
Review of Allelopathy in Green Tides: The Case of Ulva prolifera in the South Yellow Sea

 , ,
, ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
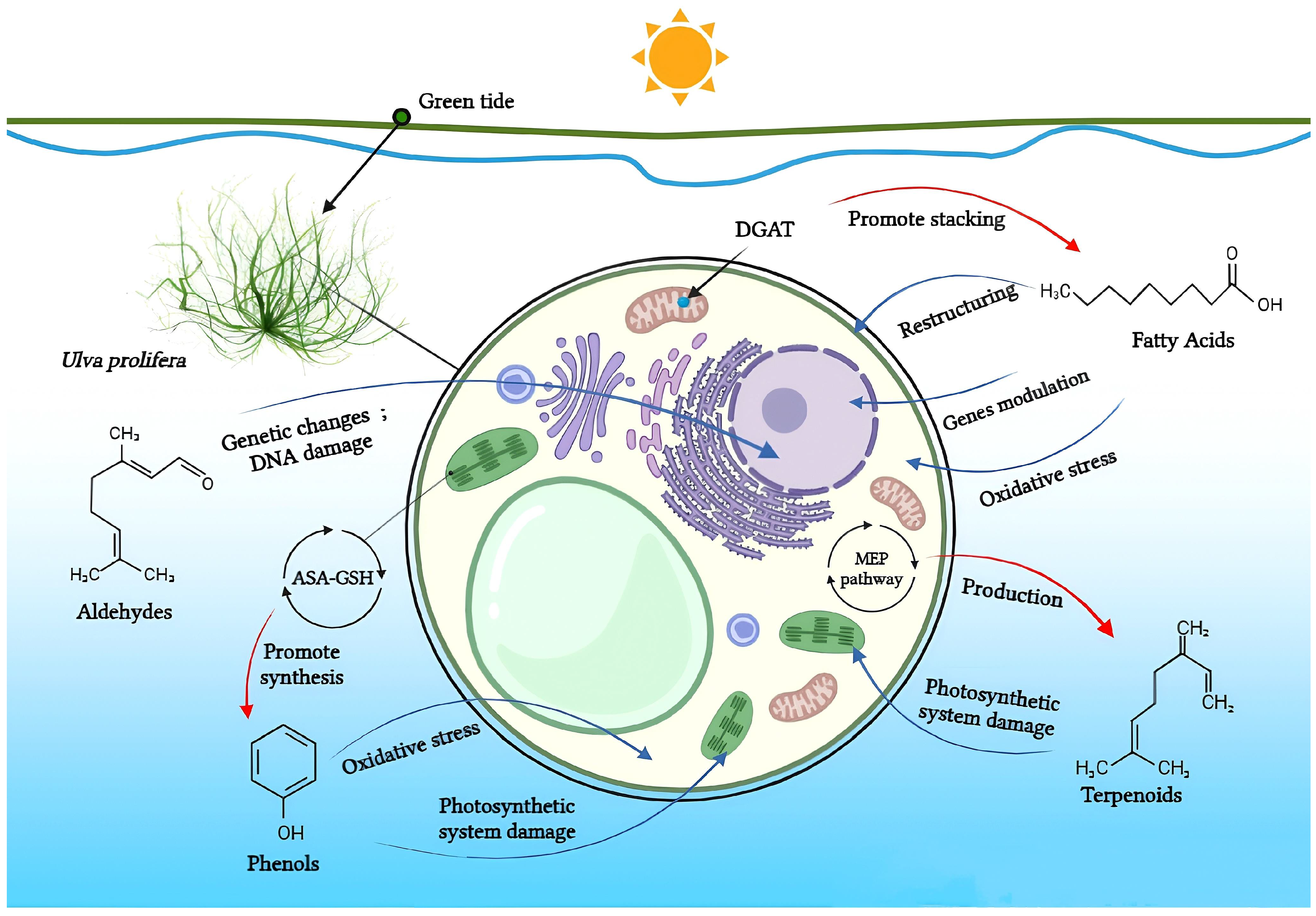
2. Allelochemicals and Allelopathy Mechanisms in the Green Tide Process
2.1. Fatty Acids
2.2. Aldehydes
2.3. Phenols
2.4. Terpenoids
2.5. Other Allelochemicals
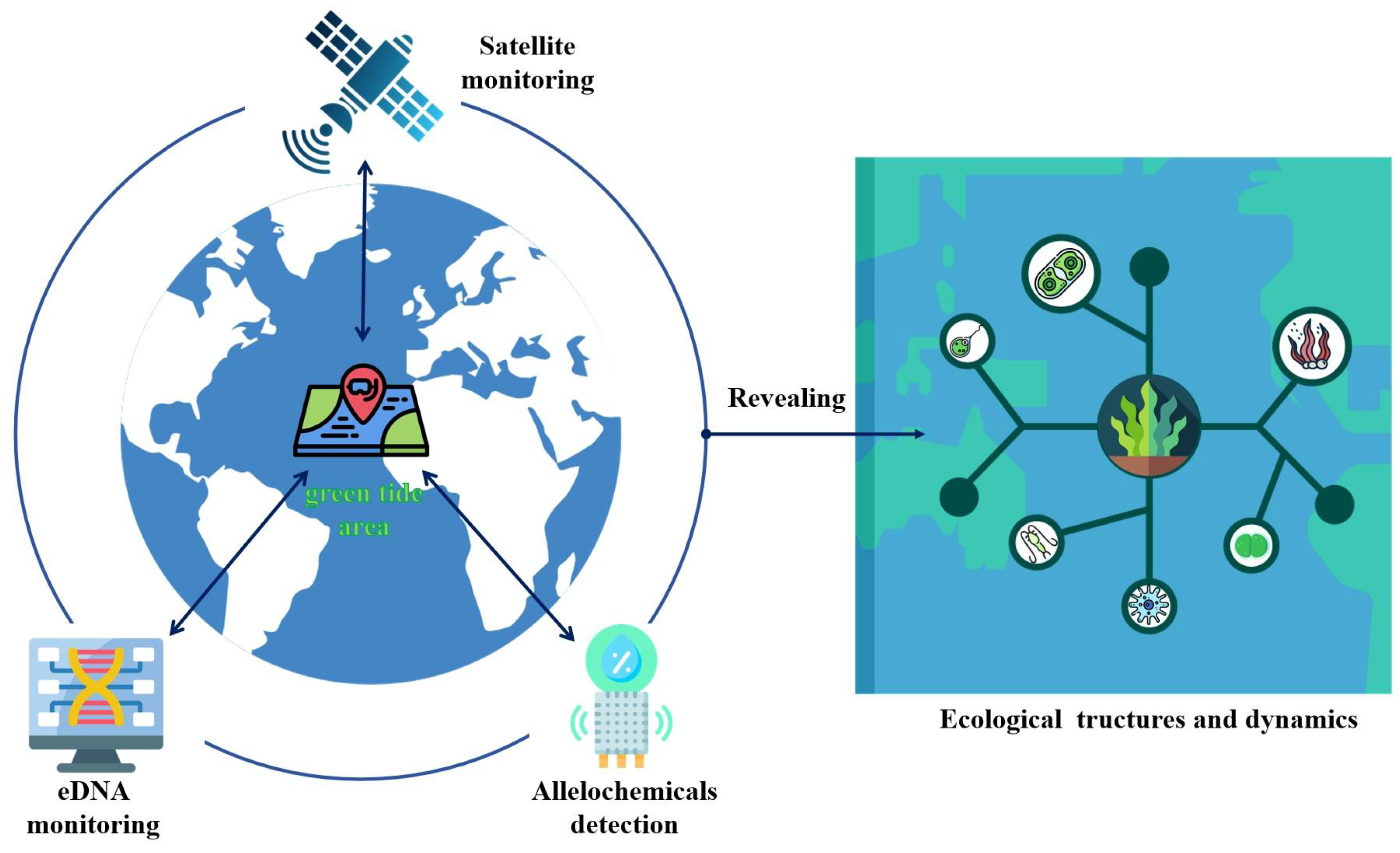
3. Rapid Detection and Application Prospect of Allelochemicals
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smetacek, V.; Zingone, A. Green and golden seaweed tides on the rise. Nature 2013, 504, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, P.; Huo, Y.; Yu, K.; He, P. The fast expansion of Pyropia aquaculture in “Sansha” regions should be mainly responsible for the Ulva blooms in Yellow Sea. Estuar. Coast. Shelf Sci. 2017, 189, 58–65. [Google Scholar] [CrossRef]
- He, P.; Duan, Y.; Qiao, L. Strategy of macroalgae eco-remediation with cases in nearshore China. J. Appl. Oceanogr. 2021, 40, 557–563. [Google Scholar]
- Li, H.; Feng, X.; Xiong, T.; He, C.; Wu, W.; Shi, Q.; Jiao, N.; Zhang, Y. Green tides significantly alter the molecular composition and properties of coastal DOC and perform dissolved carbon sequestration. Environ. Sci. Technol. 2022, 57, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-Y.; Geng, H.-X.; Zhang, Q.-C.; Li, Y.-F.; Kong, F.-Z.; Yan, T.; Zhou, M.-J.; Yang, D.; Yuan, Y.; Yu, R.-C.J.E.S.; et al. Green tides in the Yellow Sea promoted the proliferation of pelagophyte Aureococcus anophagefferens. Environ. Sci. Technol. 2022, 56, 3056–3064. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, R. The occurrence of “green tides”—A review. In Marine Benthic Vegetation: Recent Changes the Effects of Eutrophication; Springer: Berlin/Heidelberg, Germany, 1996; pp. 7–43. [Google Scholar]
- Morand, P.; Merceron, M. Macroalgal population and sustainability. J. Coast. Res. 2005, 21, 1009–1020. [Google Scholar] [CrossRef]
- Hiraoka, M.; Dan, A.; Shimada, S.; Hagihira, M.; Migita, M.; Ohno, M. Different life histories of Enteromorpha prolifera (Ulvales, Chlorophyta) from four rivers on Shikoku Island, Japan. Phycologia 2003, 42, 275–284. [Google Scholar] [CrossRef]
- Liu, D.; Keesing, J.K.; Xing, Q.; Shi, P. World’s largest macroalgal bloom caused by expansion of seaweed aquaculture in China. Mar. Pollut. Bull. 2009, 58, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huo, Y.; Yu, K.; Chen, Q.; He, Q.; Han, W.; Chen, L.; Cao, J.; Shi, D.; He, P. Growth characteristics and reproductive capability of green tide algae in Rudong coast, China. J. Appl. Phycol. 2013, 25, 795–803. [Google Scholar] [CrossRef]
- Gao, G.; Clare, A.S.; Rose, C.; Caldwell, G.S. Eutrophication and warming-driven green tides (Ulva rigida) are predicted to increase under future climate change scenarios. Mar. Pollut. Bull. 2017, 114, 439–447. [Google Scholar] [CrossRef]
- Li, Y.; Song, W.; Xiao, J.; Wang, Z.; Fu, M.; Zhu, M.; Li, R.; Zhang, X.; Wang, X. Tempo-spatial distribution and species diversity of green algae micro-propagules in the Yellow Sea during the large-scale green tide development. Harmful Algae 2014, 39, 40–47. [Google Scholar] [CrossRef]
- Zongling, W.; Mingzhu, F.; Jie, X.; Xuelei, Z.; Wei, S. Progress on the study of the Yellow Sea green tides caused by Ulva prolifera. Acta Oceanol. Sin. 2018, 40, 1–13. [Google Scholar]
- Wang, Z.-Y.; Tian, Z.-J.; Li, F.-M.; An, Z.; Hu, H.-Y. Allelopathic effects of large seaweeds on red tide dinoflagellate Gymnodinium breve. Allelopath. J. 2008, 22, 181–188. [Google Scholar]
- Xu, D.; Gao, Z.; Zhang, X.; Fan, X.; Wang, Y.; Li, D.; Wang, W.; Zhuang, Z.; Ye, N. Allelopathic interactions between the opportunistic species Ulva prolifera and the native macroalga Gracilaria lichvoides. PLoS ONE 2012, 7, e33648. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Cao, S.; Li, J.; Zhao, S.; Liu, J.; Zhuang, M.; Qin, Y.; Gao, S.; Sun, Y.; Kim, J.K. Controlling the main source of green tides in the Yellow Sea through the method of biological competition. Mar. Pollut. Bull. 2022, 177, 113561. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Liu, J.; Zhao, S.; Sun, Y.; Cui, Q.; Wu, L.; Gao, S.; Zhang, J.; He, P. Review of the development of the green tide and the process of control in the southern Yellow Sea in 2022. Estuar. Coast. Shelf Sci. 2024, 302, 108772. [Google Scholar] [CrossRef]
- Diller, J.G.P.; Hüftlein, F.; Lücker, D.; Feldhaar, H.; Laforsch, C. Allelochemical run-off from the invasive terrestrial plant Impatiens glandulifera decreases defensibility in Daphnia. Sci. Rep. 2023, 13, 1207. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xie, M.; Li, X.; Jin, H.; Yang, X.; Yan, Z.; Su, A.; Qin, B. Main allelochemicals from the rhizosphere soil of Saussurea lappa (Decne.) Sch. Bip. and their effects on plants’ antioxidase systems. Molecules 2018, 23, 2506. [Google Scholar] [CrossRef] [PubMed]
- Quan, W.; Wang, A.; Li, C.; Xie, L. Allelopathic potential and allelochemical composition in different soil layers of Rhododendron delavayi forest, southwest China. Front. Ecol. Evol. 2022, 10, 963116. [Google Scholar] [CrossRef]
- Kong, C.-H.; Xuan, T.D.; Khanh, T.D.; Tran, H.-D.; Trung, N.T. Allelochemicals and signaling chemicals in plants. Molecules 2019, 24, 2737. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathy and allelochemicals of Imperata cylindrica as an invasive plant species. Plants 2022, 11, 2551. [Google Scholar] [CrossRef] [PubMed]
- Hickman, D.T.; Rasmussen, A.; Ritz, K.; Birkett, M.A.; Neve, P. Allelochemicals as multi-kingdom plant defence compounds: Towards an integrated approach. Pest Manag. Sci. 2021, 77, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Rahaman, F.; Shukor Juraimi, A.; Rafii, M.Y.; Uddin, K.; Hassan, L.; Chowdhury, A.K.; Karim, S.M.R.; Yusuf Rini, B.; Yusuff, O.; Bashar, H.K. Allelopathic potential in rice-a biochemical tool for plant defence against weeds. Front. Plant Sci. 2022, 13, 1072723. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Mejías, F.J.; Molinillo, J.M. Recent advances in allelopathy for weed control: From knowledge to applications. Pest Manag. Sci. 2019, 75, 2413–2436. [Google Scholar] [CrossRef] [PubMed]
- Kalisz, S.; Kivlin, S.N.; Bialic-Murphy, L. Allelopathy is pervasive in invasive plants. Biol. Invasions 2021, 23, 367–371. [Google Scholar] [CrossRef]
- Zeng, R.S.; Wen, Z.; Niu, G.; Schuler, M.A.; Berenbaum, M.R. Allelochemical induction of cytochrome P450 monooxygenases and amelioration of xenobiotic toxicity in Helicoverpa zea. J. Chem. Ecol. 2007, 33, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Teneva, I.; Velikova, V.; Belkinova, D.; Moten, D.; Dzhambazov, B. Allelopathic Potential of the Cyanotoxins Microcystin-LR and Cylindrospermopsin on Green Algae. Plants 2023, 12, 1403. [Google Scholar] [CrossRef] [PubMed]
- Chia, M.A.; Bittencourt-Oliveira, M.d.C. Allelopathic interactions between phytoplankton species alter toxin production, oxidative response, and nitrogen fixation. Hydrobiologia 2021, 848, 4623–4635. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, Y.; Yuan, L.; Weber, E.; van Kleunen, M. Effect of allelopathy on plant performance: A meta-analysis. Ecol. Lett. 2021, 24, 348–362. [Google Scholar] [CrossRef]
- Singh, A.; Thakur, N.L. Allelopathic interaction among rocky intertidal invertebrates: Sponge Cinachyrella cf. cavernosa and Zooxanthellate zoanthids Zoanthus sansibaricus. Hydrobiologia 2021, 848, 4647–4659. [Google Scholar]
- Allen, J.L.; Ten-Hage, L.; Leflaive, J. Allelopathic interactions involving benthic phototrophic microorganisms. Environ. Microbiol. Rep. 2016, 8, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, Y.; Jin, Y.; Cui, T.; Wang, S.; Kong, F. The inhibitory effects of Ulva prolifera extracts on early growth of Spartina alterniflora and the underlying mechanisms. J. Environ. Manag. 2022, 319, 115639. [Google Scholar] [CrossRef] [PubMed]
- Gostyńska, J.; Pankiewicz, R.; Romanowska-Duda, Z.; Messyasz, B. Overview of allelopathic potential of Lemna minor L. obtained from a shallow eutrophic lake. Molecules 2022, 27, 3428. [Google Scholar] [CrossRef] [PubMed]
- Felpeto, A.B.; Roy, S.; Vasconcelos, V.M. Allelopathy prevents competitive exclusion and promotes phytoplankton biodiversity. Oikos 2018, 127, 85–98. [Google Scholar] [CrossRef]
- Gross, E.M. Allelopathy of aquatic autotrophs. Crit. Rev. Plant Sci. 2003, 22, 313–339. [Google Scholar] [CrossRef]
- Semmouri, I.; Janssen, C.R.; Asselman, J. Allelopathy in macroalgae: Ecological principles, research opportunities and pitfalls reviewed. J. Appl. Phycol. 2024, 36, 441–458. [Google Scholar] [CrossRef]
- Cai, J.; Ni, J.; Chen, Z.; Wu, S.; Wu, R.; He, C.; Wang, J.; Liu, Y.; Zhou, W.; Xu, J. Effects of ocean acidification and eutrophication on the growth and photosynthetic performances of a green tide alga Ulva prolifera. Front. Mar. Sci. 2023, 10, 1145048. [Google Scholar] [CrossRef]
- Lv, M.; Yuan, M.; Wang, Y.; Tang, X.; Zhao, Y. Allelopathic effects of Ulva linza on marine phytoplankton and identification of the allelochemicals. Environ. Sci. Pollut. Res. 2021, 28, 45714–45723. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhang, J.; Hu, C. Study of allelopathic effect on spore attachment of Ulva prolifera. Environ. Sci. Technol. 2021, 44, 26–34. [Google Scholar]
- Żak, A.; Kosakowska, A. Allelopathic influence of cyanobacteria Microcystis aeruginosa on green algae Chlorella vulgaris. In Insights on Environmental Changes: Where the World is Heading; Springer: Cham, Switzerland, 2014; pp. 141–150. [Google Scholar]
- Zhang, X.; Chi, X.; Wang, Y.; Zhang, J.; Zhang, Y.; Xu, D.; Fan, X.; Liang, C.; Ye, N. Characterization of a broad substrates specificity acyl-CoA: Diacylglycerol acyltransferase 1 from the green tide alga Ulva prolifera. Acta Oceanol. Sin. 2020, 39, 42–49. [Google Scholar] [CrossRef]
- Matorin, D.; Plekhanov, S.; Bratkovskaya, L.; Yakovleva, O.; Alekseev, A. The effect of phenols on the parameters of chlorophyll fluorescence and reactions of P 700 in green algae Scenedesmus quadricauda. Biophysics 2014, 59, 374–379. [Google Scholar] [CrossRef]
- Zaytseva, T.; Medvedeva, N.; Mamontova, V. Peculiarities of the effect of octyl-and nonylphenols on the growth and development of microalgae. Inland Water Biol. 2015, 8, 406–413. [Google Scholar] [CrossRef]
- Xie, Y.; Tian, L.; Han, X.; Yang, Y. Research advances in allelopathy of volatile organic compounds (VOCs) of plants. Horticulturae 2021, 7, 278. [Google Scholar] [CrossRef]
- Gu, K.; Liu, Y.; Jiang, T.; Cai, C.; Zhao, H.; Liu, X.; He, P. Molecular response of Ulva prolifera to short-term high light stress revealed by a multi-omics approach. Biology 2022, 11, 1563. [Google Scholar] [CrossRef]
- Sun, Y.-y.; Wang, H.; Guo, G.-l.; Pu, Y.-f.; Yan, B.-l.; Wang, C.-h. Isolation, purification, and identification of antialgal substances in green alga Ulva prolifera for antialgal activity against the common harmful red tide microalgae. Environ. Sci. Pollut. Res. 2016, 23, 1449–1459. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Tong, M.; Glibert, P.M. Effect of allelochemicals on photosynthetic and antioxidant defense system of Ulva prolifera. Aquat. Toxicol. 2020, 224, 105513. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhang, J.; Zhao, X.; Wang, P.; Tong, M.; Glibert, P.M. Allelopathic Inhibition by the Bacteria Bacillus cereus BE23 on Growth and Photosynthesis of the Macroalga Ulva prolifera. J. Mar. Sci. Eng. 2020, 8, 718. [Google Scholar] [CrossRef]
- Mushtaq, W.; Siddiqui, M.B.; Hakeem, K.R.; Mushtaq, W.; Siddiqui, M.B.; Hakeem, K.R. Mechanism of Action of Allelochemicals. In Allelopathy: Potential for Green Agriculture; Springer: Cham, Switzerland, 2020; pp. 61–66. [Google Scholar]
- Salek, M.; Hosseini Hooshiar, S.; Salek, M.; Poorebrahimi, M.; Jafarnejad, S. Omega-3 fatty acids: Current insights into mechanisms of action in systemic lupus erythematosus. Lupus 2023, 32, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Liu, Q. Responses of bloom-forming Heterosigma akashiwo to allelochemical linoleic acid: Growth inhibition, oxidative stress and apoptosis. Front. Mar. Sci. 2022, 8, 793567. [Google Scholar] [CrossRef]
- Pinto, M.E.A.; Silva, N.L.; de Siqueira, E.P.; dos Santos, F.J.L.; dos Santos Lima, L.A.R. Efeito alelopático dos ésteres metílicos dos ácidos graxos obtidos de óleos de milho e girassol. BBR—Biochem. Biotechnol. Rep. 2013, 2, 44–48. [Google Scholar] [CrossRef]
- González-Becerra, K.; Ramos-López, O.; Barrón-Cabrera, E.; Riezu-Boj, J.I.; Milagro, F.; Martínez-López, E.; Martínez, J. Fatty acids, epigenetic mechanisms and chronic diseases: A systematic review. Lipids Health Dis. 2019, 18, 178. [Google Scholar] [CrossRef]
- Gniazdowska, A.; Bogatek, R.J. Allelopathic interactions between plants. Multi site action of allelochemicals. Acta Physiol. Plant. 2005, 27, 395–407. [Google Scholar] [CrossRef]
- Gniazdowska, A.; Oracz, K.; Bogatek, R. Allelopatia-nowe interpretacje oddziaływań pomiędzy roślinami. Kosmos 2004, 53, 207–217. [Google Scholar]
- Ali, H.A.; Al-Darraji, M.N.; Ali, S.F. The Allelopathic in the Diatoms. Rafidain J. Sci. 2021, 30, 30–38. [Google Scholar] [CrossRef]
- Liu, Q.; Cui, R.; Lin, J.; Kang, Z.; Zhou, X. Competition relations between selected microalgae and bloom-forming Ulva prolifera. J. Mar. Biol. Assoc. 2023, 103, e43. [Google Scholar] [CrossRef]
- Zhuo, J.; Wang, H.; Du, Y.; Shi, M.; Huan, L.; Wang, G. Transcriptomic Analysis of Ulva prolifera in Response to Salt Stress. Water 2022, 15, 63. [Google Scholar] [CrossRef]
- Xie, M.-Z.; Shoulkamy, M.I.; Salem, A.M.; Oba, S.; Goda, M.; Nakano, T.; Ide, H. Aldehydes with high and low toxicities inactivate cells by damaging distinct cellular targets. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2016, 786, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Das, C.; Dey, A.; Bandyopadhyay, A. Allelochemicals: An emerging tool for weed management. In Evidence Based Validation of Traditional Medicines: A Comprehensive Approach; Springer: Singapore, 2021; pp. 249–259. [Google Scholar]
- Kaiira, M.; Chemining’wa, G.; Ayuke, F.; Baguma, Y.; Atwijukire, E. Allelopathic potential of compounds in selected crops. J. Agric. Sci. 2021, 13, 192. [Google Scholar] [CrossRef]
- Shirgapure, K.; Ghosh, P. Allelopathy a tool for sustainable weed management. Arch. Curr. Res. Int. 2020, 20, 17–25. [Google Scholar] [CrossRef]
- Chou, C.-H. Allelopathic compounds as naturally occurring herbicides. In Proceedings of the 15th Asian-Pacific Weeds Control Conference, Tsukuba, Japan, 24–28 July 1995; pp. 107–115. [Google Scholar]
- LoPachin, R.M.; Gavin, T. Molecular mechanisms of aldehyde toxicity: A chemical perspective. Chem. Res. Toxicol. 2014, 27, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Ambika, S. Multifaceted attributes of allelochemicals and mechanism of allelopathy. In Allelopathy: Current Trends and Future Applications; Springer: Berlin/Heidelberg, Germany, 2012; pp. 389–405. [Google Scholar]
- Scavo, A.; Restuccia, A.; Mauromicale, G. Allelopathy: Principles and basic aspects for agroecosystem control. In Sustainable Agriculture Reviews 28: Ecology for Agriculture; Springer: Cham, Switzerland, 2018; pp. 47–101. [Google Scholar]
- Van Alstyne, K.L.; Nelson, T.A.; Ridgway, R.L. Environmental chemistry and chemical ecology of “green tide” seaweed blooms. Integr. Comp. Biol. 2015, 55, 518–532. [Google Scholar] [CrossRef] [PubMed]
- Brannon, M.A.; Bartsch, A.F. Influence of growth substances on growth and cell division in green algae. Am. J. Bot. 1939, 26, 271–279. [Google Scholar] [CrossRef]
- Jalliffier-Merlon, E.; Marty, J.-C.; Denant, V.; Saliot, A. Phytoplanktonic sources of volatile aldehydes in the river Rhône estuary. Estuar. Coast. Shelf Sci. 1991, 32, 463–482. [Google Scholar] [CrossRef]
- Prince, E.K.; Myers, T.L.; Kubanek, J. Effects of harmful algal blooms on competitors: Allelopathic mechanisms of the red tide dinoflagellate Karenia brevis. Limnol. Oceanogr. 2008, 53, 531–541. [Google Scholar] [CrossRef]
- Hajnal-Jafari, T.I.; Đurić, S.S.; Stamenov, D.R. Influence of green algae Chlorella vulgaris on initial growth of different agricultural crops. Zb. Matice Srp. Prir. Nauk. 2016, 130, 29–33. [Google Scholar] [CrossRef]
- Misra, D.; Dutta, W.; Jha, G.; Ray, P. Interactions and regulatory functions of phenolics in soil-plant-climate nexus. Agronomy 2023, 13, 280. [Google Scholar] [CrossRef]
- He, Z.; Wang, Y.; Yan, Y.; Qin, S.; He, H.; Mao, R.; Liang, Z. Dynamic analysis of physiological indices and transcriptome profiling revealing the mechanisms of the allelopathic effects of phenolic acids on Pinellia ternata. Front. Plant Sci. 2022, 13, 1039507. [Google Scholar] [CrossRef]
- Pardo-Muras, M.; Puig, C.G.; Pedrol, N. Complex synergistic interactions among volatile and phenolic compounds underlie the effectiveness of allelopathic residues added to the soil for weed control. Plants 2022, 11, 1114. [Google Scholar] [CrossRef]
- Bashar, H.K.; Juraimi, A.S.; Ahmad-Hamdani, M.S.; Uddin, M.K.; Asib, N.; Anwar, M.P.; Rahaman, F.; Haque, M.A.; Hossain, A. Evaluation of allelopathic effects of Parthenium hysterophorus L. methanolic extracts on some selected plants and weeds. PLoS ONE 2023, 18, e0280159. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, J.; Zhu, Y.; Zhao, T.; Guo, L.; Kang, L.; Yu, J.; Du, H.; Liu, D. Weed suppression and molecular mechanisms of isochlorogenic acid A isolated from Artemisia argyi extract via an activity-guided method. J. Agric. Food Chem. 2022, 70, 1494–1506. [Google Scholar] [CrossRef]
- Zhong, Y.; Xu, J.; Zhao, X.; Qu, T.; Guan, C.; Hou, C.; Tang, X.; Wang, Y. Balancing Damage via Non-Photochemical Quenching, Phenolic Compounds and Photorespiration in Ulva prolifera Induced by Low-Dose and Short-Term UV-B Radiation. Int. J. Mol. Sci. 2022, 23, 2693. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Lin, J.; Jiang, J.; Hu, S.; Kang, C.K.; Xu, N.; Li, Y. Environmental history affects the growth and photosynthesis of a green-tide macroalgae Ulva prolifera. Aquac. Res. 2022, 53, 2509–2517. [Google Scholar] [CrossRef]
- He, Y.; Shen, S.; Yu, D.; Wang, Y.; Yin, J.; Wang, Z.; Ye, Y. The Ulva prolifera genome reveals the mechanism of green tides. J. Oceanol. Limnol. 2021, 39, 1458–1470. [Google Scholar] [CrossRef]
- Del Mondo, A.; Smerilli, A.; Ambrosino, L.; Albini, A.; Noonan, D.M.; Sansone, C.; Brunet, C. Insights into phenolic compounds from microalgae: Structural variety and complex beneficial activities from health to nutraceutics. Crit. Rev. Biotechnol. 2021, 41, 155–171. [Google Scholar] [CrossRef]
- Pei, Y.; Liu, L.; Hilt, S.; Xu, R.; Wang, B.; Li, C.; Chang, X. Root exudated algicide of Eichhornia crassipes enhances allelopathic effects of cyanobacteria Microcystis aeruginosa on green algae. Hydrobiologia 2018, 823, 67–77. [Google Scholar] [CrossRef]
- Huang, S.; Zhu, J.; Zhang, L.; Peng, X.; Zhang, X.; Ge, F.; Liu, B.; Wu, Z. Combined effects of allelopathic polyphenols on Microcystis aeruginosa and response of different chlorophyll fluorescence parameters. Front. Microbiol. 2020, 11, 614570. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, X.; Ma, Z.; Luan, Z.; Wang, Y.; Wang, Z.; Wang, L. Removal of phenolic substances from wastewater by algae. A review. Environ. Chem. Lett. 2020, 18, 377–392. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, X.; Ding, L.; Kong, C.-H. Allelopathy and allelochemicals in grasslands and forests. Forests 2023, 14, 562. [Google Scholar] [CrossRef]
- Ooka, J.K.; Owens, D.K. Allelopathy in tropical and subtropical species. Phytochem. Rev. 2018, 17, 1225–1237. [Google Scholar] [CrossRef]
- Muzell Trezzi, M.; Vidal, R.A.; Balbinot Junior, A.A.; von Hertwig Bittencourt, H.; da Silva Souza Filho, A.P. Allelopathy: Driving mechanisms governing its activity in agriculture. J. Plant Interact. 2016, 11, 53–60. [Google Scholar] [CrossRef]
- Cheng, F.; Cheng, Z. Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front. Plant Sci. 2015, 6, 160714. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, F.; Wang, M.; Moejes, F.W.; Bi, Y. Characterization and transcriptional analysis of one carbonic anhydrase gene in the green-tide-forming alga Ulva prolifera (Ulvophyceae, Chlorophyta). Phycol. Res. 2020, 68, 90–97. [Google Scholar] [CrossRef]
- He, Y.; Ye, Y.; Shen, S. Effects of light and salinity on carotenoid biosynthesis in Ulva prolifera. Acta Oceanol. Sin. 2020, 39, 50–57. [Google Scholar] [CrossRef]
- He, Y.; Ma, Y.; Du, Y.; Shen, S. Differential gene expression for carotenoid biosynthesis in a green alga Ulva prolifera based on transcriptome analysis. BMC Genom. 2018, 19, 916. [Google Scholar] [CrossRef] [PubMed]
- Nelson, T.; Lee, D.; Smith, B.; Prins, R. Are ‘Green TIDES’Harmful Algal Blooms? Allelopathic Properties of Extracts from Ulva Fenestrata and Ulvaria Obscura. J. Phycol. 2002, 38, 28–29. [Google Scholar] [CrossRef]
- Qiu, X.; Yamasaki, Y.; Shimasaki, Y.; Gunjikake, H.; Honda, M.; Kawaguchi, M.; Matsubara, T.; Nagasoe, S.; Etoh, T.; Matsui, S. Allelopathy of the raphidophyte Heterosigma akashiwo against the dinoflagellate Akashiwo sanguinea is mediated via allelochemicals and cell contact. Mar. Ecol. Prog. Ser. 2012, 446, 107–118. [Google Scholar] [CrossRef]
- Deng, R.; Wang, F.; Wang, L.; Xiong, L.; Shen, X.; Song, H. Advances in plant polysaccharides as antiaging agents: Effects and signaling mechanisms. J. Agric. Food Chem. 2023, 71, 7175–7191. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhou, Y.; Hu, C. One Step Catalytic Conversion of Polysaccharides in Ulva prolifera to Lactic Acid and Value-Added Chemicals. Catalysts 2023, 13, 262. [Google Scholar] [CrossRef]
- Ge, H.; Ni, Q.; Chen, Z.; Li, J.; Zhao, F. Effects of short period feeding polysaccharides from marine macroalga, Ulva prolifera on growth and resistance of Litopenaeus vannamei against Vibrio parahaemolyticus infection. J. Appl. Phycol. 2019, 31, 2085–2092. [Google Scholar] [CrossRef]
- Lebecque, S.; Crowet, J.-M.; du Jardin, P.; Delory, B.; Lins, L.; Deleu, M.; Fauconnier, M.-L. Interactions of allelochemicals with plant plasma membrane: A case study with alkaloids from barley. In Proceedings of the 41st New Phytologist Symposium, Nancy, France, 11–13 April 2018. [Google Scholar]
- Greer, B.; McNamee, S.E.; Boots, B.; Cimarelli, L.; Guillebault, D.; Helmi, K.; Marcheggiani, S.; Panaiotov, S.; Breitenbach, U.; Akçaalan, R. A validated UPLC–MS/MS method for the surveillance of ten aquatic biotoxins in European brackish and freshwater systems. Harmful Algae 2016, 55, 31–40. [Google Scholar] [CrossRef]
- Ma, J.; Meng, F.; Zhou, Y.; Wang, Y.; Shi, P. Distributed water pollution source localization with mobile UV-visible spectrometer probes in wireless sensor networks. Sensors 2018, 18, 606. [Google Scholar] [CrossRef]
- Hellou, J.; Ross, N.W.; Moon, T.W. Glutathione, glutathione S-transferase, and glutathione conjugates, complementary markers of oxidative stress in aquatic biota. Environ. Sci. Pollut. Res. 2012, 19, 2007–2023. [Google Scholar] [CrossRef] [PubMed]
- Denisov, I.; Lukyanenko, K.; Yakimov, A.; Kukhtevich, I.; Esimbekova, E.; Belobrov, P. Disposable luciferase-based microfluidic chip for rapid assay of water pollution. Luminescence 2018, 33, 1054–1061. [Google Scholar] [CrossRef]
- Chouler, J.; Di Lorenzo, M. Water quality monitoring in developing countries; can microbial fuel cells be the answer? Biosensors 2015, 5, 450–470. [Google Scholar] [CrossRef]
- De Fazio, R.; Dinoi, L.M.; De Vittorio, M.; Visconti, P. A sensor-based drone for pollutants detection in eco-friendly cities: Hardware design and data analysis application. Electronics 2021, 11, 52. [Google Scholar] [CrossRef]
- Jurdak, R.; Elfes, A.; Kusy, B.; Tews, A.; Hu, W.; Hernandez, E.; Kottege, N.; Sikka, P. Autonomous surveillance for biosecurity. Trends Biotechnol. 2015, 33, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Dörnhöfer, K.; Oppelt, N. Remote sensing for lake research and monitoring–Recent advances. Ecol. Indic. 2016, 64, 105–122. [Google Scholar] [CrossRef]
- Sagan, V.; Peterson, K.T.; Maimaitijiang, M.; Sidike, P.; Sloan, J.; Greeling, B.A.; Maalouf, S.; Adams, C. Monitoring inland water quality using remote sensing: Potential and limitations of spectral indices, bio-optical simulations, machine learning, and cloud computing. Earth-Sci. Rev. 2020, 205, 103187. [Google Scholar] [CrossRef]
- Geng, H.; Yu, R.; Yan, T.; Zhang, Q.; Kong, F. Using sterol biomakers to trace deposition areas of floating green algae after green tides. Oceanol. Limnol. Sin. 2018, 49, 1094–1102. [Google Scholar]
- Geng, H.; Yu, R.; Zhang, Q.; Yan, T.; Kong, F.; Zhou, M. Tracing the settlement region of massive floating green algae in the Yellow Sea. J. Oceanol. Limnol. 2019, 37, 1555–1565. [Google Scholar] [CrossRef]
- Liu, J.; Yang, X.; Li, J.; Wen, Q.; Liu, W.; Song, W.; He, P.; Zhang, J. Research progress on settled mature Ulva prolifera during the outbreak of green tide in the Yellow Sea. Environ. Pollut. Control 2020, 42, 614–618. [Google Scholar]
- Malthus, T.J.; Lehmann, E.; Ho, X.; Botha, E.; Anstee, J. Implementation of a satellite based inland water algal bloom alerting system using analysis ready data. Remote Sens. 2019, 11, 2954. [Google Scholar] [CrossRef]
- Shang, W.; Gao, Z.; Gao, M.; Jiang, X. Monitoring green tide in the Yellow Sea using high-resolution imagery and deep learning. Remote Sens. 2023, 15, 1101. [Google Scholar] [CrossRef]
- Song, Q.; Ma, C.; Liu, J.; Wei, H. Quantifying ocean surface green tides using high-spatial resolution thermal images. Opt. Express 2022, 30, 36592–36602. [Google Scholar] [CrossRef] [PubMed]
- Ortega, A.; Geraldi, N.R.; Díaz-Rúa, R.; Ørberg, S.B.; Wesselmann, M.; Krause-Jensen, D.; Duarte, C.M. A DNA mini-barcode for marine macrophytes. Mol. Ecol. Resour. 2020, 20, 920–935. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Wang, X.; Liu, J.; Cao, J.; Sun, Y.; Zhao, S.; Chen, Z.; Kim, J.K.; Zhang, J.; He, P. Harnessing the power of eDNA technology for macroalgal ecological studies: Recent advances, challenges, and future perspectives. Algal Res. 2023, 77, 103340. [Google Scholar] [CrossRef]
- Rees, H.C.; Maddison, B.C.; Middleditch, D.J.; Patmore, J.R.; Gough, K.C. The detection of aquatic animal species using environmental DNA–a review of eDNA as a survey tool in ecology. J. Appl. Ecol. 2014, 51, 1450–1459. [Google Scholar] [CrossRef]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. Genet. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Zeng, Y.; Chen, Z.; Cao, J.; Li, S.; Xia, Z.; Sun, Y.; Zhang, J.; He, P. Revolutionizing early-stage green tide monitoring: eDNA metabarcoding insights into Ulva prolifera and microecology in the South Yellow Sea. Sci. Total Environ. 2024, 912, 169022. [Google Scholar] [CrossRef]
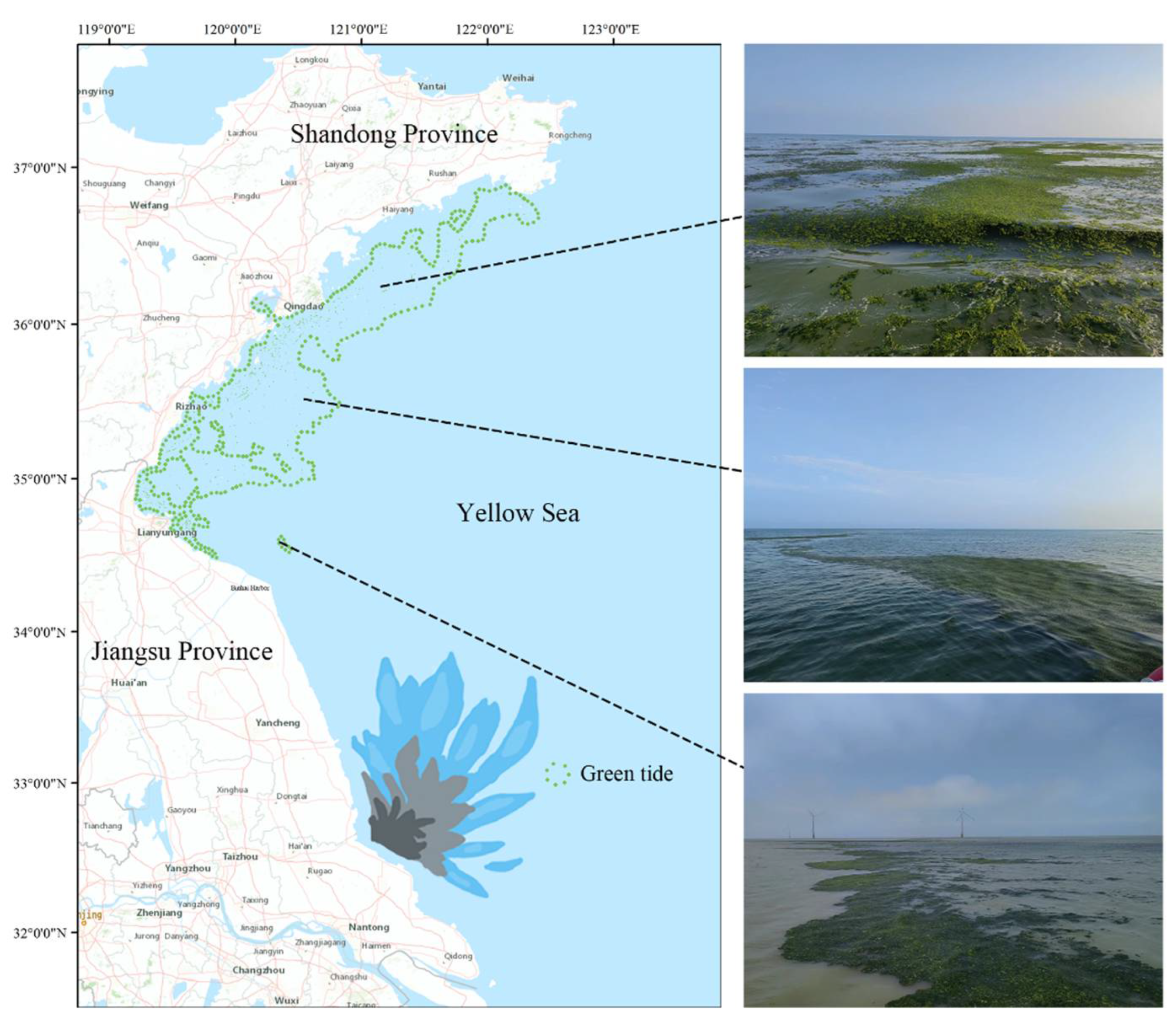

{kind=link}
{kind=link}
{kind=link}
| Allelochemical Classification | Allelochemicals | Chemical Formula | Chemical Structure | Reference |
|---|---|---|---|---|
| Fatty acid | Palmitic acid | C16H32O2 | 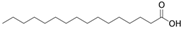 | [38,39] |
| Oleic acid | C18H34O2 | 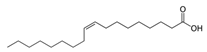 | [39] | |
| Nonanoic acid | C9H18O2 | 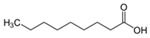 | [40] | |
| Aldehyde | Aldehyde 2-trans | / | [41] | |
| 4-trans-decadienal | / | [41] | ||
| Citral | C10H16O | 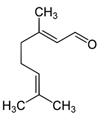 | [40] | |
| Phenol | Eugenol | C10H12O2 | 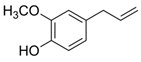 | [42] |
| Phenol | C6H5OH | 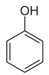 | [43] | |
| Pyrocatechol | C6H4(OH)2 | 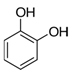 | [44] | |
| Terpenoid | Betulinic acid | C30H48O3 | 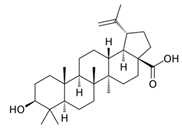 | [45] |
| Oleanolic acid | C30H48O3 | 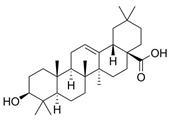 | [45] | |
| Ursolic acid | C30H48O3 | 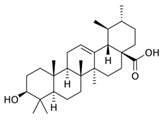 | [45] | |
| Myrcene | C10H16 | 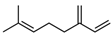 | [40] | |
| Alcohol | Sterol | / | [46] | |
| 1-Octanol | C8H18O | 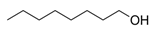 | [40] | |
| Alkaloid | Pyrrolopiperazine-2,5-dione | C7H10N2O2 | 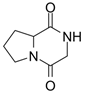 | [47] |
| Amide | N-phenethylacetamide | C10H13NO | 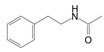 | [48,49] |
| Diketopiperazine derivatives | Cyclo(L-Pro-L-Val) | C10H16N2O2 | 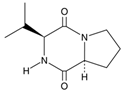 | [48,49] |
| Cyclo(L-Pro-L-Pro) | C10H14N2O2 | 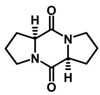 | [48,49] | |
| Amino acid | Valine | C5H11NO2 | 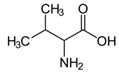 | [33] |
| Carboxylic acid | Hexanedioic acid | C6H10O4 | 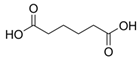 | [33] |
| Esters | Bis(2-ethylhexyl) ester | C24H38O4 | 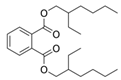 | [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Y.; Yang, X.; Xia, Z.; Chen, R.; He, F.; Zhang, J.; He, P. Review of Allelopathy in Green Tides: The Case of Ulva prolifera in the South Yellow Sea. Biology 2024, 13, 456. https://doi.org/10.3390/biology13060456
Zeng Y, Yang X, Xia Z, Chen R, He F, Zhang J, He P. Review of Allelopathy in Green Tides: The Case of Ulva prolifera in the South Yellow Sea. Biology. 2024; 13(6):456. https://doi.org/10.3390/biology13060456
Chicago/Turabian StyleZeng, Yinqing, Xinlan Yang, Zhangyi Xia, Runze Chen, Faqing He, Jianheng Zhang, and Peimin He. 2024. "Review of Allelopathy in Green Tides: The Case of Ulva prolifera in the South Yellow Sea" Biology 13, no. 6: 456. https://doi.org/10.3390/biology13060456
APA StyleZeng, Y., Yang, X., Xia, Z., Chen, R., He, F., Zhang, J., & He, P. (2024). Review of Allelopathy in Green Tides: The Case of Ulva prolifera in the South Yellow Sea. Biology, 13(6), 456. https://doi.org/10.3390/biology13060456