
Generation and Characterization of Stable Small Colony Variants of USA300 Staphylococcus aureus in RAW 264.7 Murine Macrophages

 , ,
, ,  ,
,  ,
,  , ,
, ,  ,
, 
 and
and
Abstract
1. Introduction
2. Results
2.1. Carnosine Does Not Show Antimicrobial Activity on ATCC BAA-1556_Catania
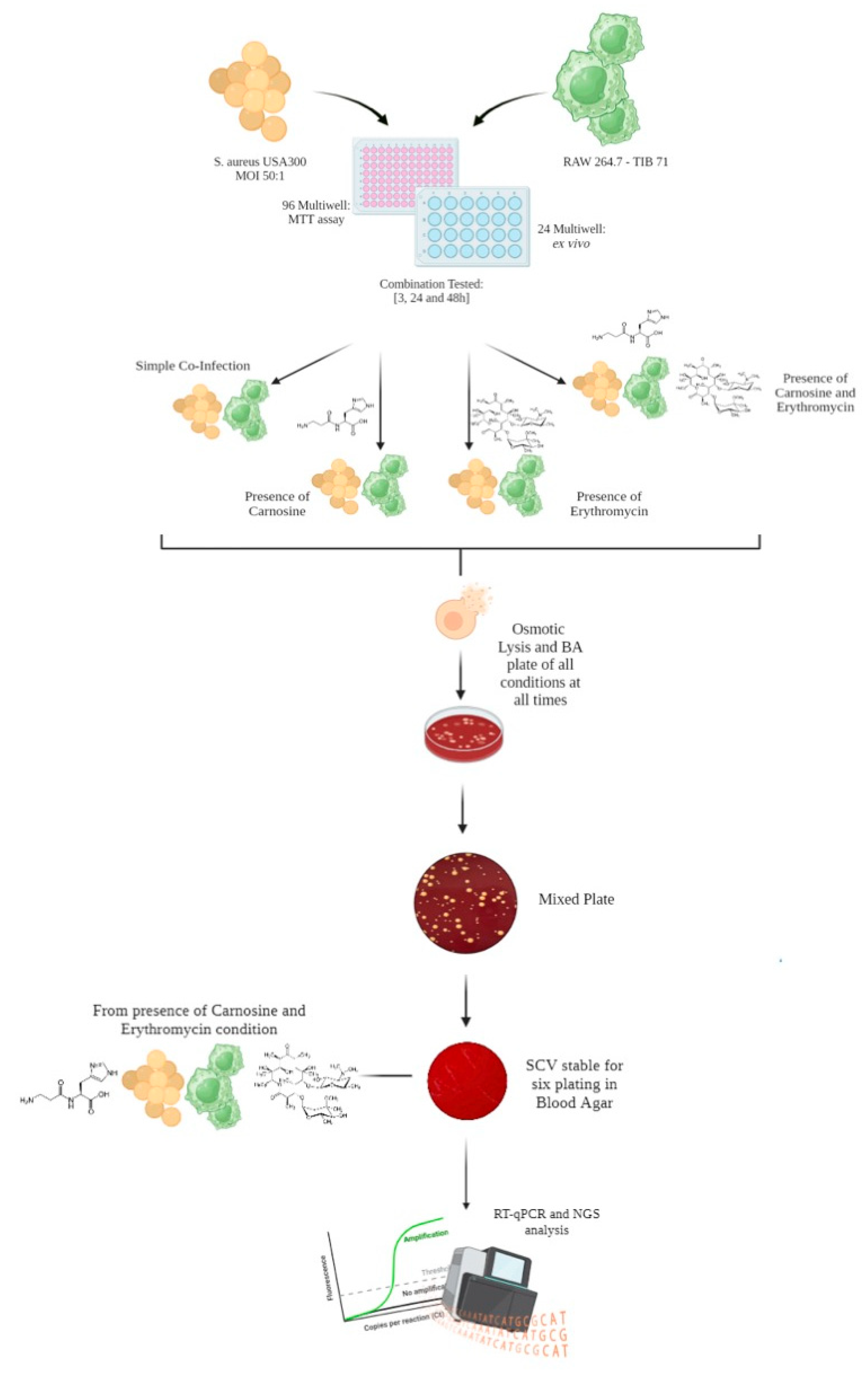
2.2. Macrophages Infection and Evaluation of Small Colony Variants (SCVs) Formation and Stability
2.3. Genome Analysis of SCV
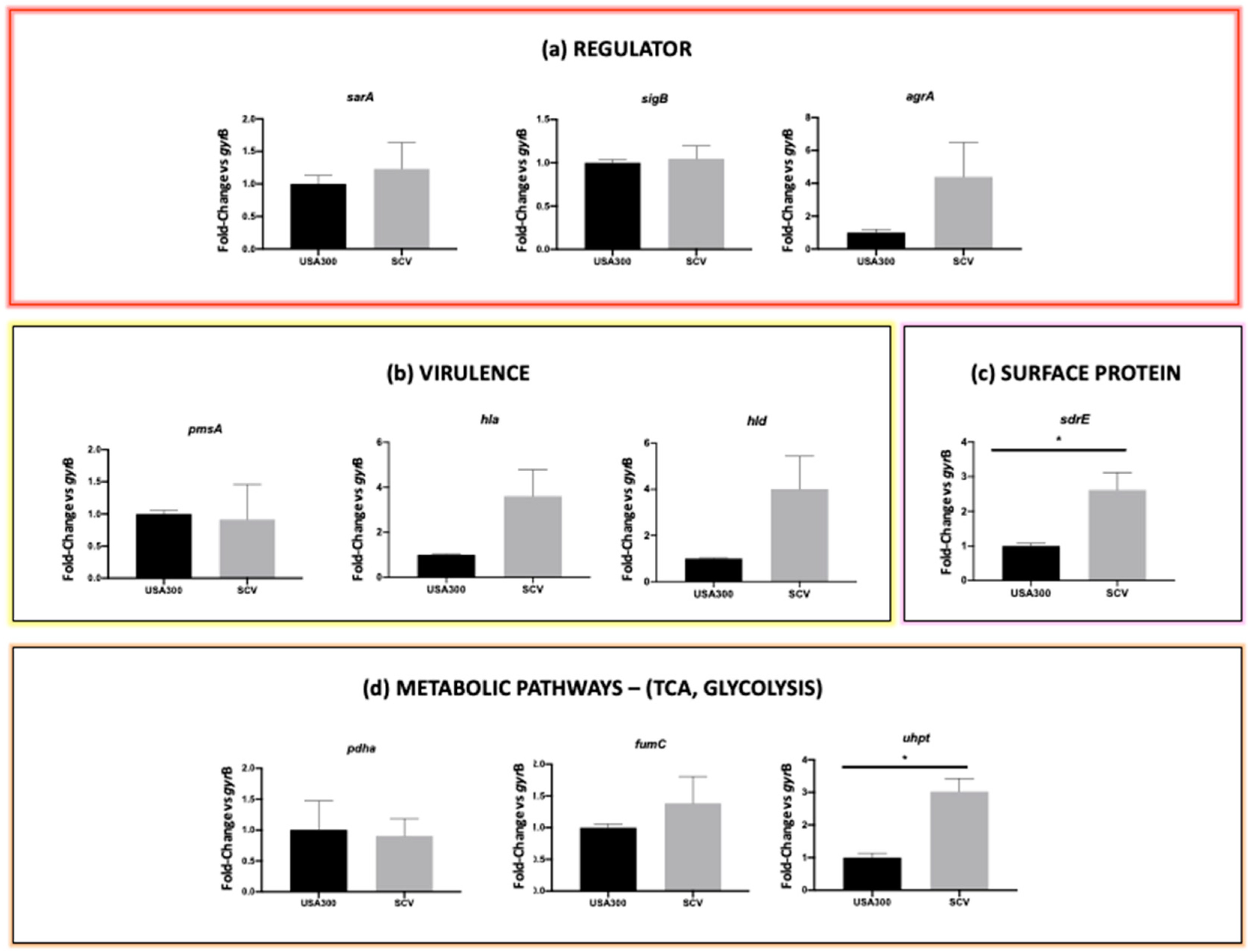
2.4. Comparative Gene Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain
4.2. Determination of Minimum Inhibitory Concentration
4.3. Infection Assay
4.3.1. Eukaryotic Cell Culture Preparation
4.3.2. Infection of RAW 264.7 Cells
4.4. Evaluation of SCV Stability
4.5. Genomic Analysis
4.5.1. DNA Extraction
4.5.2. Genome Sequencing
4.5.3. Bioinformatic Analysis
4.5.4. Data Availability Statement
4.6. Gene Expression Study
4.6.1. Primer Design
4.6.2. RNA Extraction
4.6.3. RT-qPCR
4.6.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [PubMed]
- Kahl, B.C.; Becker, K.; Löffler, B. Clinical Significance and Pathogenesis of Staphylococcal Small Colony Variants in Persistent Infections. Clin. Microbiol. Rev. 2016, 29, 401–427. [Google Scholar] [CrossRef] [PubMed]
- Tuchscherr, L.; Medina, E.; Hussain, M.; Völker, W.; Heitmann, V.; Niemann, S.; Holzinger, D.; Roth, J.; Proctor, R.A.; Becker, K.; et al. Staphylococcus aureus phenotype switching: An effective bacterial strategy to escape host immune response and establish a chronic infection. EMBO Mol. Med. 2011, 3, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Bongiorno, D.; Musso, N.; Lazzaro, L.M.; Mongelli, G.; Stefani, S.; Campanile, F. Detection of methicillin-resistant Staphylococcus aureus persistence in osteoblasts using imaging flow cytometry. Microbiologyopen 2020, 9, e1017. [Google Scholar] [CrossRef] [PubMed]
- Stracquadanio, S.; Musso, N.; Costantino, A.; Lazzaro, L.M.; Stefani, S.; Bongiorno, D. Staphylococcus aureus Internalization in Osteoblast Cells: Mechanisms, Interactions and Biochemical Processes. What Did We Learn from Experimental Models? Pathogens 2021, 10, 239. [Google Scholar] [CrossRef] [PubMed]
- Musso, N.; Caruso, G.; Bongiorno, D.; Grasso, M.; Bivona, D.A.; Campanile, F.; Caraci, F.; Stefani, S. Different Modulatory Effects of Four Methicillin-Resistant Staphylococcus aureus Clones on MG-63 Osteoblast-Like Cells. Biomolecules 2021, 11, 72. [Google Scholar] [CrossRef] [PubMed]
- Flannagan, R.S.; Heit, B.; Heinrichs, D.E. Intracellular replication of Staphylococcus aureus in mature phagolysosomes in macrophages precedes host cell death, and bacterial escape and dissemination. Cell Microbiol. 2016, 18, 514–535. [Google Scholar] [CrossRef]
- Kubica, M.; Guzik, K.; Koziel, J.; Zarebski, M.; Richter, W.; Gajkowska, B.; Golda, A.; Maciag-Gudowska, A.; Brix, K.; Shaw, L.; et al. A potential new pathway for Staphylococcus aureus dissemination: The silent survival of S. aureus phagocytosed by human monocyte-derived macrophages. PLoS ONE 2008, 3, e1409. [Google Scholar] [CrossRef]
- Horn, J.; Stelzner, K.; Rudel, T.; Fraunholz, M. Inside job: Staphylococcus aureus host-pathogen interactions. Int. J. Med. Microbiol. 2018, 308, 607–624. [Google Scholar] [CrossRef]
- Cafruny, W.A.; Bradley, S.E. Trojan Horse macrophages: Studies with the murine lactate dehydrogenase-elevating virus and implications for sexually transmitted virus infection. J. Gen. Virol. 1996, 77 Pt 12, 3005–3012. [Google Scholar] [CrossRef]
- Percivalle, E.; Sammartino, J.C.; Cassaniti, I.; Arbustini, E.; Urtis, M.; Smirnova, A.; Concardi, M.; Belgiovine, C.; Ferrari, A.; Lilleri, D.; et al. Macrophages and Monocytes: “Trojan Horses” in COVID-19. Viruses 2021, 13, 2178. [Google Scholar] [CrossRef]
- Bongiorno, D.; Musso, N.; Caruso, G.; Lazzaro, L.M.; Caraci, F.; Stefani, S.; Campanile, F. Staphylococcus aureus ST228 and ST239 as models for expression studies of diverse markers during osteoblast infection and persistence. Microbiologyopen 2021, 10, e1178. [Google Scholar] [CrossRef]
- Kalyankar, G.D.; Meister, A. Enzymatic synthesis of carnosine and related beta-alanyl and gamma-aminobutyryl peptides. J. Biol. Chem. 1959, 234, 3210–3218. [Google Scholar] [CrossRef]
- Hipkiss, A.R.; Preston, J.E.; Himsworth, D.T.; Worthington, V.C.; Keown, M.; Michaelis, J.; Lawrence, J.; Mateen, A.; Allende, L.; Eagles, P.A.; et al. Pluripotent protective effects of carnosine, a naturally occurring dipeptide. Ann. N. Y. Acad. Sci. 1998, 854, 37–53. [Google Scholar] [CrossRef]
- Hill, C.A.; Harris, R.C.; Kim, H.J.; Harris, B.D.; Sale, C.; Boobis, L.H.; Kim, C.K.; Wise, J.A. Influence of beta-alanine supplementation on skeletal muscle carnosine concentrations and high intensity cycling capacity. Amino Acids 2007, 32, 225–233. [Google Scholar] [CrossRef]
- Caruso, G.; Caraci, F.; Jolivet, R.B. Pivotal role of carnosine in the modulation of brain cells activity: Multimodal mechanism of action and therapeutic potential in neurodegenerative disorders. Prog. Neurobiol. 2019, 175, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Fleisher-Berkovich, S.; Abramovitch-Dahan, C.; Ben-Shabat, S.; Apte, R.; Beit-Yannai, E. Inhibitory effect of carnosine and N-acetyl carnosine on LPS-induced microglial oxidative stress and inflammation. Peptides 2009, 30, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Privitera, A.; Antunes, B.M.; Lazzarino, G.; Lunte, S.M.; Aldini, G.; Caraci, F. The Therapeutic Potential of Carnosine as an Antidote against Drug-Induced Cardiotoxicity and Neurotoxicity: Focus on Nrf2 Pathway. Molecules 2022, 27, 4452. [Google Scholar] [CrossRef] [PubMed]
- Fresta, C.G.; Fidilio, A.; Lazzarino, G.; Musso, N.; Grasso, M.; Merlo, S.; Amorini, A.M.; Bucolo, C.; Tavazzi, B.; Lazzarino, G.; et al. Modulation of Pro-Oxidant and Pro-Inflammatory Activities of M1 Macrophages by the Natural Dipeptide Carnosine. Int. J. Mol. Sci. 2020, 21, 776. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Privitera, A.; Saab, M.W.; Musso, N.; Maugeri, S.; Fidilio, A.; Privitera, A.P.; Pittalà, A.; Jolivet, R.B.; Lanzanò, L.; et al. Characterization of Carnosine Effect on Human Microglial Cells under Basal Conditions. Biomedicines 2023, 11, 474. [Google Scholar] [CrossRef]
- Bukharin, O.V.; Stadnikov, A.A.; Chernova, O.L.; Kirgizova, S.B.; Kovbyk, L.V.; Boldyrev, A.A. Biological importance of the anti-carnosine activity of bacteria. Zh. Mikrobiol. Epidemiol. Immunobiol. 2000, (4 Suppl.), 56–59. [Google Scholar]
- Zhou, P.; Xia, D.; Xia, Y.; Zhang, H.; Wang, Y.; Tang, T.; Xu, S. Synergistic effect of vancomycin and l-homocarnosine alleviates Staphylococcus aureus-induced osteomyelitis in rats. Biomed. Pharmacother. 2019, 111, 31–35. [Google Scholar] [CrossRef]
- Ibrahim, N.; El Said, H.; Choukair, A. Zinc carnosine-based modified bismuth quadruple therapy vs standard triple therapy for Helicobacter pylori eradication: A randomized controlled study. World J. Clin. Cases 2022, 10, 227–235. [Google Scholar] [CrossRef]
- Caruso, G.; Benatti, C.; Musso, N.; Fresta, C.G.; Fidilio, A.; Spampinato, G.; Brunello, N.; Bucolo, C.; Drago, F.; Lunte, S.M.; et al. Carnosine Protects Macrophages against the Toxicity of Aβ1-42 Oligomers by Decreasing Oxidative Stress. Biomedicines 2021, 9, 477. [Google Scholar] [CrossRef]
- Bui, L.M.G.; Hoffmann, P.; Turnidge, J.D.; Zilm, P.S.; Kidd, S.P. Prolonged Growth of a Clinical Staphylococcus aureus Strain Selects for a Stable Small-Colony-Variant Cell Type. Infect. Immun. 2015, 83, 470–481. [Google Scholar] [CrossRef]
- Melter, O.; Radojevič, B. Small colony variants of Staphylococcus aureus—Review. Folia Microbiol. 2010, 55, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Ebbole, D.J.; Zalkin, H. Cloning and characterization of a 12-gene cluster from Bacillus subtilis encoding nine enzymes for de novo purine nucleotide synthesis. J. Biol. Chem. 1987, 262, 8274–8287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wright, J.A.; Osman, A.A.; Nair, S.P. An aroD Ochre Mutation Results in a Staphylococcus aureus Small Colony Variant That Can Undergo Phenotypic Switching via Two Alternative Mechanisms. Front. Microbiol. 2017, 8, 1001. [Google Scholar] [CrossRef] [PubMed]
- Fraunholz, M.; Sinha, B. Intracellular Staphylococcus aureus: Live-in and let die. Front. Cell Infect. Microbiol. 2012, 2, 43. [Google Scholar] [CrossRef] [PubMed]
- Besier, S.; Ludwig, A.; Ohlsen, K.; Brade, V.; Wichelhaus, T.A. Molecular analysis of the thymidine-auxotrophic small colony variant phenotype of Staphylococcus aureus. Int. J. Med. Microbiol. 2007, 297, 217–225. [Google Scholar] [CrossRef]
- Dean, M.A.; Olsen, R.J.; Long, S.W.; Rosato, A.E.; Musser, J.M. Identification of point mutations in clinical Staphylococcus aureus strains that produce small-colony variants auxotrophic for menadione. Infect. Immun. 2014, 82, 1600–1605. [Google Scholar] [CrossRef] [PubMed]
- Goncheva, M.I.; Flannagan, R.S.; Sterling, B.E.; Laakso, H.A.; Friedrich, N.C.; Kaiser, J.C.; Watson, D.W.; Wilson, C.H.; Sheldon, J.R.; McGavin, M.J.; et al. Stress-induced inactivation of the Staphylococcus aureus purine biosynthesis repressor leads to hypervirulence. Nat. Commun. 2019, 10, 775. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, H.; Chen, J.; Wang, T.; Tu, S.; Zhang, X.; Wang, Q.; Yin, Y.; Zhang, Y.; Wang, X.; et al. Transcriptome and Proteome of Methicillin-Resistant Staphylococcus aureus Small-Colony Variants Reveal Changed Metabolism and Increased Immune Evasion. Microbiol. Spectr. 2023, 11, e0189822. [Google Scholar] [CrossRef]
- Wong Fok Lung, T.; Monk, I.R.; Acker, K.P.; Mu, A.; Wang, N.; Riquelme, S.A.; Pires, S.; Noguera, L.P.; Dach, F.; Gabryszewski, S.J.; et al. Staphylococcus aureus small colony variants impair host immunity by activating host cell glycolysis and inducing necroptosis. Nat. Microbiol. 2020, 5, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, A.; Fraunholz, M.J. In or out: Phagosomal escape of Staphylococcus aureus. Cell Microbiol. 2019, 21, e12997. [Google Scholar] [CrossRef]
- Tuchscherr, L.; Bischoff, M.; Lattar, S.M.; Noto Llana, M.; Pförtner, H.; Niemann, S.; Geraci, J.; Van de Vyver, H.; Fraunholz, M.J.; Cheung, A.L.; et al. Sigma Factor SigB Is Crucial to Mediate Staphylococcus aureus Adaptation during Chronic Infections. PLoS Pathog. 2015, 11, e1004870. [Google Scholar] [CrossRef]
- Kriegeskorte, A.; König, S.; Sander, G.; Pirkl, A.; Mahabir, E.; Proctor, R.A.; von Eiff, C.; Peters, G.; Becker, K. Small colony variants of Staphylococcus aureus reveal distinct protein profiles. Proteomics 2011, 11, 2476–2490. [Google Scholar] [CrossRef]
- Stoneham, S.M.; Cantillon, D.M.; Waddell, S.J.; Llewelyn, M.J. Spontaneously Occurring Small-Colony Variants of Staphylococcus aureus Show Enhanced Clearance by THP-1 Macrophages. Front. Microbiol. 2020, 11, 1300. [Google Scholar] [CrossRef] [PubMed]
- Azarian, T.; Daum, R.S.; Petty, L.A.; Steinbeck, J.L.; Yin, Z.; Nolan, D.; Boyle-Vavra, S.; Hanage, W.P.; Salemi, M.; David, M.Z. Intrahost Evolution of Methicillin-Resistant Staphylococcus aureus USA300 Among Individuals With Reoccurring Skin and Soft-Tissue Infections. J. Infect. Dis. 2016, 214, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhai, L.; Qin, T.; Hu, L.; Wang, J.; Zhang, Z.; Sui, C.; Zhang, L.; Zhou, D.; Lv, M.; et al. Time-Course Transcriptome Analysis of the Lungs of Mice Challenged with Aerosols of Methicillin-Resistant Staphylococcus aureus USA300 Clone Reveals Inflammatory Balance. Biomolecules 2023, 13, 347. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Conly, J.; McClure, J.-A.; Kurwa, H.A.; Zhang, K. Arginine Catabolic Mobile Element in Evolution and Pathogenicity of the Community-Associated Methicillin-Resistant Staphylococcus aureus Strain USA300. Microorganisms 2020, 8, 275. [Google Scholar] [CrossRef] [PubMed]
- EUCAST: Clinical Breakpoints and Dosing of Antibiotics. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 12 July 2022).
- Bongiorno, D.; Bivona, D.A.; Cicino, C.; Trecarichi, E.M.; Russo, A.; Marascio, N.; Mezzatesta, M.L.; Musso, N.; Privitera, G.F.; Quirino, A.; et al. Omic insights into various ceftazidime-avibactam-resistant Klebsiella pneumoniae isolates from two southern Italian regions. Front. Cell Infect. Microbiol. 2022, 12, 1010979. [Google Scholar] [CrossRef] [PubMed]
- Pinzauti, D.; Iannelli, F.; Pozzi, G.; Santoro, F. DNA isolation methods for Nanopore sequencing of the Streptococcus mitis genome. Microb. Genom. 2022, 8, 000764. [Google Scholar] [CrossRef]
- Teodori, L.; Colombini, L.; Cuppone, A.M.; Lazzeri, E.; Pinzauti, D.; Santoro, F.; Iannelli, F.; Pozzi, G. Complete Genome Sequence of Lactobacillus crispatus Type Strain ATCC 33820. Microbiol. Resour. Announc. 2021, 10, e00634-21. [Google Scholar] [CrossRef]
- Pinzauti, D.; De Giorgi, S.; Fox, V.; Lazzeri, E.; Messina, G.; Santoro, F.; Iannelli, F.; Ricci, S.; Pozzi, G. Complete Genome Sequences of Mycobacterium chimaera Strains 850 and 852, Isolated from Heater-Cooler Unit Water. Microbiol. Resour. Announc. 2022, 11, e01021-21. [Google Scholar] [CrossRef]
- Krueger, F.; Trim Galore. 7 July 2022. Available online: https://github.com/FelixKrueger/TrimGalore (accessed on 12 July 2022).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Colombini, L.; Cuppone, A.M.; Tirziu, M.; Lazzeri, E.; Pozzi, G.; Santoro, F.; Iannelli, F. The Mobilome-Enriched Genome of the Competence-Deficient Streptococcus pneumoniae BM6001, the Original Host of Integrative Conjugative Element Tn5253, Is Phylogenetically Distinct from Historical Pneumococcal Genomes. Microorganisms 2023, 11, 1646. [Google Scholar] [CrossRef]
- Pidwill, G.R.; Gibson, J.F.; Cole, J.; Renshaw, S.A.; Foster, S.J. The Role of Macrophages in Staphylococcus aureus Infection. Front. Immunol. 2021, 11, 620339. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Product | Genome Nucleotide Position | Nucleotide Position in Cds | Codon | Amino acid Change | Type of Mutation |
|---|---|---|---|---|---|---|
| purR | Pur operon repressor | 532,783 | T803A | ATC>AAC | I267N | Missense |
| Intergenic | - | 909,332 | C>T | - | - | - |
| aroK | Shikimate kinase | 1,659,182 | G306A | TGG>TAG | W102* | Nonsense |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bivona, D.; Bonomo, C.; Colombini, L.; Bonacci, P.G.; Privitera, G.F.; Caruso, G.; Caraci, F.; Santoro, F.; Musso, N.; Bongiorno, D.; et al. Generation and Characterization of Stable Small Colony Variants of USA300 Staphylococcus aureus in RAW 264.7 Murine Macrophages. Antibiotics 2024, 13, 264. https://doi.org/10.3390/antibiotics13030264
Bivona D, Bonomo C, Colombini L, Bonacci PG, Privitera GF, Caruso G, Caraci F, Santoro F, Musso N, Bongiorno D, et al. Generation and Characterization of Stable Small Colony Variants of USA300 Staphylococcus aureus in RAW 264.7 Murine Macrophages. Antibiotics. 2024; 13(3):264. https://doi.org/10.3390/antibiotics13030264
Chicago/Turabian StyleBivona, Dalida, Carmelo Bonomo, Lorenzo Colombini, Paolo G. Bonacci, Grete F. Privitera, Giuseppe Caruso, Filippo Caraci, Francesco Santoro, Nicolò Musso, Dafne Bongiorno, and et al. 2024. "Generation and Characterization of Stable Small Colony Variants of USA300 Staphylococcus aureus in RAW 264.7 Murine Macrophages" Antibiotics 13, no. 3: 264. https://doi.org/10.3390/antibiotics13030264
APA StyleBivona, D., Bonomo, C., Colombini, L., Bonacci, P. G., Privitera, G. F., Caruso, G., Caraci, F., Santoro, F., Musso, N., Bongiorno, D., Iannelli, F., & Stefani, S. (2024). Generation and Characterization of Stable Small Colony Variants of USA300 Staphylococcus aureus in RAW 264.7 Murine Macrophages. Antibiotics, 13(3), 264. https://doi.org/10.3390/antibiotics13030264