
The Perpetual Vector Mosquito Threat and Its Eco-Friendly Nemeses

 ,
,  and
and 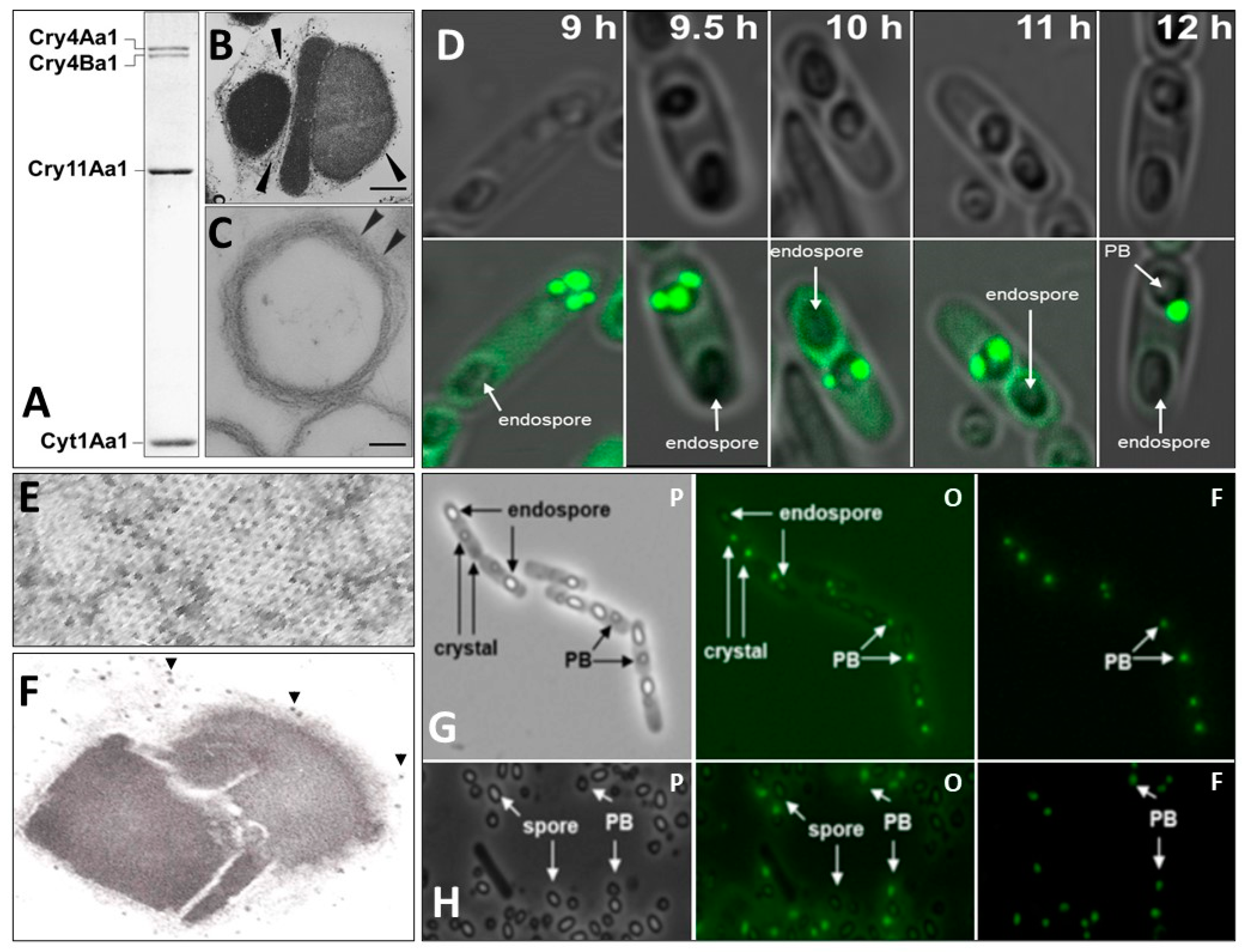{kind=link}
Simple Summary
Abstract
1. Introduction
1.1. A Glimpse of the Toll of Mosquito-Borne Diseases in Human Populations
1.2. A Glimpse of the Toll of Mosquito-Borne Diseases in Animals
1.2.1. West Nile Virus (WNV)
1.2.2. Japanese Encephalitis Virus (JEV)
2. The Expanding Mosquito Range, Climate Change, and Computational Modeling
3. Integrated Pest Management Programs (IPMPs) for Mosquito Control
3.1. Synthetic Pesticides
3.2. Avoidance of Pesticide through Olfaction
4. Effective “Green” Technologies
4.1. Sterile Insect Technique (SIT)
4.2. Wolbachia and Incompatible Insect Technique (IIT)
Wolbachia Can Enhance Pathogen Proliferation
4.3. Combined SIT–IIT
4.4. Release of Insect Carrying a Dominant Lethal Gene (RIDL)
RIDL OX513A—Success and Concerns
4.5. CRISPR/Cas9 Gene Drive
4.6. CRISPR-Based Engineering of Mosquitoes Refractive to Pathogens
4.6.1. CRISPR/Cas9
4.6.2. CRISPR/Cas13
5. Eco-Friendly Lysinibacillus sphaericus and Bacillus thuringiensis subsp. israelensis
5.1. Brief History of Lysinibacillus (Formerly Bacillus) sphaericus (Ls)
5.1.1. Structural Characteristics of Tpp1Aa1/Tpp2Aa1
5.1.2. Mechanism of Toxicity of Tpp1A1/Tpp2Aa1
5.2. Brief Historical Account of Bacillus thuringiensis Strains—A Collective of Highly Specific Insect Larvicides
5.2.1. Bacillus thuringiensis subsp. israelensis, Bti
5.2.2. Bti, the Most Robust and Efficacious Natural Mosquito Larvicidal Bacterium Known
5.2.3. Structural Characteristics of Bti’s Larvicidal Proteins
5.2.4. Mechanism of Toxicity of Bti’s Larvicidal Proteins
5.2.5. Prospects for Engineering More Robust Strains of Bti
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vector-Borne Diseases—World Health Organization (WHO). Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 15 February 2024).
- Kalluri, S.; Gilruth, P.; Rogers, D.; Szczur, M. Surveillance of arthropod vector-borne infectious diseases using remote sensing techniques: A review. PLoS Pathog. 2007, 3, e116. [Google Scholar] [CrossRef]
- WHO 2020. Elimination of human onchocerciasis: Progress report, 2020. Wkly. Epidemiol. Rec. 2021, 96, 557–567. [Google Scholar]
- Weaver, S.C.; Charlier, C.; Vasilakis, N.; Lecuit, M. Zika, Chikungunya, and other emerging vector-borne viral diseases. Annu. Rev. Med. 2018, 69, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Young, P.R. Arboviruses: A family on the move. In Dengue and Zika: Control and Antiviral Treatment Strategies; Hilgenfeld, R., Vasudevan, S., Eds.; Springer: New York, NY, USA, 2018; pp. 1–10. [Google Scholar]
- WHO 2023. World Malaria Report 2023. Available online: https://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2023 (accessed on 15 February 2024).
- Idro, R.; Marsh, K.; John, C.C.; Newton, C.R.J. Cerebral malaria: Mechanisms of brain injury and strategies for improved neurocognitive outcome. Pediatr. Res. 2010, 64, 267–274. [Google Scholar] [CrossRef]
- Beare, N.A.V. Cerebral malaria -using the retina to study the brain. Eye 2023, 37, 2379–2384. [Google Scholar] [CrossRef]
- Wolfe, C.M.; Barry, A.; Campos, A.; Farham, B.; Achu, D.; Juma, E.; Kalu, A.; Impouma, B. Control, elimination, and eradication efforts for neglected tropical diseases in the World Health Organization African region over the last 30 years—A scoping review. Int. J. Infect. Dis. 2024, 141, 106943. [Google Scholar] [CrossRef] [PubMed]
- Neglected Tropical Diseases—World Health Organization (WHO). Available online: www.who.int/news-room/questions-and-answers/item/neglected-tropical-diseases (accessed on 15 February 2024).
- Schaub, B.; Gueneret, M.; Jolivet, E.; Decatrelle, V.; Yazza, S.; Gueye, H.; Monthieux, A.; Juve, M.L.; Gautier, M.; Najioullah, F.; et al. Ultrasound imaging for identification of cerebral damage in congenital Zika virus syndrome: A case series. Lancet Child Adolesc. Heath. 2017, 1, 45–155. [Google Scholar] [CrossRef] [PubMed]
- Lessa, C.L.S.; Hodel, K.V.S.; Gonçalves, M.d.S.; Machado, B.A.S. Dengue as a Disease Threatening Global Health: A Narrative Review Focusing on Latin America and Brazil. Trop. Med. Infect. Dis. 2023, 8, 241. [Google Scholar] [CrossRef]
- WHO 2023 Dengue Stats Dengue and Severe Dengue. Available online: https://www.who.int/news-room/dengue-and-severe-dengue (accessed on 15 February 2024).
- Poungou, N.; Sevidzem, S.L.; Koumba, A.A.; Koumba, C.R.Z.; Mbehang, P.; Onanga, R.; Zahouli, J.Z.B.; Maganga, G.D.; Djogbénou, L.S.; Borrmann, S.; et al. Mosquito-Borne Arboviruses Occurrence and Distribution in the Last Three Decades in Central Africa: A Systematic Literature Review. Microorganisms 2024, 12, 4. [Google Scholar] [CrossRef]
- Johnston, B.L.; Conly, J.M. West Nile virus– Where did it come from and where might it go? Can. J. Infect. Dis. 2020, 11, 4. [Google Scholar] [CrossRef]
- Kramer, L.A.; Ciota, A.T.; Kilpatrick, A.M. Introduction, Spread, and Establishment of West Nile Virus in the Americas. J. Med. Entomol. 2021, 56, 1448–1455. [Google Scholar] [CrossRef] [PubMed]
- Ronca, S.E.; Ruff, J.C.; Murray, K.O. A 20-year historical review of West Nile virus since its initial emergence in North America: Has West Nile virus become a neglected tropical disease? PLoS Negl. Trop. Dis. 2021, 15, e0009190. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; LaDeau, S.L.; Marra, P.P. Ecology of West Nile virus transmission and its impact on birds in te Western Hemisphere. Auk 2007, 124, 1121–1136. [Google Scholar] [CrossRef]
- Habarugira, G.; Suen, W.W.; Hobson-Peters, J.; Hall, R.A.; Bielefeldt-Ohmann, H. West Nile Virus: An Update on Pathobiology, Epidemiology, Diagnostics, Control and “One Health” Implications. Pathogens 2020, 9, 589. [Google Scholar] [CrossRef]
- Pandit, P.S.; Doyle, M.M.; Smart, K.M.; Young, C.C.W.; Drape, G.W.; Johnson, C.K. Predicting wildlife reservoirs and global vulnerability to zoonotic Flaviviruses. Nat. Commun. 2018, 9, 5425. [Google Scholar] [CrossRef] [PubMed]
- Root, J.J.; Bosco-Lauth, A.M. West Nile Virus associations in wild animals: An update. Viruses 2019, 11, 11459. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Daszak, P.; Goodman, S.J.; Rogg, H.; Kramer, L.D.; Cedeño, V.; Cunningham, A.A. Predicting pathogen introduction: West Nile virus spread to Galápagos. Conserv. Biol. 2006, 20, 1224–1231. [Google Scholar] [CrossRef]
- Rappole, J.H.; Derrickson, S.R.; Hubálek, Z. Migratory Birds and Spread of West Nile Virus in the Western Hemisphere. Emerg. Infect. Dis. 2000, 6, 319–328. [Google Scholar] [CrossRef]
- Vannice, K.S.; Hills, S.L.; Schwartz, L.M.; Barrett, A.D.; Heffelfinger, J.; Hombach, J.; Letson, G.W.; Solomon, T.; Marfin, A.A. Japanese Encephalitis Vaccination Experts Panel. The future of Japanese encephalitis vaccination: Expert recommendations for achieving and maintaining optimal JE control. NPJ Vaccines 2021, 6, 82. [Google Scholar] [CrossRef]
- Park, S.L.; Huang, Y.S.; Vanlandingham, D.L. Re-Examining the Importance of Pigs in the Transmission of Japanese Encephalitis Virus. Pathogens 2022, 11, 575. [Google Scholar] [CrossRef]
- Park, S.L.; Huang, Y.S.; Lyons, A.C.; Ayers, V.B.; Hettenbach, S.M.; McVey, D.S.; Noronha, L.E.; Burton, K.R.; Higgs, S.; Vanlandingham, D.L. Infection of Feral Phenotype Swine with Japanese Encephalitis Virus. Vector Borne Zoonotic Dis. 2023, 23, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Yakob, L.; Hu, W.; Frentiu, F.D.; Gyawali, N.; Hugo, L.E.; Johnson, B.; Lau, C.; Furuya-Kanamori, L.; Magalhaes, R.S.; Devine, G. Japanese Encephalitis Emergence in Australia: The Potential Population at Risk. Clin. Infect. Dis. 2023, 76, 335–337. [Google Scholar] [CrossRef]
- Monath, T.P. Japanese encephalitis: Risk of emergence in the unted states. Viruses 2024, 16, 54. [Google Scholar] [CrossRef] [PubMed]
- Martens, W.J.; Niessen, L.W.; Rotmans, J.; Jetten, T.H.; McMichael, A.J. Potential impact of global climate change on malaria risk. Environ. Health Perspect 1995, 103, 458–464. [Google Scholar] [CrossRef]
- Kraemer, M.U.G.; Reiner, R.C., Jr.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Lippi, C.A.; Stewart-Ibarra, A.M.; Loor, M.E.F.B.; Zambrano, J.E.D.; Lopez, N.A.E.; Blackburn, J.K.; Ryan, S.J. Geographic shifts in Aedes aegypti habitat suitability in Ecuador using larval surveillance data and ecological niche modeling: Implications of climate change for public health vector control. PLoS Negl. Trop. Dis. 2019, 13, e0007322. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.J.; Carlson, C.J.; Mordecai, E.A.; Johnson, L.R. Global expansion and redistribution of Aedes-borne virus transmission risk with climate change. PLoS Negl. Trop. Dis. 2019, 13, e0007213. [Google Scholar] [CrossRef]
- Mordecai, E.A.; Ryan, S.J.; Caldwell, J.M.; Shah, M.M.; LaBeaud, A.D. Climate change could shift disease burden from malaria to arboviruses in Africa. Lancet Planet Health 2020, 4, e416–e423. [Google Scholar] [CrossRef]
- Colón-González, F.J.; Sewe, M.O.; Tompkins, A.M.; Sjödin, H.; Casallas, A.; Rocklöv, J.; Caminade, C.; Lowe, R. Projecting the risk of mosquito-borne diseases in a warmer and more populated world: A multi-model, multi-scenario intercomparison modelling study. Lancet Planet Health 2021, 5, e404–e414. [Google Scholar] [CrossRef]
- Mora, C.; McKenzie, T.; Gaw, I.M.; Dean, J.M.; von Hammerstein, H.; Knudson, T.A.; Setter, R.O.; Smith, C.Z.; Webster, K.M.; Patz, J.A.; et al. Over half of known human pathogenic diseases can be aggravated by climate change. Nat. Clim. Chang. 2022, 12, 869–875. [Google Scholar] [CrossRef]
- Brugueras, S.; Fernández-Martínez, B.; Martínez-de la Puente, J.; Figuerola, J.; Porro, T.M.; Rius, C.; Larrauri, A.; Gómez-Barroso, D. Environmental drivers, climate change and emergent diseases transmitted by mosquitoes and their vectors in southern Europe: A systematic review. Environ. Res. 2020, 191, 110038. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.E.; Mahmud, A.S.; Millar, I.F.; Rajeev, A.; Rasambainarivo, F.; Rice, B.L.; Takahashii, S.; Tatem, A.J.; Wagner, C.E.; Wang, L.-F.; et al. Infectious disease in the era of global change. Nat. Rev. Microbiol. 2022, 20, 193–205. [Google Scholar] [CrossRef]
- Gona, P.N.; More, A.F. Bacterial pathogens and climate change. Lancet 2022, 400, 2161–2163. [Google Scholar] [CrossRef]
- Semenza, J.C.; Rocklöv, J.; Ebi, K.L. Climate Change and Cascading Risks from Infectious Disease. Infect. Dis Ther. 2022, 11, 1371–1390. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, A.A.; Susanna, D.; Febrian, T. The relationship between climate change and malaria in South-East Asia: A systematic review of the evidence. F1000 Res. 2023, 11, 1555. [Google Scholar] [CrossRef] [PubMed]
- Shah, I.H.; Manzoor, M.A.; Jinhui, W.; Li, X.; Hameed, M.K.; Rehaman, A.; Li, P.; Zhang, Y.; Niu, Q.; Chang, L. Comprehensive review: Effects of climate change and greenhouse gases emission relevance to environmental stress on horticulture crops and managements. J. Environ. Manag. 2024, 351, 119978. [Google Scholar] [CrossRef]
- Samuel, G.H.; Adelman, Z.N.; Myles, K.M. Temperature-dependent effects on the rteplication and transmission of arthropod-borne viruses in their insect hosts. Curr. Opin. Insect Sci. 2016, 16, 108–113. [Google Scholar] [CrossRef]
- Richards, S.L.; Mores, C.N.; Lord, C.C.; Tabachnick, W.J. Impact of extrinsic incubation temperature and virus exposure on vector competence of Culex pipiens quinquefasciatus say (diptera: Culicidae) for west nile virus. Vector Borne Zoonotic Dis. 2007, 7, 629–636. [Google Scholar] [CrossRef]
- Alto, B.W.; Bettinardi, D. Temperature and dengue virus infection in mosquitoes: Independent effects on the immature and adult stages. Am. J. Trop. Med. Hyg. 2013, 88, 497–505. [Google Scholar] [CrossRef]
- Couper, L.I.; Farner, J.E.; Lyberger, K.P.; Lee, A.S.; Mordecai, E.A. Mosquito thermal tolerance is remarkably constrained across a large climatic range. bioRxiv 2023. bioRxiv 2023.03.02.530886 Update in Proc. Biol. Sci. 2024, 291, 20232457. [Google Scholar] [CrossRef]
- Bell, G. Evolutionary rescue. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 605–627. [Google Scholar] [CrossRef]
- Gomulkiewicz, R.; Shaw, R.G. Evolutionary rescue beyond the models. Phil. Trans. R Soc. B 2012, 368, 20120093. [Google Scholar] [CrossRef]
- Carlson, C.J.; Bannon, E.; Mendenhall, E.; Newfield, T.; Bansal, S. Rapid range shifts in African Anopheles mosquitoes over the last century. Biol. Lett. 2023, 19, 20220365. [Google Scholar] [CrossRef]
- Hernandez-Valencia, J.C.; Muñoz-Laiton, P.; Gómez, G.F.; Correa, M.M. A Systematic Review on the Viruses of Anopheles Mosquitoes: The Potential Importance for Public Health. Trop. Med. Infect. Dis. 2023, 8, 459. [Google Scholar] [CrossRef] [PubMed]
- Masson-Delmotte, Z.; Pirani, C. Climate change 2021, the physical science basis. In Intergovernmental Panel on Climate Change; World Meteorological Organization: Geneva, Switzerland, 2021; Available online: https://report.ipcc.ch/ar6/wg1/IPCC_AR6_WGI_FullReport.pdf (accessed on 16 February 2024).
- Zardini, A.; Menegale, F.; Gobbi, A.; Manica, M.; Guzzetta, G.; d’Andrea, V.; Marziano, V.; Trentini, F.; Montarsi, F.; Caputo, B.; et al. Estimating the potential risk of transmission of arboviruses in the Americas and Europe: A modelling study. Lancet Planet Health 2024, 8, e30–e40. [Google Scholar] [CrossRef]
- Gossner, C.M.; Ducheyne, E.; Schafner, F. Increased risk for autochthonous vector-borne infections transmitted by Aedes albopictus in continental europe. Eurosurveillance 2018, 23, 2–7. [Google Scholar] [CrossRef]
- ECDC (European Centre for Disease Prevention and Control and European Food Safety Authority). Aedes albopictus—Current Known Distribution: September 2020. Mosquito Maps. Available online: https://ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data/mosquito-maps (accessed on 15 February 2024).
- Oliveira, S.; Rocha, J.; Sousa, C.A.; Capinha, C. Wide and increasing suitability for Aedes albopictus in Europe is congruent across distribution models. Sci. Rep. 2021, 11, 9916. [Google Scholar] [CrossRef] [PubMed]
- Vector Disease Control International. Available online: https://www.vdci.net/blog/the-key-components-of-an-integrated-mosquito-management-program-0/#:~:text=The%20Key%20Components%20of%20an%20Integrated%20Mosquito%20Management,Public%20Education%20..%207%207.%20Record%20keeping%20 (accessed on 5 March 2024).
- Organização Pan-Americana da Saúde. Documento Operacional Para a Execução do Manejo Integrado de Vetores Adaptado ao Contexto das Américas; OPAS: Washington, DC, USA, 2019. [Google Scholar]
- Lees, R.S.; Fornadel, C.; Snetselaar, J.; Wagman, J.; Spiers, A. Insecticides for Mosquito Control: Improving and Validating Methods to Strengthen the Evidence Base. Insects 2023, 14, 116. [Google Scholar] [CrossRef]
- Kleinschmidt, I.; Rowland, M. Insecticides and malaria. In Ecology and Control of Vector-Borne Diseases Volume 6. Innovative Strategies for Vector Control; Wageningen Academic Publishers: Wageningen, The Netherlands, 2021. [Google Scholar] [CrossRef]
- Saeed-ul-Hassan, S.; Tariq, I.; Khalid, A.; Karim, S. Comparative clinical study on the efectiveness of homeopathic combination remedy with standard maintenance therapy for dengue fever. Trop. J. Pharm. Res. 2013, 12, 767–770. [Google Scholar]
- Katritzky, A.R.; Wang, Z.; Slavov, S.; Tsikolia, M.; Dobchev, D.; Akhmedov, N.G.; Hall, C.D.; Bernier, U.R.; Clark, G.G.; Linthicum, K.J. Synthesis and bioassay of improved mosquito repellents predicted from chemical structure. Proc. Natl. Acad. Sci. USA 2008, 105, 7359–7364. [Google Scholar] [CrossRef]
- Pal, M.; Kumar, A.; Tewari, K.S. Chemical composition and mosquito repellent activity of the essential oil of Plectranthusincanus link. Factauniversitatis-Ser. Phys. Chem. Technol. 2011, 9, 57–64. [Google Scholar] [CrossRef]
- Onyett, H. Preventing mosquito and tick bites: A Canadian update. Paediatr. Child Health 2014, 19, 329–333. [Google Scholar]
- Tripathi, A.K.; Upadhyay, S.; Bhuiyan, M.; Bhattacharya, P.R. A review on prospects of essential oils as biopesticide in insectpest management. J. Pharmacogn. Phytother. 2009, 1, 52–63. [Google Scholar]
- Mansouri, A.; Cregut, M.; Abbes, C.; Durand, M.J.; Landoulsi, A.; Thouand, G. The environmental issues of DDT pollution and bioremediation: A multidisciplinary review. Appl. Biochem. Biotechnol. 2017, 181, 309–339. [Google Scholar] [CrossRef]
- Harada, T.; Takeda, M.; Kojima, S.; Tomiyama, N. Toxicity and Carcinogenicity of Dichlorodiphenyltrichloroethane (DDT). Toxicol. Res. 2016, 32, 3221–3233. [Google Scholar] [CrossRef]
- Fu, H.; Tang, P.; Wang, R.; Li, S.; Liu, H.; Yang, Y.; Wu, Z. Advances in organophosphorus pesticides pollution: Current status and challenges in ecotoxicological, sustainable agriculture, and degradation strategies. J. Hazard. Mater. Adv. 2022, 424, 127494. [Google Scholar] [CrossRef]
- Ahamad, A.; Kumar, J. Pyrethroid pesticides: And overview on classification, toxicological assessment and monitoring. J. Hazard. Mater. Adv. 2023, 10, 100284. [Google Scholar] [CrossRef]
- Buckingham, S.D.; Ihara, M.; Sattelle, D.B.; Matsuda, K. Mechanisms of Action, Resistance and Toxicity of Insecticides Targeting GABA Receptors. Curr. Med. Chem. 2017, 24, 2935–2945. [Google Scholar] [CrossRef] [PubMed]
- Corbel, V.; Duchon, S.; Zaim, M.; Hougard, J.M. A Po-tential Neonicotinoid Insecticide Against Resistant Mosquitoes. J. Med. Entomol. 2004, 41, 712–717. [Google Scholar] [CrossRef] [PubMed]
- Ashu, F.A.; Fouet, C.; Ambadiang, M.M.; Penlap-Beng, V.; Kamdem, C. Vegetable oilbased surfactants are adjuvants that enhance the efficacy of neonicotinoid insecticides and can bias sus-ceptibility testing in adult mosquitoes. PLoS Negl. Trop. Dis. 2023, 17, e0011737. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Ihara, M.; Sattelle, D.B. Neonicotinoid Insecticides: Molecular Targets, Resistance, and Toxicity. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 241–255. [Google Scholar] [CrossRef]
- Romi, R.; Proietti, S.; Di Luca, M.; Cristofaro, M. Laboratory evaluation of the bioinsecticide Spi-nosad for mosquito control. J. Am. Mosq. Control Assoc. 2006, 22, 93–96. [Google Scholar] [CrossRef]
- Santos, V.S.V.; Pereira, B.B. Properties, toxicity and current applications of the biolarvicide spi-nosad. J. Toxicol Env. Health B Crit. Rev. 2020, 23, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Crouse, G.D.; Benko, Z.; Demeter, D.; Giampietro, N.C.; Lambert, W.; Brown, A.V. The spinosyns, spinosad, spinetoram, and synthetic spinosyn mimics–Discovery, exploration, and evolution of a natural product chemistry and the impact of computational tools. Pest. Manag. Sci. 2021, 77, 3637–3649. [Google Scholar] [CrossRef] [PubMed]
- Kirst, H. The spinosyn family of insecticides: Realizing the potential of natural products re-search. J. Antibiot. 2010, 63, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Black, B.C.; Hollingworth, R.M.; Ahmmadsahib, K.I.; Kukel, C.D.; Donovan, S. Insecticidal action and mitochondrial uncoupling activity of AC-303,630 and related halogenated pyrroles. Pestic. Biochem. Physiol. 1994, 50, 115–128. [Google Scholar] [CrossRef]
- Oxborough, R.M.; N’Guessan, R.; Jones, R.; Kitau, J.; Ngufor, C.; Malone, D.; Mosha, F.W.; Rowland, M.W. The activity of the pyrrole insecticide chlorfenapyr in mosquito bioassay: Towards a more rational testing and screening of non-neurotoxic insecticides for malaria vector control. Malar. J. 2015, 14, 124. [Google Scholar] [CrossRef]
- Mosha, F.W.; Lyimo, I.N.; Oxborough, R.M.; Malima, R.; Tenu, F.; Matowo, J.; Feston, E.; Mndeme, R.; Magesa, S.M.; Rowland, M. Experimental hut evaluation of the pyrrole insecticide chlorfenapyr on bed nets for the control of Anopheles arabiensis and Culex quinquefasciatus. Trop. Med. Int. Health 2008, 13, 644–652. [Google Scholar] [CrossRef]
- Che-Mendoza, A.; González-Olvera, G.; Medina-Barreiro, A.; Arisqueta-Chablé, C.; Bibiano-Marin, W.; Correa-Morales, F.; Kirstein, O.D.; Manrique-Saide, P.; Vazquez-Prokopec, G.M. Efficacy of targeted indoor residual spraying with the pyrrole insecticide chlorfenapyr against pyrethroid-resistant Aedes aegypti. PLoS Negl. Trop. Dis. 2021, 15, e0009822. [Google Scholar] [CrossRef]
- Lawler, S.P. Environmental safety review of methoprene and bacterially-derived pesti-cides commonly used for sustained mosquito control. Ecotoxicol. Environ. Saf. 2017, 139, 335–343. [Google Scholar] [CrossRef]
- Mmbaga, A.T.; Lwetoijera, D.W. Current and future opportunities of autodissemination of pyriproxyfen approach for malaria vector control in urban and rural Africa. Wellcome Open Res. 2023, 8, 119. [Google Scholar] [CrossRef]
- Hustedt, J.C.; Boyce, R.; Bradley, J.; Hii, J.; Alexander, N. Use of pyriproxyfen in control of Aedes mosquitoes: A systematic review. PLoS Negl. Trop. Dis. 2020, 14, e0008205. [Google Scholar] [CrossRef] [PubMed]
- Liu, A. Insectcide resistance in Mosquitoes: Impact, mechanisms, and research directions. Ann. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Cuervo-Parra, J.A.; Romero Cortes, T.; Ramirez-Lepe, M. Insect Resist; Trdan, S., Ed.; Tech Publishers: Rijeka, Croatia, 2016; Chapter 9. [Google Scholar]
- Kaur, K.; Choudhary, D.; Bali, S.; Bandral, S.S.; Singh, V.; Ahmad, M.A.; Rani, N.; Singh, T.G.; Chandrasekaran, B. Pesticides: An alarming detrimental to health and environment. Sci. Total Environ. 2023, 915, 170113. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Mittal, P.K.; Kumar, G.; Dhiman, R.C. Insecticide susceptibility status of Aedes aegypti and Anopheles stephensi larvae against temephos in Delhi, India. Int. J. Mosq. Res. 2014, 1, 69–73. [Google Scholar]
- Moyes, C.L.; Vontas, J.; Martins, A.J.; Ng, L.C.; Koou, S.Y.; Dusfour, I.; Raghavendra, K.; Pinto, J.; Corbel, V.; David, J.P.; et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl. Trop. Dis. 2017, 11, e0005625. [Google Scholar] [CrossRef]
- Asgarian, T.S.; Vatandoost, H.; Hanafi-Bojd, A.A.; Nikpoor, F. Worldwide Status of Insecticide Resistance of Aedes aegypti and Ae. albopictus, Vectors of Arboviruses of Chikungunya, Dengue, Zika and Yellow Fever. J. Arthropod Borne Dis. 2023, 17, 1–27. [Google Scholar] [CrossRef]
- Azarm, A.; Nasrabadi, M.; Shahidi, F.; Dehghan, A.; Nikpoor, F.; Zahraie-Ramazani, A.; Molaeezadeh, S.M.; Bozorgomid, F.; Tashakori, G.; Vatandoost, H. Insecticide Resistance in the West Nile Encephalitis, Japanese Encephalitis, Avian Malaria and Lymphatic Elephantiasis Vector, Culex pipiens complex (Diptera: Culicidae) in Iran. J. Arthropod Borne Dis. 2021, 15, 349–357. [Google Scholar] [CrossRef]
- Suh, P.; Elanga-Ndille, E.; Tchouakui, M.; Sandeu, M.M.; Tagne, D.; Wondji, C.; Ndo, C. Impact of insecticide resistance on malaria vector competence: A literature review. Malar. J. 2023, 22, 19. [Google Scholar] [CrossRef]
- Nauen, R.; Bass, C.; Feyereisen, V.J. The role of cytochrome P450s in insect toxicology and resistance. Annu. Rev. Entomol. 2022, 67, 105–124. [Google Scholar] [CrossRef]
- Che-Mendoza, A.; Penilla, R.P.; Rodriguez, S.A. Insecticide resistance and glutathione S-transferases in mosquitoes: A review. Afr. J. Biotechnol. 2009, 8. [Google Scholar]
- Pavlidi, N.; Vontas, J.; Van Leeuwen, T. The role of glutathione S-transferases 9GSTs) in insecticide resistance in crop pests and disease vectore. Curr. Opin. Insect Sci. 2018, 27, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Karunaratne, S.H.P.P.; De Silva, W.A.P.P.; Weeraratne, T.C.; Surendran, S.N. Insecticide resistance in mosquitoes: Development, mechanisms and monitoring. Ceylon J. Sci. 2018, 47, 299–309. [Google Scholar] [CrossRef]
- Senthil-Nathan, S. A review of resistance mechanisms of synthetic insecticides and botanicals, phytochemicals, and essential oli as alternative larvicidal agents against mosquitoes. Front. Physiol. 2020, 10, 1591. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhang, L.; Li, T.; Zhang, L.; He, L.; Dong, K.; Liu, N. Evolutionary Adaptation of the Amino Acid and Codon Usage of the Mosquito Sodium Channel following Insecticide Selection in the Field Mosquitoes. PLoS ONE 2012, 7, e47609. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, L.; Reid, W.R.; Xu, Q.; Dong, K.; Liu, N. Multiple mutations and mutation combinations in the sodium channel of permethrin resistant mosquitoes, Culex Quinquefasciatus. Sci. Rep. 2012, 2, 781. [Google Scholar] [CrossRef] [PubMed]
- Alout, H.; Djogbénou, L.; Berticat, C.; Chandre, F.; Weill, M. Comparison of Anopheles gambiae and Culex pipiens acetycholinesterase 1 biochemical properties. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 150, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Engdahl, C.; Knutsson, S.; Fredriksson, S.A.; Linusson, A. Acetylcholinesterases from the disease vectors Aedes aegypti and Anopheles gambiae: Functional characterization and comparisons with vertebrate orthologues. PLoS ONE 2015, 10, e0138598. [Google Scholar] [CrossRef] [PubMed]
- Weill, M.; Fort, P.; Berthomieu, A.; Dubois, M.P.; Pasteur, N.; Raymond, M. A novel acetylcholinesterase gene in mosquitoes codes for the insecticide target and is non–homologous to the ace gene Drosophila. Proc. R. Soc. London. Ser. B Biol. Sci. 2002, 269, 2007–2016. [Google Scholar] [CrossRef]
- Weill, M.; Lutfalla, G.; Mogensen, K.; Chandre, F.; Berthomieu, A.; Berticat, C.; Pasteur, N.; Philips, A.; Fort, P.; Raymond, M. Insecticide resistance in mosquito vectors. Nature 2003, 423, 136–137. [Google Scholar] [CrossRef]
- Du, W.; Awolola, T.S.; Howell, P.; Koekemoer, L.L.; Brooke, B.D.; Benedict, M.Q.; Coetzee, M.; Zheng, L. Independent mutations in the Rdl locus confer dieldrin resistance to Anopheles gambiae and An. arabiensis. Insect Mol. Biol. 2005, 14, 179–183. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H.; Anthony, N.; Aronstein, K.; Rocheleau, T.; Stilwell, G. Cyclodiene insecticide resistance: From molecular to population genetics. Annu. Rev. Entomol. 2000, 45, 449–466. [Google Scholar] [CrossRef]
- Davari, B.; Vatandoost, H.; Oshaghi, M.A.; Ladonni, H.; Enayati, A.A.; Shaeghi, M.; Basseri, H.R.; Rassi, Y.; Hanafi-Bojd, A.A. Selection of Anopheles stephensi with DDT and dieldrin and cross-resistance spectrum to pyrethroids and fipronil. Pestic. Biochem. Physiol. 2007, 89, 97–103. [Google Scholar] [CrossRef]
- Wondji, C.S.; Dabire, R.K.; Tukur, Z.; Irving, H.; Djouaka, R.; Morgan, J.C. Identification and distribution of a GABA receptor mutation conferring dieldrin resistance in the malaria vector Anopheles funestus in Africa. Insect Biochem. Mol. Biol. 2011, 41, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Cupp, E.W.; Micher, K.M.; Guo, A.; Liu, N. Insecticide resistance and cross-resistance in Alabama and Florida strains of Culex quinquefaciatus. J. Med. Entomol. 2004, 1, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xu, Q.; Zhang, L.; Liu, N. Chlorpyrifos resistance in the mosquito Culex quinquefasciatus. J. Med. Entomol. 2005, 42, 815–820. [Google Scholar] [CrossRef]
- Bhat, R.; Axtell, R.; Mitra, A.; Miranda, M.; Lock, C.; Tsien, R.W.; Steinman, L. Inhibitory role for GABA in autoimmune inflammation. Proc. Natl Acad. Sci. USA 2010, 107, 2580–2585. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.W.; Thompson, A.J.; Corletto, F.; Reckless, J.; Loke, J.C.; Lapaque, N.; Grant, A.J.; Mastroeni, P.; Grainger, D.J.; Padgett, C.L.; et al. Anaesthetic impairment of immune function is mediated via GABA(A) receptors. PLoS ONE 2011, 6, e17152. [Google Scholar] [CrossRef]
- Fuks, J.M.; Arrighi, R.B.; Weidner, J.M.; Kumar Mendu, S.; Jin, Z.; Wallin, R.P.; Rethi, B.; Birnir, B.; Barragan, A. GABAergic signaling is linked to a hypermigratory phenotype in dendritic cells infected by Toxoplasma gondii. PLoS Pathog. 2012, 8, e1003051. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, R.; Zhang, B.; Zhao, T.; Wang, P.; Liang, G.; Cheng, G. Blood meal acquisition enhances arbovirus replication if mosquitoes through activation of the GABAergic system. Nat. Commun. 2017, 8, 1262. [Google Scholar] [CrossRef]
- Meister, S.; Kanzok, S.M.; Zheng, X.-L.; Luna, C.; Li, T.-R.; Hoa, N.T.; Clayton, J.R.; White, K.P.; Kafatos, F.C.; Christophides, G.K.; et al. Immune signaling pathways regulating bacterial and malaria parasite infection of the mosquito Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2005, 102, 11420–11425. [Google Scholar] [CrossRef]
- Lee, W.-S.; Webster, J.A.; Madzokere, E.T.; Stephenson, E.B.; Herrero, L.J. Mosquito antiviral defense mechanisms: A delicate balance between innate immunity and persistent viral infection. Parasite Vectors 2019, 12, 165. [Google Scholar] [CrossRef] [PubMed]
- Tikhe, C.V.; Dimopoulos, G. Mosquito antiviral immune pathways. Evelopmental Comp. Immunol. 2021, 116, 103964. [Google Scholar] [CrossRef]
- Wheelwright, M.; Whittle, C.R.; Riabinina, O. Olfactory sensation across mosquito species. Cell Tissue Res. 2021, 383, 75–90. [Google Scholar] [CrossRef]
- WHO Tube Test 2022. Standard Operating Procedure for Testing Insecticide Susceptibility of Adult Mosquitoes in WHO Tube Tests. Available online: https://www.who.int/publications/i/item/9789240043831 (accessed on 15 February 2024).
- Praulins, G.; McDermott, D.P.; Spiers, A.; Lees, R.S. Reviewing the WHO Tube Bioassay Methodology: Accurate method reporting and numbers of mosquitos are key to producing robust results. Insects 2022, 13, 544. [Google Scholar] [CrossRef]
- Sougoufara, S.; Yorkston-Dives, H.; Aklee, N.M.; Rus, A.C.; Zairi, J.; Tripet, F. Standardised bioassays reveal that mosquitoes learn to avoid compounds used in chemical vector control after a single sub-lethal exposure. Sci. Rep. 2022, 12, 2206. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Jeffries, C.L.; Walker, T. Biological control of mosquito vectors: Past, present, and future. Insects 2016, 7, 52. [Google Scholar] [CrossRef]
- Kaura, T.; Mewara, A.; Zaman, K.; Sehgal, R. Comparative efficacy of natural aquatic predators for biological control of mosquito larvae: A neglected tool for vector control. J. Vector Borne Dis. 2023, 60, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Bourtzis, K.; Mreysen, M.J.B. Sterile Insect Technique (SIT) and its applications. Insects 2021, 12, 638. [Google Scholar] [CrossRef]
- Ankita Agarwal, A.; Sarma, D.; Chaurasia, D.; Maan, H.S. Novel molecular approaches to combat vectors and vector-borne viruses: Special focus on RNA interference (RNAi) mechanism. Acta Trop. 2022, 233, 106539. [Google Scholar] [CrossRef]
- Nikolouli, K.; Colinet, H.; Renault, D.; Enriquez, T.; Mouton, L.; Gibert, P.; Sassu, F.; Cáceres, C.; Stauffer, C.; Pereira, R.; et al. 1018. Sterile insect technique and Wolbachia symbiosis as potential tools for the control of the invasive species Drosophila suzukii. J. Pest Sci. 2018, 91, 489–503. [Google Scholar] [CrossRef]
- Minwuyelet, A.; Petronio, G.P.; Yewhalaw, D.; Sciarretta, A.; Magnifico, I.; Nicolosi, D.; Di Marco, R.; Atenafu, G. Symbiotic Wolbachia in mosquitoes and its role in reducing the transmission of mosquito-borne diseases: Updates and prospects. Front. Microbiol. 2023, 14, 1267832. [Google Scholar] [CrossRef]
- Gantz, V.M.; Jasinskiene, N.; Tatarenkova, O.; Fazekas, A.; Macias, V.M.; Bier, E.; James, A.A. Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proc. Natl. Acad. Sci. USA 2015, 112, E6736–E6743. [Google Scholar] [CrossRef]
- Crickmore, N.; Berry, C.; Panneerselvam, S.; Mishra, R.; Connor, T.R.; Bonning, B.C. A structure-based nomenclature for Bacillus thuringiensis and other bacteria-derived pesticidal proteins. J. Invertebr. Pathol. 2021, 186, 107438. [Google Scholar] [CrossRef]
- Rudd, S.R.; Miranda, L.S.; Curtis, H.R.; Bigot, Y.; Diaz-Mendoza, M.; Hice, R.; Nizet, V.; Park, H.W.; Blaha, G.; Federici, B.A.; et al. The Parasporal Body of Bacillus thuringiensis subsp. israelensis: A Unique Phage Capsid-Associated Prokaryotic Insecticidal Organelle. Biology 2023, 12, 1421. [Google Scholar] [CrossRef]
- Knipling, E. Possibilities of insect control or eradication through the use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Knipling, E.F. Suppression of pest Lepidoptera by releasing partially sterile males: A theoretical appraisal. Bioscience 1970, 20, 465–470. [Google Scholar] [CrossRef]
- Patterson, R.S.; Weidhaas, D.E.; Ford, H.R.; Lofgren, C.S. Suppression and Elimination of an Island Population of Culex pipiens quinquefasciatus with Sterile Males. Science 1970, 168, 1368–1370. [Google Scholar] [CrossRef]
- Weidhaas, D.E.; Breeland, S.G.; Lofgren, C.S.; Dame, D.A.; Kaiser, R. Release of Chemosterilized Males for the Control of Anopheles albimanus in El Salvador. IV. Dynamics of the Test Population. Am. J. Trop. Med. Hyg. 1974, 23, 298–308. [Google Scholar] [CrossRef]
- Benedict, M.Q. Sterile Insect Technique: Lessons from the Past. J. Med. Entomol. 2021, 58, 1974–1979. [Google Scholar] [CrossRef]
- Bellini, R.; Medici, A.; Puggioli, A.; Balestrino, F.; Carrieri, M. Pilot Field Trials with Aedes albopictus Irradiated Sterile Males inItalian Urban Areas. J. Med. Entomol. 2013, 50, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Gato, R.; Menéndez, Z.; Prieto, E.; Argilés, R.; Rodríguez, M.; Baldoquín, W.; Hernández, Y.; Pérez, D.; Anaya, J.; Fuentes, I.; et al. Sterile Insect Technique: Successful Suppression of an Aedes aegypti Field Population in Cuba. Insects 2021, 12, 469. [Google Scholar] [CrossRef] [PubMed]
- Manana, P.N.; Jewett, S.; Zikhali, J.; Dlamini, D.; Mabaso, N.; Mlambo, Z.; Ngobese, R.; Munhenga, G. “Maskandi experience”: Exploring the use of a cultural song for community engagement in preparation for a pilot Sterile Insect Technique release programme for malaria vector control in KwaZulu-Natal Province, South Africa 2019. Malar. J. 2023, 20, 204. [Google Scholar] [CrossRef] [PubMed]
- Oliva, C.F.; Benedict, M.Q.; Collins, C.M.; Baldet, T.; Bellini, R.; Bossin, H.; Bouyer, J.; Corbel, V.; Facchinelli, L.; Fouque, F.; et al. Sterile Insect Technique (SIT) against Aedes Species Mosquitoes: A Roadmap and Good Practice Framework for Designing, Implementing and Evaluating Pilot Field Trials. Insects 2021, 12, 191. [Google Scholar] [CrossRef] [PubMed]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia?—A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef]
- Telschow, A.; Hilgenboecker, K.; Hammerstein, P.; Werren, J.H. Dobzhansky-muller and wolbachia-induced incompatibilities in a diploid genetic system. PLoS ONE 2014, 23, e95488. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.; Sullivan, W. The cellular lives of Wolbachia. Nat. Rev. Microbiol. 2023, 21, 750–766. [Google Scholar] [CrossRef]
- Laven, H. Eradication of Culex pipiens fatigans through cytoplasmic incompatibility. Nature 1967, 216, 383–384. [Google Scholar] [CrossRef]
- Pagendam, D.E.; Trewin, B.J.; Snoad, N.; Ritchie, S.A.; Hoffmann, A.A.; Staunton, K.M.; Paton, C.; Beebe, N. Modelling the Wolbachia incompatible insect technique: Strategies for effective mosquito population elimination. BMC Biol. 2020, 18, 161. [Google Scholar] [CrossRef]
- Schairer, C.E.; Najera, J.; James, A.A.; Akbari, O.S.; Bloss, C.S. Oxitec and MosquitoMate in the United States: Lessons for the future of gene drive mosquito control. Pathog. Glob. Health 2021, 115, 365–376. [Google Scholar] [CrossRef]
- Crawford, J.E.; Clarke, D.W.; Criswell, V.; Desnoyer, M.; Cornel, D.; Deegan, B.; Gong, K.; Hopkins, K.C.; Howell, P.; Hyde, J.S.; et al. Efficient production of male Wolbachia-infected Aedes aegypti mosquitoes enables largescale suppression of wild populations. Nat. Biotechnol. 2020, 38, 482–492. [Google Scholar] [CrossRef]
- Mains, J.W.; Kelly, P.H.; Dobson, K.L.; Petrie, W.D.; Dobson, S.L. Localized control of Aedes aegypti (Diptera: Culicidae) in Miami, FL, via inundative releases of Wolbachia-infected male mosquitoes. J. Med. Entomol. 2019, 56, 1296–1303. [Google Scholar] [CrossRef]
- Project Wolbachia–Singapore Consortium Wolbachiamediated sterility suppresses Aedes aegypti populations in the urban tropics. Medrxiv 2021. Medrxiv 2021-06. [CrossRef]
- Zheng, X.; Zhang, D.; Li, Y.; Yang, C.; Wu, Y.; Liang, X.; Xi, Z. Incompatible and sterile insect techniques combined eliminate mosquitoes. Nature 2019, 572, 56–61. [Google Scholar] [CrossRef]
- O’Neill, S.L.; Ryan, P.A.; Turley, A.P.; Wilson, G.; Retzki, K.; Iturbe-Ormaetxe, I.; Dong, Y.; Kenny, N.; Paton, C.J.; Ritchie, S.A.; et al. Scaled deployment of Wolbachia to protect the community from dengue and other Aedes transmitted arboviruses. Gates Open Res. 2019, 2, 36. [Google Scholar] [CrossRef]
- Indriani, C.; Tantowijoyo, W.; Rancès, E.; Andari, B.; Prabowo, E.; Yusdi, D.; Ansari, M.R.; Wardana, D.S.; Supriyati, E.; Nurhayati, I.; et al. Reduced dengue incidence following deployments of Wolbachia-infected Aedes aegypti in Yogyakarta, Indonesia: A quasi-experimental trial using controlled interrupted time series analysis. Gates Open Res. 2020, 4, 50. [Google Scholar] [CrossRef]
- Pinto, S.B.; Riback, T.I.S.; Sylvestre, G.; Costa, G.; Peixoto, J.; Dias, F.B.S.; Tanamas, S.K.; Simmons, C.P.; Dufault, S.M.; Ryan, P.A.; et al. Effectiveness of Wolbachia-infected mosquito deployments in reducing the incidence of dengue and chikungunya in Niterói, Brazil: A quasi-experimental study. PLoS Negl. Trop. Dis. 2021, 15, e0009556. [Google Scholar] [CrossRef]
- Garcia, G.A.; Sylvestre, G.; Aguiar, R.; da Costa, G.B.; Martins, A.J.; Lima, J.B.P.; Petersen, M.T.; Lourenço-de-Oliveira, R.; Shadbolt, M.F.; Rašić, G.; et al. Matching the genetics of released and local Aedes aegypti populations is critical to assure Wolbachia invasion. PLoS Negl. Trop. Dis. 2019, 13, e0007023. [Google Scholar] [CrossRef]
- Nazni, W.A.; Hoffmann, A.A.; NoorAfizah, A.; Cheong, Y.L.; Mancini, M.V.; Golding, N.; Kamarul, G.M.R.; Arif, M.A.K.; Thohir, H.; NurSyamimi, H.; et al. Establishment of Wolbachia strain wAlbB in Malaysian populations of Aedes aegypti for dengue control. Curr. Biol. 2019, 29, 4241–4248.e5. [Google Scholar] [CrossRef]
- Ryan, P.A.; Turley, A.P.; Wilson, G.; Hurst, T.P.; Retzki, K.; Brown-Kenyon, J.; Hodgson, L.; Kenny, N.; Cook, H.; Montgomery, B.L.; et al. Establishment of wMel Wolbachia in Aedes aegypti mosquitoes and reduction of local dengue transmission in Cairns and surrounding locations in northern Queensland, Australia. Gates Open Res. 2020, 3, 1547. [Google Scholar] [CrossRef]
- Caragata, E.P.; Dutra, H.L.C.; Sucupira, P.H.F.; Ferreira, A.G.A.; Luciano, A.; Moreira, L.A. Wolbachia as translational science: Controlling mosquito-borne pathogens. Trends Parasitol. 2021, 37, 1050–1066. [Google Scholar] [CrossRef]
- Yen, J.H.; Barr, A.R. New hypothesis of te cause of cytoplasmic incompatibility in Culex pipiens L. Nature 1971, 232, 657–658. [Google Scholar] [CrossRef]
- Atyame, C.M.; Pasteur, N.; Dumas, E.; Tortosa, P.; Tantely, M.L.; Pocquet, N.; Licciardi, S.; Bheecarry, A.; Zumbo, B.; Weill, M.; et al. Cytoplasmic incompatibility as a means of controlling Culex pipiens quinquefasciatus mosquito in the islands of the south-western indian ocean. PLoS Negl. Trop. Dis. 2011, 5, e1440. [Google Scholar] [CrossRef]
- Atyame, C.M.; Cattel, J.; Lebon, C.; Flores, O.; Dehecq, J.S.; Weill, M.; Gouagna, L.C.; Tortosa, P. Wolbachia-based population control strategy targeting Culex quinquefasciatus mosquitoes proves effcient under semi-feld conditions. PLoS ONE 2015, 10, e0119288. [Google Scholar] [CrossRef]
- da Moura, A.J.F.; Valadas, V.; Leal, S.d.V.; Sabino, E.M.; Sousa, C.A.; Pinto, J. Screening of natural Wolbachia infection in mosquitoes (Diptera: Culicidae) fro the Cape Verde Islands. Parasites Vectors 2023, 16, 142. [Google Scholar] [CrossRef]
- Glaser, R.L.; Meola, M.A. The native Wolbachia endosymbionts of Drosophila melanogaster and Culex quinquefasciatus increase host resistance to West Nile virus infection. PLoS ONE 2010, 5, e11977. [Google Scholar] [CrossRef]
- Hughes, G.L.; Koga, R.; Xue, P.; Fukatsu, T.; Rasgon, J.L. Wolbachia infections are virulent and inhibit the human malaria parasite Plasmodium falciparum in Anopheles gambiae. PLoS Pathog. 2011, 7, e1002043. [Google Scholar] [CrossRef]
- Bian, G.; Joshi, D.; Dong, Y.; Lu, P.; Zhou, G.; Pan, X.; Dimopoulos, G.; Xi, Z. Wolbachia invades Anopheles stephensi populations and induces refractoriness to Plasmodium infection. Science 2013, 340, 748–751. [Google Scholar] [CrossRef]
- Graham, R.I.; Grzywacz, D.; Mushobozi, W.L.; Wilson, K. Wolbachia in a major African crop pest increases susceptibility to viral disease rather than protects. Ecol. Lett. 2012, 15, 993–1000. [Google Scholar] [CrossRef]
- Hughes, G.L.; Vega-Rodriguez, J.; Xue, P.; Rasgon, J.L. Wolbachia strain wAlbB enhances infection by the rodent malaria parasite Plasmodium berghei in Anopheles gambiae mosquitoes. Appl. Env. Microbiol. 2012, 78, 1491–1495. [Google Scholar] [CrossRef]
- Murdock, C.C.; Blanford, S.; Hughes, G.L.; Rasgon, J.L.; Thomas, M.B. Temperature alters malaria transmission blocking by Wolbachia. Sci. Rep. 2013, 4, 3932. [Google Scholar] [CrossRef]
- Baton, L.A.; Pacidônio, E.C.; Gonçalves, D.D.S.; Moreira, L.A. wFlu: Characterization and evaluation of a native Wolbachia from the mosquito Aedes fluviatilis as a potential vector control agent. PLoS ONE 2013, 8, e59619. [Google Scholar] [CrossRef]
- Zélé, F.; Nicot, A.; Berthomieu, A.; Weill, M.; Duron, O.; Rivero, A. Wolbachia increases susceptibility to Plasmodium infection in a natural system. Proc. Biol. Sci. 2013, 281, 20132837. [Google Scholar] [CrossRef]
- Hussain, M.; Lu, G.; Torres, S.; Edmonds, J.H.; Kay, B.H.; Khromykh, A.A.; Asgari, S. Effect of Wolbachia on replication of West Nile virus in a mosquito cell line and adult mosquitoes. J. Virol. 2013, 87, 851–858. [Google Scholar] [CrossRef]
- Dodson, B.L.; Hughes, G.L.; Paul, O.; Matacchiero, A.C.; Kramer, L.D.; Rasgon, J.L. Wolbachia Enhances West Nile Virus (WNV) Infection in the Mosquito Culex tarsalis. PLoS Negl. Trop. Dis. 2014, 8, e2965. [Google Scholar] [CrossRef]
- Hughes, G.L.; Rivero, A.; Rasgon, J.L. Wolbachia Can Enhance Plasmodium Infection in Mosquitoes: Implications for Malaria Control? PLoS Pathog. 2012, 10, e1004182. [Google Scholar] [CrossRef]
- Amuzu, H.E.; Tsyganov, K.; Koh, C.; Herbert, R.I.; Powell, D.R.; McGraw, E.A. Wolbachia enhances insect-specific flavivirus infection in Aedes aegypti mosquitoes. Ecol. Evol. 2018, 8, 5441–5454. [Google Scholar] [CrossRef]
- Moretti, R.; Calvitti, M. Issues with combining incompatible and sterile insect techniques. Nature 2021, 590, E1–E2. [Google Scholar] [CrossRef]
- Kittayapong, P.; Ninphanomchai, S.; Limohpasmanee, W.; Chansang, C.; Chansang, U.; Mongkalangoon, P. Combined sterile insect technique and incompatible insect technique: The first proof-of-concept to suppress Aedes aegypti vector populations in semi-rural settings in Thailand. PLoS Negl. Trop. Dis. 2019, 13, e0007771. [Google Scholar] [CrossRef]
- Ong, J.; Ho, S.H.; Soh, S.X.H.; Wong, Y.; Ng, Y.; Vasquez, K.; Lai, Y.L.; Setoh, Y.X.; Chong, C.S.; Lee, V.; et al. Assessing the efficacy of male Wolbachia-infected mosquito deployments to reduce dengue incidence in Singapore: Study protocol for a cluster-randomized controlled trial. Trials 2022, 23, 1023. [Google Scholar] [CrossRef]
- Vorsino, A.E.; Xi, Z. A mass rearing cost calculator for the control of Culex quinquefasciatus in Hawai’i using the incompatible insect technique. Parasit Vectors 2022, 15, 453. [Google Scholar] [CrossRef]
- Martín-Park, A.; Che-Mendoza, A.; Contreras-Perera, Y.; Pérez-Carrillo, S.; Puerta-Guardo, H.; Villegas-Chim, J.; Guillermo-May, G.; Medina-Barreiro, A.; Delfín-González, H.; Méndez-Vales, R.; et al. Pilot trial using mass field-releases of sterile males produced with the incompatible and sterile insect techniques as part of integrated Aedes aegypti control in Mexico. PLoS Negl. Trop. Dis. 2022, 16, e0010324. [Google Scholar] [CrossRef]
- Thomas, D.; Donnelly, C.; Wood, R.; Alphey, L. Insect population control using a dominant, repressible, lethal genetic system. Science 2000, 287, 2474–2476. [Google Scholar] [CrossRef]
- Alphey, L.A. Re-engineering the sterile insect technique. Insect Biochem. Mol. Biol. 2002, 32, 1243–1247. [Google Scholar] [CrossRef]
- Zhao, Y.; Schetelig, M.F.; Handler, A.M. Genetic breakdown of a Tet-off conditional lethality system for insect population control. Nat. Commun. 2020, 11, 3095. [Google Scholar] [CrossRef]
- Qsim, M.; Ashfaq, U.A.; Yousaf, M.Z.; Masoud, M.S.; Rasul, I.; Noor, N.; Hussain, A. Genetically Modified Aedes aegypti to Control Dengue: A Review. Crit. Rev. Eukaryot. Gene Expr. 2017, 27, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Viktor, A.G. Genetic Methods of Insect Control: History and Current State. Russ. J. Biol. Invasions 2021, 12, 167–175. [Google Scholar] [CrossRef]
- Wise de Valdez, M.R.; Nimmo, D.; Betz, J.; Gong, H.F.; James, A.A.; Alphey, L.; Black, W.C., 4th. Genetic elimination of dengue vector mosquitoes. Proc. Natl. Acad. Sci. USA 2011, 108, 4772–4775. [Google Scholar] [CrossRef]
- Wilke, A.B.; Marrelli, M.T. Genetic control of mosquitoes: Population suppression strategies. Rev. Inst. Med. Trop. São Paulo 2012, 54, 287–292. [Google Scholar] [CrossRef]
- Seirin Lee, S.; Baker, R.E.; Gaffney, E.A.; White, S.M. Modelling Aedes aegypti mosquito control via transgenic and sterile insect techniques: Endemics and emerging outbreaks. J. Theor. Biol. 2013, 331, 78–90. [Google Scholar] [CrossRef]
- Carvalho, D.O.; Nimmo, D.; Naish, N.; McKemey, A.R.; Gray, P.; Wilke, A.B.; Marrelli, M.T.; Virginio, J.F.; Alphey, L.; Capurro, M.L. Mass production of genetically modified Aedes aegypti for field releases in Brazil. J. Vis. Exp. 2014, 4, e3579. [Google Scholar] [CrossRef]
- Leftwich, P.T.; Bolton, M.; Chapman, T. Evolutionary biology and genetic techniques for insect control. Evol. Appl. 2016, 9, 212–230. [Google Scholar] [CrossRef]
- Mansor, S.M.; Haninah, U.A.; Lacroix, R.; Angamuthu, C.; Ravindran, T.; Seshadri, V.S.; Sekaran, D.S.; Lee, H.L.; Murad, S.; Nam, W.S.; et al. Similar vertical transmission rates of dengue and chikungunya viruses in a transgenic and a non-transformed Aedes aegypti (L.) laboratory strain. Trop. Biomed. 2016, 33, 120–134. [Google Scholar]
- Massonnet-Bruneel, B.; Corre-Catelin, N.; Lacroix, R.; Lees, R.S.; Hoang, K.P.; Nimmo, D.; Alphey, L.; Reiter, P. Fitness of transgenic mosquito Aedes aegypti males carryning a dominant lethal genetic system. PLoS ONE 2013, 8, e62711. [Google Scholar] [CrossRef]
- Harris, A.F.; McKemey, A.R.; Nimmo, D.; Curtis, Z.; Black, I.; Morgan, S.A.; Oviedo, M.N.; Lacroix, R.; Naish, N.; Morrison, N.I.; et al. Successful suppression of a field mosquito population by sustained release of engineered male mosquitoes. Nat. Biotech. 2012, 30, 828–830. [Google Scholar] [CrossRef]
- Harris, A.F.; Nimmo, D.; McKemey, A.R.; Kelly, N.; Scaife, S.; Donnelly, C.A.; Beech, C.; Petrie, W.D.; Alphey, L. Field performance of engineered male mosquitoes. Nat. Biotech. 2011, 29, 1034–1037. [Google Scholar] [CrossRef]
- Carvalho, D.O.; McKemey, A.R.; Garziera, L.; Lacroix, R.; Donnelly, C.A.; Alphey, L.; Malavasi, A.; Margareth Capurro, M.L. Suppression of a field population of Aedes aegypti in Brazil by sustained release of transgenic male mosquitoes. PLoS Negl. Trop. Dis. 2015, 9, e0003864. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.R.; Kotsakiozi, P.; Costa-da-Silva, A.L.; Ioshino, R.S.; Garziera, L.; Pedrosa, M.C.; Malavasi, A.; Virginio, J.F.; Capurro, M.L.; Powell, J.R. Transgenic Aeded aegypti mosqutoes transfer genes into natrral population. Sci. Rep. 2019, 9, 13047. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.B.; Dasgupta, S.K.; Gorman, K.; Pickl-Herk, A.; Puinean, M.; McKemey, A.; Char, B.; Zehr, U.B.; Barwale, S.R. Elimination of a closed population of the yellow fever mosquito, Aedes aegypti, through releases of self-limiting male mosquitoes. PLoS Negl. Trop. Dis. 2022, 16, e0010315. [Google Scholar] [CrossRef]
- Servick, K. GM mosquito study draws fire. Science 2019, 365, 1234. [Google Scholar] [CrossRef] [PubMed]
- Oxitec responds to article entitled ‘Transgenic Aedes aegypti mosquitoes transfers genes into natural populations’. Available online: https://www.oxitec.com/en/news/oxitec-response-scientific-reports-article (accessed on 15 February 2024).
- Jiang, F.; Doudna, J.A. CRISPR–Cas9 Structures and Mechanisms. Annu. Rev. Biophys. 2017, 46, 505–529. [Google Scholar] [CrossRef] [PubMed]
- Hille, F.; Charpentier, E. CRISPR-Cas: Biology, mechanisms and relevance. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150496. [Google Scholar] [CrossRef]
- Hall, A.B.; Basu, S.; Jiang, X.; Qi, Y.; Timoshevskiy, V.A.; Biedler, J.K.; Sharakhova, M.V.; Elahi, R.; Anderson, M.A.; Chen, X.G.; et al. A male-determining factor in the mosquito Aedes aegypti. Science 2015, 348, 1268–1270. [Google Scholar] [CrossRef]
- Hammond, A.; Galizi, R.; Kyrou, K.; Simoni, A.; Siniscalchi, C.; Katsanos, D.; Gribble, M.; Baker, D.; Marois, E.; Russell, S.; et al. A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nat. Biotechnol. 2016, 34, 78–83. [Google Scholar] [CrossRef]
- Galizi, R.; Doyle, L.A.; Menichelli, M.; Bernardini, F.; Deredec, A.; Burt, A.; Stoddard, B.L.; Windbichler, N.; Crisanti, A. A synthetic sex ratio distortion system for the control of the human malaria mosquito. Nat. Commun. 2014, 5, 3977. [Google Scholar] [CrossRef]
- Galizi, R.; Hammond, K.K.; Taxiarchi, C.; Bernardini, F.; O’Loughlin, S.M.; Papathanos, P.-A.; Nolan, T.; Windbichler, N.; Crisanti, A. 2018. A CRISPR-Cas9 sex-ratio distortion system for genetic control. Sci. Rep. 2018, 6, 31139. [Google Scholar] [CrossRef] [PubMed]
- Kyrou, K.; Hammond, A.M.; Galizi, R.; Kranjc, N.; Burt, A.; Beaghton, A.K.; Nolan, T.; Crisanti, A. A CRISPR-Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nat. Biotechnol. 2018, 36, 1062–1066. [Google Scholar] [CrossRef]
- Connolly, J.B.; Romeis, J.; Devos, Y.; Glandorf, D.C.M.; Turner, G.; Coulibaly, M.B. Gene drive in species complexes: Defining target organisms. Trends Biotechnol. 2023, 41, 154–164. [Google Scholar] [CrossRef]
- Bottino-Rojas, V.; James, A.A. Use of Insect Promoters in Genetic Engineering to Control Mosquito-Borne Diseases. Biomolecules 2022, 13, 16. [Google Scholar] [CrossRef]
- Nourani, L.; Mehrizi, A.A.; Pirahmadi, S.; Pourhashem, Z.; Asadollahi, E.; Jahangiri, B. CRISPR/Cas advancements for genome editing, diagnosis, therapeutics, and vaccine development for Plasmodium parasites, and genetic engineering of Anopheles mosquito vector. Infect. Genet Evol. 2023, 109, 105419. [Google Scholar] [CrossRef]
- James, S.L.; Quemada, H.; Benedict, M.Q.; Dass, B. Requirements for market entry of gene drive-modified mosquitoes for control of vector-borne diseases: Analogies to other biologic and biotechnology products. Front. Bioeng. Biotechnol. 2023, 11, 1205865. [Google Scholar] [CrossRef]
- D’Amato, R.; Taxiarchi, C.; Galardini, M.; Trusso, A.; Minuz, R.L.; Grilli, S.; Somerville, A.G.T.; Shittu, D.; Khalil, A.S.; Galizi, R.; et al. Anti-CRISPR Anopheles mosquitoes inhibit gene drive spread under challenging behavioural conditions in large cages. Nat. Commun. 2024, 15, 952. [Google Scholar] [CrossRef]
- Jones, S. Modidying mosquities: Gene-drive technology in mosquitoes is maturing, but there are questions to answer before the malaria fighting technique can be set loose. Nature 2023, 618, S29–S31. [Google Scholar] [CrossRef]
- James, S.; Santos, M. The promise and challenge of genetic biocontrol approaches for malaria elimination. Trop. Med. Infect. Dis. 2023, 8, 201. [Google Scholar] [CrossRef] [PubMed]
- James, S.L.; Dass, B.; Quemada, H. Regulatory and policy considerations fro the implementation of gene drive mosquitoes to prevent malaria transmission. Transgenic Res. 2023, 32, 17–32. [Google Scholar] [CrossRef]
- Dong, Y.; Simões, M.L.; Marois, E.; Dimopoulos, G. CRISPR/Cas9 -mediated gene knockout of Anopheles gambiae FREP1 suppresses malaria parasite infection. PLoS Pathog. 2018, 14, e1006898. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Schleicher, T.R.; Dong, Y.; Park, H.B.; Lan, J.; Cresswell, P.; Crawford, J.; Dimopoulos, G.; Fikrig, E. Disruption of mosGILT in Anopheles gambiae impairs ovarian development and Plasmodium infection. J. Exp. Med. 2020, 217, e20190682. [Google Scholar] [CrossRef]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L. C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef]
- Smargon, A.A.; Cox, D.B.; Pyzocha, N.K.; Zheng, K.; Slaymaker, I.M.; Gootenberg, J.S.; Abudayyeh, O.A.; Essletzbichler, P.; Shmakov, S.; Makarova, K.S.; et al. Cas13b is a type VI-B CRISPRassociated RNA-guided RNase differentially regulated by accessory proteins Csx27 and Csx 28. Mol. Cell 2017, 65, 618–630. [Google Scholar] [CrossRef]
- Koonin, E.V.; Makarova, K.S.; Zhang, F. Diversity, classification and evolution of CRISPR-Cas systems. Curr. Opin. Microbiol. 2017, 37, 67–78. [Google Scholar] [CrossRef]
- Huang, Z.; Fang, J.; Zhou, M.; Gong, Z.; Xiang, T. CRISPR-Cas13: A new technology for the rapid detection of pathogenic microorganisms. Front. Microbiol. 2022, 13, 1011399. [Google Scholar] [CrossRef]
- Morelli, K.H.; Wu, Q.; Gosztyla, M.L.; Liu, H.; Yao, M.; Zhang, C.; Chen, J.; Marina, R.J.; Lee, K.; Jones, K.L.; et al. An RNA-targeting CRISPR-Cas13d system alleviates disease-related phenotypes in Huntington’s disease models. Nat. Neurosci. 2023, 26, 27–38. [Google Scholar] [CrossRef]
- Huynh, N.; Depner, N.; Larson, R.; King-Jones, K. A versatile toolkit for CRISPR-Cas13-based RNA manipulation in Drosophila. Genome Biol. 2020, 21, 279. [Google Scholar] [CrossRef]
- Tng, P.Y.L.; Carabajal Paladino, L.; Verkuijl, S.A.N.; Purcell, J.; Merits, A.; Leftwich, P.T.; Fragkoudis, R.; Noad, R.; Alphey, L. Cas13b-dependent and Cas13b-independent RNA knockdown of viral sequences in mosquito cells following guide RNA expression. Commun. Biol. 2020, 3, 413. [Google Scholar] [CrossRef]
- Chen, P.; Chen, M.; Chen, Y.; Jing, X.; Zhang, N.; Zhou, X.; Li, X.; Long, G.; Hao, P. Targeted inhibition of Zika virus infection in human cells by CRISPR-Cas13b. Virus Research. 2022, 31, 198707. [Google Scholar] [CrossRef]
- Dalla Benetta, E.; López-Denman, A.J.; Li, H.H.; Masri, R.A.; Brogan, D.J.; Bui, M.; Yang, T.; Li, M.; Dunn, M.; Klein, M.J.; et al. Engineered Antiviral Sensor Targets Infected Mosquitoes. CRISPR J. 2023, 6, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Panneerselvam, S.; Mishra, S.; Berry, C.; Crickmore, N.; Bonning, B.C. BPPRC database: A web-based tool to access and analyse bacterial pesticidal proteins. Database 2022, 2022, baac022. [Google Scholar] [CrossRef] [PubMed]
- Federici, B.A.; Park, H.-W.; Bideshi, D.K. Overview of the basic biology of Bacillus thuringiensis with emphasis on genetic engineering of bacterial larvicides for mosquito control. Open Toxinol. J. 2010, 3, 83–100. [Google Scholar] [CrossRef]
- Kellen, W.R.; Clark, T.B.; Lindegren, J.E.; Ho, B.C.; Rogoff, M.H.; Singer, S. Bacillus sphaericus Neide as a pathogen of mosquitoes. J. Invertebr. Pathol. 1965, 7, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Park, H.W.; Bideshi, D.K.; Federici, B.A. Properties and applied use of the mosquitocidal bacterium, Bacillus sphaericus. J. Asia Pac. Entomol. 2010, 13, 159–168. [Google Scholar] [CrossRef]
- Berry, C. The bacterium, Lysinibacillus sphaericus, as an insect pathogen. J. Invertebr. Pathol. 2012, 109, 1–10. [Google Scholar] [CrossRef]
- Weiser, J. A mosquito-virulent Bacillus sphaericus in adult Simulium damnosum from northern Nigeria. Zentralbl. Mikrobiol. 1984, 139, 57–60. [Google Scholar] [CrossRef]
- Colletier, J.P.; Sawaya, M.R.; Gingery, M.; Rodriguez, J.A.; Cascio, D.; Brewster, A.S.; Michels-Clark, T.; Hice, R.H.; Coquelle, N.; Boutet, S.; et al. De novo phasing with X-ray laser reveals mosquito larvicide BinAB structure. Nature 2016, 539, 43–47. [Google Scholar] [CrossRef]
- Degiacomi, M.T.; Iacovache, I.; Pernot, L.; Chami, M.; Kudryashev, M.; Stahlberg, H.; van der Goot, F.G.; Dal Peraro, M. Molecular assembly of the aerolysin pore reveals a swirling membrane-insertion mechanism. Nat. Chem. Biol. 2013, 9, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Williamson, L.J.; Galchenkova, M.; Best, H.L.; Bean, R.J.; Munke, A.; Awel, S.; Pena, G.; Knoska, J.; Schubert, R.; Dörner, K.; et al. Structure of the Lysinibacillus sphaericus Tpp49Aa1 pesticidal protein elucidated from natural crystals using MHz-SFX. Proc. Natl. Acad. Sci. USA 2023, 120, e2203241120. [Google Scholar] [CrossRef]
- Opota, O.; Charles, J.F.; Warot, S.; Pauron, D.; Darboux, I. Identification and characterization of the receptor for Bacillus sphaericus binary toxin in the malaria vector mosquito. Anopheles Gambiae. Comp. Biochem. Physiol.-B Biochem. Mol. Biol. 2008, 149, 419–427. [Google Scholar] [CrossRef]
- Silva-Filha, M.H.N.L.; Romão, T.P.; Rezende, T.M.T.; Carvalho, K.d.S.; Gouveia de Menezes, H.S.; Alexandre do Nascimento, N.; Soberón, M.; Bravo, A. Bacterial Toxins Active against Mosquitoes: Mode of Action and Resistance. Toxins 2021, 13, 523. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Kumar, V. Mosquito-larvicidal binary (BinA/B) proteins for mosquito control programs–Advancements, challenges, and possibilities. Curr. Res. Insect Sci. 2022, 2, 10028. [Google Scholar] [CrossRef]
- Ferriera, L.M.; Romao, T.P.; Pompillo de-Melo-Neto, O.; Silva-Filha, M.H.N.L. The orthologue of the Cpm1/Cqm1 receptor in Aedes aegypti is expressed as a midgut GPI-anchored a-glucosidase, which does not bind to the insecticidal binary toxin. Insect. Biochem. Mol. Biol. 2010, 40, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Charles, J.F. Ultrastructural midgut events in Culicidae larvae fed with Bacillus sphaericus 2297 spore/crystal complex. Ann. Inst. Pasteur. Microbiol. 1987, 138, 471–484. [Google Scholar] [CrossRef]
- de Melo, J.V.; Vasconcelos, R.H.; Furtado, A.F.; Peixoto, C.A.; Silva-Filha, M.H. Ultrastructural analysis of midgut cells from Culex quinquefasciatus (Diptera: Culicidae) larvae resistant to Bacillus sphaericus. Micron 2008, 39, 1342–1350. [Google Scholar] [CrossRef]
- Silva Filha, M.H.N.L.; Peixoto, C.A. Immunocytochemical localization of the Bacillus sphaericus toxin components in Culex quinquefasciatus (Diptera: Culicidae) larvae midgut. Pest. Biochem. Physiol. 2003, 77, 138–146. [Google Scholar] [CrossRef]
- Singh, G.J.; Gill, S.S. An electron microscope study of the toxic action of Bacillus sphaericus in Culex quinquefasciatus larvae. J. Invertebr. Pathol. 1988, 52, 237–247. [Google Scholar] [CrossRef]
- Tangsongcharoen, C.; Chomanee, N.; Promdonkoy, B.; Boonserm, P. Lysinibacillus sphaericus binary toxin induces apoptosis in susceptible Culex quinquefasciatus larvae. J. Invertebr. Pathol. 2015, 128, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Lekakarn, H.; Promdonkoy, B.; Boonserm, P. Interaction of Lysinibacillus sphaericus binary toxin with mosquito larval gut cells: Binding and internalization. J. Invertebr. Pathol. 2015, 132, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Tangsongcharoen, C.; Jupatanakul, N.; Promdonkoy, B.; Dimopoulos, G.; Boonserm, P. Molecular analysis of Culex quinquefasciatus larvae responses to Lysinibacillus sphaericus Bin toxin. PLoS ONE 2017, 12, e0175473. [Google Scholar] [CrossRef] [PubMed]
- Ishiwata, S. On a new type of severe flacherie (sotto disease). Dainihon Sansi Kaiho. 1901, 114, 1–5. (In Japanese) [Google Scholar]
- Berliner, E. Ueber die schlaffsucht der Ephestia kuhniella und Bac. thuringiensis n. sp. Z Angew. Entomol. 1915, 2, 21–56. [Google Scholar] [CrossRef]
- Krieg, A.; Huger, A.M.; Langenbrunch, G.A.; Schnetter, W. Bacillus thuringiensis var. tenebrionis: Ein neuer, gegenüber Larven von Coleopteren wirksamer Pathotyp. Z Angew. Entomol. 1983, 96, 500–508. [Google Scholar] [CrossRef]
- Meadows, M.P. Bacillus thuringiensis in the environment: Ecology and risk assessment. In Bacillus thuringiensis, an Environmental Biopesticide: Theory and Practice; Entwistle, P.F., Cory, J.S., Bailey, M.J., Higgs, S., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 1993; pp. 193–220. [Google Scholar]
- Wang, K.; Shu, C.; Bravo, A.; Soberón, M.; Zhang, H.; Crickmore, N.; Zhang, J. Development of an Online Genome Sequence Comparison Resource for Bacillus cereus sensu lato Strains Using the Efficient Composition Vector Method. Toxins 2023, 15, 393. [Google Scholar] [CrossRef]
- Kumar, P.; Kamle, M.; Borah, R.; Mahato, D.K.; Sharma, B. Bacillus thuringiensis as microbial biopesticides: Uses and application for sustainable agriculture. Egypt. J. Biol. Pest Control. 2019, 31, 95. [Google Scholar] [CrossRef]
- Yamamoto, T. Engineering of Bacillus thuringiensis insecticidal protens. J. Pestic. Sci. 2022, 47, 47–58. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Resig, D.D. Management of insect pests with Bt crops in the United States. Annu. Rev. Entomol. 2023, 68, 31–49. [Google Scholar] [CrossRef]
- Goldberg, L.J.; Margalit, J. A bacterial spore demonstrating rapid larvicidal activity against Anopheles sergentii, Uranotaenia unguiculata, Culex univittatus, Aedes aegypti, and Culex pipiens. Mosq. News 1977, 37, 317–324. [Google Scholar]
- Lacey, L.A.; Undeen, A.H. Microbial control of blackflies and mosquitoes. Ann. Rev. Entomol. 1986, 31, 265–296. [Google Scholar] [CrossRef]
- Derua, Y.A.; Kweka, E.J.; Kisinza, W.N.; Githeko, A.K.; Mosha, F.W. Bacterial larvicides used for malaria vector control in sub-Saharan Africa: Review of their effectiveness and operational feasibility. Parasit Vectors 2019, 12, 426. [Google Scholar] [CrossRef] [PubMed]
- Bacillus thuringiensis israelensis (Bti) Larvicides|Mosquito Control. Available online: https://www.arbico-organics.com (accessed on 15 February 2024).
- Glare, T.R.; O’Callaghan, M. Env Health Impacts of Bti. 1998. Environmental and Health Impacts of Bacillus Thuringiensis Isr. Available online: https://www.smsl.co.nz (accessed on 15 February 2024).
- PAHO. Available online: https://www.paho.org/en/topics/onchocerciasis-river-blindness#:~:text=The%20disease%20is%20commonly%20known,introduced%20through%20the%20slave%20trade (accessed on 15 February 2024).
- World Health Organization (WHO). World Health Organization Fact Sheets: Onchocerciasis; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Robacker, D.C.; Martinez, A.J.; Garcia, J.A.; Diaz, M.; Romero, C. Toxicity of Bacillus thuringiensis to Mexican fruit fly (Diptera: Tephritidae). J. Econ. Entomol. 1996, 89, 104–110. [Google Scholar] [CrossRef]
- Vidal-Quist, J.C.; Castañera, P.; González-Cabrera, J. Cyt1Aa protein from Bacillus thuringiensis (Berliner) serovar israelensis is active against the Mediterranean fruit fly, Ceratitis capitata (Wiedemann). Pest Manag. Sci. 2010, 66, 949–955. [Google Scholar] [CrossRef]
- Porcar, M.; Grenier, A.M.; Federici, B.; Rahbé, Y. Effects of Bacillus thuringiensis δ-endotoxins on the Pea aphid (Acyrthosiphon pisum). Appl. Environ. Microbiol. 2009, 75, 4897–4900. [Google Scholar] [CrossRef]
- Walters, F.S.; English, L.H. Toxicity of Bacillus thuringiensis δ-endotoxins toward the potato aphid in an artificial diet bioassay. Entomol. Exp. Appl. 1995, 77, 211–216. [Google Scholar] [CrossRef]
- Harris, M.A.; Oetting, R.D.; Gardner, W.A. Use of entomopathogenic nematodes and a new monitoring technique for control of fungus gnats, Bradysia coprophila (Diptera: Sciaridae), in floriculture. Biol. Control 1995, 5, 412–418. [Google Scholar] [CrossRef]
- Taylor, M.D.; Willey, R.D.; Noblet, R. A 24-h potato-based toxicity test for evaluating Bacillus thuringiensis var. israelensis (H-14) against darkwinged fungus gnat Bradysia impatiens Johannsen (Diptera: Sciaridae) larvae. Int. J. Pest Manag. 2007, 53, 77–81. [Google Scholar] [CrossRef]
- Monnerat, R.; Martins, E.; Praça, L.; Dumas, V.; Berry, C. Activity of a Brazilian strain of Bacillus thuringiensis israelensis against the cotton boll weevil Anthonomus grandis Boheman (Coleoptera: Tenebrionidae). Neotrop. Entomol. 2012, 41, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Federici, B.A.; Bauer, L.S. Cyt1Aa protein of Bacillus thuringiensis is toxic to the cottonwood leaf beetle, Chrysomela scripta, and suppresses high levels of resistance to Cry3Aa. Appl. Environ. Microbiol. 1998, 64, 4368–4371. [Google Scholar] [CrossRef] [PubMed]
- Horak, P.; Weiser, J.; Mikes, L.; Kolarova, L. The effect of Bacillus thuringiensis M-exotoxin on trematode cercariae. J. Invertebr. Pathol. 1996, 68, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.A.; Salem, H.H.; Wang, X.M.; Huang, T.H.; Xie, Q.D.; Zhang, Z.Y. Effect of Bacillus thuringiensis var. israelensis endotoxin on the intermediate snail host of Schistosoma japonicum. Curr. Res. Baacteriol. 2010, 3, 37–41. [Google Scholar] [CrossRef]
- Sharma, R.D. Bacillus thuringiensis: A biocontrol agent of Meloidogyne incognita on barley. Nematol. Bras. 1994, 18, 79–84. [Google Scholar]
- Diaz-Mendoza, M.; Bideshi, D.K.; Federici, B.A. A 54 -kilodalton proteins encoded by pBtoxis is required for parasporal body structural integrity in Bacillus thuringiensis subsp. israelensis. J. Bacteriol. 2012, 194, 1562–1571. [Google Scholar] [CrossRef]
- Berry, C.; O’Neil, S.; Ben-Dov, E.; Jones, A.F.; Murphy, L.; Quail, M.A.; Holden, M.T.; Harris, D.; Zaritsky, A.; Parkhill, J. Complete sequence and organization of pBtoxis, the toxin-coding plasmid of Bacillus thuringiensis subsp. israelensis. Appl. Environ. Microbiol. 2002, 68, 5082–5095. [Google Scholar] [CrossRef]
- Tetreau, G.; Sawaya, M.R.; De Zitter, E.; Andreeva, E.A.; Banneville, A.S.; Schibrowsky, N.A.; Coquelle, N.; Brewster, A.S.; Grünbein, M.L.; Kovacs, G.N.; et al. De novo determination of mosquitocidal Cry11Aa and Cry11Ba structures from naturally-occurring nanocrystals. Nat Commun. 2022, 13, 4376. [Google Scholar] [CrossRef]
- Boonserm, P.; Davis, P.; Ellar, D.J.; Li, J. Crystal structure of the mosquito-larvicidal toxin Cry4Ba and its biological implications. J. Mol. Biol. 2005, 348, 363–382. [Google Scholar] [CrossRef]
- Boonserm, P.; Mo, M.; Angsuthanasombat, C.; Lescar, J. Structure of the functional form of the mosquito larvicidal Cry4Aa toxin from Bacillus thuringiensis at a 2.8-angstrom resolution. J. Bacteriol. 2006, 188, 3391–3401. [Google Scholar] [CrossRef]
- Tetreau, G.; Banneville, A.S.; Andreeva, E.A.; Brewster, A.S.; Hunter, M.S.; Sierra, R.G.; Teulon, J.M.; Young, I.D.; Burke, N.; Grünewald, T.A.; et al. Serial femtosecond crystallography on in vivo-grown crystals drives elucidation of mosquitocidal Cyt1Aa1 bioactivation cascade. Nat. Commun. 2020, 11, 1153. [Google Scholar] [CrossRef]
- Ben-Dov, E. Bacillus thuringiensis subsp. israelensis and its dipteran-specific toxins. Toxins 2014, 6, 1222–1243. [Google Scholar] [CrossRef]
- Palma, L.; Munoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis toxins: And overview of their biocidal actiity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef]
- Pinos, D.; Andrés-Garrido, A.; Ferré, J.; Hernández-Martínez, P. Response mechanisms of invertebrates to Bacillus thuringiensis and its pesticidal proteins. Microbiol Mol Biol Rev. 2021, 85, 10–11228. [Google Scholar] [CrossRef]
- Heckel, D.G. How do toxins from Bacillus thuringiensis kill insects? And evolutionary perspective. Arch. Insect. Biochem. Physiol. 2022, 4, e2167. [Google Scholar] [CrossRef] [PubMed]
- Stalinski, R.; Laporte, F.; Tetreau, G.; Depres, L. Receptors are affected by selection with each Bacillys thuringiensis Cry toxin but not with the Bti ull mixture in Aedes aegypti. Infect. Genet. Evol. 2016, 44, 218–227. [Google Scholar] [CrossRef]
- Wu, D.; Federici, B.A. A 20-kilodalton protein preserves cell viability and promotes CytA crystal formation in Bacillus thu-ringiensis. J. Bacteriol. 1993, 175, 5276–5280. [Google Scholar] [CrossRef] [PubMed]
- Manasherob, R.; Zaritsk, A.; Ben-Dov, E.; Saxena, D.; Barak, Z.; Einav, M. Effect of accessory proteins P19 and P20 on cy-tolytic activity of Cyt1Aa1 from Bacillus thuringiensis subsp. israelensis in Escherichia coli. Curr. Microbiol. 2001, 43, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Albeck, S.; Ben-Dov, E.; Cahan, R.; Firer, M.; Zaritsky, A.; Dym, O. Cyt1Aa toxin: Crystal structure reveals implications for its membrane-perforating function. J. Mol. Biol. 2011, 413, 804–814. [Google Scholar] [CrossRef]
- Guerchicoff, A.; Delécluse, A.; Rubinstein, C.P. The Bacillus thuringiensis cyt genes for hemolytic endotoxins constitute a gene family. Appl. Environ. Microbiol. 2001, 67, 1090–1096. [Google Scholar] [CrossRef]
- Anaya, P.; Onofre, J.; Torres-Quintero, M.C.; Sánchez, J.; Gill, S.S.; Bravo, A.; Soberón, M. Oligomerization is a key step for Bacillus thuringiensis Cyt1Aa insecticidal activity but not for toxicity against red blood cells. Insect. Biochem. Mol. Biol. 2020, 119, 103317. [Google Scholar] [CrossRef] [PubMed]
- Bideshi, D.K.; Park, H.-W.; Hice, R.D.; Wirth, M.C.; Federici, B.A. Highly effective broad spectrum chimeric larvicide that targets vector mosquitoes using a lipophilic protein. Sci. Rep. 2017, 7, 11282. [Google Scholar] [CrossRef] [PubMed]
- Georghiou, G.P.; Wirth, M.C. Influence of Exposure to Single versus Multiple Toxins of Bacillus thuringiensis Subsp. israelensis on Development of Resistance in the Mosquito Culex Quinquefasciatus (Diptera: Culicidae). Appl. Environ. Microbiol. 1997, 63, 1095–1101. [Google Scholar] [PubMed]
- Wirth, M.C.; Park, H.W.; Walton, W.E.; Federici, B.A. Cyt1A of Bacillus thuringiensis Delays Evolution of Resistance to Cry11A in the Mosquito Culex quinquefasciatus. Appl. Environ. Microbiol. 2005, 71, 185–189. [Google Scholar] [CrossRef]
- Federici, B.A.; Park, H.W.; Bideshi, D.K.; Wirth, M.C.; Johnson, J.J. Recombinant Bacteria for Mosquito Control. J. Exp. Biol. 2003, 206, 3877–3885. [Google Scholar] [CrossRef]
- Perez, C.; Fernandez, L.E.; Sun, J.G.; Folch, J.L.; Gill, S.S.; Soberon, M.; Bravo, A. Bacillus thuringiensis subsp israelensis Cyt1Aa syn-ergizes Cry11Aa toxin by functioning as a membrane-bound receptor. Proc. Natl. Acad. Sci. USA 2005, 102, 18303–18308. [Google Scholar] [CrossRef] [PubMed]
- Elleuch, J.; Jacca, S.; Darriet, F.; Chandre, F.; Tounsi, F.; Zghal, R.Z. Cry4Ba and Cyt1Aa proteins from Bacillus thuringiensis israelensis: Interactions and toxicity mechanism against Aedes aegypti. Toxicon 2015, 104, 83–90. [Google Scholar] [CrossRef]
- Wirth, M.C.; Federici, B.A.; Walton, W.E. Cyt1A from Bacillus thuringiensis synergizes activity of Bacillus sphaericus against Aedes aegypti (Diptera: Culicidae). Appl. Environ. Microbiol. 2000, 66, 1093–1097. [Google Scholar] [CrossRef]
- Wirth, M.C.; Walton, W.E.; Federici, B.A. Cyt1A from Bacillus thuringiensis restores toxicity of Bacillus sphaericus against resistant Culex quinquefasciatus (Diptera: Culicidae). J. Med. Entomol. 2000, 37, 401–407. [Google Scholar] [CrossRef]
- Nascimento, N.A.; Torres-Quintero, M.C.; Molina, S.L.; Pacheco, S.; Romão, T.P.; Pereira-Neves, A.; Soberón, M.; Bravo, A.; Silva-Filha, M.H. Functional Bacillus thuringiensis Cyt1Aa is necessary to synergize Lysinibacillus sphaericus binary toxin (Bin) against Bin-resistant and-refractory mosquito species. Appl. Environ. Microbiol. 2020, 86, e02770-19. [Google Scholar] [CrossRef]
- Knowles, B.H.; Ellar, D.J. Colloid-osmotic lysis is a general feature of the mechanism of action of Bacillus thuringiensis δ-endotoxins with different insect specificity. Biochim. Biophys. 1987, 924, 509–518. [Google Scholar] [CrossRef]
- Drobniewski, F.A.; Ellar, D.J. Investigation of the membrane lesion induced in vitro by two mosquitocidal δ-endotoxins of Bacillus thuringiensis. Curr. Microbiol. 1988, 16, 195–199. [Google Scholar] [CrossRef]
- Knowles, B.H.; Blatt, M.R.; Tester, M.; Horsnell, J.M.; Carroll, J.; Menestrina, G.; Ellar, D.J. A cytolytic δ-endotoxin from Bacillus thuringiensis var. israelensis forms cation-selective channels in planar lipid bilayers. FEBS Lett. 1989, 244, 259–262. [Google Scholar] [CrossRef]
- Butko, P. Cytolytic toxin Cyt1A and its mechanism of membrane damage: Data and hypotheses. Appl. Environ. Microbiol. 2003, 69, 2415–2422. [Google Scholar] [CrossRef] [PubMed]
- Soberon, M.; Lopez-Diaz, J.A.; Bravo, A. Cyt toxins produced by Bacillus thuringiensis: A protein fold conserved in several pathogenic microorganisms. Peptides 2013, 41, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Promdonkoy, B.; Ellar, D.J. Structure-function relationships of a membrane pore forming toxin revealed by reversion mutagenesis. Mol. Membr. Biol. 2005, 22, 327–337. [Google Scholar] [CrossRef]
- Carvalho, K.d.S.; Crespo, M.M.; Araújo, A.P.; da Silva, R.S.; de Melo-Santos, M.A.V.; de Oliveira, C.M.F.; Silva-Filha, M.H.N.L. Long-term exposure of Aedes aegypti to Bacillus thuringiensis svar. israelensis did not involve altered susceptibility to this microbial larvicide or to other control agents. Parasites Vectors 2018, 11, 673. [Google Scholar] [CrossRef]
- Park, H.W.; Bideshi, D.K.; Wirth, M.C.; Johnson, J.J.; Walton, W.E.; Federici, B.A. Recombinant larvicidal bacteria with markedly improved efficacy against culex vectors of west nile virus. Am. J. Trop. Med. Hyg. 2005, 72, 732–738. [Google Scholar] [CrossRef]
- Sakano, Y.; Park, H.-W.; Bideshi, D.K.; Ge, B.; Federici, B.A. Contributions of 5’-UTR and 3’-UTR cis elements to Cyt1Aa synthesis in Bacillus thuringiensis subsp. israelensis. J. Invertebr. Pathol. 2017, 149, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Agaisse, H.; Lereclus, D. STAB-SD: A Shine-Dalgarno sequence in the 5′ untranslated region is a determinant of mRNA stability. Mol. Microbiol. 1996, 20, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-W.; Ge, B.; Bauer, L.S.; Federici, B.A. Optimization of Cry3A yields in Bacillus thuringiensis by use of sporulation-dependent promoters in combination with the STAB-SD mRNA sequence. Appl. Environ. Microbiol. 1998, 64, 3932–3938. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-W.; Delecluse, A.; Federici, B.A. Construction and characterization of a recombinant Bacillus thuringiensis strain that produces Cry11B. J. Invertebr. Pathol. 2001, 78, 37–44. [Google Scholar] [CrossRef]
- Juárez-Hernández, E.O.; Casados-Vázquez, L.E.; del Rincón-Castro, M.C.; Salcedo-Hernández, R.; Bideshi, D.K.; Barboza-Corona, J.E. Bacillus thuringiensis subsp. israelensis producing endochitinase ChiA74Δsp inclusions and its improved activity against Aedes aegypti. J. Appl. Microbiol. 2015, 119, 1692–1699. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miranda, L.S.; Rudd, S.R.; Mena, O.; Hudspeth, P.E.; Barboza-Corona, J.E.; Park, H.-W.; Bideshi, D.K. The Perpetual Vector Mosquito Threat and Its Eco-Friendly Nemeses. Biology 2024, 13, 182. https://doi.org/10.3390/biology13030182
Miranda LS, Rudd SR, Mena O, Hudspeth PE, Barboza-Corona JE, Park H-W, Bideshi DK. The Perpetual Vector Mosquito Threat and Its Eco-Friendly Nemeses. Biology. 2024; 13(3):182. https://doi.org/10.3390/biology13030182
Chicago/Turabian StyleMiranda, Leticia Silva, Sarah Renee Rudd, Oscar Mena, Piper Eden Hudspeth, José E. Barboza-Corona, Hyun-Woo Park, and Dennis Ken Bideshi. 2024. "The Perpetual Vector Mosquito Threat and Its Eco-Friendly Nemeses" Biology 13, no. 3: 182. https://doi.org/10.3390/biology13030182
APA StyleMiranda, L. S., Rudd, S. R., Mena, O., Hudspeth, P. E., Barboza-Corona, J. E., Park, H.-W., & Bideshi, D. K. (2024). The Perpetual Vector Mosquito Threat and Its Eco-Friendly Nemeses. Biology, 13(3), 182. https://doi.org/10.3390/biology13030182