1. Introduction
Analog CMOS hardware has the potential to reduce energy consumption of deep neural networks by orders of magnitude, but the in situ training of networks implemented on such hardware is challenging. Once the chip has been programmed with the correct weight values for a task, typically no further learning occurs. We introduce a biologically-inspired knowledge transfer approach for neural networks that offers potential for in situ learning on the physical chip. In our method, the weight matrices of a spiking neural network [
1,
2,
3,
4,
5] are initialized with values learned via offline (i.e., off-chip) methods, and the system is exposed to an analogous—but distinct—learning task. The bias inputs of the chip’s spiking neurons are manipulated such that the network’s outputs adapt to the new learning task.
This approach has applications for autonomous, power-constrained devices that must adapt to unanticipated circumstances, including vision and navigation in unmanned aerial vehicles (UAVs) deployed into unpredictable environments; fine-grained haptic controls for robotic manipulators; dynamically adaptive prosthetic devices; and bio-cybernetic interfaces. In these real-world domains, the system must deploy with initial knowledge relevant to its target environment, then adapt to near-optimal behavior given minimal training examples, a feat beyond the capability of current learning algorithms or hardware platforms. Neuromodulatory tuning offers a path toward implementing such abilities on physical CMOS chips. The key contributions of our work are as follows:
We introduce a novel transfer learning variant, called neuromodulatory tuning, that is able to match the performance of traditional fine-tuning approaches with orders of magnitude fewer weight updates. This lends itself naturally to easier, lower power implementation on physical chips, especially because the proposed CMOS implementation of our the fine-tuning method does not involve writing to memory hardware.
We provide a biologically-inspired motivation for this tuning method based on recent findings in neuroscience, and discuss additional insights gleaned from modulatory neurotransmitter behaviors in biological brains that may prove valuable for neuromorphic computing hardware.
We demonstrate in both traditional (non-analog) feed-forward architectures and spiking neural network simulations that neuromodulatory tuning methods are able to approach or exceed the performance of traditional fine-tuning methods on a number of transfer learning tasks in the domain of image recognition, while overall task performance must still be improved, the trends and potential of the method are encouraging.
We outline the mechanisms by which neuromodulatory tuning can feasibly be implemented on CMOS hardware. We present an analog spiking neuron with neuromodulatory tuning capabilities. Post-layout simulations demonstrate energy/spike rates as low as 1.08 pJ.
The remainder of this paper adheres to the following structure: We begin by providing a general background on transfer learning, artificial neural networks, and neuromorphic hardware in
Section 2. We then outline the motivating principles and neurobiological foundations of the current work (
Section 3.1) and present our biologically inspired tuning method (
Section 3.2). A preliminary analysis follows (
Section 4), showing performance comparisons of NT versus TFT in digital computation environments across a variety of learning rates and transfer tasks. Lastly, we present our spiking neuron design (
Section 5) with confirming evidence that our neuromodulatory tuning method can be used as an acceptable proxy for traditional fine-tuning in analog CMOS environments (
Section 6). Conclusions are presented in
Section 7.
4. Modeling and Analysis
We first probe the capabilities and weaknesses of neuromodulatory tuning (NT) in a traditional deep learning setting. Using a pre-trained VGG-19 network architecture, we fine-tune the model on three image recognition tasks. VGG-19 was trained on ImageNet [
56], an image classification dataset composed of 1000 different image categories. The first dataset we use in our evaluation is STL-10, a subset of ImageNet with only 10 image categories [
57]. We expect traditional fine-tuning (TFT) and neuromodulatory tuning (NT) to achieve high accuracies on STL-10 since the data is a subset of the original training data. Next, we evaluate neuromodulatory tuning on a more difficult food classification task, Food-11 [
58], which contains images of 11 different types of food none of which match any of ImageNet’s classes. Finally, we examine the capability of neuromodulatory tuning to learn blood cell classification (BCCD) [
59], which is a task very distinct from ImageNet containing 4 classes of blood cells images. We hypothesize that as the difficulty of the tasks increase, NT will be less effective in tuning the model to solve the given task, but still comparable to TFT.
For simplicity, fine tuning is applied only to the VGG-19 classifier layers, a process which lowers the fine-tuned classification accuracy but facilitates our comparisons to spiking neural network implementations in
Section 5.1. Additionally, it is common practice to only fine tune select layers of VGG models in recent literature [
60,
61]. We then apply neuromodulatory tuning to the same layers that were fine-tuned (i.e., classification layers only) and compare the performance of traditional fine tuning (TFT) to neuromodulatory tuning (NT), as shown in
Table 3.
To visualize the comparison between neuromodulatory tuning (NT) and traditional fine-tuning (TFT), we create two model architectures, one with hyper-parameters configured for NT and the other for TFT. We use the existing train and validation partitions in the STL-10, Food-11 and BCCD datasets to train and evaluate the classifier layers of the pre-trained VGG-19 model. We resize the data in each of the datasets to be images of size 256 × 256 to be compatible with VGG-19. Using an NVIDIA GeForce RTX 2080 Ti GPU, we fine-tune both models for 10 epochs, with various training set sizes and learning rates.
We set the batch size to 64 training instances in all experiments with neural networks. The effect of batch size on model performance has been studied in depth in recent literature. Kandel and Castelli [
62] study the effect of varying batch size and learning rate on VGG-16, and also provide a literature review which details several papers concerning the properties of training batch sizes. From these sources, it is clear that batch size and learning rates are dependent, but the measure of dependence often differs depending on the given task, model, and optimizer. Thus, we run a quick experimental analysis of the effect of batch size for a given learning rate on VGG-19 and the Food-11 dataset in
Table 4. The learning rate for NT is set to be 0.01 and it is set to 0.0001 for TFT, since these learning rates performed well in preliminary results. As evident from the results in
Table 4, we see that batch size does not effect the validation accuracy of NT or TFT models significantly. Therefore, we can fix batch size to 64 in the remainder of our experiments with varying learning rates.
To perform gradient descent we use Cross Entropy Loss and the Adam optimizer. After tuning, we iterate through the entire predefined validation set to find the mean loss and accuracy for a specific model (NT or TFT) and learning rate.
Our results show that algorithm performance between traditional fine-tuning (TFT) and neuromodulatory tuning (NT) is largely on par, a result that remains consistent across a wide variety of learning rates.
Table 3 provide our experimental data that highlights the best-performing learning rates for NT (lr = 0.01) and TFT (lr = 0.0001). Interestingly, the optimal learning rate for each tuning algorithm differs, and the average performance of NT across multiple learning rates is higher than that of TFT. TFT achieves the highest validation accuracies overall, but critically, not by much. This is important because it means we can retain much of TFT’s learning accuracy while using four orders of magnitude fewer trainable parameters, a circumstance that makes NT far more feasible than TFT to implement on neuromorphic hardware.
Recognizing our success in the results presented above, we further reduced the number of tunable parameters. The reduction in parameters was biologically motivated such that each tunable parameter matches to a single neuron in the classifier layers of VGG-19. Specifically, our initial results as reported in
Table 3 include a set of tunable parameters applied after the VGG-19 convolutional layers but before the data was passed into the VGG-19 classifier.
Table 5 shows the same experiment repeated with this additional layer of parameters removed, resulting in an even smaller number of trainable parameters—a critical factor for potential implementation of such methods within the space constraints of physical analog chips. We found that this reduction in parameters did decrease the accuracy of the network on each task, but only slightly. As this reduced parameter count is more analogous to biological neuromodulatory transmitters, we use this NT configuration in future experiments in
Section 5.1.
6. Results
Our long-term objective is to enable low-power analog learning behaviors in situ on physical analog chips. This requires both a viable mechanism for potential in situ learning that does not require large amounts of surface area for gradient calculations and a validated circuit design that can realistically implement that mechanism. We present neuromodulatory tuning as a possible mechanism for this objective, and here provide results showing its performance in simulated (digital) spiking neural networks (
Section 6.1) and a full chip design for its eventual implementation on physical CMOS hardware (
Section 6.2).
6.1. Neuromodulatory Tuning on Spiking Neural Networks
To validate the performance of neuromodulatory tuning in spiking neural networks (distinct from the traditional feed-foward networks shown in
Section 4), we apply neuromodulatory tuning (NT) and traditional fine-tuning (TFT) to the SNN-VGG classification layers using the STL-10, Food-11, and BCCD datasets for comparison. We fix the batch size at 64 for all training, since our experiment with batch sizes (shown in
Table 6) reveals that batch size does not impact the model performance dramatically. Both the Food-11 and BCCD datasets are singularly distinct from the ImageNet data [
56] which was used to train VGG-19. VGG-19 therefore lacks output classes corresponding to labels from the Food-11 and BCCD datasets. To create the necessary output layer size, we added one extra fully connected layer at the end of each model. This extra layer functions as the output layer for corresponding classes in Food-11 and BCCD. Different from Food-11 and BCCD, STL-10 is a subset of ImageNet. Since VGG-19 is trained on ImageNet, VGG-19 contains classes that are contained within in STL-10 labels. Therefore, we do not add extra layers for the SNN STL-10 experiments. All SNN models were trained on an AMD Ryzen Threadripper 1920X 12-Core Processor. Results are shown in
Table 7 and
Table 8.
As expected, performance is poor when no tuning is applied. This is partially because SNN architectures, comprised of leaky integrate-and-fire neurons, differ drastically from traditional deep networks in both signal accumulation and signal propagation, resulting in almost 0% accuracy on all three transfer tasks. Tuning improves this accuracy, achieving up to 88% accuracy with TFT and 50% with NT on some tasks with certain learning rates. According to our results shown in
Table 7, NT underperforms on the STL-10 dataset comparing to TFT, has equal performance to TFT on BCCD, and outperforms TFT on Food-11, which suggests that neuromodulatory tuning can positively impact learning behaviors on brain-like architectures.
Our performance comparison of the algorithms is influenced by differences between the three datasets. STL-10 is the subset of the dataset used to train VGG-19, so tasks in STL-10 is more native to the network. In contrast, Food11 and BCCD are foreign to the VGG-19 network, so those tasks will require VGG-19 to make adjustments in larger magnitudes or completely re-learn the task. Given that neuromodulatory tuning outperforms TFT on Food11, a foreign dataset, and that TFT requires changes of larger magnitudes, NT is superior for these cases. There are accuracies below random guessing, this might be caused by the low learning rate for NT and the absence of feed-forward to spiking network conversion algorithm for TFT.
Comparing two different types of NT, performs better than on STL-10 dataset, and has equal performance with on Food-11 and BCCD dataset.
According to
Table 8, TFT requires over 120 million parameters adjustment to achieve such performance, so the adjustments are impossible to implement on the physical chips. In contrast, NT method only requires 9000–20,000 adjustments, which is implementable on physical chips. Note, the parameter values for NT differ slightly in
Table 8 from
Table 5 due to the difference in implementing a spiking network versus a feed-forward network.
6.2. Analog Neuromorphic Hardware Simulation
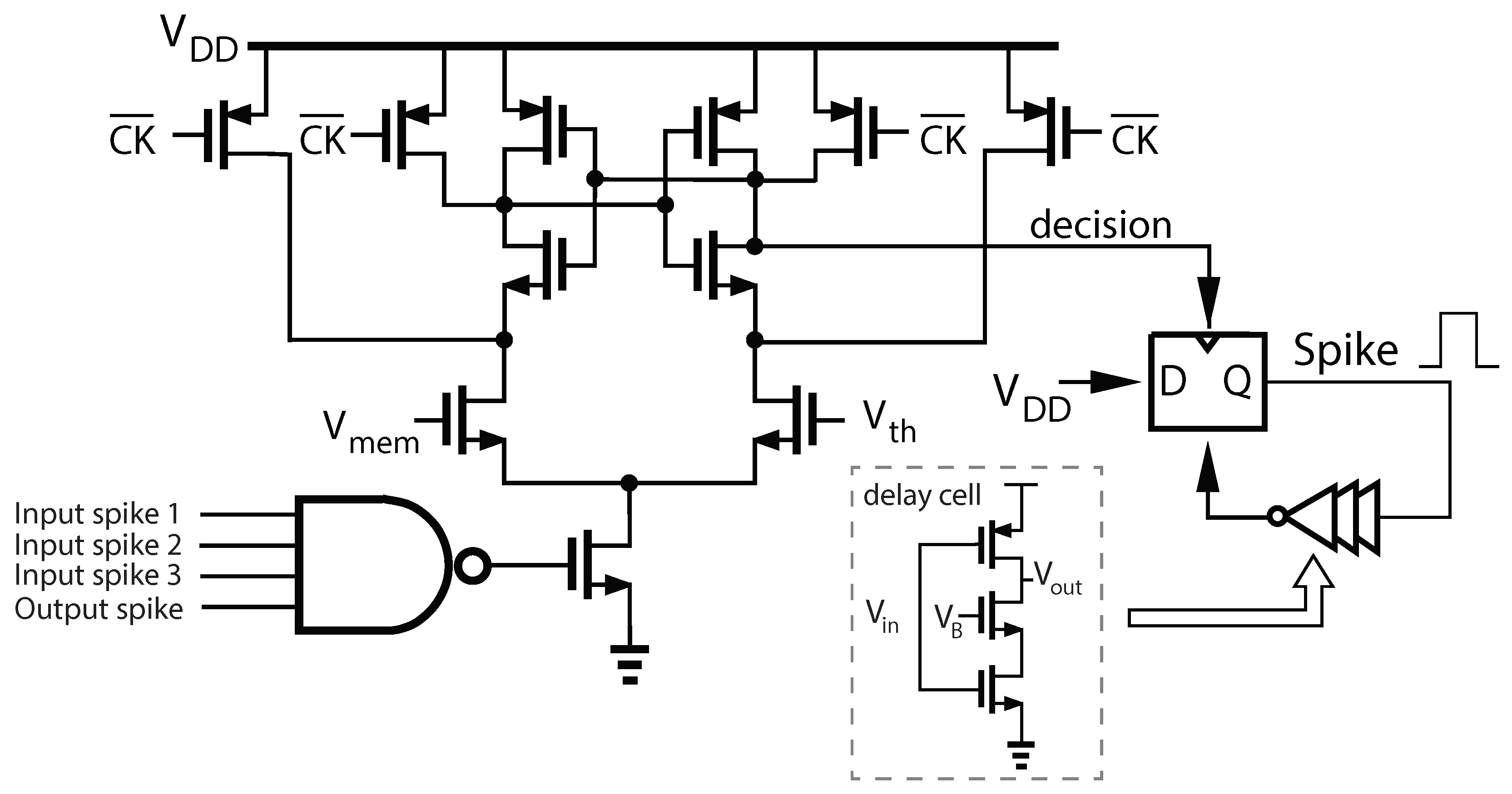
The goal of this work is to develop a low-power CMOS chip architecture that implements neuromodulatory tuning. In addition to presenting the neuromodulatory tuning algorithm and exploring its performance, we also present a complete neuron design to implements this algorithm on analog CMOS hardware.
Figure 7 shows the layout of the proposed neuron implementing NT fine tuning. The entire neuron, synapse and weight storage occupies only 598 μm
, with the neuron core (including membrane capacitor) occupying only 132 nm
. We have validated the simulation results from
Section 6.1 using post-layout simulations in Cadence Virtuoso to model an XOR task using spiking neurons. Two neurons were chosen to be the inputs to the XOR “gate” and another designated as the output. A train of 10 spikes to an input neuron constituted a “1”. No input spikes constituted a “0”. The spikes propagated through the network according to the trained weights. The output was “0” if less than three spikes were observed at the output, otherwise the output was a “1”. The analog simulation showed 2 spikes at the output for a 0, and 4 for a 1.
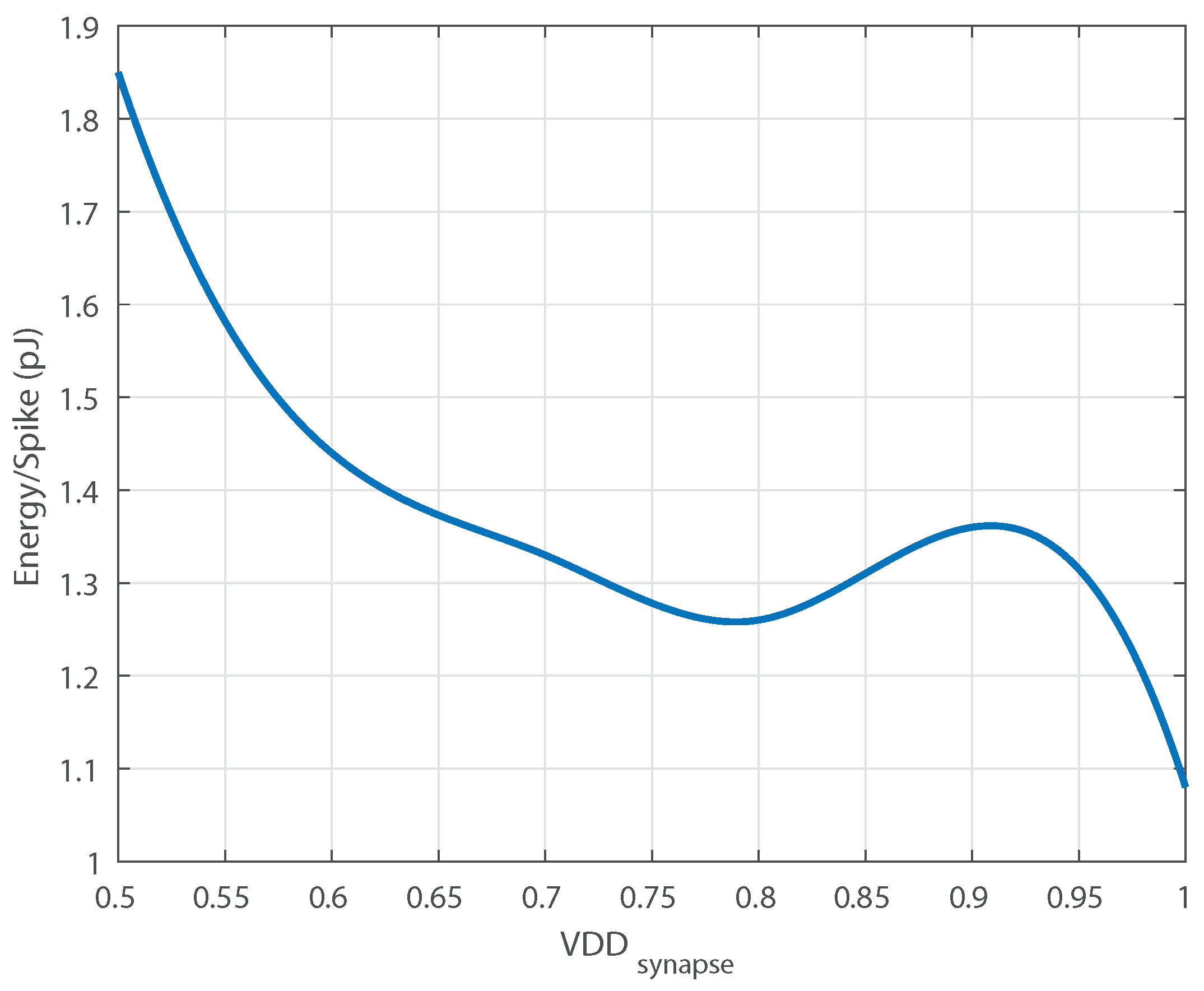
The proposed neuron achieves performance competitive with the state-of-the-art in standalone neuron circuits (see
Table 9). The total power for the neuron core varies with spike rate.
Figure 7 shows the distribution of power for two spike rates and
Figure 8 shows the energy/spike vs. spike rate.
7. Conclusions
Low-power analog machine learning has the potential to revolutionize multiple disciplines, but only if novel and physically-implementable learning algorithms are developed that enable in situ behavior modification on physical analog hardware. This paper presents a novel task transfer algorithm, termed neuromodulatory tuning, for machine learning based on biologically-inspired principles. On image recognition tasks, neuromodulatory tuning performs on test cases as well as traditional fine-tuning methods while requiring four orders of magnitude fewer active training parameters (although the total number of weights is comparable between methods). We verify this result using both deep forward networks and spiking neural network architectures. We also present a circuit design for a neuron that immplements neuromodulatory tuning, a potential layout for the use of such neurons on an analog chip, and a post-layout verification of its capabilities.
Neuromodulatory tuning has the advantage of being well-suited for implementation on neuromorphic hardware, enabling circuit implementations that support life-long learning for applications that require energy-efficient adaptation to constantly changing conditions, such as robotics, unmanned air vehicle guidance, and prosthetic limb controllers. Future research in this area should focus on probing the performance of NT in domains beyond image recognition; exploring the possibility of paired bias links in which multiple neurons connect to a single power domain region; and designing improved SNN update algorithms with stronger convergence properties.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}