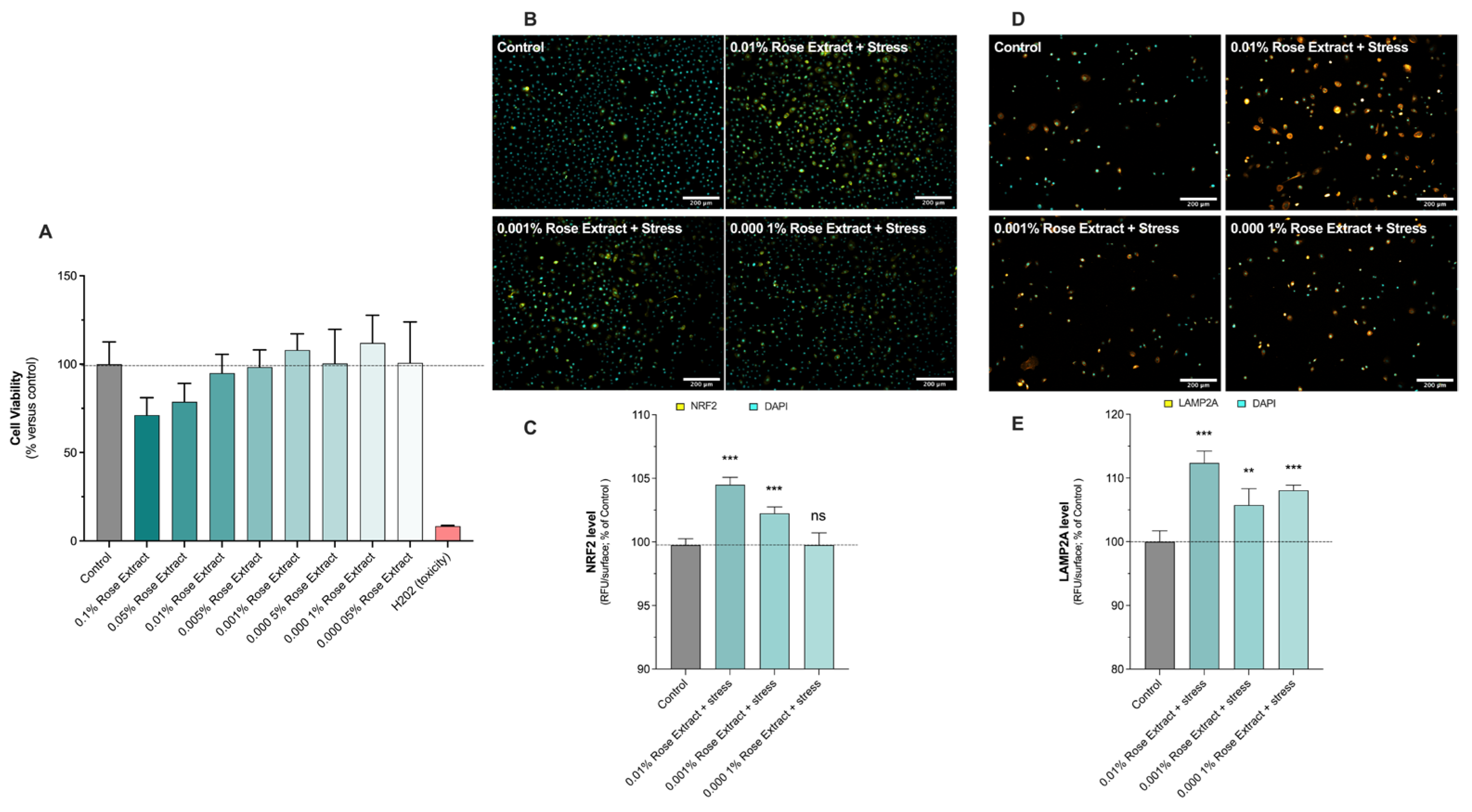
3.1. Rose Extract Induces No Cytotoxic Effect on Primary Human Keratinocytes and Stimulates Antioxidant Defense and Autophagy
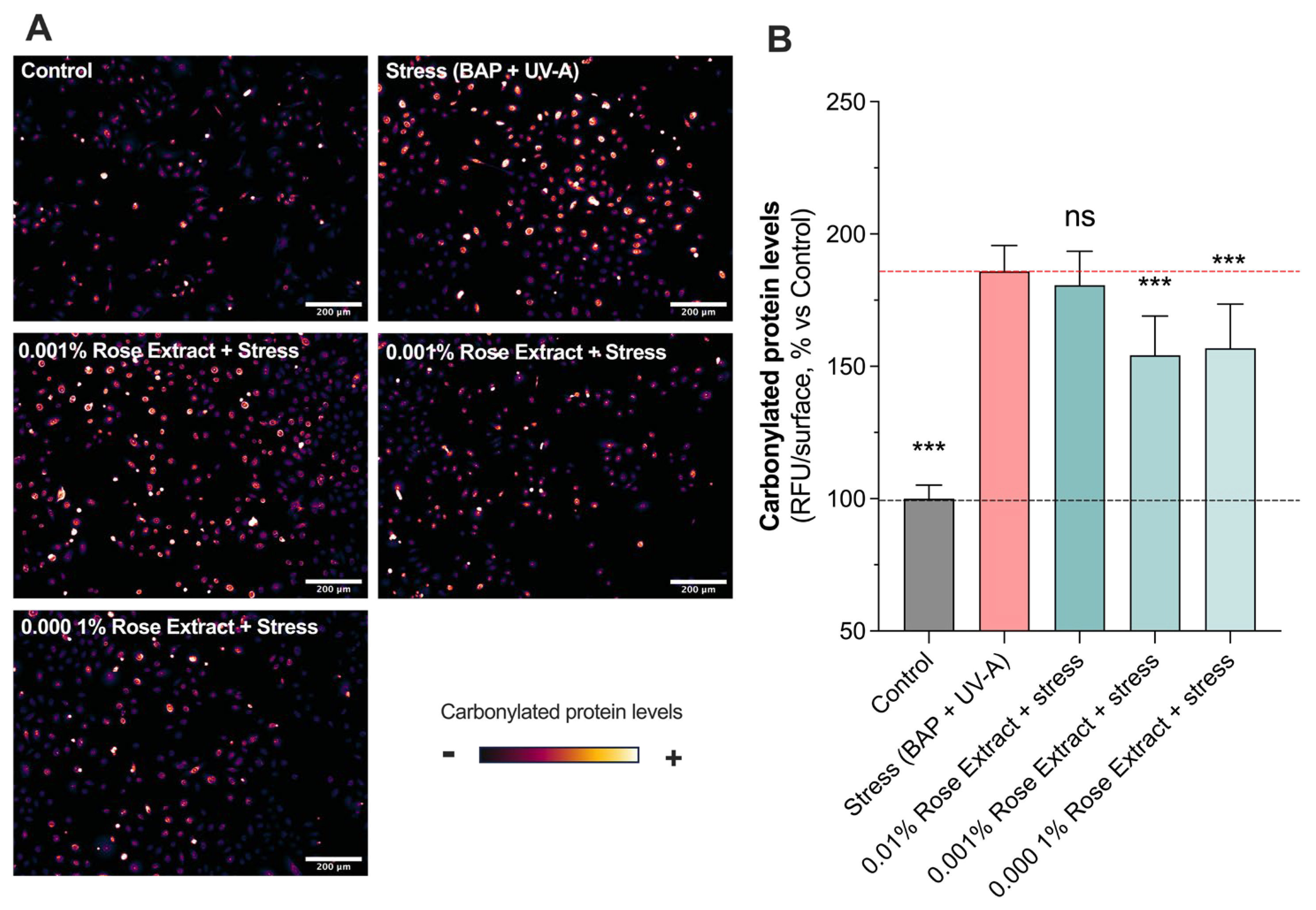
3.2. Treatment with Rose Extract Counteracts Pollution (BAP + UV-A)-Induced Oxidative Damage
3.3. Treatment with Rose Extract Counteracts Pollution (BAP + UV-A)-Mediated Cellular Senescence and Cell Growth Arrest
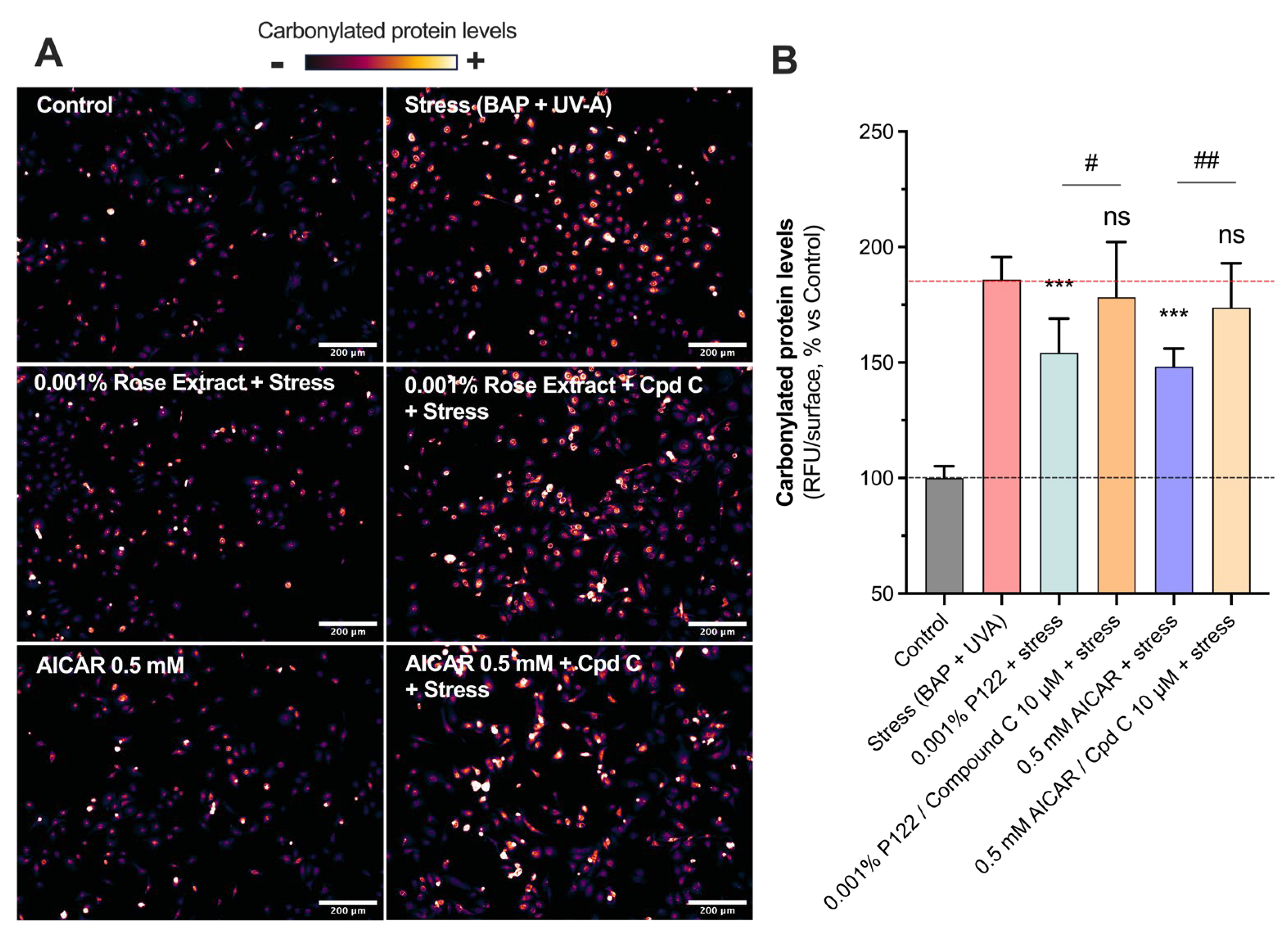
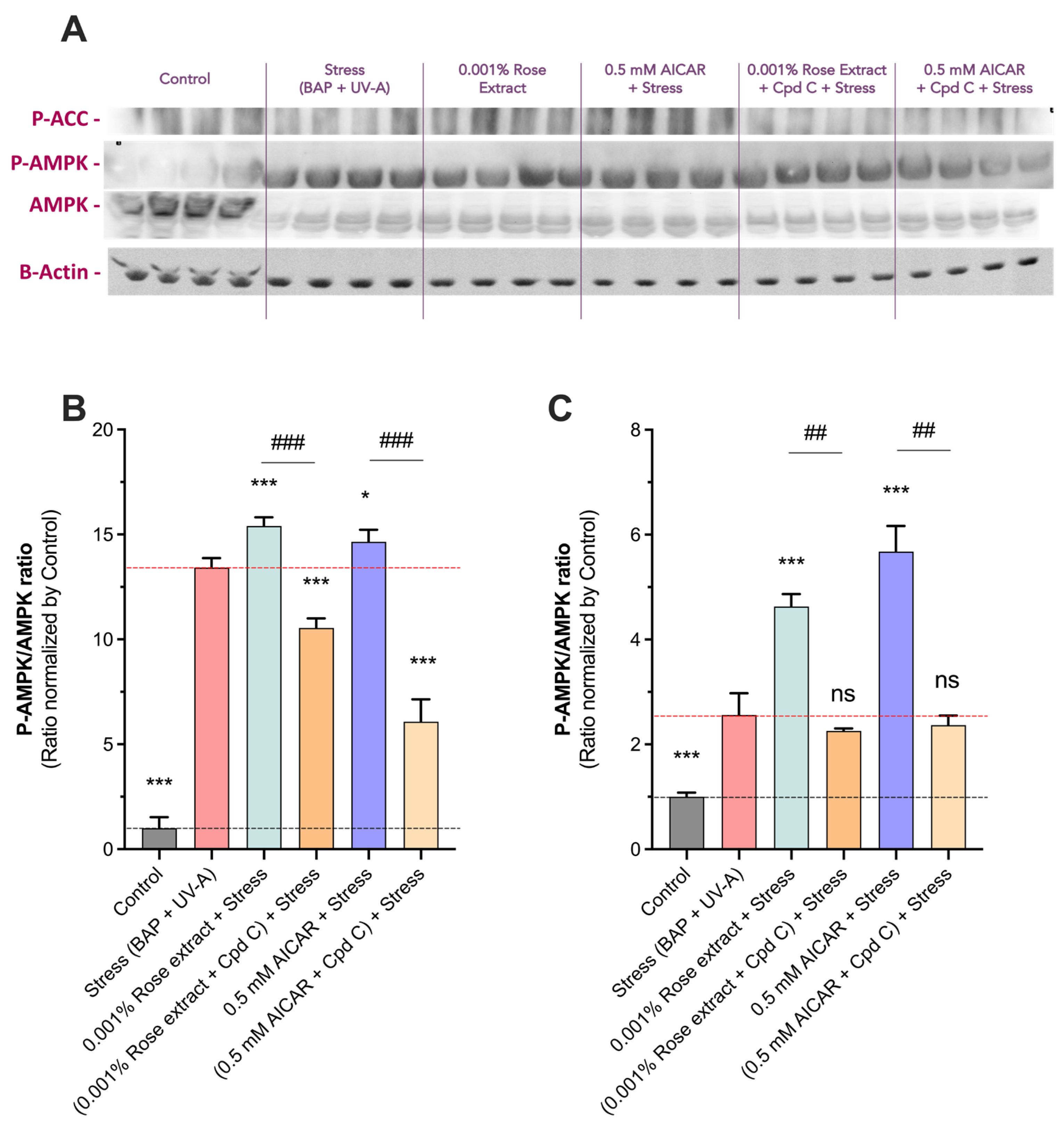
3.4. The Beneficial Antioxidant Effect of Rose Extract on Pollution (BAP + UV-A) Exposure Is Mediated by an AMPK-Dependent Mechanism
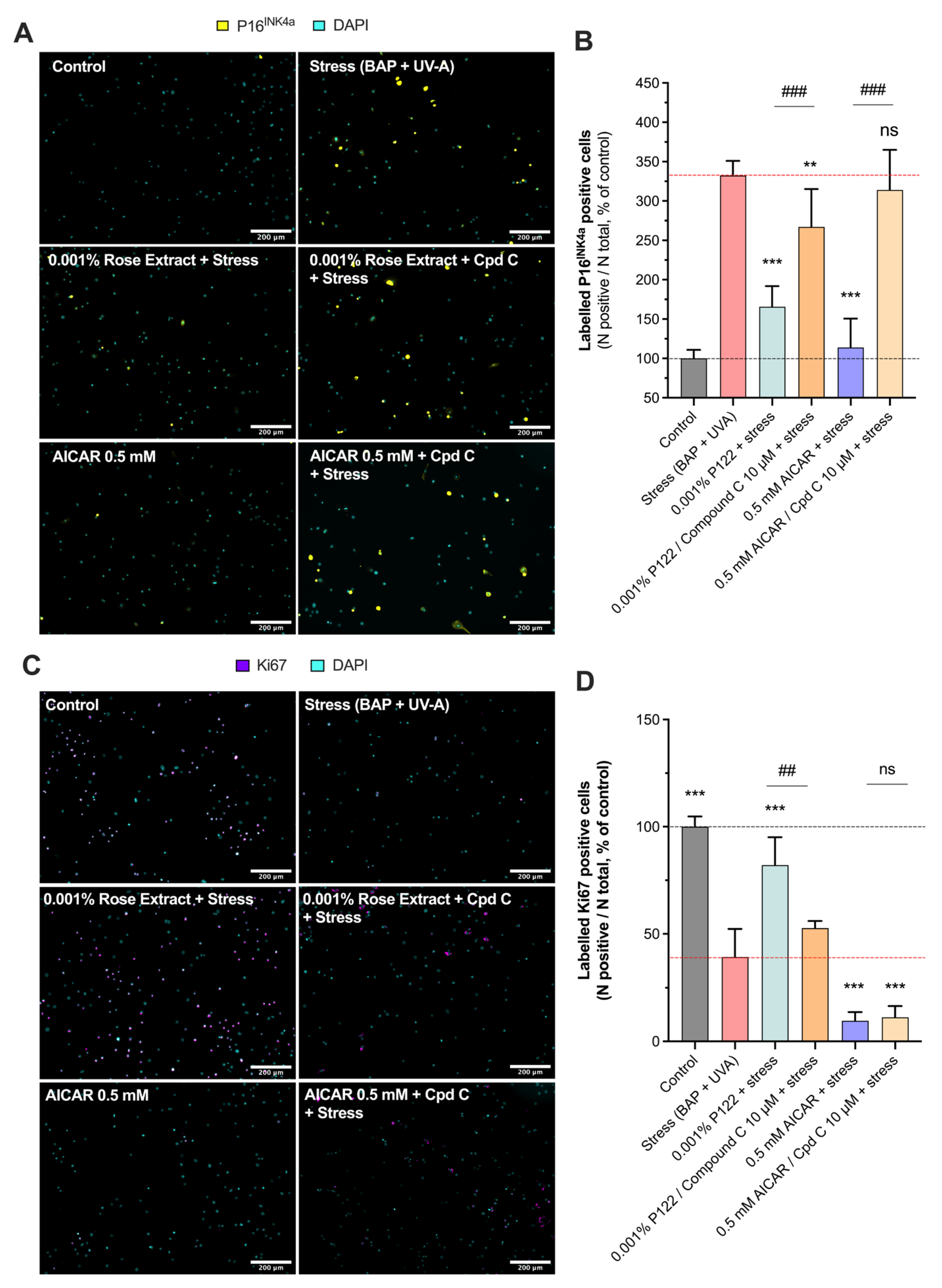
3.5. The Anti-Cell Senescence and Cell Growth Arrest Effect of Rose Extract Are Mediated by the AMPK Pathway
3.6. Rose Extract Stimulates the AMPK Pathway, Causing a Beneficial Effect
3.7. Volatile and Non-Volatile Compositions of Rose Extract
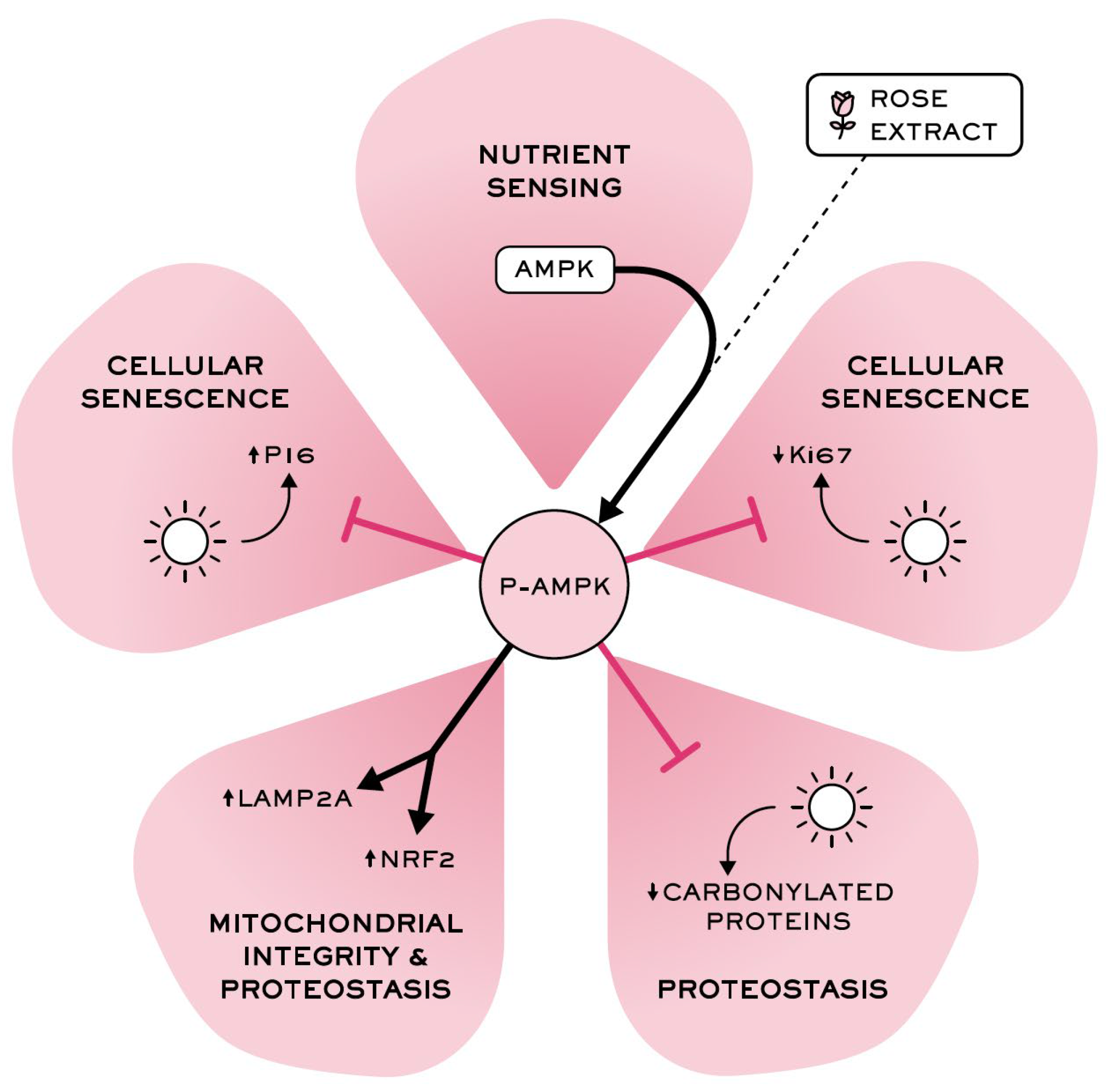
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The Skin Aging Exposome. J. Dermatol. Sci. 2017, 85, 152–161. [Google Scholar] [CrossRef]
- Xu, W.; Luo, Y.; Yin, J.; Huang, M.; Luo, F. Targeting AMPK Signaling by Polyphenols: A Novel Strategy for Tackling Aging. Food Funct. 2023, 14, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Hamon, M.-P.; Ahmed, E.K.; Baraibar, M.A.; Friguet, B. Proteome Oxidative Modifications and Impairment of Specific Metabolic Pathways During Cellular Senescence and Aging. Proteomics 2020, 20, e1800421. [Google Scholar] [CrossRef] [PubMed]
- Celebi Sozener, Z.; Ozdel Ozturk, B.; Cerci, P.; Turk, M.; Gorgulu Akin, B.; Akdis, M.; Altiner, S.; Ozbey, U.; Ogulur, I.; Mitamura, Y.; et al. Epithelial Barrier Hypothesis: Effect of the External Exposome on the Microbiome and Epithelial Barriers in Allergic Disease. Allergy 2022, 77, 1418–1449. [Google Scholar] [CrossRef] [PubMed]
- Vineis, P.; Robinson, O.; Chadeau-Hyam, M.; Dehghan, A.; Mudway, I.; Dagnino, S. What Is New in the Exposome? Environ. Int. 2020, 143, 105887. [Google Scholar] [CrossRef]
- Azzimonti, B.; Ballacchino, C.; Zanetta, P.; Cucci, M.A.; Monge, C.; Grattarola, M.; Dianzani, C.; Barrera, G.; Pizzimenti, S. Microbiota, Oxidative Stress, and Skin Cancer: An Unexpected Triangle. Antioxidants 2023, 12, 546. [Google Scholar] [CrossRef]
- Md Jaffri, J. Reactive Oxygen Species and Antioxidant System in Selected Skin Disorders. Malays. J. Med. Sci. 2023, 30, 7–20. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of Aging: An Expanding Universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef]
- Baraibar, M.A.; Friguet, B. Oxidative Proteome Modifications Target Specific Cellular Pathways during Oxidative Stress, Cellular Senescence and Aging. Exp. Gerontol. 2013, 48, 620–625. [Google Scholar] [CrossRef]
- Baraibar, M.A.; Ladouce, R.; Friguet, B. Proteomic Quantification and Identification of Carbonylated Proteins upon Oxidative Stress and during Cellular Aging. J. Proteom. 2013, 92, 63–70. [Google Scholar] [CrossRef]
- Le Boulch, M.; Ahmed, E.K.; Rogowska-Wrzesinska, A.; Baraibar, M.A.; Friguet, B. Proteome Oxidative Carbonylation during Oxidative Stress-Induced Premature Senescence of WI-38 Human Fibroblasts. Mech. Ageing Dev. 2018, 170, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Pajares, M.; Rojo, A.I.; Arias, E.; Díaz-Carretero, A.; Cuervo, A.M.; Cuadrado, A. Transcription Factor NFE2L2/NRF2 Modulates Chaperone-Mediated Autophagy through the Regulation of LAMP2A. Autophagy 2018, 14, 1310–1322. [Google Scholar] [CrossRef] [PubMed]
- Mo, C.; Wang, L.; Zhang, J.; Numazawa, S.; Tang, H.; Tang, X.; Han, X.; Li, J.; Yang, M.; Wang, Z.; et al. The Crosstalk between Nrf2 and AMPK Signal Pathways Is Important for the Anti-Inflammatory Effect of Berberine in LPS-Stimulated Macrophages and Endotoxin-Shocked Mice. Antioxid. Redox Signal 2014, 20, 574–588. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Abramov, A.Y. The Emerging Role of Nrf2 in Mitochondrial Function. Free Radic. Biol. Med. 2015, 88, 179–188. [Google Scholar] [CrossRef]
- Liu, X.; Liao, X.; Rao, X.; Wang, B.; Zhang, J.; Xu, G.; Jiang, X.; Qin, X.; Chen, C.; Zou, Z. The Lysosomal Membrane Protein LAMP-2 Is Dispensable for PINK1/Parkin-Mediated Mitophagy. FEBS Lett. 2020, 594, 823–840. [Google Scholar] [CrossRef]
- Ng, T.B.; Gao, W.; Li, L.; Niu, S.M.; Zhao, L.; Liu, J.; Shi, L.S.; Fu, M.; Liu, F. Rose (Rosa rugosa)-Flower Extract Increases the Activities of Antioxidant Enzymes and Their Gene Expression and Reduces Lipid Peroxidation. Biochem. Cell Biol. 2005, 83, 78–85. [Google Scholar] [CrossRef]
- Sparkman, O.D. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy Robert P. Adams. J. Am. Soc. Mass. Spectrom. 2005, 16, 1902–1903. [Google Scholar] [CrossRef]
- Dührkop, K.; Shen, H.; Meusel, M.; Rousu, J.; Böcker, S. Searching Molecular Structure Databases with Tandem Mass Spectra Using CSI:FingerID. Proc. Natl. Acad. Sci. USA 2015, 112, 12580–12585. [Google Scholar] [CrossRef]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A Rapid Tool for Turning Tandem Mass Spectra into Metabolite Structure Information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef]
- Baker, D.J.; Wijshake, T.; Tchkonia, T.; LeBrasseur, N.K.; Childs, B.G.; van de Sluis, B.; Kirkland, J.L.; van Deursen, J.M. Clearance of p16Ink4a-Positive Senescent Cells Delays Ageing-Associated Disorders. Nature 2011, 479, 232–236. [Google Scholar] [CrossRef]
- Safwan-Zaiter, H.; Wagner, N.; Wagner, K.-D. P16INK4A—More Than a Senescence Marker. Life 2022, 12, 1332. [Google Scholar] [CrossRef]
- Alessio, N.; Aprile, D.; Cappabianca, S.; Peluso, G.; Di Bernardo, G.; Galderisi, U. Different Stages of Quiescence, Senescence, and Cell Stress Identified by Molecular Algorithm Based on the Expression of Ki67, RPS6, and Beta-Galactosidase Activity. Int. J. Mol. Sci. 2021, 22, 3102. [Google Scholar] [CrossRef]
- Han, X.; Tai, H.; Wang, X.; Wang, Z.; Zhou, J.; Wei, X.; Ding, Y.; Gong, H.; Mo, C.; Zhang, J.; et al. AMPK Activation Protects Cells from Oxidative Stress-Induced Senescence via Autophagic Flux Restoration and Intracellular NAD+ Elevation. Aging Cell 2016, 15, 416–427. [Google Scholar] [CrossRef]
- Wang, S.; Li, H.; Yuan, M.; Fan, H.; Cai, Z. Role of AMPK in Autophagy. Front. Physiol. 2023, 13, 2994. [Google Scholar]
- Philippe, C.; Pinson, B.; Dompierre, J.; Pantesco, V.; Viollet, B.; Daignan-Fornier, B.; Moenner, M. AICAR Antiproliferative Properties Involve the AMPK-Independent Activation of the Tumor Suppressors LATS 1 and 2. Neoplasia 2018, 20, 555–562. [Google Scholar] [CrossRef]
- Albrecht, D.; Ceschin, J.; Dompierre, J.; Gueniot, F.; Pinson, B.; Daignan-Fornier, B. Chemo-Genetic Interactions Between Histone Modification and the Antiproliferation Drug AICAR Are Conserved in Yeast and Humans. Genetics 2016, 204, 1447–1460. [Google Scholar] [CrossRef]
- Shin, E.J.; Han, A.; Lee, M.; Song, Y.-R.; Lee, K.M.; Nam, T.-G.; Lee, P.; Lee, S.-Y.; Lim, T.-G. Extraction Conditions for Rosa Gallica Petal Extracts with Anti-Skin Aging Activities. Food Sci. Biotechnol. 2019, 28, 1439–1446. [Google Scholar] [CrossRef]
- Hipp, M.S.; Kasturi, P.; Hartl, F.U. The Proteostasis Network and Its Decline in Ageing. Nat. Rev. Mol. Cell Biol. 2019, 20, 421–435. [Google Scholar] [CrossRef]
- Jeong, D.; Qomaladewi, N.P.; Lee, J.; Park, S.H.; Cho, J.Y. The Role of Autophagy in Skin Fibroblasts, Keratinocytes, Melanocytes, and Epidermal Stem Cells. J. Investig. Dermatol. 2020, 140, 1691–1697. [Google Scholar] [CrossRef]
- Kaushik, S.; Cuervo, A.M. The Coming of Age of Chaperone-Mediated Autophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 365–381. [Google Scholar] [CrossRef] [PubMed]
- Raz, Y.; Guerrero-Ros, I.; Maier, A.; Slagboom, P.E.; Atzmon, G.; Barzilai, N.; Macian, F. Activation-Induced Autophagy Is Preserved in CD4+ T-Cells in Familial Longevity. J. Gerontol. Ser. A 2017, 72, 1201–1206. [Google Scholar] [CrossRef]
- Ansary, T.M.; Hossain, M.R.; Kamiya, K.; Komine, M.; Ohtsuki, M. Inflammatory Molecules Associated with Ultraviolet Radiation-Mediated Skin Aging. Int. J. Mol. Sci. 2021, 22, 3974. [Google Scholar] [CrossRef]
- Gonos, E.S.; Kapetanou, M.; Sereikaite, J.; Bartosz, G.; Naparło, K.; Grzesik, M.; Sadowska-Bartosz, I. Origin and Pathophysiology of Protein Carbonylation, Nitration and Chlorination in Age-Related Brain Diseases and Aging. Aging 2018, 10, 868–901. [Google Scholar] [CrossRef]
- Nyström, T. Role of Oxidative Carbonylation in Protein Quality Control and Senescence. EMBO J. 2005, 24, 1311–1317. [Google Scholar] [CrossRef]
- Low, E.; Alimohammadiha, G.; Smith, L.A.; Costello, L.F.; Przyborski, S.A.; von Zglinicki, T.; Miwa, S. How Good Is the Evidence That Cellular Senescence Causes Skin Ageing? Ageing Res. Rev. 2021, 71, 101456. [Google Scholar] [CrossRef]
- Baker, D.J.; Childs, B.G.; Durik, M.; Wijers, M.E.; Sieben, C.J.; Zhong, J.; Saltness, R.A.; Jeganathan, K.B.; Verzosa, G.C.; Pezeshki, A.; et al. Naturally Occurring p16Ink4a-Positive Cells Shorten Healthy Lifespan. Nature 2016, 530, 184–189. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of Metabolism and Mitochondrial Homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef]
- Smitha Grace, S.R.; Chandran, G.; Chauhan, J.B. Terpenoids: An Activator of “Fuel-Sensing Enzyme AMPK” with Special Emphasis on Antidiabetic Activity. In Plant and Human Health, Volume 2: Phytochemistry and Molecular Aspects; Ozturk, M., Hakeem, K.R., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 227–244. ISBN 978-3-030-03344-6. [Google Scholar]
- The Potential Therapeutic Value of Terpenes. Available online: https://accscience.com/journal/ITPS/7/3/10.36922/itps.0332 (accessed on 11 February 2025).
- Izcara, S.; Perestrelo, R.; Morante-Zarcero, S.; Sierra, I.; Câmara, J.S. Volatilomic Fingerprinting from Edible Flowers. Unravelling Some Impact Compounds Behind Its Attractiveness. Food Biosci. 2022, 50, 102188. [Google Scholar] [CrossRef]
- Tamilmani, P.; Sathibabu Uddandrao, V.V.; Chandrasekaran, P.; Saravanan, G.; Brahma Naidu, P.; Sengottuvelu, S.; Vadivukkarasi, S. Linalool Attenuates Lipid Accumulation and Oxidative Stress in Metabolic Dysfunction-Associated Steatotic Liver Disease via Sirt1/Akt/PPRA-α/AMPK and Nrf-2/HO-1 Signaling Pathways. Clin. Res. Hepatol. Gastroenterol. 2023, 47, 102231. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, Z.; Riethoven, J.-J.; Xia, Y.; Miner, J.; Fromm, M. Conjugated Linoleic Acid Activates AMP-Activated Protein Kinase and Reduces Adiposity More Effectively When Used with Metformin in Mice. J. Nutr. 2009, 139, 2244–2251. [Google Scholar] [CrossRef] [PubMed]
- Watt, M.J.; Steinberg, G.R.; Chen, Z.-P.; Kemp, B.E.; Febbraio, M.A. Fatty Acids Stimulate AMP-Activated Protein Kinase and Enhance Fatty Acid Oxidation in L6 Myotubes. J. Physiol. 2006, 574, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Ido, Y.; Duranton, A.; Lan, F.; Weikel, K.A.; Breton, L.; Ruderman, N.B. Resveratrol Prevents Oxidative Stress-Induced Senescence and Proliferative Dysfunction by Activating the AMPK-FOXO3 Cascade in Cultured Primary Human Keratinocytes. PLoS ONE 2015, 10, e0115341. [Google Scholar] [CrossRef] [PubMed]
- Lan, F.; Weikel, K.A.; Cacicedo, J.M.; Ido, Y. Resveratrol-Induced AMP-Activated Protein Kinase Activation Is Cell-Type Dependent: Lessons from Basic Research for Clinical Application. Nutrients 2017, 9, 751. [Google Scholar] [CrossRef]
- Lee, S.-H.; Seo, H.-S.; Seo, S.J.; Kim, C.-D.; Hong, S.-P. Screening of Plant-Derived Natural Extracts to Identify a Candidate Extract Capable of Enhancing Lipid Synthesis in Keratinocytes. Ann. Dermatol. 2022, 34, 331–339. [Google Scholar] [CrossRef]
- Čižmárová, B.; Hubková, B.; Tomečková, V.; Birková, A. Flavonoids as Promising Natural Compounds in the Prevention and Treatment of Selected Skin Diseases. Int. J. Mol. Sci. 2023, 24, 6324. [Google Scholar] [CrossRef]
- Li, J.; Huang, Y.-C.; Deng, J.-M.; Yu, M.; Zouboulis, C.C.; Wang, G.-L.; Wang, J. Tea (Camellia sinensis) Seed Saponins Act as Sebosuppression Agents via the AMPK/mTOR Pathway. J. Cosmet. Dermatol. 2025, 24, e16793. [Google Scholar] [CrossRef]
- Prasanth, M.I.; Sivamaruthi, B.S.; Chaiyasut, C.; Tencomnao, T. A Review of the Role of Green Tea (Camellia sinensis) in Antiphotoaging, Stress Resistance, Neuroprotection, and Autophagy. Nutrients 2019, 11, 474. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host | Target | Provider/Ref. | Application |
|---|---|---|---|
| Rabbit | P_ACCS79 | Abcam (Paris, France) ab222774 | Western blot |
| Rabbit | P_AMPKT183/T172 | Abcam ab23875 | Western blot |
| Mouse | AMPK | Abcam ab110036 | Western blot |
| Rabbit | Actin | Abcam ab179467 | Western blot |
| Rabbit | P16 | Invitrogen (Les Ulis, France) MA5–27905 | In situ visualization |
| Rabbit | LAMP2A | Invitrogen 10332473 | In situ visualization |
| Rabbit | Ki67 | Abcam ab92742 | In situ visualization |
| Rabbit | NRF2 | Abcam ab137550 | In situ visualization |
| Goat | Anti-mouse Alexafluor 488 | Invitrogen A11001 | Secondary antibody |
| Goat | Anti-Rabbit Alexafluor 467 | Invitrogen A21244 | Secondary antibody |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gouin, O.; Cavagnino, A.; Azadiguian, G.; Jäger, S.; Comte, G.; Bendahmane, M.; Breton, L.; Baraibar, M.A.; Black, A.F. Exploring Skin Longevity Pathways: Rosa hybrid Extract-Mediated AMP-Activated Protein Kinase Activation, Antioxidant, and Autophagic Mechanisms in Human Keratinocytes. Cosmetics 2025, 12, 57. https://doi.org/10.3390/cosmetics12020057
Gouin O, Cavagnino A, Azadiguian G, Jäger S, Comte G, Bendahmane M, Breton L, Baraibar MA, Black AF. Exploring Skin Longevity Pathways: Rosa hybrid Extract-Mediated AMP-Activated Protein Kinase Activation, Antioxidant, and Autophagic Mechanisms in Human Keratinocytes. Cosmetics. 2025; 12(2):57. https://doi.org/10.3390/cosmetics12020057
Chicago/Turabian StyleGouin, Olivier, Andrea Cavagnino, Gayané Azadiguian, Sibylle Jäger, Gilles Comte, Mohammed Bendahmane, Lionel Breton, Martin A. Baraibar, and Annie F. Black. 2025. "Exploring Skin Longevity Pathways: Rosa hybrid Extract-Mediated AMP-Activated Protein Kinase Activation, Antioxidant, and Autophagic Mechanisms in Human Keratinocytes" Cosmetics 12, no. 2: 57. https://doi.org/10.3390/cosmetics12020057
APA StyleGouin, O., Cavagnino, A., Azadiguian, G., Jäger, S., Comte, G., Bendahmane, M., Breton, L., Baraibar, M. A., & Black, A. F. (2025). Exploring Skin Longevity Pathways: Rosa hybrid Extract-Mediated AMP-Activated Protein Kinase Activation, Antioxidant, and Autophagic Mechanisms in Human Keratinocytes. Cosmetics, 12(2), 57. https://doi.org/10.3390/cosmetics12020057