Macroalgae as a Source of Valuable Antimicrobial Compounds: Extraction and Applications

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract

1. Introduction
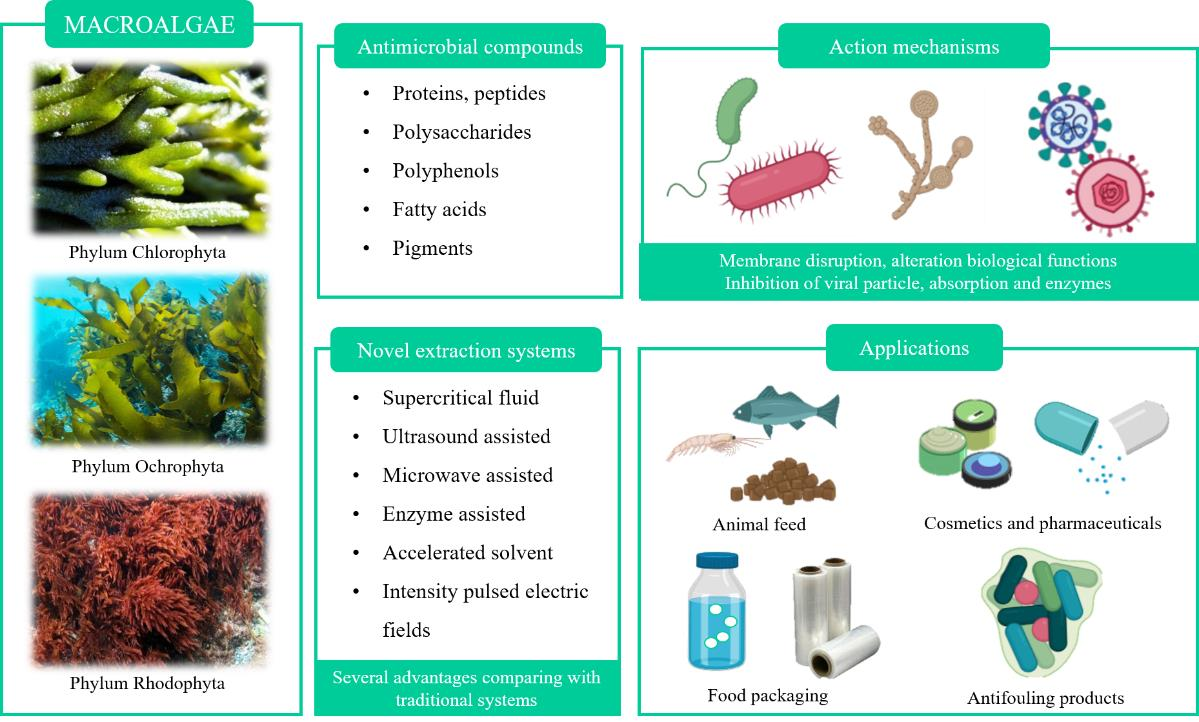
2. Macroalgae as a Promising Source of Valuable Antimicrobial Compounds
2.1. Protein and Peptides
2.2. Polysaccharides
2.3. Fatty Acids
2.4. Polyphenolic Compounds
2.5. Pigments
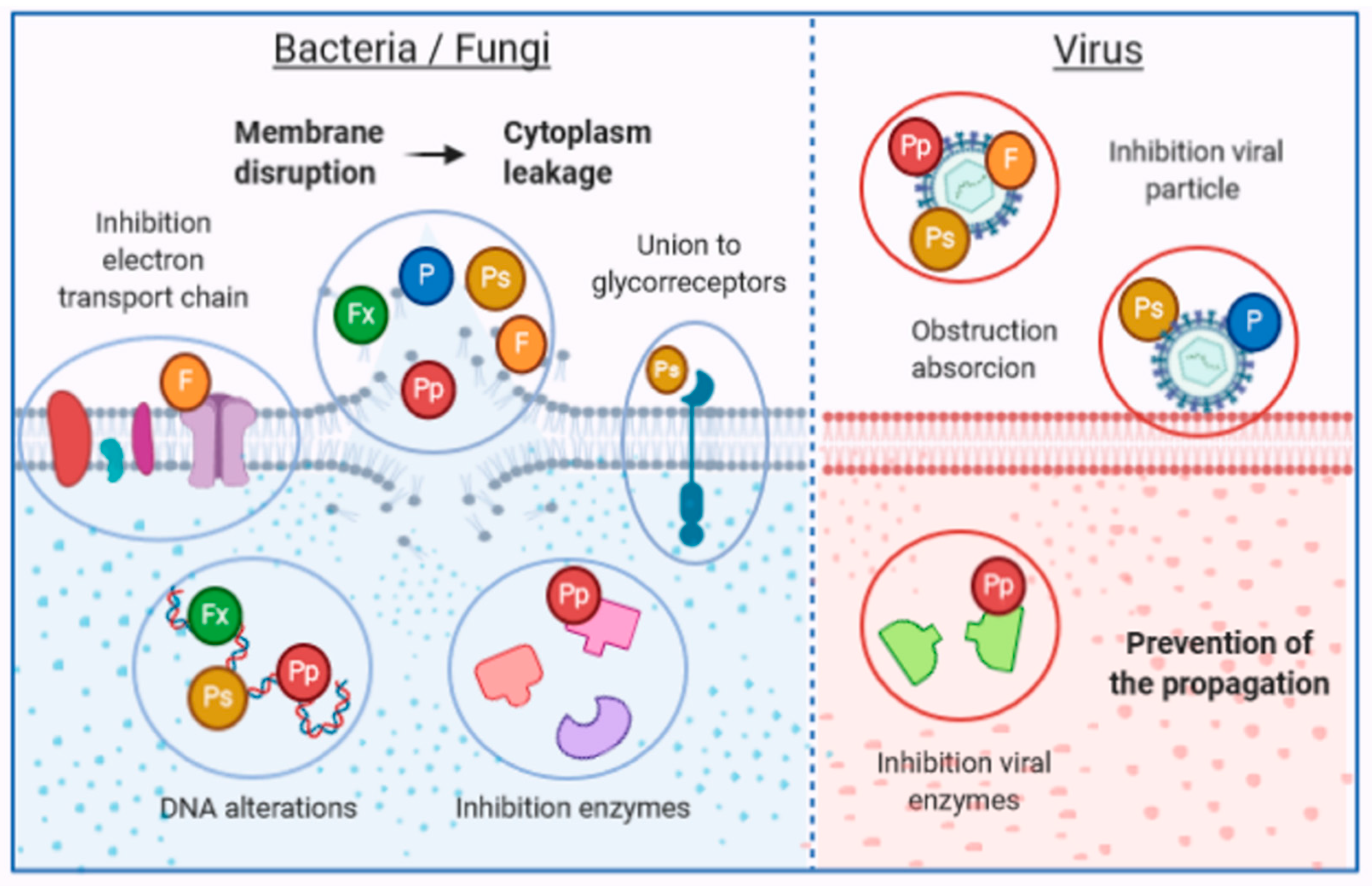
3. Mechanisms of Action of Antimicrobial Compounds
4. Novel Liquid–Liquid and Solid–Liquid Extraction Technologies to Efficiently Extract Algal Bioactive Compounds
4.1. Supercritical Fluid Extraction (SFE)
4.2. Ultrasound Assisted Extraction (UAE)
4.3. Microwave Assisted Extraction (MAE)
4.4. Enzymatic-Assisted Extractions (EAE)
4.5. Accelerated Solvent Extraction (ASE)
4.6. Intensity Pulsed Electric Fields (IPEF)
5. Applications
5.1. Food Industry and Animal Feed
5.2. Cosmetics and Pharmaceuticals
5.3. Anti-Fouling
6. New Approaches and Future Perspectives
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Generic | |
| ATCC | American type culture collection |
| DW | Dry weight |
| FW | Fresh weight |
| HVS | Herpes simplex virus |
| MIC | Minimal inhibition concentration |
| MRSA | Methicillin-resistant Staphylococcus aureus |
| MOI | Multiplicity of infection |
| NPs | Nanoparticles |
| RSM | Response surface methodology |
| RT | Room temperature |
| SARS | Severe acute respiratory syndrome |
| UV | Ultraviolet |
| Extraction techniques | |
| ASE | Accelerated solvent extraction |
| EAE | Enzyme-assisted extraction |
| EUAE | Enzymatic ultrasound assisted extraction |
| HAE | Heat-assisted extraction |
| IPEF | Intensity pulsed electric fields |
| MAE | Microwave-assisted extraction |
| PLE | Pressurized liquid extraction |
| SFE | Supercritical fluid extraction |
| UAE | Ultrasound assisted extraction |
| Compounds | |
| DEt | Diethyl ether |
| DCM | Dichloromethane |
| DPPH | 1,1-diphenyl-2-picryl hydrazyl |
| EtOAc | Ethyl acetate |
| EtOH | Ethanol |
| H2O | Water |
| Hx | Hexane |
| MeOH | Methanol |
| n-BuOH | n- Butanol |
| n-Hx | n-Hexane |
| PUFAs | Poly-unsaturated fatty acids |
| Bacteria species | |
| Acinetobacter (A.): A. baumannii. Actinobacillus (A.): A. actinomycetemcomitas. Actinomyces (A.): A. naeslundii, A. odontolyticus. Aeromonas (A.): A. hydrophyla, A. salmonicida. Aspergillus (A.): A. flavus, A. fumigatus, A. niger. Bacillus (B.): B. cereus, B. mycoides, B. pumilus, B. subtilis. Candida (C.): C. albicans, C. dublinensis, C. famata, C. glabrata, C. guilliermondi, C. kefyr, C. krusei, C. membranaefaciens, C. parapsilosis. Campylobacter (C.): C. jejuni. Chromobacterium (C.): C. violaceum. Colletotrichum (C.): C. lindemuthianum. Cryptococcus (C.): C. neoformans. Cultibacterium (C.): C. acnes. Enterobacter (E.): E. aerogens, E. cloacae. Enterococcus (E.): E. faecalis, E. faecium. Epidermophyton (E.): E. flocossum. Erwinia (E.): E. amylovora. Escherichia (E.): E. coli. Fusarium (F.): F. graminearum, F. culmorum, F. oxysporum, F. solani. Fusobacterium (F.): F. nucleatum. Klebsiella (K.): K. oxytoca, K. pneumonia. Lactobacillus (L.): L. brevis. Listeria (L.): L. innocua, L. monocytogenes. Leishmania (L.): L. amazonensis. Macrophomina (M.): M. phaseolina. Methicillin-resistant Staphylococcus aureus (MRSA). Micrococcus (M.): M. luteus. Microsporum (M.): M. canis, M. gypseum. Penicillium (P.): P. verrucosum. Porphyromonas (P.): P. gingivalis. Pseudoalteromonas (P.): P. bacteriolytica. Proteus (P.): P. vulgaris. Pseudomonas (P.): P. aeruginosa, P. cepacia, P. fluorescens. Rhizoctonia (R.): R. solani. Rhodococcus (R.): R. rhodochrous. Saccharomyces (S.): S. cerevisiae. Salmonella (S.): S. abony, S. choleraesuis, S. gallinarum, S. typhi, S. typhimurium. Septoria (S.): S. glycines. Serratia (S.): S. marcescens. Shigella (S.): S. boydii. Staphylococcus (S.): S. aureus, S. enterica, S. epidermidis, S. lentus, S. xylosus. Streptococcus (S.): S. agalactiae, S. epidermis, S. mutans, S. pneumonia, S. pyogenes, S. sobrinus. Streptomyces (S.): S. purpurascens. Syspastospora (S.): S. parasitica. Trichophyton (T.): T. interdigitale, T. mentagrophytes, T. rubrum, T. tonsurans, Trichosporon (T.): T. mucoides. Trypanosoma (T.): T. cruzi. Vibrio (V.): V. alginolyticus, V. anguillarum, V. cholerae, V. fluvialis, V. haryevi, V. ordalii, V. parahaemolyticus, V. salmonicida, V. splendidus, V. tapetis, V. vulnificus. Yersinia (Y.): Y. ruckeri. | |
| Macroalgae species | |
| Chlorophyta: Boodlea (B.): B. coacta. Caulerpa (C.): C. peltata, C. prolifera, C. racemosa, C. scalpelliformis, C. sertularioides. Cladophora (C.): C. glomerata, C. rivularis, C. rupestris, C. socialis, C. vagabunda. Codium (C.): C. bursa, C. elongatum, C. fragile, C. iyengarii, C. tomentosum. Enteromorpha (E.): E. linza, E. prolifera. Halimeda (H.): H. tuna. Monostroma (M.): M. latissimum. Rhizoclonium (R.): R. implexum. Ulva (U.): U. fasciata, U. flexuosa, U. intestinalis, U. lactuca, U. meridionalis, U. ohnoi, U. pertusa, U. prolifera, U. rigida, U. reticulata. Valonopsis (V.): V. pachynema. Rhodophyta: Acanthophora (A.): A. specifera. Actinotrichia (A.): A. fragilis. Ascophyllum (A.): A. nodosum. Ceramium (C.): C. ciliatum, C. rubrum. Chaetomorpha (C.): C. antemmina, C. linum. Chondrus (C.): C. crispus. Corallina (C.): C. elongata, C. officinalis. Digenea (D.): D. simplex. Dumontia (D.): D. contorta. Eucheuma (E.): E. serra. Galaxaura (G.): G. cylindriea, G. marginata. Gelidium (G.): G. amansii, G. corneum. Gigartina (G.): G. skottsbergii. Gracilaria (G.): G. birdiae, G. changii, G. corticata, G. dura, G. edulis, G. gracilis, G. lemaneiformis, G. multipartita, G. ornata, G. vermiculophylla. Gracilariopsis (G.): G. longissima. Grateloupia (G.): G. doryphora, G. lithophilia. Hypnea (H.): H. esperi, H. flagelliformis, H. musciformis. Jania (J.): J. rubens. Kappaphycus (K.): K. alvarezii. Laurencia (L.): L. aldingensis, L. brandenii, L. catarinensis, L. dendroidea, L. intricata, L. johnstonii, L. obtusa, L. paniculata, L. papillosa, L. translucida. Mastocarpus (M.): M. stellatus. Melanothamnus (M.): M. afaqhusainii. Osmundaria (O.): O. obtusiloba. Osmundea (O.): O. pinnatifida. Palmaria (P.): P. palmata. Polysiphonia (P.): P. elongata. Porphyra (P.): P. dioica, P. haitanensis. Portieria (P.): P. hornemannii. Pterocladia (P.): P. capillacea. Pterocladiella (P.): P. capillacea. Rissoella (R.): R. verruculosa. Rhodomela (R.): R. confervoides. Sarcodia (S.): S. ceylonensis. Ochrophyta: Bifurcaria (B.):B. bifurcata. Canistrocarpus (C.): C. cervicornis. Cladosiphon (C.): C. okamuranus. Cladostephus (C.): C. hirsutus. Carpophyllum (C.): C. flexuosum, C. plumosum. Colpomenia (C.): C. sinuosa. Cutleria (C.): C. multifida. Cystoseira (C.): C. barbata, C. indica, C. mediterranea, C. myrica, C. nodicaulis, C. sedoides, C. trinodis, C. usneoides. Dictyopteris (D.): D. membranacea. Dictyota (D.): D. dichotoma, D. flabellata, D. indica. Dilophus (D.): D. fasciola. Ecklonia (E.): E. arborea, E. cava, E. kurome, E. radiata. Eisenia (E.): E. bicyclis. Feldmannia (F.): F. irregularis. Fucus (F.): F. evanescens, F. serratus, F. spiralis, F. vesiculosus. Gymnogongrus(G.): G. martinensis. Himanthalia (H.): H. elongata. Hormophysa (H.): H. cuneiformes. Iyengaria (I.): I. stellata. Jolyna (J.): J. laminarioides. Laminaria (L.): L. digitata, L. hyperborea, L. japonica. Lessonia (L.): L. nigrecens, L. trabeculata. Nizamuddinia (N.): N. zanardinii. Padina (P.): P. australis, P. concrescens, P. gymnospora. Pelvetia (P.): P. canaliculata. Porolithon (P.): P. antillarum, P. boergesenii. Saccharina (S.): S. latissima, S. longicruris. Sargassum (S.): S. angustifolium, S. boveanum, S. cinereum, S. dentifolium, S. hystrix, S. ilicifolium, S. incisifolium, S. latifolium, S. marginatum, S. muticum, S. myriocystum, S. oligocystum, S. pallidum, S. platycarpum, S. polycystum, S. portieriatum, S. swartzii, S. tenerrimum, S. thunbergii, S. variegatum, S. vulgare, S. wightii. Scytosiphon (S.): S. lomentaria. Solieria (S.): S. chordalis, S. filiformis, S. robusta. Spatoglossum (S.): S. asperum. Spyridia (S.): S. filamentosa. Stoechospermum (S.): S. marginatum. Taonia (T.): T. atomaria. Turbinaria (T.): T. conoides, T. decurrens, T. ornata, T. triquetra. Undaria (U.): U. pinnafitida. | |
References
- Barbosa, F.; Pinto, E.; Kijjoa, A.; Pinto, M.; Sousa, E. Targeting antimicrobial drug resistance with marine natural products. Int. J. Antimicrob. Agents 2020, 5, 106005. [Google Scholar] [CrossRef] [PubMed]
- Gnanavel, V.; Roopan, S.M.; Rajeshkumar, S. Aquaculture: An overview of chemical ecology of seaweeds (food species) in natural products. Aquaculture 2019, 507, 1–6. [Google Scholar] [CrossRef]
- Lourenço-Lopes, C.; Fraga-Corral, M.; Jimenez-Lopez, C.; Pereira, A.G.; Garcia-Oliveira, P.; Carpena, M.; Prieto, M.A.; Simal-Gandara, J. Metabolites from Macroalgae and Its Applications in the Cosmetic Industry: A Circular Economy Approach. Resources 2020, 9, 101. [Google Scholar] [CrossRef]
- Leandro, A.; Pereira, L.; Gonçalves, A.M.M. Diverse applications of marine macroalgae. Mar. Drugs 2020, 18, 17. [Google Scholar] [CrossRef]
- Spalding, H.L.; Amado-Filho, G.M.; Bahia, R.G.; Ballantine, D.L.; Fredericq, S.; Leichter, J.J.; Nelson, W.A.; Slattery, M.; Tsuda, R.T. Macroalgae. In Mesophotic Coral Ecosystems; Springer: Cham, Switzerland, 2019; ISBN 978-3-319-92734-3. [Google Scholar]
- Pepper, I.L.; Gentry, T.J. Microorganisms Found in the Environment. In Environmental Microbiology: Third Edition; Academic Press: Cambridge, MA, USA, 2015; pp. 9–36. ISBN 9780123946263. [Google Scholar]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Anyanwu, R.C.; Rodriguez, C.; Durrant, A.; Olabi, A.G. Micro-Macroalgae Properties and Applications. Ref. Modul. Mater. Sci. Mater. Eng. 2018, 1–28. [Google Scholar] [CrossRef]
- Praveen, M.A.; Parvathy, K.R.K.; Balasubramanian, P.; Jayabalan, R. An overview of extraction and purification techniques of seaweed dietary fibers for immunomodulation on gut microbiota. Trends Food Sci. Technol. 2019, 92, 46–64. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Gobe, G.; Masci, P.; Osborne, S.A. Marine bioactive compounds and health promoting perspectives; innovation pathways for drug discovery. Trends Food Sci. Technol. 2016, 50, 44–55. [Google Scholar] [CrossRef]
- Gurpilhares, D.d.B.; Cinelli, L.P.; Simas, N.K.; Pessoa, A.; Sette, L.D. Marine prebiotics: Polysaccharides and oligosaccharides obtained by using microbial enzymes. Food Chem. 2019, 280, 175–186. [Google Scholar] [CrossRef]
- Belghit, I.; Rasinger, J.D.; Heesch, S.; Biancarosa, I.; Liland, N.; Torstensen, B.; Waagbø, R.; Lock, E.J.; Bruckner, C.G. In-depth metabolic profiling of marine macroalgae confirms strong biochemical differences between brown, red and green algae. Algal Res. 2017, 26, 240–249. [Google Scholar] [CrossRef]
- Vuong, D.; Kaplan, M.; Lacey, H.J.; Crombie, A.; Lacey, E.; Piggott, A.M. A study of the chemical diversity of macroalgae from South Eastern Australia. Fitoterapia 2018, 126, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Bedoux, G.; Hardouin, K.; Burlot, A.S.; Bourgougnon, N. Bioactive components from seaweeds: Cosmetic Applications and Future Development; Elsevier: Amsterdam, The Netherlands, 2014; Volume 71, ISBN 9780124080621. [Google Scholar]
- Hamed, I.; Özogul, F.; Özogul, Y.; Regenstein, J.M. Marine Bioactive Compounds and Their Health Benefits: A Review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 446–465. [Google Scholar] [CrossRef]
- Silva, M.; Vieira, L.; Almeida, A.P.; Kijjoa, A. The Marine Macroalgae of the Genus Ulva: Chemistry, Biological Activities and Potential Applications. J. Oceanogr. Mar. Res. 2013, 1, 1–6. [Google Scholar] [CrossRef]
- Mendes, M.; Pereira, R.; Sousa Pinto, I.; Carvalho, A.P.; Gomes, A.M. Antimicrobial activity and lipid profile of seaweed extracts from the North Portuguese Coast. Int. Food Res. J. 2013, 20, 3337–3345. [Google Scholar]
- Buschmann, A.H.; Camus, C.; Infante, J.; Neori, A.; Israel, Á.; Hernández-González, M.C.; Pereda, S.V.; Gomez-Pinchetti, J.L.; Golberg, A.; Tadmor-Shalev, N.; et al. Seaweed production: Overview of the global state of exploitation, farming and emerging research activity. Eur. J. Phycol. 2017, 52, 391–406. [Google Scholar] [CrossRef]
- Sharma, K.; Guleria, S.; Razdan, V.K.; Babu, V. Synergistic antioxidant and antimicrobial activities of essential oils of some selected medicinal plants in combination and with synthetic compounds. Ind. Crop. Prod. 2020, 154, 112569. [Google Scholar] [CrossRef]
- Abu-Ghannam, N.; Rajauria, G. Antimicrobial activity of compounds isolated from algae. In Functional Ingredients from Algae for Foods and Nutraceuticals; Woodhead Publishing Limited: Shaston, UK, 2013; pp. 287–306. ISBN 9780857095121. [Google Scholar]
- Roca, I.; Akova, M.; Baquero, F.; Carlet, J.; Cavaleri, M.; Coenen, S.; Cohen, J.; Findlay, D.; Gyssens, I.; Heure, O.E.; et al. The global threat of antimicrobial resistance: Science for intervention. New Microbes New Infect. 2015, 6, 22–29. [Google Scholar] [CrossRef]
- Yılmaz Öztürk, B.; Yenice Gürsu, B.; Dağ, İ. Antibiofilm and antimicrobial activities of green synthesized silver nanoparticles using marine red algae Gelidium corneum. Process Biochem. 2020, 89, 208–219. [Google Scholar] [CrossRef]
- Bhowmick, S.; Mazumdar, A.; Moulick, A.; Adam, V. Algal metabolites: An inevitable substitute for antibiotics. Biotechnol. Adv. 2020, 43, 107571. [Google Scholar] [CrossRef]
- Hornsey, I.S.; Hide, D. The production of antimicrobial compounds by British marine algae I. Antibiotic-producing marine algae. Br. Phycol. J. 1974, 9, 353–361. [Google Scholar] [CrossRef]
- Pina-Pérez, M.C.; Rivas, A.; Martínez, A.; Rodrigo, D. Antimicrobial potential of macro and microalgae against pathogenic and spoilage microorganisms in food. Food Chem. 2017, 235, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Chellat, M.F.; Raguž, L.; Riedl, R. Targeting Antibiotic Resistance. Angew. Chemie Int. Ed. 2016, 55, 6600–6626. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Platforms for antibiotic discovery. Nat. Rev. Drug Discov. 2013, 12, 371–387. [Google Scholar] [CrossRef] [PubMed]
- Raguraman, V.; Stanley, A.L.; MubarakAli, D.; Narendrakumar, G.; Thirugnanasambandam, R.; Kirubagaran, R.; Thajuddin, N. Unraveling rapid extraction of fucoxanthin from Padina tetrastromatica: Purification, characterization and biomedical application. Process Biochem. 2018, 73, 211–219. [Google Scholar] [CrossRef]
- Chemat, F.; Abert-Vian, M.; Fabiano-Tixier, A.S.; Strube, J.; Uhlenbrock, L.; Gunjevic, V.; Cravotto, G. Green extraction of natural products. Origins, current status, and future challenges. TrAC Trends Anal. Chem. 2019, 118, 248–263. [Google Scholar] [CrossRef]
- Belwal, T.; Chemat, F.; Venskutonis, P.R.; Cravotto, G.; Jaiswal, D.K.; Bhatt, I.D.; Devkota, H.P.; Luo, Z. Recent advances in scaling-up of non-conventional extraction techniques: Learning from successes and failures. TrAC Trends Anal. Chem. 2020, 127, 115895. [Google Scholar] [CrossRef]
- Baweja, P.; Kumar, S.; Sahoo, D.; Levine, I. Biology of Seaweeds. In Seaweed in Health and Disease Prevention; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 41–106. ISBN 9780128027936. [Google Scholar]
- Cloutier, M.; Mantovani, D.; Rosei, F. Antibacterial Coatings: Challenges, Perspectives, and Opportunities. Trends Biotechnol. 2015, 33, 637–652. [Google Scholar] [CrossRef]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Antibacterial derivatives of marine algae: An overview of pharmacological mechanisms and applications. Mar. Drugs 2016, 14. [Google Scholar] [CrossRef]
- Manilal, A.; Sujith, S.; Sabarathnam, B.; Kiran, G.S.; Selvin, J.; Shakir, C.; Lipton, A.P. Biological activity of the red alga Laurencia brandenii. Acta Bot. Croat. 2011, 70, 81–90. [Google Scholar] [CrossRef]
- dos Santos Amorim, R.d.N.; Rodrigues, J.A.G.; Holanda, M.L.; Quinderé, A.L.G.; de Paula, R.C.M.; Melo, V.M.M.; Benevides, N.M.B. Antimicrobial effect of a crude sulfated polysaccharide from the red seaweed Gracilaria ornata. Brazilian Arch. Biol. Technol. 2012, 55, 171–181. [Google Scholar] [CrossRef]
- Cortés, Y.; Hormazábal, E.; Leal, H.; Urzúa, A.; Mutis, A.; Parra, L.; Quiroz, A. Novel antimicrobial activity of a dichloromethane extract obtained from red seaweed Ceramium rubrum (Hudson) (Rhodophyta: Florideophyceae) against Yersinia ruckeri and Saprolegnia parasitica, agents that cause diseases in salmonids. Electron. J. Biotechnol. 2014, 17, 126–131. [Google Scholar] [CrossRef]
- Sasidharan, S.; Darah, I.; Noordin, M.K.M.J. In vitro antimicrobial activity against Pseudomonas aeruginosa and acute oral toxicity of marine algae Gracilaria changii. N. Biotechnol. 2010, 27, 390–396. [Google Scholar] [CrossRef] [PubMed]
- de Alencar, D.B.; de Carvalho, F.C.T.; Rebouças, R.H.; dos Santos, D.R.; dos Santos Pires-Cavalcante, K.M.; de Lima, R.L.; Baracho, B.M.; Bezerra, R.M.; Viana, F.A.; dos Fernandes Vieira, R.H.S.; et al. Bioactive extracts of red seaweeds Pterocladiella capillacea and Osmundaria obtusiloba (Floridophyceae: Rhodophyta) with antioxidant and bacterial agglutination potential. Asian Pac. J. Trop. Med. 2016, 9, 372–379. [Google Scholar] [CrossRef]
- Baliano, A.P.; Pimentel, E.F.; Buzin, A.R.; Vieira, T.Z.; Romão, W.; Tose, L.V.; Lenz, D.; de Andrade, T.U.; Fronza, M.; Kondratyuk, T.P.; et al. Brown seaweed Padina gymnospora is a prominent natural wound-care product. Rev. Bras. Farmacogn. 2016, 26, 714–719. [Google Scholar] [CrossRef]
- Moubayed, N.M.S.; Al Houri, H.J.; Al Khulaifi, M.M.; Al Farraj, D.A. Antimicrobial, antioxidant properties and chemical composition of seaweeds collected from Saudi Arabia (Red Sea and Arabian Gulf). Saudi J. Biol. Sci. 2017, 24, 162–169. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, J.H.; Jin, H.J.; Lee, S.Y. Antimicrobial activity of ethanol extracts of Laminaria japonica against oral microorganisms. Anaerobe 2013, 21, 34–38. [Google Scholar] [CrossRef]
- Akremi, N.; Cappoen, D.; Anthonissen, R.; Verschaeve, L.; Bouraoui, A. Phytochemical and in vitro antimicrobial and genotoxic activity in the brown algae Dictyopteris membranacea. S. Afr. J. Bot. 2017, 108, 308–314. [Google Scholar] [CrossRef]
- Tajbakhsh, S.; Pouyan, M.; Zandi, K.; Bahramian, P.; Sartavi, K.; Fouladvand, M.; Asayesh, G.; Barazesh, A. In vitro study of antibacterial activity of the alga Sargassum oligocystum from the Persian Gulf. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 293–298. [Google Scholar]
- Salem, W.M.; Galal, H.; Nasr El-deen, F. Screening for antibacterial activities in some marine algae from the red sea (Hurghada, Egypt). African J. Microbiol. Res. 2011, 5, 2160–2167. [Google Scholar] [CrossRef]
- Chiao-Wei, C.; Siew-Ling, H.; Ching-Lee, W. Antibacterial activity of Sargassum polycystum C. Agardh and Padina australis Hauck (Phaeophyceae). Afr. J. Biotechnol. 2011, 10, 14125–14131. [Google Scholar] [CrossRef]
- Assaw, S.; Rosli, N.L.; Azmi, N.A.M.; Mazlan, N.W.; Ismail, N. Antioxidant and antibacterial activities of polysaccharides and methanolic crude extracts of local edible red seaweed Gracilaria sp. Malaysian Appl. Biol. 2018, 47, 135–144. [Google Scholar]
- Mashjoor, S.; Yousefzadi, M.; Esmaeili, M.A.; Rafiee, R. Cytotoxicity and antimicrobial activity of marine macro algae (Dictyotaceae and Ulvaceae) from the Persian Gulf. Cytotechnology 2016, 68, 1717–1726. [Google Scholar] [CrossRef] [PubMed]
- Sirbu, R.; Stanciu, G.; Tomescu, A.; Ionescu, A.M.; Cadar, E. Evaluation of antioxidant and antimicrobial activity in relation to total phenolic content of green algae from Black Sea. Rev. Chim. 2019, 70, 1197–1203. [Google Scholar] [CrossRef]
- Ismail, A.; Ktari, L.; Ben Redjem Romdhane, Y.; Aoun, B.; Sadok, S.; Boudabous, A.; El Bour, M. Antimicrobial Fatty Acids from Green Alga Ulva rigida (Chlorophyta). Biomed Res. Int. 2018, 2018. [Google Scholar] [CrossRef]
- Abdel-Latif, H.H.; Shams El-Din, N.G.; Ibrahim, H.A.H. Antimicrobial activity of the newly recorded red alga Grateloupia doryphora collected from the Eastern Harbor, Alexandria, Egypt. J. Appl. Microbiol. 2018, 125, 1321–1332. [Google Scholar] [CrossRef]
- Omar, H.H.; Shiekh, H.M.; Gumgumjee, N.M.; El-Kazan, M.M.; El-Gendy, A.M. Antibacterial activity of extracts of marine algae from the Red Sea of Jeddah, Saudi Arabia. Afr. J. Biotechnol. 2012, 11, 13576–13585. [Google Scholar] [CrossRef]
- Al Khazan, M.M.; Omar, H.H.; Gumgumjee, N.M.; Shiekh, H.M.; El-Gendy, A.M. Marine macroalgae as a potential source of bioactive natural products with antibacterial activity. Main Group Chem. 2016, 15, 139–151. [Google Scholar] [CrossRef]
- KOSANIĆ, M.; RANKOVIĆ, B.; Stanojković, T. Evaluation of antioxidant, antimicrobial and anticancer effects of three selected marine macroalgae. Rom. Biotechnol. Lett. 2018, 1–10. [Google Scholar]
- Ambreen; Hira, K.; Tariq, A.; Ruqqia, R.; Sultana, V.; Jehan, A. Evaluation of biochemical component and antimicrobial activity of some seaweeeds occurring at Karachi coast. Pakistan J. Bot. 2012, 44, 1799–1803. [Google Scholar]
- Cavallo, R.A.; Acquaviva, M.I.; Stabili, L.; Cecere, E.; Petrocelli, A.; Narracci, M. Antibacterial activity of marine macroalgae against fish pathogenic Vibrio species. Cent. Eur. J. Biol. 2013, 8, 646–653. [Google Scholar] [CrossRef]
- Stabili, L.; Acquaviva, M.I.; Biandolino, F.; Cavallo, R.A.; De Pascali, S.A.; Fanizzi, F.P.; Narracci, M.; Cecere, E.; Petrocelli, A. Biotechnological potential of the seaweed Cladophora rupestris (Chlorophyta, Cladophorales) lipidic extract. New Biotechnol. 2014, 31, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Stabili, L.; Acquaviva, M.I.; Biandolino, F.; Cavallo, R.A.; de Pascali, S.A.; Fanizzi, F.P.; Narracci, M.; Petrocelli, A.; Cecere, E. The lipidic extract of the seaweed Gracilariopsis longissima (Rhodophyta, Gracilariales): A potential resource for biotechnological purposes? New Biotechnol. 2012, 29, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Manikandan, S.; Ganesapandian, S.; Singh, M.; Sangeetha, N.; Kumaraguru, A.K. Antimicrobial activity of seaweeds against multi drug resistant strains. Int. J. Pharmacol. 2011, 7, 522–526. [Google Scholar] [CrossRef]
- Osman, M.E.H.; Abushady, A.M.; Elshobary, M.E. In vitro screening of antimicrobial activity of extracts of some macroalgae collected from Abu-Qir bay Alexandria, Egypt. Afr. J. Biotechnol. 2010, 9, 7203–7208. [Google Scholar] [CrossRef]
- Águila-Ramírez, R.N.; Arenas-González, A.; Hernández-Guerrero, C.J.; González-Acosta, B.; Borges-Souza, J.M.; Véron, B.; Pope, J.; Hellio, C. Antimicrobial and antifouling activities achieved by extracts of seaweeds from Gulf of California, Mexico. Hidrobiologica 2012, 22, 8–15. [Google Scholar]
- Lee, J.-H.; Eom, S.-H.; Lee, E.-H.; Jung, Y.-J.; Kim, H.-J.; Jo, M.-R.; Son, K.-T.; Lee, H.-J.; Kim, J.H.; Lee, M.-S.; et al. In vitro antibacterial and synergistic effect of phlorotannins isolated from edible brown seaweed Eisenia bicyclis against acne-related bacteria. ALGAE 2014. [Google Scholar] [CrossRef]
- Tajbakhsh, S.; Ilkhani, M.; Rustaiyan, A.; Larijani, K.; Sartavi, K.; Tahmasebi, R.; Asayesh, G. Antibacterial effect of the brown alga Cystoseira trinodis. J. Med. Plant Res. 2011, 5, 4654–4657. [Google Scholar]
- Ertürk, Ö.; Taş, B. Antibacterial and antifungal effects of some marine algae. Kafkas Univ. Vet. Fak. Derg. 2011, 17, 121–124. [Google Scholar]
- Ghania, A.; Nabila, B.B.; Larbi, B.; Elisabeth, M.; Philippe, G.; Mariem, B.; Khadidja, K.K.; Wacila, B.R.; Fawzia, A.B. Antimicrobial and antiparasitic activities of three algae from the northwest coast of Algeria. Nat. Prod. Res. 2019, 33, 742–745. [Google Scholar] [CrossRef]
- Stein, E.M.; Colepicolo, P.; Afonso, F.A.K.; Fujii, M.T. Screening for antifungal activities of extracts of the Brazilian seaweed genus Laurencia (Ceramiales, Rhodophyta). Braz. J. Pharmacogn. 2011, 21, 290–295. [Google Scholar] [CrossRef]
- Aoun, Z.B.; Said, R.B.; Farhat, F. Anti-inflammatory, antioxidant and antimicrobial activities of aqueous and organic extracts from Dictyopteris membranacea. Bot. Mar. 2010, 53, 259–264. [Google Scholar] [CrossRef]
- Rosaline, X.D.; Sakthivelkumar, S.; Rajendran, K.; Janarthanan, S. Screening of selected marine algae from the coastal Tamil Nadu, South India for antibacterial activity. Asian Pac. J. Trop. Biomed. 2012, 2. [Google Scholar] [CrossRef]
- Goecke, F.; Labes, A.; Wiese, J.; Imhoff, J.F. Dual effect of macroalgal extracts on growth of bacteria in Western Baltic Sea. Rev. Biol. Mar. Oceanogr. 2012, 47, 75–86. [Google Scholar] [CrossRef]
- Milović, S.; Kundaković, T.; Mačić, V.; Stanković, J.A.; Grozdanić, N.; Đuričić, I.; Stanković, I.; Stanojković, T. Anti α-glucosidase, antitumour, antioxidative, antimicrobial activity, nutritive and health protective potential of some seaweeds from the Adriatic coast of Montenegro. Farmacia 2017, 65, 731–740. [Google Scholar]
- Arulkumar, A.; Rosemary, T.; Paramasivam, S.; Rajendran, R.B. Phytochemical composition, in vitro antioxidant, antibacterial potential and GC-MS analysis of red seaweeds (Gracilaria corticata and Gracilaria edulis) from Palk Bay, India. Biocatal. Agric. Biotechnol. 2018, 15, 63–71. [Google Scholar] [CrossRef]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish seaweeds. Int. Food Res. J. 2010, 17, 205–220. [Google Scholar] [CrossRef]
- Arumugam, G.; Rajendran, R. Anti-candidal activity and synergetic interaction of antifungal drugs with differential extract of brown algae Stocheospermum marginatum. Biocatal. Agric. Biotechnol. 2019, 19, 101145. [Google Scholar] [CrossRef]
- Taskin, E.; Caki, Z.; Ozturk, M.; Taskin, E. Assessment of in vitro antitumoral and antimicrobial activities of marine algae harvested from the eastern Mediterranean sea. Afr. J. Biotechnol. 2010, 9, 4272–4277. [Google Scholar] [CrossRef]
- Kosanić, M.; Ranković, B.; Stanojković, T. Biological activities of two macroalgae from Adriatic coast of Montenegro. Saudi J. Biol. Sci. 2015, 22, 390–397. [Google Scholar] [CrossRef]
- Osman, N.A.H.K.; Siam, A.A.; El-manawy, I.M.; Jeon, Y. Anti-microbial and Anti-diabetic Activity of Six Seaweeds Collected from the Red Sea, Egypt. Catrina Int. J. Environ. Sci. 2019, 19, 55–60. [Google Scholar]
- Kang, O.; Brice, O.; Lee, Y.; Chae, H. Antibacterial activity of Ecklonia cava against methicillin-resistant Staphylococcus aureus and Salmonella spp. Foodborne Pathog. Dis. 2010, 7, 435–441. [Google Scholar] [CrossRef]
- Abdel-Raouf, N.; Mohamed, H.; Mostafa, S.; Ibraheem, I. Controlling of Microbial Growth by Using Cystoseira barbata Extract. Egypt. J. Bot. 2017, 57, 469–477. [Google Scholar] [CrossRef]
- Bhuyar, P.; Rahim, M.H.; Sundararaju, S.; Maniam, G.P.; Govindan, N. Antioxidant and antibacterial activity of red seaweed; Kappaphycus alvarezii against pathogenic bacteria. Glob. J. Environ. Sci. Manag. 2020, 5, 47–58. [Google Scholar] [CrossRef]
- Guedes, E.A.C.; dos Santos Araújo, M.A.; Souza, A.K.P.; de Souza, L.I.O.; de Barros, L.D.; de Albuquerque Maranhão, F.C.; Sant’Ana, A.E.G. Antifungal Activities of Different Extracts of Marine Macroalgae Against Dermatophytes and Candida Species. Mycopathologia 2012, 174, 223–232. [Google Scholar] [CrossRef]
- Alves, C.; Pinteus, S.; Simões, T.; Horta, A.; Silva, J.; Tecelão, C.; Pedrosa, R. Bifurcaria bifurcata: A key macro-alga as a source of bioactive compounds and functional ingredients. Int. J. Food Sci. Technol. 2016, 51, 1638–1646. [Google Scholar] [CrossRef]
- Jassbi, A.R.; Mohabati, M.; Eslami, S.; Sohrabipour, J.; Miri, R. Biological activity and chemical constituents of red and brown algae from the Persian Gulf. Iran. J. Pharm. Res. 2013, 12, 339–348. [Google Scholar] [CrossRef]
- Shanmugam, S.K.; Kumar, Y.; Yar, K.M.S.; Gupta, V.; de Clercq, E. Antimicrobial and cytotoxic activities of Turbinaria conoides (J.Agardh) Kuetz. Iran. J. Pharm. Res. 2010, 9, 411–416. [Google Scholar] [CrossRef]
- Kosanić, M.; Ranković, B.; Stanojković, T. Brown macroalgae from the Adriatic Sea as a promising source of bioactive nutrients. J. Food Meas. Charact. 2019, 13, 330–338. [Google Scholar] [CrossRef]
- Chan, Y.S.; Ong, C.W.; Chuah, B.L.; Khoo, K.S.; Chye, F.Y.; Sit, N.W. Antimicrobial, antiviral and cytotoxic activities of selected marine organisms collected from the coastal areas of Malaysia. J. Mar. Sci. Technol. 2018, 26, 128–136. [Google Scholar] [CrossRef]
- Deveau, A.M.; Miller-Hope, Z.; Lloyd, E.; Williams, B.S.; Bolduc, C.; Meader, J.M.; Weiss, F.; Burkholder, K.M. Antimicrobial activity of extracts from macroalgae Ulva lactuca against clinically important Staphylococci is impacted by lunar phase of macroalgae harvest. Lett. Appl. Microbiol. 2016, 62, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Rahelivao, M.P.; Gruner, M.; Andriamanantoanina, H.; Bauer, I.; Knölker, H.J. Brown Algae (Phaeophyceae) from the Coast of Madagascar: Preliminary Bioactivity Studies and Isolation of Natural Products. Nat. Products Bioprospect. 2015, 5, 223–235. [Google Scholar] [CrossRef]
- Mickymaray, S.; Alturaiki, W. Antifungal efficacy of marine macroalgae against fungal isolates from bronchial asthmatic cases. Molecules 2018, 23, 3032. [Google Scholar] [CrossRef]
- Selim, S.; Amin, A.; Hassan, S.; Hagazey, M. Antibacterial, cytotoxicity and anticoagulant activities from Hypnea esperi and Caulerpa prolifera marine algae. Pak. J. Pharm. Sci. 2015, 28, 525–530. [Google Scholar] [PubMed]
- Rahelivao, M.P.; Gruner, M.; Andriamanantoanina, H.; Andriamihaja, B.; Bauer, I.; Knölker, H.J. Red algae (Rhodophyta) from the coast of Madagascar: Preliminary bioactivity studies and isolation of natural products. Mar. Drugs 2015, 13, 4197–4216. [Google Scholar] [CrossRef]
- Rajauria, G.; Jaiswal, A.K.; Abu-Gannam, N.; Gupta, S. Antimicrobial, antioxidant and free radical-scavenging capacity of brown seaweed Himanthalia elongata from western coast of Ireland. J. Food Biochem. 2013, 37, 322–335. [Google Scholar] [CrossRef]
- Higgins Hoare, A.; Tan, S.P.; McLoughlin, P.; Mulhare, P.; Hughes, H. The Screening and Evaluation of Fucus serratus and Fucus vesiculosus Extracts against Current Strains of MRSA Isolated from a Clinical Hospital Setting. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Van Tran, T.T.; Truong, H.B.; Tran, N.H.V.; Quach, T.M.T.; Nguyen, T.N.; Bui, M.L.; Yuguchi, Y.; Thanh, T.T.T. Structure, conformation in aqueous solution and antimicrobial activity of ulvan extracted from green seaweed Ulva reticulata. Nat. Prod. Res. 2018, 32, 2291–2296. [Google Scholar] [CrossRef]
- Trigui, M.; Gasmi, L.; Zouari, I.; Tounsi, S. Seasonal variation in phenolic composition, antibacterial and antioxidant activities of Ulva rigida (Chlorophyta) and assessment of antiacetylcholinesterase potential. J. Appl. Phycol. 2013, 25, 319–328. [Google Scholar] [CrossRef]
- Vijayavel, K.; Martinez, J.A. In vitro antioxidant and antimicrobial activities of two Hawaiian marine Limu: Ulva fasciata (Chlorophyta) and Gracilaria salicornia (Rhodophyta). J. Med. Food 2010, 13, 1494–1499. [Google Scholar] [CrossRef]
- Zouaoui, B.; Ghalem, B.R. The Phenolic Contents and Antimicrobial Activities of Some Marine Algae from the Mediterranean Sea (Algeria). Russ. J. Mar. Biol. 2017, 43, 491–495. [Google Scholar] [CrossRef]
- Vieira, E.F.; Soares, C.; Machado, S.; Correia, M.; Ramalhosa, M.J.; Oliva-teles, M.T.; Paula Carvalho, A.; Domingues, V.F.; Antunes, F.; Oliveira, T.A.C.; et al. Seaweeds from the Portuguese coast as a source of proteinaceous material: Total and free amino acid composition profile. Food Chem. 2018, 269, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Cherry, P.; Yadav, S.; Strain, C.R.; Allsopp, P.J.; Mcsorley, E.M.; Ross, R.P.; Stanton, C. Prebiotics from seaweeds: An ocean of opportunity? Mar. Drugs 2019, 17, 327. [Google Scholar] [CrossRef] [PubMed]
- Ciko, A.M.; Jokić, S.; Šubarić, D.; Jerković, I. Overview on the application of modern methods for the extraction of bioactive compounds from marine macroalgae. Mar. Drugs 2018, 16, 348. [Google Scholar] [CrossRef] [PubMed]
- Bak, U.G.; Nielsen, C.W.; Marinho, G.S.; Gregersen, Ó.; Jónsdóttir, R.; Holdt, S.L. The seasonal variation in nitrogen, amino acid, protein and nitrogen-to-protein conversion factors of commercially cultivated Faroese Saccharina latissima. Algal Res. 2019, 42, 101576. [Google Scholar] [CrossRef]
- Stack, J.; Tobin, P.R.; Gietl, A.; Harnedy, P.A.; Stengel, D.B.; FitzGerald, R.J. Seasonal variation in nitrogenous components and bioactivity of protein hydrolysates from Porphyra dioica. J. Appl. Phycol. 2017, 29, 2439–2450. [Google Scholar] [CrossRef]
- Pangestuti, R.; Kim, S. Seaweed proteins, peptides, and amino acids. In Seaweed Sustainablity; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 125–140. ISBN 9780124186972. [Google Scholar]
- Cerná, M. Seaweed Proteins and Amino Acids as Nutraceuticals. In Advances in Food and Nutrition Research; Elsevier Inc.: Amsterdam, The Netherlands, 2011; Volume 64, pp. 297–312. ISBN 9780123876690. [Google Scholar]
- Admassu, H.; Abdalbasit, M.; Yang, R.; Zhao, W.; War, W. Bioactive Peptides Derived from Seaweed Protein and Their Health Benefits: Antihypertensive, Antioxidant, and Antidiabetic Properties. J. Food Sci. 2018, 83, 6–16. [Google Scholar] [CrossRef]
- Beaulieu, L.; Bondu, S.; Doiron, K.; Rioux, L.; Turgeon, S.L. Characterization of antibacterial activity from protein hydrolysates of the macroalga Saccharina longicruris and identification of peptides implied in bioactivity. J. Funct. Foods 2015, 17, 685–697. [Google Scholar] [CrossRef]
- Cordeiro, R.A.; Gomes, V.M.; Carvalho, A.F.U.; Melo, V.M.M. Effect of proteins from the red seaweed Hypnea musciformis (Wulfen) lamouroux on the growth of human pathogen yeasts. Braz. Arch. Biol. Technol. 2006, 49, 915–921. [Google Scholar] [CrossRef]
- Holanda, M.L.; Melo, V.M.M.; Silva, L.M.C.M.; Amorim, R.C.N.; Pereira, M.G.; Benevides, N.M.B. Differential activity of a lectin from Solieria filiformis against human pathogenic bacteria. Brazilian J. Med. Biol. Res. 2005, 38, 1769–1773. [Google Scholar] [CrossRef]
- Xu, S.; Huang, X.; Cheong, K. Recent Advances in Marine Algae Polysaccharides: Isolation, Structure, and Activities. Mar. Drugs 2017, 15, 388. [Google Scholar] [CrossRef] [PubMed]
- Berri, M.; Olivier, M.; Holbert, S.; Dupont, J.; Demais, H.; Le Goff, M.; Collen, P.N. Ulvan from Ulva armoricana (Chlorophyta) activates the PI3K/Akt signalling pathway via TLR4 to induce intestinal cytokine production. Algal Res. 2017, 28, 39–47. [Google Scholar] [CrossRef]
- Cho, M.; You, S. Sulfated Polysaccharides from Green Seaweeds. In Springer Handbook of Marine Biotechnology; Springer: Berlin/Heidelberg, Germany, 2015; pp. 941–953. [Google Scholar]
- Bouhlal, R.; Haslin, C.; Chermann, J.C.; Colliec-Jouault, S.; Sinquin, C.; Simon, G.; Cerantola, S.; Riadi, H.; Bourgougnon, N. Antiviral activities of sulfated polysaccharides isolated from Sphaerococcus coronopifolius (Rhodophytha, Gigartinales) and Boergeseniella thuyoides (Rhodophyta, Ceramiales). Mar. Drugs 2011, 9, 1187–1209. [Google Scholar] [CrossRef] [PubMed]
- Ermakova, S.; Men’shova, R.; Vishchuk, O.; Kim, S.M.; Um, B.H.; Isakov, V.; Zvyagintseva, T. Water-soluble polysaccharides from the brown alga Eisenia bicyclis: Structural characteristics and antitumor activity. Algal Res. 2013, 2, 51–58. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; De Morais, A.M.B.; De Morais, R.M.S.C. Marine polysaccharides from algae with potential biomedical applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef] [PubMed]
- O’Doherty, J.V.; McDonnell, P.; Figat, S. The effect of dietary laminarin and fucoidan in the diet of the weanling piglet on performance and selected faecal microbial populations. Livest. Sci. 2010, 134, 208–210. [Google Scholar] [CrossRef]
- Belattmania, Z.; Engelen, A.H.; Pereira, H.; Serrão, E.A.; Custódio, L.; Varela, J.C.; Zrid, R.; Reani, A.; Sabour, B. Fatty acid composition and nutraceutical perspectives of brown seaweeds from the Atlantic coast of Morocco. Int. Food Res. J. 2018, 25, 1520–1527. [Google Scholar]
- Kendel, M.; Wielgosz-Collin, G.; Bertrand, S.; Roussakis, C.; Bourgougnon, N.B.; Bedoux, G. Lipid composition, fatty acids and sterols in the seaweeds ulva armoricana, and solieria chordalis from brittany (France): An analysis from nutritional, chemotaxonomic, and antiproliferative activity perspectives. Mar. Drugs 2015, 13, 5606–5628. [Google Scholar] [CrossRef]
- Schmid, M.; Guihéneuf, F.; Stengel, D.B. Fatty acid contents and profiles of 16 macroalgae collected from the Irish Coast at two seasons. J. Appl. Phycol. 2014, 26, 451–463. [Google Scholar] [CrossRef]
- Cardoso, C.; Ripol, A.; Afonso, C.; Freire, M.; Varela, J.; Quental-Ferreira, H.; Pousão-Ferreira, P.; Bandarra, N. Fatty acid profiles of the main lipid classes of green seaweeds from fish pond aquaculture. Food Sci. Nutr. 2017, 5, 1186–1194. [Google Scholar] [CrossRef]
- El Baz, F.K.; El Baroty, G.S.; Abd El Baky, H.H.; Abd El-Salam, O.I.; Ibrahim, E.A. Structural characterization and biological activity of sulfolipids from selected marine algae. Grasas y Aceites 2013, 64, 561–571. [Google Scholar] [CrossRef]
- Mekinić, I.G.; Skroza, D.; Šimat, V.; Hamed, I.; Čagalj, M.; Perković, Z.P. Phenolic content of brown algae (Pheophyceae) species: Extraction, identification, and quantification. Biomolecules 2019, 9, 244. [Google Scholar] [CrossRef]
- Gómez-Guzmán, M.; Rodríguez-Nogales, A.; Algieri, F.; Gálvez, J. Potential role of seaweed polyphenols in cardiovascular-associated disorders. Mar. Drugs 2018, 16, 250. [Google Scholar] [CrossRef] [PubMed]
- Cassani, L.; Gomez-zavaglia, A.; Jimenez-lopez, C.; Prieto, M.A.; Simal-gandara, J. Seaweed-based natural ingredients: Stability of phlorotannins during extraction, storage, passage through the gastrointestinal tract and potential incorporation into functional foods. Food Res. Int. 2020, 109676. [Google Scholar] [CrossRef]
- Montero, L.; del Pilar Sánchez-Camargo, A.; Ibáñez, E.; Gilbert-López, B. Phenolic Compounds from Edible Algae: Bioactivity and Health Benefits. Curr. Med. Chem. 2017, 25, 4808–4826. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave assisted extraction of phenolic compounds from four economic brown macroalgae species and evaluation of their antioxidant activities and inhibitory effects on α-amylase, α-glucosidase, pancreatic lipase and tyrosinase. Food Res. Int. 2018, 113, 288–297. [Google Scholar] [CrossRef]
- Nagayama, K.; Iwamura, Y.; Shibata, T.; Hirayama, I.; Nakamura, T. Bactericidal activity of phlorotannins from the brown alga Ecklonia kurome. J. Antimicrob. Chemother. 2002, 50, 889–893. [Google Scholar] [CrossRef]
- Aryee, A.N.; Agyei, D.; Akanbi, T.O. Recovery and utilization of seaweed pigments in food processing. Curr. Opin. Food Sci. 2018, 19, 113–119. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- Karpinski, T.M.; Adamczak, A. Fucoxanthin—An Antibacterial Carotenoid. Antioxidants 2019, 8, 239. [Google Scholar] [CrossRef]
- Bogolitsyn, K.; Dobrodeeva, L.; Druzhinina, A.; Ovchinnikov, D.; Parshina, A.; Shulgina, E. Biological activity of a polyphenolic complex of Arctic brown algae. J. Appl. Phycol. 2019, 31, 3341–3348. [Google Scholar] [CrossRef]
- Wei, Y.; Liu, Q.; Xu, C.; Yu, J.; Zhao, L.; Guo, Q. Damage to the Membrane Permeability and Cell Death of Vibrio parahaemolyticus Caused by Phlorotannins with Low Molecular Weight from Sargassum thunbergii. J. Aquat. Food Prod. Technol. 2016, 25, 323–333. [Google Scholar] [CrossRef]
- Eom, S.H.; Lee, D.S.; Jung, Y.J.; Park, J.H.; Choi, J.I.; Yim, M.J.; Jeon, J.M.; Kim, H.W.; Son, K.T.; Je, J.Y.; et al. The mechanism of antibacterial activity of phlorofucofuroeckol-A against methicillin-resistant Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2014, 98, 9795–9804. [Google Scholar] [CrossRef] [PubMed]
- Cherian, C.; Jannet Vennila, J.; Sharan, L. Marine bromophenols as an effective inhibitor of virulent proteins (peptidyl arginine deiminase, gingipain R and hemagglutinin A) in Porphyromas gingivalis. Arch. Oral Biol. 2019, 100, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Lee, K.B.; Oh, S.M.; Lee, B.H.; Chee, H.Y. Antifungal activities of dieckol isolated from the marine brown alga Ecklonia cava against Trichophyton rubrum. J. Appl. Biol. Chem. 2010, 53, 504–507. [Google Scholar] [CrossRef]
- Lopes, G.; Pinto, E.; Andrade, P.B.; Valentão, P. Antifungal Activity of Phlorotannins against Dermatophytes and Yeasts: Approaches to the Mechanism of Action and Influence on Candida albicans Virulence Factor. PLoS ONE 2013, 8, e72203. [Google Scholar] [CrossRef]
- Ryu, Y.B.; Jeong, H.J.; Yoon, S.Y.; Park, J.Y.; Kim, Y.M.; Park, S.J.; Rho, M.C.; Kim, S.J.; Lee, W.S. Influenza virus neuraminidase inhibitory activity of phlorotannins from the edible brown alga Ecklonia cava. J. Agric. Food Chem. 2011, 59, 6467–6473. [Google Scholar] [CrossRef]
- Morán-Santibañez, K.; Peña-Hernáncez, M.A.; Cruz-Suárez, L.E.; Ricque-Marie, D.; Skouta, R.; Vasquez, A.H.; Rodríguez-Padilla, C.; Trejo-Avila, L.M. Virucidal and Synergistic Activity of Polyphenol-Rich Extracts of Seaweeds against Measles Virus. Viruses 2018, 10, 465. [Google Scholar] [CrossRef]
- Liu, M.; Liu, Y.; Cao, M.; Liu, G.; Chen, Q.; Sun, L.; Chen, H. Antibacterial activity and mechanisms of depolymerized fucoidans isolated from Laminaria japonica. Carbohydr. Polym. 2017, 172, 294–305. [Google Scholar] [CrossRef]
- Id, J.J.; Jung, M.; Jeong, I.; Yamazaki, K.; Kawai, Y. Antimicrobial and Antibiofilm Activities of Sulfated Polysaccharides from Marine Algae against Dental. Mar. Drugs 2018, 16, 301. [Google Scholar] [CrossRef]
- Kadam, S.U.; Donnell, C.P.O.; Rai, D.K.; Hossain, M.B.; Burgess, C.M.; Walsh, D.; Tiwari, B.K. Laminarin from Irish brown seaweeds Ascophyllum nodosum and Laminaria hyperborea: Ultrasound assisted extraction, characterization and bioactivity. Mar. Drugs 2015, 13, 4270–4280. [Google Scholar] [CrossRef] [PubMed]
- Zeid, A.H.A.; Aboutabl, E.A.; Sleem, A.A.; El-rafie, H.M. Water soluble polysaccharides extracted from Pterocladia capillacea and Dictyopteris membranacea and their biological activities. Carbohydr. Polym. 2014, 113, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Siahaan, E.A.; Pangestuti, R.; Kim, S.-K. Seaweeds: Valuable Ingredients for the Pharmaceutical Industries; Springer: Cham, Siwtzerland, 2018; ISBN 9783319690759. [Google Scholar]
- Singh, R.S.; Walia, A.K. Lectins from red algae and their biomedical potential. J. Appl. Phycol. 2018, 30, 1833–1858. [Google Scholar] [CrossRef] [PubMed]
- Cheung, R.C.F.; Wong, J.H.; Pan, W.; Chan, Y.S.; Yin, C.; Dan, X.; Ng, T.B. Marine lectins and their medicinal applications. Appl. Microbiol. Biotechnol. 2015, 99, 3755–3773. [Google Scholar] [CrossRef]
- El Shafay, S.M.; Ali, S.S.; El-Sheekh, M.M. Antimicrobial activity of some seaweeds species from Red sea, against multidrug resistant bacteria. Egypt. J. Aquat. Res. 2016, 42, 65–74. [Google Scholar] [CrossRef]
- Kasanah, N.; Amelia, W.; Mukminin, A.; Triyanto; Isnansetyo, A. Antibacterial activity of Indonesian red algae Gracilaria edulis against bacterial fish pathogens and characterization of active fractions. Nat. Prod. Res. 2019, 33, 3303–3307. [Google Scholar] [CrossRef]
- Anjali, K.P.; Sangeetha, B.M.; Devi, G.; Raghunathan, R.; Dutta, S. Bioprospecting of seaweeds (Ulva lactuca and Stoechospermum marginatum): The compound characterization and functional applications in medicine-a comparative study. J. Photochem. Photobiol. B Biol. 2019, 200, 111622. [Google Scholar] [CrossRef]
- de Felício, R.; de Albuquerque, S.; Young, M.C.M.; Yokoya, N.S.; Debonsi, H.M. Trypanocidal, leishmanicidal and antifungal potential from marine red alga Bostrychia tenella J. Agardh (Rhodomelaceae, Ceramiales). J. Pharm. Biomed. Anal. 2010, 52, 763–769. [Google Scholar] [CrossRef]
- Wang, H.; Li, Y.; Shen, W.; Rui, W.; Ma, X.; Cen, Y. Antiviral activity of a sulfoquinovosyldiacylglycerol ( SQDG ) compound isolated from the green alga Caulerpa racemosa. Bot. Mar. 2007, 50, 185–190. [Google Scholar] [CrossRef]
- Rajauria, G.; Abu-ghannam, N. Isolation and Partial Characterization of Bioactive Fucoxanthin from Himanthalia elongata Brown Seaweed: A TLC-Based Approach. Int. J. Anal. Chem. 2013, 2013. [Google Scholar] [CrossRef]
- Gerasimenko, N.I.; Martyyas, E.A.; Logvinov, S.V.; Busarova, N.G. Biological activity of lipids and photosynthetic pigments of Sargassum pallidum C. Agardh. Appl. Biochem. Microbiol. 2014, 50, 73–81. [Google Scholar] [CrossRef]
- Mendiola, J.A.; Santoyo, S.; Cifuentes, A.; Reglero, G.; Ibáñez, E.; Javier Señoráns, F. Antimicrobial activity of sub- and supercritical CO2 extracts of the green alga Dunaliella salina. J. Food Prot. 2008, 71, 2138–2143. [Google Scholar] [CrossRef] [PubMed]
- Tyskiewicz, K.; Tyskiewicz, R.; Konkol, M.; Rój, E.; Jaroszuk-Sciseł, J.; Skalicka-Wozniak, K. Antifungal Properties of Fucus vesiculosus L. Supercritical Fluid Extract against Fusarium culmorum and Fusarium oxysporum. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Alboofetileh, M.; Rezaei, M.; Tabarsa, M.; Rittà, M.; Donalisio, M.; Mariatti, F.; You, S.G.; Lembo, D.; Cravotto, G. Effect of different non-conventional extraction methods on the antibacterial and antiviral activity of fucoidans extracted from Nizamuddinia zanardinii. Int. J. Biol. Macromol. 2019, 124, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Munir, N.; Rafique, M.; Altaf, I.; Sharif, N.; Naz, S. Antioxidant and antimicrobial activities of extracts from selected algal species. Bangladesh J. Bot. 2018, 47, 53–61. [Google Scholar]
- Sabeena, S.F.; Alagarsamy, S.; Sattari, Z.; Al-Haddad, S.; Fakhraldeen, S.; Al-Ghunaim, A.; Al-Yamani, F. Enzyme-assisted extraction of bioactive compounds from brown seaweeds and characterization. J. Appl. Phycol. 2020, 32, 615–629. [Google Scholar] [CrossRef]
- Meillisa, A.; Siahaan, E.A.; Park, J.N.; Woo, H.C.; Chun, B.S. Effect of subcritical water hydrolysate in the brown seaweed Saccharina japonica as a potential antibacterial agent on food-borne pathogens. J. Appl. Phycol. 2013, 25, 763–769. [Google Scholar] [CrossRef]
- Otero, P.; Quintana, S.E.; Reglero, G.; Fornari, T.; García-Risco, M.R. Pressurized Liquid Extraction (PLE) as an innovative green technology for the effective enrichment of galician algae extracts with high quality fatty acids and antimicrobial and antioxidant properties. Mar. Drugs 2018, 16. [Google Scholar] [CrossRef]
- Plaza, M.; Santoyo, S.; Jaime, L.; García-Blairsy Reina, G.; Herrero, M.; Señoráns, F.J.; Ibáñez, E. Screening for bioactive compounds from algae. J. Pharm. Biomed. Anal. 2010, 51, 450–455. [Google Scholar] [CrossRef]
- Heavisides, E.; Rouger, C.; Reichel, A.F.; Ulrich, C.; Wenzel-Storjohann, A.; Sebens, S.; Tasdemir, D. Seasonal variations in the metabolome and bioactivity profile of fucus vesiculosus extracted by an optimised, pressurised liquid extraction protocol. Mar. Drugs 2018, 16, 503. [Google Scholar] [CrossRef]
- Sosa-Hernández, J.E.; Escobedo-Avellaneda, Z.; Iqbal, H.M.N.; Welti-Chanes, J. State-of-the-art extraction methodologies for bioactive compounds from algal biome to meet bio-economy challenges and opportunities. Molecules 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Gallego, R.; Bueno, M.; Herrero, M. Sub- and supercritical fl uid extraction of bioactive compounds from plants, food-by-products, seaweeds and microalgae—An update. Trends Anal. Chem. 2019, 116, 198–213. [Google Scholar] [CrossRef]
- Saravana, P.S.; Getachew, A.T.; Cho, Y.J.; Choi, J.H.; Park, Y.B.; Woo, H.C.; Chun, B.S. Influence of co-solvents on fucoxanthin and phlorotannin recovery from brown seaweed using supercritical CO2. J. Supercrit. Fluids 2017, 120, 295–303. [Google Scholar] [CrossRef]
- Bogolitsyn, K.G.; Kaplitsin, P.A.; Dobrodeeva, L.K.; Druzhinina, A.S.; Ovchinnikov, D.V.; Parshina, A.E.; Shulgina, E.V. Fatty Acid Composition and Biological Activity of Supercritical Extracts from Arctic Brown Algae Fucus vesiculosus. Russ. J. Phys. Chem. B 2017, 11, 1144–1152. [Google Scholar] [CrossRef]
- Fabrowska, J.; Messyasz, B.; Pankiewicz, R.; Wilińska, P.; Łęska, B. Seasonal differences in the content of phenols and pigments in thalli of freshwater Cladophora glomerata and its habitat. Water Res. 2018, 135, 66–74. [Google Scholar] [CrossRef]
- Tiwari, B.K. Ultrasound: A clean, green extraction technology. TrAC Trends Anal. Chem. 2015, 71, 100–109. [Google Scholar] [CrossRef]
- Adam, F.; Abert-Vian, M.; Peltier, G.; Chemat, F. “Solvent-free” ultrasound-assisted extraction of lipids from fresh microalgae cells: A green, clean and scalable process. Bioresour. Technol. 2012, 114, 457–465. [Google Scholar] [CrossRef]
- Chemat, F.; Rombaut, N.; Sicaire, A.G.; Meullemiestre, A.; Fabiano-Tixier, A.S.; Abert-Vian, M. Ultrasound assisted extraction of food and natural products. Mechanisms, techniques, combinations, protocols and applications. A review. Ultrason. Sonochem. 2017, 34, 540–560. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; Smyth, T.J.; O’Donnell, C.P. Optimization of ultrasound assisted extraction of bioactive components from brown seaweed Ascophyllum nodosum using response surface methodology. Ultrason. Sonochem. 2015, 23, 308–316. [Google Scholar] [CrossRef]
- Rodríguez-Bernaldo de Quirós, A.; Lage-Yusty, M.A.; López-Hernández, J. Determination of phenolic compounds in macroalgae for human consumption. Food Chem. 2010, 121, 634–638. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; Rajauria, G.; Tiwari, B.; Sweeney, T.; O’Doherty, J. Extraction and yield optimisation of fucose, glucans and associated antioxidant activities from laminaria digitata by applying response surface methodology to high intensity ultrasound-assisted extraction. Mar. Drugs 2018, 16. [Google Scholar] [CrossRef] [PubMed]
- Topuz, O.K.; Gokoglu, N.; Yerlikaya, P.; Ucak, I.; Gumus, B. Optimization of Antioxidant Activity and Phenolic Compound Extraction Conditions from Red Seaweed (Laurencia obtuse). J. Aquat. Food Prod. Technol. 2015, 25, 414–422. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; Ummat, V.; Tiwari, B.; Rajauria, G. Exploring ultrasound, microwave and ultrasound-microwave assisted extraction technologies to increase the extraction of bioactive compounds and antioxidants from brown macroalgae. Mar. Drugs 2020, 18, 172. [Google Scholar] [CrossRef]
- Simsek, M.; Sumnu, G.; Sahin, S. Microwave Assisted Extraction of Phenolic Compounds from Sour Cherry Pomace. Sep. Sci. Technol. 2012, 47, 1248–1254. [Google Scholar] [CrossRef]
- Zhang, H.F.; Yang, X.H.; Wang, Y. Microwave assisted extraction of secondary metabolites from plants: Current status and future directions. Trends Food Sci. Technol. 2011, 22, 672–688. [Google Scholar] [CrossRef]
- Mirzadeh, M.; Arianejad, M.R.; Khedmat, L. Antioxidant, antiradical, and antimicrobial activities of polysaccharides obtained by microwave-assisted extraction method: A review. Carbohydr. Polym. 2020, 229, 115421. [Google Scholar] [CrossRef]
- Boulho, R.; Marty, C.; Freile-Pelegrín, Y.; Robledo, D.; Bourgougnon, N.; Bedoux, G. Antiherpetic (HSV-1) activity of carrageenans from the red seaweed Solieria chordalis (Rhodophyta, Gigartinales) extracted by microwave-assisted extraction (MAE). J. Appl. Phycol. 2017, 29, 2219–2228. [Google Scholar] [CrossRef]
- Ren, B.; Chen, C.; Li, C.; Fu, X.; You, L.; Liu, R.H. Optimization of microwave-assisted extraction of Sargassum thunbergii polysaccharides and its antioxidant and hypoglycemic activities. Carbohydr. Polym. 2017, 173, 192–201. [Google Scholar] [CrossRef]
- Cao, C.; Huang, Q.; Zhang, B.; Li, C.; Fu, X. Physicochemical characterization and in vitro hypoglycemic activities of polysaccharides from Sargassum pallidum by microwave-assisted aqueous two-phase extraction. Int. J. Biol. Macromol. 2018, 109, 357–368. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Xue, Y.T. Optimization of microwave assisted extraction, chemical characterization and antitumor activities of polysaccharides from porphyra haitanensis. Carbohydr. Polym. 2019, 206, 179–186. [Google Scholar] [CrossRef]
- He, J.; Xu, Y.; Chen, H.; Sun, P. Extraction, structural characterization, and potential antioxidant activity of the polysaccharides from four seaweeds. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef]
- Le, B.; Golokhvast, K.S.; Yang, S.H.; Sun, S. Optimization of microwave-assisted extraction of polysaccharides from ulva pertusa and evaluation of their antioxidant activity. Antioxidants 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Tsubaki, S.; Oono, K.; Hiraoka, M.; Onda, A.; Mitani, T. Microwave-assisted hydrothermal extraction of sulfated polysaccharides from Ulva spp. and Monostroma latissimum. Food Chem. 2016, 210, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Safari, P.; Rezaei, M.; Shaviklo, A.R. The optimum conditions for the extraction of antioxidant compounds from the Persian gulf green algae (Chaetomorpha sp.) using response surface methodology. J. Food Sci. Technol. 2015, 52, 2974–2981. [Google Scholar] [CrossRef] [PubMed]
- López-Hortas, L.; Gannon, L.; Moreira, R.; Chenlo, F.; Domínguez, H.; Torres, M.D. Microwave hydrodiffusion and gravity (MHG) processing of Laminaria ochroleuca brown seaweed. J. Clean. Prod. 2018, 197, 1108–1116. [Google Scholar] [CrossRef]
- Zhang, R.; Yuen, A.K.L.; Magnusson, M.; Wright, J.T.; de Nys, R.; Masters, A.F.; Maschmeyer, T. A comparative assessment of the activity and structure of phlorotannins from the brown seaweed Carpophyllum flexuosum. Algal Res. 2018, 29, 130–141. [Google Scholar] [CrossRef]
- Abdelhamid, A.; Lajili, S.; Elkaibi, M.A.; Ben Salem, Y.; Abdelhamid, A.; Muller, C.D.; Majdoub, H.; Kraiem, J.; Bouraoui, A. Optimized Extraction, Preliminary Characterization and Evaluation of the in Vitro Anticancer Activity of Phlorotannin-Rich Fraction from the Brown Seaweed, Cystoseira sedoides. J. Aquat. Food Prod. Technol. 2019, 28, 892–909. [Google Scholar] [CrossRef]
- Xiao, X.; Si, X.; Yuan, Z.; Xu, X.; Li, G. Isolation of fucoxanthin from edible brown algae by microwave-assisted extraction coupled with high-speed countercurrent chromatography. J. Sep. Sci. 2012, 35, 2313–2317. [Google Scholar] [CrossRef]
- Rodriguez-Jasso, R.M.; Mussatto, S.I.; Pastrana, L.; Aguilar, C.N.; Teixeira, J.A. Microwave-assisted extraction of sulfated polysaccharides (fucoidan) from brown seaweed. Carbohydr. Polym. 2011, 86, 1137–1144. [Google Scholar] [CrossRef]
- Lorbeer, A.J.; Lahnstein, J.; Fincher, G.B.; Su, P.; Zhang, W. Kinetics of conventional and microwave-assisted fucoidan extractions from the brown alga, Ecklonia radiata. J. Appl. Phycol. 2015, 27, 2079–2087. [Google Scholar] [CrossRef]
- Zhong-rui Li Influence of preparation methods on the yield, components and antioxidant activities of polysaccharides from Palmaria palmata. African J. Pharm. Pharmacol. 2012, 6, 1157–1165. [CrossRef]
- Fabrowska, J.; Messyasz, B.; Szyling, J.; Walkowiak, J.; Łęska, B. Isolation of chlorophylls and carotenoids from freshwater algae using different extraction methods. Phycol. Res. 2018, 66, 52–57. [Google Scholar] [CrossRef]
- Puri, M.; Sharma, D.; Barrow, C.J. Enzyme-assisted extraction of bioactives from plants. Trends Biotechnol. 2012, 30, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Sarker, M.Z.I.; Ghafoor, K.; Happy, R.A.; Ferdosh, S. Bioactive Compounds and Extraction Techniques. In Recovering Bioactive Compounds from Agricultural Wastes; Nguyen, V.T., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 33–53. [Google Scholar]
- Casas, M.P.; Conde, E.; Domínguez, H.; Moure, A. Ecofriendly extraction of bioactive fractions from Sargassum muticum. Process Biochem. 2019, 79, 166–173. [Google Scholar] [CrossRef]
- Rodrigues, D.; Sousa, S.; Silva, A.; Amorim, M.; Pereira, L.; Rocha-Santos, T.A.P.; Gomes, A.M.P.; Duarte, A.C.; Freitas, A.C. Impact of enzyme- and ultrasound-assisted extraction methods on biological properties of red, brown, and green seaweeds from the Central West Coast of Portugal. J. Agric. Food Chem. 2015, 63, 3177–3188. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.; Sánchez-Camargo, A.d.P.; Cifuentes, A.; Ibáñez, E. Plants, seaweeds, microalgae and food by-products as natural sources of functional ingredients obtained using pressurized liquid extraction and supercritical fluid extraction. TrAC Trends Anal. Chem. 2015, 71, 26–38. [Google Scholar] [CrossRef]
- Harrysson, H.; Hayes, M.; Eimer, F.; Carlsson, N.G.; Toth, G.B.; Undeland, I. Production of protein extracts from Swedish red, green, and brown seaweeds, Porphyra umbilicalis Kützing, Ulva lactuca Linnaeus, and Saccharina latissima (Linnaeus) J. V. Lamouroux using three different methods. J. Appl. Phycol. 2018, 30, 3565–3580. [Google Scholar] [CrossRef]
- Sun, H.; Ge, X.; Lv, Y.; Wang, A. Application of accelerated solvent extraction in the analysis of organic contaminants, bioactive and nutritional compounds in food and feed. J. Chromatogr. A 2012, 1237, 1–23. [Google Scholar] [CrossRef]
- Tierney, M.S.; Smyth, T.J.; Hayes, M.; Soler-Vila, A.; Croft, A.K.; Brunton, N. Influence of pressurised liquid extraction and solid-liquid extraction methods on the phenolic content and antioxidant activities of Irish macroalgae. Int. J. Food Sci. Technol. 2013, 48, 860–869. [Google Scholar] [CrossRef]
- Fayad, S.; Nehmé, R.; Tannoury, M.; Lesellier, E.; Pichon, C.; Morin, P. Macroalga Padina pavonica water extracts obtained by pressurized liquid extraction and microwave-assisted extraction inhibit hyaluronidase activity as shown by capillary electrophoresis. J. Chromatogr. A 2017, 1497, 19–27. [Google Scholar] [CrossRef]
- Montero, L.; Sánchez-Camargo, A.P.; García-Cañas, V.; Tanniou, A.; Stiger-Pouvreau, V.; Russo, M.; Rastrelli, L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Anti-proliferative activity and chemical characterization by comprehensive two-dimensional liquid chromatography coupled to mass spectrometry of phlorotannins from the brown macroalga Sargassum muticum collected on North-Atlantic coasts. J. Chromatogr. A 2016, 1428, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Anaëlle, T.; Serrano Leon, E.; Laurent, V.; Elena, I.; Mendiola, J.A.; Stéphane, C.; Nelly, K.; Stéphane, L.B.; Luc, M.; Valérie, S.P. Green improved processes to extract bioactive phenolic compounds from brown macroalgae using Sargassum muticum as model. Talanta 2013, 104, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Carullo, D.; Abera, B.D.; Casazza, A.A.; Donsì, F.; Perego, P.; Ferrari, G.; Pataro, G. Effect of pulsed electric fields and high pressure homogenization on the aqueous extraction of intracellular compounds from the microalgae Chlorella vulgaris. Algal Res. 2018, 31, 60–69. [Google Scholar] [CrossRef]
- La, H.J.; Choi, G.G.; Cho, C.; Seo, S.H.; Srivastava, A.; Jo, B.H.; Lee, J.Y.; Jin, Y.S.; Oh, H.M. Increased lipid productivity of Acutodesmus dimorphus using optimized pulsed electric field. J. Appl. Phycol. 2016, 28, 931–938. [Google Scholar] [CrossRef]
- Polikovsky, M.; Fernand, F.; Sack, M.; Frey, W.; Müller, G.; Golberg, A. Towards marine biorefineries: Selective proteins extractions from marine macroalgae Ulva with pulsed electric fields. Innov. Food Sci. Emerg. Technol. 2016, 37, 194–200. [Google Scholar] [CrossRef]
- Gupta, S.; Abu-Ghannam, N. Recent developments in the application of seaweeds or seaweed extracts as a means for enhancing the safety and quality attributes of foods. Innov. Food Sci. Emerg. Technol. 2011, 12, 600–609. [Google Scholar] [CrossRef]
- Sofi, S.A.; Singh, J.; Rafiq, S.; Ashraf, U.; Dar, B.N.; Nayik, G.A. A Comprehensive Review on Antimicrobial Packaging and its Use in Food Packaging. Curr. Nutr. Food Sci. 2018, 14, 305–312. [Google Scholar] [CrossRef]
- Poveda-castillo, G.C.; Rodrigo, D.; Mart, A.; Consuelo, M. Bioactivity of Fucoidan as an Antimicrobial Agent in a New Functional Beverage. Beverages 2018, 4, 64. [Google Scholar] [CrossRef]
- Øverland, M.; Mydland, L.T.; Skrede, A. Marine macroalgae as a source of protein and bioactive compounds in feed for monogastric animals. J. Sci. Food Agric. 2019, 99, 13–24. [Google Scholar] [CrossRef]
- Mohan, K.; Ravichandran, S.; Muralisankar, T.; Uthayakumar, V.; Chandirasekar, R.; Seedevi, P.; Abirami, R.G.; Rajan, D.K. Application of marine-derived polysaccharides as immunostimulants in aquaculture: A review of current knowledge and further perspectives. Fish Shellfish Immunol. 2019, 86, 1177–1193. [Google Scholar] [CrossRef]
- Madhiyazhagan, P.; Murugan, K.; Kumar, A.N.; Nataraj, T.; Dinesh, D.; Panneerselvam, C.; Subramaniam, J.; Kumar, P.M.; Suresh, U.; Roni, M.; et al. Sargassum muticum synthesized silver nanoparticles: An effective control tool against mosquito vectors and bacterial pathogens. Parasitol. Res. 2015, 114, 4305–4317. [Google Scholar] [CrossRef]
- Pugazhendhi, A.; Prabakar, D.; Jacob, J.M.; Karuppusamy, I.; Saratale, R.G. Synthesis and characterization of silver nanoparticles using Gelidium amansii and its antimicrobial property against various pathogenic bacteria. Microb. Pathog. 2018, 114, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Senthamil Selvi, S.; Govindaraju, M. Seaweed-mediated biosynthesis of silver nanoparticles using Gracilaria corticata for its antifungal activity against Candida spp. Appl. Nanosci. 2013, 3, 495–500. [Google Scholar] [CrossRef]
- de Aragão, A.P.; de Oliveira, T.M.; Quelemes, P.V.; Perfeito, M.L.G.; Araújo, M.C.; Santiago, J.d.A.S.; Cardoso, V.S.; Quaresma, P.; de Souza de Almeida Leite, J.R.; da Silva, D.A. Green synthesis of silver nanoparticles using the seaweed Gracilaria birdiae and their antibacterial activity. Arab. J. Chem. 2019, 12, 4182–4188. [Google Scholar] [CrossRef]
- Selvaraj, P.; Neethu, E.; Rathika, P.; Jayaseeli, J.P.R.; Jermy, B.R.; AbdulAzeez, S.; Borgio, J.F.; Dhas, T.S. Antibacterial potentials of methanolic extract and silver nanoparticles from marine algae. Biocatal. Agric. Biotechnol. 2020, 28, 101719. [Google Scholar] [CrossRef]
- Fatima, R.; Priya, M.; Indurthi, L.; Radhakrishnan, V.; Sudhakaran, R. Biosynthesis of silver nanoparticles using red algae Portieria hornemannii and its antibacterial activity against fish pathogens. Microb. Pathog. 2020, 138, 103780. [Google Scholar] [CrossRef] [PubMed]
- El-Rafie, H.M.; El-Rafie, M.H.; Zahran, M.K. Green synthesis of silver nanoparticles using polysaccharides extracted from marine macro algae. Carbohydr. Polym. 2013, 96, 403–410. [Google Scholar] [CrossRef]
- Stalin Dhas, T.; Ganesh Kumar, V.; Karthick, V.; Jini Angel, K.; Govindaraju, K. Facile synthesis of silver chloride nanoparticles using marine alga and its antibacterial efficacy. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2014, 120, 416–420. [Google Scholar] [CrossRef]
- Arockiya Aarthi Rajathi, F.; Parthiban, C.; Ganesh Kumar, V.; Anantharaman, P. Biosynthesis of antibacterial gold nanoparticles using brown alga, Stoechospermum marginatum (kützing). Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2012, 99, 166–173. [Google Scholar] [CrossRef]
- Dhas, T.S.; Sowmiya, P.; Kumar, V.G.; Ravi, M.; Suthindhiran, K.; Borgio, J.F.; Narendrakumar, G.; Kumar, V.R.; Karthick, V.; Kumar, C.M.V. Antimicrobial effect of Sargassum plagiophyllum mediated gold nanoparticles on Escherichia coli and Salmonella typhi. Biocatal. Agric. Biotechnol. 2020, 26, 101627. [Google Scholar] [CrossRef]
- Salem, D.M.S.A.; Ismail, M.M.; Aly-Eldeen, M.A. Biogenic synthesis and antimicrobial potency of iron oxide (Fe3O4) nanoparticles using algae harvested from the Mediterranean Sea, Egypt. Egypt. J. Aquat. Res. 2019, 45, 197–204. [Google Scholar] [CrossRef]
- Mashjoor, S.; Yousefzadi, M.; Zolgharnain, H.; Kamrani, E.; Alishahi, M. Organic and inorganic nano-Fe3O4: Alga Ulva flexuosa-based synthesis, antimicrobial effects and acute toxicity to briny water rotifer Brachionus rotundiformis. Environ. Pollut. 2018, 237, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Kumaresan, M.; Vijai Anand, K.; Govindaraju, K.; Tamilselvan, S.; Ganesh Kumar, V. Seaweed Sargassum wightii mediated preparation of zirconia (ZrO 2 ) nanoparticles and their antibacterial activity against gram positive and gram negative bacteria. Microb. Pathog. 2018, 124, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Pugazhendhi, A.; Prabhu, R.; Muruganantham, K.; Shanmuganathan, R.; Natarajan, S. Anticancer, antimicrobial and photocatalytic activities of green synthesized magnesium oxide nanoparticles (MgONPs) using aqueous extract of Sargassum wightii. J. Photochem. Photobiol. B Biol. 2019, 190, 86–97. [Google Scholar] [CrossRef]
- Nagarajan, S.; Arumugam Kuppusamy, K. Extracellular synthesis of zinc oxide nanoparticle using seaweeds of gulf of Mannar, India. J. Nanobiotechnol. 2013, 11, 1–11. [Google Scholar] [CrossRef]
- Dang, H.; Lovell, C.R. Microbial Surface Colonization and Biofilm Development in Marine Environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef]
- Batista, D.; Carvalho, A.P.; Costa, R.; Coutinho, R.; Dobretsov, S. Extracts of macroalgae from the Brazilian coast inhibit bacterial quorum sensing. Bot. Mar. 2014, 57, 441–447. [Google Scholar] [CrossRef]
- Ying-ying, S.; Hui, W.; Gan-lin, G.; Yin-fang, P. Green alga Ulva pertusa — a new source of bioactive compounds with antialgal activity. Environ. Sci. Pollut. Res. 2015, 22, 10351–10359. [Google Scholar] [CrossRef]
- Nakajima, N.; Sugimoto, N.; Ohki, K.; Kamiya, M. Diversity of phlorotannin profiles among sargassasacean species affecting variation and abundance of epiphytes. Eur. J. Phycol. 2016, 51, 307–316. [Google Scholar] [CrossRef]
- Dahms, H.U.; Dobretsov, S. Antifouling Compounds from Marine Macroalgae. Mar. Drugs 2017, 15, 265. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Batista, D.; Dobretsov, S.; Coutinho, R. Extracts of seaweeds as potential inhibitors of quorum sensing and bacterial growth. J. Appl. Phycol. 2017, 29, 789–797. [Google Scholar] [CrossRef]
- Sethupathy, S.; Shanmuganathan, B.; Kasi, P.D.; Karutha Pandian, S. Alpha-bisabolol from brown macroalga Padina gymnospora mitigates biofilm formation and quorum sensing controlled virulence factor production in Serratia marcescens. J. Appl. Phycol. 2016, 28, 1987–1996. [Google Scholar] [CrossRef]
- Du, Y.; Sun, J.; Gong, Q.; Wang, Y.; Fu, P.; Zhu, W. New α-Pyridones with Quorum-Sensing Inhibitory Activity from Diversity-Enhanced Extracts of a Streptomyces sp. Derived from Marine Algae. J. Agric. Food Chem. 2018, 66, 1807–1812. [Google Scholar] [CrossRef] [PubMed]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Khanna, P.; Kaur, A.; Goyal, D. Algae-based metallic nanoparticles: Synthesis, characterization and applications. J. Microbiol. Methods 2019, 163, 105656. [Google Scholar] [CrossRef] [PubMed]
- Sri, V.; Prakash, S.; Ramasubburayan, R. Enzyme and Microbial Technology Seaweeds: A resource for marine bionanotechnology. Enzyme Microb. Technol. 2016, 95, 45–57. [Google Scholar] [CrossRef]
- Sudhakar, M.S.; Aggarwal, A.; Sah, M.K. Engineering biomaterials for the bioremediation: Advances in nanotechnological approaches for heavy metals removal from natural resources. In Emerging Technologies in Environmental Bioremediation; Elsevier Inc.: Amsterdam, The Netherlands, 2020; pp. 323–340. ISBN 9780128198605. [Google Scholar]
- Gómez-Ordóñez, E.; Jiménez-Escrig, A.; Rupérez, P. Dietary fibre and physicochemical properties of several edible seaweeds from the northwestern Spanish coast. Food Res. Int. 2010, 43, 2289–2294. [Google Scholar] [CrossRef]
- Olszewska, M.A.; Gędas, A.; Simões, M. Antimicrobial polyphenol-rich extracts: Applications and limitations in the food industry. Food Res. Int. 2020, 134, 109214. [Google Scholar] [CrossRef]
- Lourenço-Lopes, C.; Garcia-Oliveira, P.; Carpena, M.; Fraga-Corral, M.; Jimenez-Lopez, C.; Pereira, A.G.; Prieto, M.A.; Simal-Gándara, J. Scientific Approaches on Extraction, Purification and Stability for the Commercialization of Fucoxanthin Recovered from Brown Algae. Foods 2020, 9, 1113. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Alga Species | Conditions | Bacteria Tested | Main Outcomes | Ref. |
|---|---|---|---|---|
| L. brandenii—(India) | HAE, MeOH: CHCl3 (6:4), 35 °C, 120 rpm, 7 days | S. aureus/B. subtilis/M.luteus/R. rhodochrous/E. coli/P.aeruginosa/Vibrio cholerae/Salmonella typhi/Streptococcus pneumoniae | All organisms were inhibited. High activity against B. subtilis whereas it was moderate against E. coli | [35] |
| G. ornata—(Brazil) | HAE, H2O, 25 °C, 24 h | B. subtilis/S. aureus/E. aerogens/E. coli/P. aeruginosa/S. choleraesuis/S. typhi | Only exhibits inhibition to E. coli | [36] |
| C. rubrum—(Chile) | HAE (3x), 96% EtOH, 24 h. | S. parasitica/Y. ruckeri | Antibacterial and antifungal activities against fish pathogens | [37] |
| G. changii—(Malaysia) | Solid liquid extraction, MeOH, 4 days. | P. aeruginosa | Minimal inhibitory concentration (MIC) 6. 25 mg/mL | [38] |
| P. capillacea/O. obtusiloba—(Brazil) | Solid liquid extraction, cold Hx and 70% EtOH. | E. coli/S. aureus/Salmonella sp./V. harveyi | No antimicrobial activity. | [39] |
| P. gymnospora—(Brazil) | Percolation with MeOH | S. aureus | MIC 500 µg/mL | [40] |
| S. latifolium/S. platycarpum/C. socialis—(Arabian Gulf) | HAE extraction, MeOH and AcO 25 °C, 150 rpm, 7 days | S. aureus/S. xylosus/MRSA/E. faecalis/B. subtilis/E. coli/P. aeruginosa/Salmonella sp./K. pneumoniae/C. albicans | Higher activity against Gram positive bacteria than Gram negative | [41] |
| L. japonica—(Korea, Japan, China) | HAE (x3), EtOH, 25 °C, 1 day. | S. mutans/S. sobrinus/A. naeslundii/A. odontolyticus/A. actinomycetemcomitas/F. nucleatum/P.gingivalis | Inhibitory activity against all microorganisms | [42] |
| D. membranacea—(Mediterranean Sea) | Column extraction, EtOH, AcO and MeOH/DCM | S. aureus/S. agalactiae/B. subtilis/E. faecium/E. faecalis/E. coli/C. albicans | EtOH and AcO showed higher antimicrobial activity | [43] |
| S. oligocystum—(Persian Gulf) | HAE, hot and cold H2O and glycerin | S. aureus/S. epidermidis/P. aeruginosa/E. coli | Hot water extract exhibited activity against S. aureus, S. epidermidis, and P. aeruginosa | [44] |
| C. myrica/C. trinodis/P. gymnospora/S. dentifolium/S. hystrix/A. fragilis/C. racemosa/C. fragile—(Red sea) | HAE, MeOH, 25 °C, 50 rpm, 7 days. | E. coli/S. aureus/E. faecalis/Salmonella sp./B. cereus/P. aeruginosa. | MeOH extracts P. gymnospora and C. fragile showed the highest activities | [45] |
| S. polycystum/P. australis—(Malasya) | HAE, Hx, DCM, MeOH, 72 h | S. aureus/B. cereus/E. coli/E. coli/P. aeruginosa | S. polycystum extracts exhibited higher bacteriostatic activity | [46] |
| Gracilaria sp.—(Malaysia farmed algae) | HAE, MeOH, 48 h | B. subtilis/S. aureus/S. epidermidis/E. coli/V. cholera/E. cloacae. | Moderate antibacterial activity but S. aureus, S epidermidis, E. cloacae were not inhibited | [47] |
| P. antillarum/P. boergesenii/U. flexuosa—(Persian Gulf) | HAE, EtOAc, MeOH; 48 h | B. subtilis/B. pumulis/E. faecalis/S. aureus/S. epidermidis/E. coli/K. pneumoniae/P. aeruginosa/A. niger/C. albicans/S. cerevisiae | A. niger, P. aeruginosa were not inhibited. P. antillarum extracts do not have effect in K. pneumoniae | [48] |
| U. lactuca/U. intestinales/C. vagabunda—(Black sea) | HAE, EtOH 70% | S. aureus/E. coli | Antibacterial activity was higher in E. coli | [49] |
| U. rigida—(Turkey) | HAE, DCM; then DCM/MeOH | S. agalactiae./S. aureus./E. faecalis/Micrococcus sp./V. tapetis/V. anguillarum/V. alginolyticus E. coli/P. cepacia/P. fluorescens/P. aeruginosa/A. salmonicida/A. hydrophila/S. typhimurium | No significant variation with seasons. The most sensitive bacteria were A. salmonicida, S. typhimurium, S. agalactiae, A. hydrophila, P. cepacia, S. aureus and E. faecalis | [50] |
| G. doryphora—(Egypt) | HAE, MeOH, EtOH, EtOAc, 72 h, 150 rpm. | B. subtilis/E. faecalis/S. aureus/E. coli/P. aeruginosa/C. albicans | Inhibitory effects except against E. coli | [51] |
| E. prolifera/U. reticulata/C. myrica/P. pavonica/T. triquetra/S. portieriatum/G. multipartita—(Saudi Arabia) | HAE, PeEt, DEt, EtOAc, MeOH, 30 °C, 120 rpm, 24 h | B. subtilis/MRSA/S aureus/E. coli/K. pneumoniae/P. aeruginosa | P. pavonica and T. triquetra extracts showed better activity. In some cases, inhibitory effects changed with seasons | [52] |
| G. multipartita/U. reticulata/S. marginatum—(Turkey) | HAE, Hx, DCM, MeOH, 72 h | B. subtilis/MRSA/S. aureus/E. coli/K. pneumoniae/P. aeruginosa | B. subtilis, MRSA, and E. coli were susceptible | [53] |
| L. obtusa/C. elongatum/C. multifida—(Adriact sea) | Soxhlet extraction, AcO | B. mycoides/B. subtilis/S. aureus/E. coli/K. pneumoniae/A. flavus/A. fumigatus/C. albicans/P. verrucosum | All the tested extracts showed antimicrobial activity | [54] |
| C. iyengarii/S. asperum/S. marginatum/C. indica/S. variegatum/S. swartzii/S. tenerrimum/S. ilicifolium/I. stellata/S. robusta/H. tuna/R. implexum/D. dichotoma var. intricata/D. indica/M. afaqhusainii/J. laminarioides—(Pakistan) | HAE, EtOH, 1 week | B. subtilis/S. aureus/E. coli/S. typhi/P. aeruginosa/R. solani/Macrophomina phaseolina/F. solani/F. oxysporum | Brown species have shown more potential than red algal species. The highest antibacterial activity was found in EtOH extract of D. dichotoma var intricata and D. indica against S. typhimurium | [55] |
| C. linum/C. rupestris/G. dura/G. gracilis/G. longissima/U. prolifera—(Italy) | Soxhlet extraction, CHCl3/MeOH, 60 °C, 24 h | V. ordalii/V. salmonicida V. alginolyticus/V. splendidus/V. harveyi/V. vulnificus | Different susceptibilities to algal extracts were detected. G. longissima was the most effective | [56] |
| C. rupestris—(Mediterranean Sea) | HAE, MeOH, CHCl3, H2O | Enterococcus sp./S. agalactiae/V. cholerae | Results showed seasonal variety | [57] |
| G. longissima—(Mediterranean Sea) | Soxhlet extraction, CHCl3/MeOH (2:1), 60 °C, 24 h | P. aeruginosa/Enterococcus sp./S. agalactiae/V. salmonicida/V. fluvialis/V. vulnificus/V. cholerae/V. alginolyticus/C. albicans/C. famata/C. glabrata | Moderate antimicrobial effect except on V. salmonicida and fungal species | [58] |
| C. antemmina/C. peltata/C. scalpelliformis/D. dichotoma/S. marginatum/A. specifera/G. lithophilia/G. corticata—(India) | HAE, MeOH | E. coli/P. aeruginosa/S. aureus/K. pneumoniae | G. lithophila presents the most promising results | [59] |
| J. rubens/C. elongata/P. capillacea/U. fasciata/U. lactuca/E. compressa/E. linza/S. vulgare/C. sinuosa—(Egypt) | HAE, EtOH 70%, MeOH 70% AcO 70%, 150 rpm, 72 h | B. subtilis/S. aureus/E. coli/S. typhi/K. pneumoniae/C. albicans. | In all the tests, AcO showed the biggest inhibition halos | [60] |
| D. flabellata/P. concrescens/L. johnstonii/G. martinensis/U. lactuca/C. fragile—(Mexico) | HAE, AcO:MeOH | E. coli/S. aureus/B. cereus/B. subtilis/S. epidermidis | L. johnstonii, D. flabellata and U. lactuca presented activity against pathogenic bacteria tested | [61] |
| E. bicyclis—(South Korea) | HAE, MeOH, 70 °C, 3 h | C. acnes/S. aureus/S. epidermidis/P. aeruginosa | Inhibitory effects except against P. aeruginosa | [62] |
| C. trinodis—(Persian Gulf)—(Persian Gulf) | HAE, DEt:EtOH:Hx | S. aureus/S. epidermidis/E. coli/P. aeruginosa | The best Inhibitory effect was against S. epidermidis was the worst against P. aeruginosa | [63] |
| C. glomerata,/E. linza/U. rigida/C. barbata/P. pavonica/C. ciliatum/C. officinalis—(Black sea Turkey) | HAE, 95% EtOH | S. aureus/B. cereus/A. niger/S typhimurium/L. monocytogenes/E. coli/C. albicans/P. aeruginosa | All alga extracts present antimicrobial activity | [64] |
| S. vulgare/C. hirsutus/R. verruculosa—(Coast of Algeria) | Soxhlet extraction, MeOH, MeOH: CHCl3, 6h | B. cereus/S. aureus/M. luteus/P. aeruginosa/E. coli/K. pneumoniae/C. albicans | Positive antimicrobial results against S. aureus and B. cereus | [65] |
| Laurencia ssp. (aldingensis/catarinensis/dendroidea/intricata/translucida) –(Brazil) | HAE, Hx, CHCl3, MeOH, H2O | C. albicans/C. parapsilosis/C. neoformans | L. aldingensis showed the best antifungal effects | [66] |
| D. membranacea—(Tunisia) | HAE, H2O, CHCl3, EtOAc | S. aureus/S. epidermidis/L. monocytogenes/M. luteus/E. faecium/E. coli/P. aeruginosa/S. typhimurium/C. albicans/C. kefyr/C. krusei/C. dubliniensis/C. glabrata | Inhibitory effects against M. luteus, S. aureus, S. epidermidis, L. monocytogenes, C. krusei, C. dubliniensis and C. kefyr | [67] |
| S. wightii/C. linum/P. gymnospora—(India) | HAE, Hx, EtOAc, AcO, MeOH | P. aeruginosa/S. typhi/E. amylovora/E. aerogens/P. vulgaris/K. pneumonia/E. coli/MRSA/B. subtilis/E. faecalis | EtOAc and AcO extracts were more efficient, but no inhibitory effects were observed against S. paratyphi and K. pneumonia. | [68] |
| Fucus spp./P. elongata/Rhodomela confervoides/S. latissima./C. rupestris/D. contorta/F. vesiculosus/C. rubrum/M. stellatus/L. digitata—(Germany) | HAE, DCM | E. amylovora/E. coli/P. aeruginosa/B. subtilis/S. lentus | The macroalgae presented antibacterial activity against at least one of the test strains | [69] |
| H. tuna/C. barbata/C. bursa—(Montenegro) | HAE, DCM:MeOH, 48 h | E. coli/S. aureus/B. subtilis/E. faecalis/C. albicans | C. barbata demonstrated as having the best antimicrobial activity for S. aureus and B. subtilis | [70] |
| G. corticata/G. edulis—(India) | HAE, DMSO, 70% MeOH, 130 rpm, 16 h | E. coli/Photobacterium sp./P. fluorescens/S. aureus/B. subtilis | MeOH and DMSO extracts inhibited B. subtilis | [71] |
| L. digitata/S. latissima/H. elongata/P. palmata/C. crispus—(Ireland) | HAE, MeOH, EtOH, AcO, 2 h | L. monocytogenes/S. abony/E. faecalis/P. aeruginosa | The extraction of antimicrobials from macroalgae were solvent dependent | [72] |
| S. marginatum—(India) | HAE, DCM, EtOAc, AcO, MeOH | Candida spp. | Low antifungal properties. | [73] |
| S. lomentaria/P. pavonica/C.mediterranea/H. musciformis/S. filamentosa—(Turkey) | HAE, MeOH, 8 h, 200 rpm | S. aureus/S. typhimurium/E. coli/E. faecalis/C. albicans | S. lomentaria inhibited S. typhimurium. C. mediterrranea inhibited C. albicans | [74] |
| U. lactuca/E. intestinalis—(Adriatic coast of Montenegro) | HAE, Hx, DCM, MeOH, 72 h | B. mycoides/B. subtilis/E. coli/K. pneumoniae/S. aureus/A. flavus/A. fumigatus/C. albicans/P. purpurascens/P. verrucosum | Inhibitory effects were observed against B. mycoides and B. subtilis | [75] |
| A. fragilis/C.a myrica/H. cuneiformes/L. papillosa/S. cinereum/T turbinata—(Egypt) | HAE, 80% MeOH, 25 °C | B. subtilis/S. aureus/E. coli/C. albicans | H. cuneiformis extract showed stronger activity | [76] |
| E.cava—(Korea) | HAE, EtOH, n-Hx, DCM, EtOAc, n-BuOH, H2O | S. aureus/MRSA/S. typhi/S. enteritidis/S. gallinarum | EtOH had antibacterial activity S. aureus, MRSA and Salmonella spp. | [77] |
| C. barbata—(Red Sea, Egypt) | Soxhlet extraction, EtOH | B. subtilis/S. aureus/M. luteus/E. coli/P. aeruginosa/Serratia. marcescens/S. typhi/Vibrio sp./A. hydrophila/C. albicans | Inhibitory activity except against M. luteus | [78] |
| K. alvarezii—(Malaysia) | HAE, EtOH, H2O | E. coli/B. cereus | B. cereus was inhibited but no E. coli. | [79] |
| U. lactuca/D. dichotoma/P. gymnospora/S. vulgare/H. musciformis/D. simplex—(Brazil) | HAE, DCM, MeOH, EtOH, H2O | T. rubrum/T. tonsurans/T. mentagrophytes/M. canis/M. gypseum/E. flocossum/C. albicans/C. krusei/C. guilliermondi/C. parapsilosis/ | EtOH and MeOH extracts were the most effective | [80] |
| B. bifurcata—(Portugal) | HAE, MeOH, DCM, 12 h | E. coli/P. aeruginosa/B. subtilis/S. aureus/S. cerevisiae | MeOH extracts had inhibitory effects in all the microorganisms | [81] |
| H. flagelliformis/C. myrica/S. boveanum—(Persian Gulf) | HAE, DCM, 48 h | E. coli:/K. pneumonia/S. typhi/S. aureus/S. epidemidis/B. subtilis/A. niger/C. albicans | The antimicrobial activity was solvent-dependent | [82] |
| T. conoides—(India) | HAE, n-Hx, MeOH and EtOH: H2O, 72 h. | S. aureus/S. epidermidis/E. coli/P. aeruginosa/A. niger/C. albicans | MeOH and EtOH: H2O extracts were the most effective against the microorganisms studied | [83] |
| D. dichotoma/P. pavonica/S. vulgare—(Adriatic Sea) | HAE, AcO, 50 °C;4 h | B. mycoides/B. subtilis/S. aureus/E. coli/K. pneumoniae/A. flavus/A. fumigatus/C. albicans/P. purpurescens/P. verrucosum | All crude extracts have a statistically significant inhibitory effect on microbial growth | [84] |
| C. racemosa/C. sertularioides/K. alvarezii—(Malaysian coast) | HAE, Hx, CHCl3, EtOAc, EtOH, MeOH, H2O, 1 day | B. cereus/S. aureus/A. baumannii/E. coli/K. pneumoniae/P. aeruginosa/C. albicans/C. parapsilosis/C. krusei/C. neoformans/A. fumigatus/T. interdigitale | Inhibitory effects except against A. fumigatus | [85] |
| U. lactuca—(Gulf of Maine) | HAE, MeOH, 70 °C | S. aureus/S. epidermidis | Inhibitory effects against both species | [86] |
| T. ornata/T. decurrens/T. conoides/S. polycystum/S. incisifolium/S. ilicifolium/H.a cuneiformis—(Madagascar) | HAE, MeOH, EtOAc | B. cereus/S. aureus/S. pneumoniae/E. cloacae/K. oxytoca/S. boydii/E. coli/S. enteridis/P. aeruginosa/C. albicans/C. membranaefaciens/C. neoformans/T. mucoides | Antimicrobial tests of the crude extracts revealed a strong activity against S. aureus and S. pneumoniae | [87] |
| A. specifera/Cladophoropsis sp./L. paniculata/Tydemania sp./U. prolifera | Soxhlet extraction, EtOH and PeEt, 24 h | C. albicans/A. niger/Mucor sp./Paeciliomyces sp. | EtOH extract of L. paniculata showed the best antimicrobial activity | [88] |
| H. esperi/C. prolifera—(Egypt) | Soxhlet extraction, MeOH, 40 °C, 24 h | E. coli/P. aeruginosa/S. typhimurium/A. hydrophila/B. subtilis/S. aureus | Inhibitory effects against B. subtilis and S. aureus growth but no against P. aeruginosa and S. typhimurium, | [89] |
| Grateloupia sp./G. corticata/Halymenia sp./Metamastophora sp./Spyridia sp. | HAE, MeOH, 24 h | E. cloacae/K. oxytoca/E. coli/S. enteridis/B. cereus/S. aureus/S. pneumoniae/C. albicans. | All the crude extracts obtained can inhibit microbe’s growth. | [90] |
| H. elongata—(Ireland) | HAE, H2O, MeOH, 40 °C, 100 rpm, 2 h | L. monocytogenes/S. abony/E. faecalis/P. aeruginosa | 60% MeOH extract showed the best results. | [91] |
| F. serratus/F. vesiculosus—(Ireland) | HAE, H2O, MeOH, EtOAc, AcO | MRSA 28 strains | Both species present antibacterial activity against several MRSA strains. | [92] |
| U. reticulata—(Vietnam) | HAE, MeOH:CHCl3: H2O | B. cereus/S. faecalis/E. cloace/S. aureus/E. coli/P. aeruginosa/V. haveyi | U. reticulata showed high antimicrobial activity, against E. cloace and against E. coli. | [93] |
| U. rigida—(Tunisia) | HAE, EtOH:H2O, 48 | B. subtilis/B. cerus/S. aureus/S. epidermis/E. faecalis/L. monocytogenes/E. coli/P. aeruginosa/K. pneumoniae/A. niger/F. graminearum/F. culmorum/F. oxysporum/C. albicans | Antimicrobial activity varied depending on the season | [94] |
| U. fasciata/G. salicornia—(Honolulu, USA) | HAE, EtOH | E. faecalis/V. alginolyticus/V. cholerae/S. aureus/S. typhimurium/E. coli | U. fasciata had significantly higher antimicrobial activity compared to G. salicornia | [95] |
| U. lactuca/D. dichotoma/C. elongata—(Algeria) | HAE, MeOH, DEt, CHCl3 | E. coli/S. aureus/Salmonella/C. albicans/Penicillium sp. | CHCl3 extracts of U. lactuca and C. elongata had the highest activity against E. coli and Salmonella sp. MeOH of all species showed antifungal activity for C. albicans. | [96] |
| Type | Compounds | Macroalgae | Antimicrobial Activity | Ref. |
|---|---|---|---|---|
| Polyphenols | Phlorotannins | F. vesiculosus | Alteration of the cell membrane and cell destruction of S. aureus, S. pneumonia and P. aeruginosa | [129] |
| Phlorotannins | S. thunbergii | Alteration of the cell membrane, cytoplasm’s leakage and cell destruction of V. parahaemolyticus | [130] | |
| Phlorofucofuroeckol | E. bicyclis | Cell membrane damage and suppression of genes related to methicillin resistance in S. aureus | [131] | |
| Bromophenols | K. alvarezii | Downregulation of pathogenic genes of P. gingivalis | [132] | |
| Dieckol | E. clava | Alteration of cell integrity and metabolism of T. rubrum | [133] | |
| Phlorotannins | C. nodicaulis, C. usneoides, F. spiralis | Alterations of the cell wall composition, increased mitochondrial respiration. Inhibition of the formation of the germ tube of C. albicans | [134] | |
| Phlorotannins | E. clava | Inhibition of the enzyme neuraminidase of the Influenza A virus | [135] | |
| Polyphenolic rich extracts | E. arborea, S. filiformis | Inhibition of the viral particle | [136] | |
| Polysaccharides | Depolymerized fucoidans | L. japonica | Interaction with protein of the cell membrane and cellular rupture of E. coli and S. aureus | [137] |
| Fucoidan | F. vesiculosus | Inhibition of dental plaque bacteria and foodborne pathogens. | [138] | |
| Laminarin rich extracts | A. nodosum, L. hyperborea | Inhibition of S. aureus, L. monocytogenes, E. coli and S. typhimurium. | [139] | |
| Water soluble polysaccharide extracts | P. capillacae, D. membranacea | Inhibition of F. oxysporium Inhibition of C. albicans and M. phaseli | [140] | |
| Sulfated polysaccharides | G. skottbergii | Obstruction of herpes simplex virus type 1 and 2 attachment to the cells | [141] | |
| Interference with fusion between HIV infected cells. Inhibition of the viral enzyme reverse transcriptase | ||||
| C. okamuranus | Inhibition of dengue virus by interaction with the glycoprotein of the viral envelop | |||
| Proteins & peptides | Lectins | E. serra, G. marginata | Inhibition of the growth of V. vulnificus and V. pelagicus. Interaction between lectins and components of the bacterial cell wall | [25] |
| Protein hydrolysate fraction | S. longicruris | Inhibition of S. aureus growth | [105] | |
| Lectins | S. filiformis | Inhibition of several Gram-negative bacteria by interaction with compounds of the cell wall | [142] | |
| Lectins | H. musciformis | Inhibition of T. rubrum and C. lindemuthianum | ||
| Lectins | B. coacta, Griffithsia sp. | Antiviral effects against HIV, Hepatitis C virus and SARS-CoV by preventing the entry in the host cells | [143] | |
| Fatty acids | Bioactive fraction | S. vulgare, S. fusiforme | Perforation of the cell wall of S. aureus and K. neumoniae, cytoplasmic leakage and cell death | [144] |
| Bioactive fraction | G. edulis | Rupture of cell membrane of Vibrio spp and A. hydrophila | [145] | |
| Bioactive fraction | S. marginatum, U. lactuca | Fatty acids could be involved in the inhibition S. aureus, E. coli and P. vulgaris | [146] | |
| Bioactive fraction | B. tenella | Inhibition of C. cladosporioides and C. sphaerospermum by disrupting the cell membrane | [147] | |
| Sulfoquinovosyldia- cylglycerol | C. racemosa | Antiviral effects against HSV type 2 by disturbing the initial stages of the viral life cycle | [148] | |
| Pigments | Fucoxanthin | H. elongata | Inhibition of L. monocytogenes | [149] |
| Fucoxanthin | Commercial extract | Inhibition of several pathogenic bacteria by increasing cell membrane permeability, leakage of cytoplasm and inhibition of nucleic acid | [128] |
| Method | Conditions | Macroalgae | Bioactive Compound | Main Outcomes | Ref. |
|---|---|---|---|---|---|
| SFE | 314 bar, 10 °C | D. salina | Fucoidans | Inhibition of E. coli, S. aureus, and C. albicans, growth | [151] |
| 300 bar, 50 °C | F. vesiculosus | Fucosterol | Inhibition of Fusarium sp. | [152] | |
| UAE | 200 W, 20 kHz; 55 °C; 20 min | N. zanardinii | Fucoidans | No activity against E. coli, L. monocytogenes P. aeruginosa and S. aureus | [153] |
| MAE | 1500 W; 150 °C; 10 min (× 2) | N. zanardinii | Fucoidans | Positive activity against E. coli | [153] |
| 50 °C, 500 W, 10 min, MeOH, EtOH | Oedogonium sp./Stigeoclonium sp./Ulothrix sp./Nitzschia sp. | n.d. | All extracts inhibited at least one microorganism tested | [154] | |
| UMAE | 65 °C, 3 h (x 2) | N. zanardinii | Fucoidans | Inhibition of P. aeruginosa, but no effect on E. coli L. monocytogenes and S. aureus | [153] |
| EAE | Alcalase: 2.5 mL (2.4 U/g), pH 8, 50 °C, 24 h Flavourzyme: 2.5 mL (500 U/g), pH 7, 50 °C, 24 h Cellulase: 2.5 g (3 U/mg), pH 4.5, 50 °C, 24 h Viscozyme: 2.5 mL 100 fungal β-glucanase U/mL, pH 4.5, 50 °C, 24 h | N. zanardinii | Fucoidans | No activity against E. coli L. monocytogenes, P. aeruginosa and S. aureus | [153] |
| Viscozyme L: pH 4.5 (0.1M, AB), 50 °C; AMG 300 L: pH 4.5 (0.1M, AB), 60 °C; Celluclast: pH 4.5 (0.1M, AB), 50 °C; Termamyl: pH 6 (0.1 M, SPB), 60 °C; Ultraflo: pH 6 (0.1 M, SPB), 40 °C; Flavourzyme: pH 7 (0.1 M, SPB), 50 °C; Alcalase: pH 8 (0.1 M, SPB), 50 °C; Neutrase: pH 8 (0.1 M, SPB), 50 °C; | S. boveanum, S. angustifolium, P. gymnospora, C. cervicornis, C. sinuosa, I. stellate, F. irregularis | Polyphenols/ polysaccharides | F. irregularis extracts obtained with Viscozyme, Celluclas and Flavourzyme inhibited S. aureus. P. gymnospora and C. sinuosa extracts obtained with Celluclast inhibited E. feacalis. | [155] | |
| EUAE | 65 °C, 3 h (× 2) | N. zanardinii | Fucoidans | Inhibition of P. aeruginosa, but no effect on E. coli L. monocytogenes and S. aureus | [153] |
| SWE | 1500 W, 150 °C, 10 min (× 2) | N. zanardinii | Fucoidans | Inhibition of E. coli and P. aeruginosa, but no effect on L. monocytogenes and S. aureus | [153] |
| SWH | 200–280 °C, 1.3–6.0 MPa, Catalyst -Acetic acid | L. japonica | n.d. | Strong antibacterial activity against S. typhimurium and E. coli, | [156] |
| PLE | Hx, EtOAc, AcO, EtOH, EtOH: H2O (50:50) | U. intestinalis, U. lactuca, F. vesiculosus, D. dichotoma, C. baccata, H. elongata | Fatty acids | F. vesiculosus extract exhibited the best antimicrobial properties | [157] |
| Hx, EtOH, W; 200 °C, 20 min | H. elongata | Fatty acids/pigments | All extracts presented antimicrobial activity against S. aureus, E. coli, C. albicans and A. niger | [158] | |
| H2O, MeOH, DCM Temperature (20, 40, 60 °C) | F. vesiculosus | Phlorotannins/phosphatidylcholine/ betaine/ lipids/chlorophylls/carotenoids | F. vesiculos extract was only effective as an antimicrobial agent to MRSA | [159] |
| Macroalgae | NPs | Size (nm) | Antimicrobial Activity | Ref. |
|---|---|---|---|---|
| S. muticum | Ag | 43–79 | Growth inhibition of B. subtilis, K. pneumoniae and S. typhi | [211] |
| G. amansii | Ag | 27–54 | Antifouling activity against P. aeruginosa, V. parahaemolyticus, E. coli, A. hydrophila, B. pumilus and S. aureus | [212] |
| G. corneum | Ag | 20–50 | Antimicrobial and antibiofilm activity against C. albicans and E. coli | [22] |
| G. corticata | Ag | 18–46 | Antifungal activity against Candida spp. | [213] |
| G. birdiae | Ag | 20–95 | Antimicrobial activity against E. coli | [214] |
| S. wightii | Ag | 55–70 | Maximum growth inhibition against M. luteus > S. marcescens | [215] |
| V. pachynema | Ag | 30–40 | Moderate growth inhibition against M. luteus > S. marcescens | [215] |
| P. hornemannii | Ag | 70–75 | Antimicrobial activity against the fish pathogens: V. parahaemolyticus, V. vulnificus, V. harveyii and V. anguillarum | [216] |
| U. faciata | Ag | 7 | Bacterial reduction in textile fabrics against S. aureus and E. coli | [217] |
| P. capillacea | Ag | 7 | Bacterial reduction in textile fabrics against S. aureus and E. coli | [217] |
| J. rubens | Ag | 12 | Bacterial reduction in textile fabrics against S. aureus and E. coli | [217] |
| C. sinusa | Ag | 20 | Bacterial reduction in textile fabrics against S. aureus and E. coli | [217] |
| S. plagiophyllum | AgCl | 18–42 | Growth inhibition of E.a coli | [218] |
| S. marginatum | Au | 18–94 | Growth inhibition of P. aeruginosa, K. oxytoca, E. faecalis, K. pneumoniae, V. parahaemolyticus, V. cholerae, S. typhi, S. paratyphi, and P. vulgaris | [219] |
| S. plagiophyllum | Au | 65–66 | Antibacterial activity against S. typhi and E. coli by bacteria membrane lysis | [220] |
| C. sinuosa | Fe3O4 | 11–34 | Excellent antifungal activity against A. flavus and F. oxysporum. Antibacterial activity against E. coli, P. aeruginosa, S. typhi, V. cholera, B. subtilis and S. aureus | [221] |
| P. capillacea | Fe3O4 | 16–23 | Antibacterial activity against E. coli, P. aeruginosa, S. typhi, V. cholera, B. subtilis and S. aureus | [221] |
| U. flexuosa | Fe3O4 | 12 | Antibacterial activity against B. subtilis, S. aureus, E. coli, E. faecalis and S.s epidermidis | [222] |
| S. wightii | ZrO2 | 4.8 | Enhancement of the antibacterial activity against B. subtilis, E. coli and S. typhi | [223] |
| S. wightii | MgO | 68 | Antibacterial activity against S. pneumonia, MRSA 11, MRSA 56, E. coli, P. aeruginosa and A. baumannii | [224] |
| S. myriocystum | ZnO | 76–186 | Growth inhibition against Gram (+) bacteria: S. mutans > M. luteus and Gram (-): Neisseria gonorrohea > V. cholera > K. pneumonia. Antifungal activity against C. albicans > A. niger | [225] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.; Silva, S.A.; Carpena, M.; Garcia-Oliveira, P.; Gullón, P.; Barroso, M.F.; Prieto, M.A.; Simal-Gandara, J. Macroalgae as a Source of Valuable Antimicrobial Compounds: Extraction and Applications. Antibiotics 2020, 9, 642. https://doi.org/10.3390/antibiotics9100642
Silva A, Silva SA, Carpena M, Garcia-Oliveira P, Gullón P, Barroso MF, Prieto MA, Simal-Gandara J. Macroalgae as a Source of Valuable Antimicrobial Compounds: Extraction and Applications. Antibiotics. 2020; 9(10):642. https://doi.org/10.3390/antibiotics9100642
Chicago/Turabian StyleSilva, Aurora, Sofia A. Silva, M. Carpena, P. Garcia-Oliveira, P. Gullón, M. Fátima Barroso, M.A. Prieto, and J. Simal-Gandara. 2020. "Macroalgae as a Source of Valuable Antimicrobial Compounds: Extraction and Applications" Antibiotics 9, no. 10: 642. https://doi.org/10.3390/antibiotics9100642
APA StyleSilva, A., Silva, S. A., Carpena, M., Garcia-Oliveira, P., Gullón, P., Barroso, M. F., Prieto, M. A., & Simal-Gandara, J. (2020). Macroalgae as a Source of Valuable Antimicrobial Compounds: Extraction and Applications. Antibiotics, 9(10), 642. https://doi.org/10.3390/antibiotics9100642