


Mycobacterium avium ssp. paratuberculosis and Crohn’s Disease—Diagnostic Microbiological Investigations Can Inform New Therapeutic Approaches



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. The Pathogenic Potential of MAP
3. Isolation of MAP in Humans
4. The Importance of the Diagnostic Laboratory
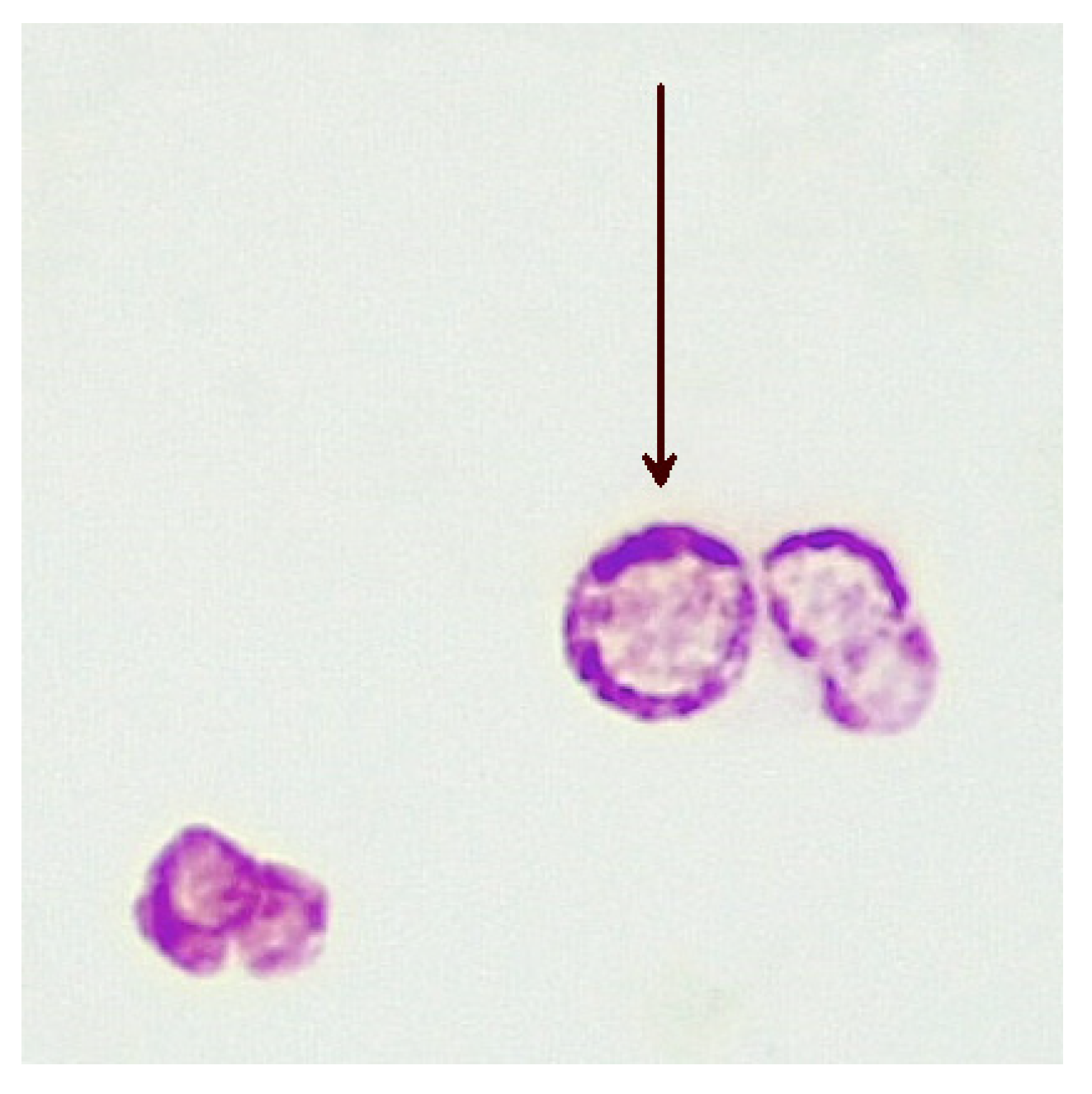
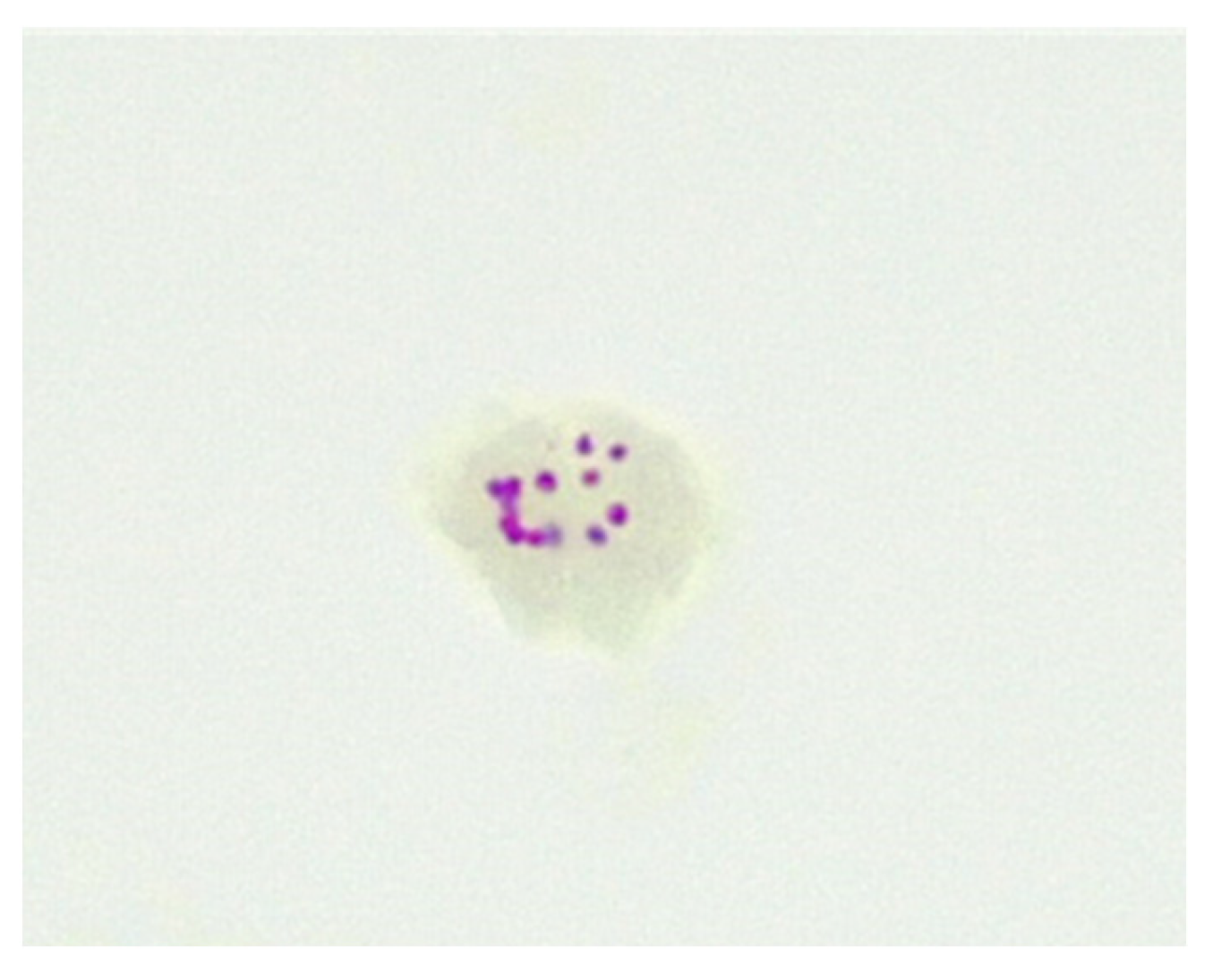
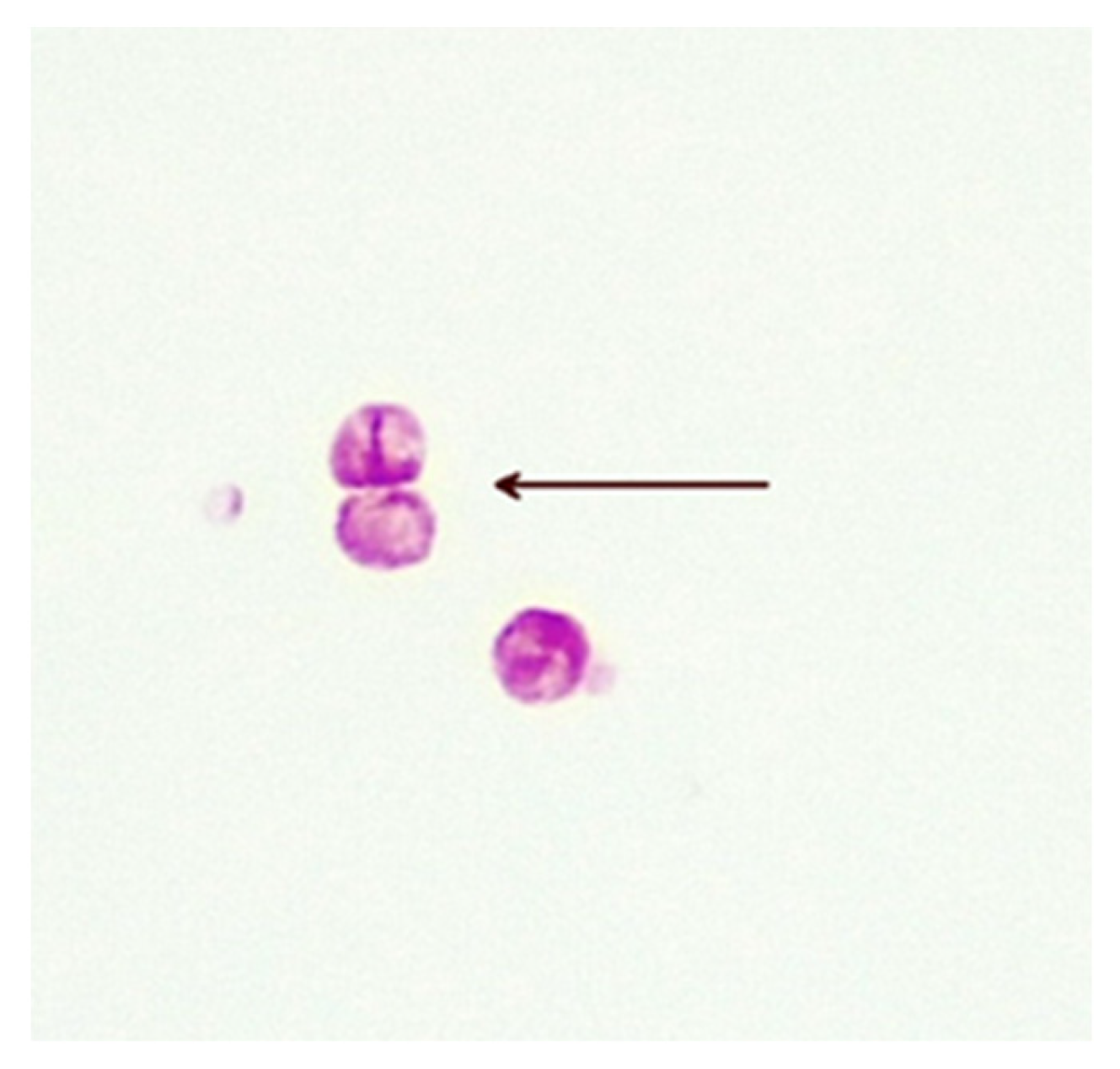
5. Cell-Wall-Deficient Mycobacteria in Crohn’s Disease
6. Quantitation of Opportunistic Mycobacteria
7. Antimycobacterial Therapy for MAP Infection
7.1. Testing for Antimycobacterial Susceptibilities
7.2. Therapy for CD with Antimycobacterial Antibiotics
8. Methods for Antimicrobial Susceptibility Testing
8.1. Molecular Detection of Resistance Genes
8.2. Breakpoint Minimum Inhibitory Concentration (MIC)
8.3. Alamar Blue
8.4. Pour Plate Counts
8.5. Plate Counts Using Surface Inoculation
8.6. Animal Testing (Koch’s Postulates)
8.7. qPCR
9. Potential Therapeutic Options for MAP Acquired CD
9.1. Dietary Regulation of the Gut Microbiome
9.2. Probiotic Supplements
9.3. Faecal Microbiome Transplant (FMT)
9.4. Gut Sterilisation
9.5. Bacteriophage Therapy
9.6. Infection Control
9.7. Vaccination
9.8. Immune Therapy
10. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Twort, F.W.; Ingram, G.L.Y.; Hill, L.E. A method for isolating and cultivating the mycobacterium enteritidis chronicæ pseudotuberculosæ bovis, Jöhne, and some experiments on the preparation of a diagnostic vaccine for pseudo-tuberculous enteritis of bovines. Proc. R. Soc. Lond. Ser. B Contain. Pap. A Biol. Character 1997, 84, 517–542. [Google Scholar] [CrossRef]
- Chiodini, R.J.; Van Kruiningen, H.J.; Thayer, W.R.; Coutu, J.A. Spheroplastic phase of mycobacteria isolated from patients with Crohn’s disease. J. Clin. Microbiol. 1986, 24, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Timms, V.J.; Daskalopoulos, G.; Mitchell, H.M.; Neilan, B.A. The Association of Mycobacterium avium subsp. paratuberculosis with Inflammatory Bowel Disease. PLoS ONE 2016, 11, e0148731. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.V.; Chauhan, D.S.; Singh, S.V.; Kumar, V.; Singh, A.; Yadav, A.; Yadav, V.S. Current status of Mycobacterium avium subspecies paratuberculosis infection in animals & humans in India: What needs to be done? Indian J. Med. Res. 2016, 144, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Ekundayo, T.C.; Okoh, A.I. Systematic Assessment of Mycobacterium avium Subspecies paratuberculosis Infections from 1911–2019: A Growth Analysis of Association with Human Autoimmune Diseases. Microorganisms 2020, 8, 1212. [Google Scholar] [CrossRef]
- Mintz, M.J.; Lukin, D.J. Mycobacterium avium subspecies paratuberculosis (MAP) and Crohn’s disease: The debate continues. Transl. Gastroenterol. Hepatol. 2023, 8, 28. [Google Scholar] [CrossRef]
- Magro, F.; Moreira, P.L.; Catalano, G.; Alves, C.; Roseira, J.; Estevinho, M.M.; Silva, I.; Dignass, A.; Peyrin-Biroulet, L.; Danese, S.; et al. Has the therapeutical ceiling been reached in Crohn’s disease randomized controlled trials? A systematic review and meta-analysis. United Eur. Gastroenterol. J. 2023, 11, 202–217. [Google Scholar] [CrossRef]
- Singh, A.V.; Singh, S.V.; Singh, P.K.; Sohal, J.S.; Singh, M.K. High prevalence of Mycobacterium avium subspecies paratuberculosis (‘Indian bison type’) in animal attendants suffering from gastrointestinal complaints who work with goat herds endemic for Johne’s disease in India. Int. J. Infect. Dis. 2011, 15, e677–e683. [Google Scholar] [CrossRef]
- Ananthakrishnan, A.N.; Adler, J.; Chachu, K.A.; Nguyen, N.H.; Siddique, S.M.; Weiss, J.M.; Sultan, S.; Velayos, F.S.; Cohen, B.L.; Singh, S. AGA Clinical Practice Guideline on the Role of Biomarkers for the Management of Crohn’s Disease. Gastroenterology 2023, 165, 1367–1399. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, E.W. Original mycobacterial sin. Tubercle 1970, 51, 316–321. [Google Scholar] [CrossRef]
- Markova, N.; Slavchev, G.; Michailova, L. Presence of mycobacterial L-forms in human blood: Challenge of BCG vaccination. Human. Vaccines Immunother. 2015, 11, 1192–1200. [Google Scholar] [CrossRef]
- Dimova, T.; Terzieva, A.; Djerov, L.; Dimitrova, V.; Nikolov, A.; Grozdanov, P.; Markova, N. Mother-to-newborn transmission of mycobacterial L-forms and Vδ2 T-cell response in placentobiome of BCG-vaccinated pregnant women. Sci. Rep. 2017, 7, 17366. [Google Scholar] [CrossRef]
- Chiodini, R.J.; Van Kruiningen, H.J.; Merkal, R.S.; Thayer, W.R.; Coutu, J.A. Characteristics of an unclassified Mycobacterium species isolated from patients with Crohn’s disease. J. Clin. Microbiol. 1984, 20, 966–971. [Google Scholar] [CrossRef]
- Sechi, L.A.; Scanu, A.M.; Molicotti, P.; Cannas, S.; Mura, M.; Dettori, G.; Fadda, G.; Zanetti, S. Detection and Isolation of Mycobacterium avium subspecies paratuberculosis from intestinal mucosal biopsies of patients with and without Crohn’s disease in Sardinia. Am. J. Gastroenterol. 2005, 100, 1529–1536. [Google Scholar] [CrossRef]
- Richter, E.; Wessling, J.; Lügering, N.; Domschke, W.; Rüsch-Gerdes, S. Mycobacterium avium subsp. paratuberculosis Infection in a Patient with HIV, Germany. Emerg. Infect. Dis. 2002, 8, 729–731. [Google Scholar] [CrossRef]
- Estevinho, M.M.; Cabeda, J.; Santiago, M.; Machado, E.; Silva, R.; Duro, M.; Pita, I.; Morais, R.; Macedo, G.; Bull, T.J.; et al. Viable Mycobacterium avium subsp. paratuberculosis Colonizes Peripheral Blood of Inflammatory Bowel Disease Patients. Microorganisms 2023, 11, 1520. [Google Scholar] [CrossRef]
- Reller, L.B.; Weinstein, M.P.; Peterson, L.R.; Hamilton, J.D.; Baron, E.J.; Tompkins, L.S.; Miller, J.M.; Wilfert, C.M.; Tenover, F.C.; Thomson, R.B., Jr. Role of Clinical Microbiology Laboratories in the Management and Control of Infectious Diseases and the Delivery of Health Care. Clin. Infect. Dis. 2001, 32, 605–610. [Google Scholar] [CrossRef]
- Mahomed, S.; Dlamini-Mvelase, N.R.; Dlamini, M.; Mlisana, K. Failure of BACTECTM MGIT 960TM to detect Mycobacterium tuberculosis complex within a 42-day incubation period. Afr. J. Lab. Med. 2017, 6, 537. [Google Scholar] [CrossRef] [PubMed]
- Dane, H.; Stewart, L.D.; Grant, I.R. Culture of Mycobacterium avium subsp. paratuberculosis: Challenges, limitations and future prospects. J. Appl. Microbiol. 2023, 134, lxac017. [Google Scholar] [CrossRef] [PubMed]
- Aitken, J.M.; Phan, K.; Bodman, S.E.; Sharma, S.; Watt, A.; George, P.M.; Agrawal, G.; Tie, A.B.M. A Mycobacterium species for Crohn’s disease? Pathology 2021, 53, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Bishop, P.J.; Neumann, G. The history of the Ziehl-Neelsen stain. Tubercle 1970, 51, 196–206. [Google Scholar] [CrossRef]
- Piersimoni, C.; Nista, D.; Bornigia, S.; Gherardi, G. Unreliable Detection of Mycobacterium xenopi by the Nonradiometric Bactec MGIT 960 Culture System. J. Clin. Microbiol. 2009, 47, 804–806. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bansal-Mutalik, R.; Nikaido, H. Mycobacterial outer membrane is a lipid bilayer and the inner membrane is unusually rich in diacyl phosphatidylinositol dimannosides. Proc. Natl. Acad. Sci. USA 2014, 111, 4958–4963. [Google Scholar] [CrossRef] [PubMed]
- Sueoka, E.; Nishiwaki, S.; Okabe, S.; Iida, N.; Suganuma, M.; Yano, I.; Aoki, K.; Fujiki, H. Activation of protein kinase C by mycobacterial cord factor, trehalose 6-monomycolate, resulting in tumor necrosis factor-alpha release in mouse lung tissues. Jpn. J. Cancer Res. 1995, 86, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Thouvenel, L.; Rech, J.; Guilhot, C.; Bouet, J.-Y.; Chalut, C. In vivo imaging of MmpL transporters reveals distinct subcellular locations for export of mycolic acids and non-essential trehalose polyphleates in the mycobacterial outer membrane. Sci. Rep. 2023, 13, 7045. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Meshcheryakov, V.A.; Poce, G.; Chng, S.-S. MmpL3 is the flippase for mycolic acids in mycobacteria. Proc. Natl. Acad. Sci. USA 2017, 114, 7993–7998. [Google Scholar] [CrossRef] [PubMed]
- Zang, X.; Dang, G.; Cai, Z.; Shao, M.; Tang, Y.; Cao, J.; Cui, Z.; Liu, S. Extracellular DNase MAP3916c attacks the neutrophil extracellular traps and is needed for Mycobacterium avium subsp. paratuberculosis virulence. Vet. Microbiol. 2022, 273, 109529. [Google Scholar] [CrossRef] [PubMed]
- Quigley, J.; Lewis, K. Noise in a Metabolic Pathway Leads to Persister Formation in Mycobacterium tuberculosis. Microbiol. Spectr. 2022, 10, e0294822. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Madiraju, M.V.V.S.; Fol, M.; Lofton, H.; Maloney, E.; Reynolds, R.; Rajagopalan, M. Mycobacterium tuberculosis cells growing in macrophages are filamentous and deficient in FtsZ rings. J. Bacteriol. 2006, 188, 1856–1865. [Google Scholar] [CrossRef]
- Lazenby, J.J.; Li, E.S.; Whitchurch, C.B. Cell wall deficiency—An alternate bacterial lifestyle? Microbiology 2022, 168, 001218. [Google Scholar] [CrossRef]
- Marquetoux, N.; Ridler, A.; Heuer, C.; Wilson, P. What counts? A review of in vitro methods for the enumeration of Mycobacterium avium subsp. paratuberculosis. Vet. Microbiol. 2019, 230, 265–272. [Google Scholar] [CrossRef]
- Kuenstner, J.T.; Potula, R.; Bull, T.J.; Grant, I.R.; Foddai, A.; Naser, S.A.; Bach, H.; Zhang, P.; Yu, D.; Lu, X.; et al. Presence of Infection by Mycobacterium avium subsp. paratuberculosis in the Blood of Patients with Crohn’s Disease and Control Subjects Shown by Multiple Laboratory Culture and Antibody Methods. Microorganisms 2020, 8, 2054. [Google Scholar] [CrossRef]
- Juste, R.A.; Elguezabal, N.; Pavón, A.; Garrido, J.M.; Geijo, M.; Sevilla, I.; Cabriada, J.L.; Tejada, A.; García-Campos, F.; Casado, R.; et al. Association between Mycobacterium avium subsp. paratuberculosis DNA in blood and cellular and humoral immune response in inflammatory bowel disease patients and controls. Int. J. Infect. Dis. 2009, 13, 247–254. [Google Scholar] [CrossRef]
- Masur, H. Recommendations on Prophylaxis and Therapy for Disseminated Mycobacterium avium Complex Disease in Patients Infected with the Human Immunodeficiency Virus. N. Engl. J. Med. 1993, 329, 898–904. [Google Scholar] [CrossRef]
- Zaw, M.T.; Emran, N.A.; Lin, Z. Mutations inside rifampicin-resistance determining region of rpoB gene associated with rifampicin-resistance in Mycobacterium tuberculosis. J. Infect. Public Health 2018, 11, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Patangia, D.V.; Anthony Ryan, C.; Dempsey, E.; Paul Ross, R.; Stanton, C. Impact of antibiotics on the human microbiome and consequences for host health. Microbiologyopen 2022, 11, e1260. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-H.; Xiao, Y.-X.; Hsiao, H.-C.; Jou, R. Development and Assessment of a Novel Whole-Gene-Based Targeted Next-Generation Sequencing Assay for Detecting the Susceptibility of Mycobacterium tuberculosis to 14 Drugs. Microbiol. Spectr. 2022, 10, e0260522. [Google Scholar] [CrossRef]
- Ängeby, K.; Juréen, P.; Kahlmeter, G.; Hoffner, S.E.; Schön, T. Challenging a dogma: Antimicrobial susceptibility testing breakpoints for Mycobacterium tuberculosis. Bull. World Health Organ. 2012, 90, 693. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J.; Douarre, P.; Coffey, A.; Buckley, J.; Cashman, B.; O’Farrell, K.; O’Mahony, J. Optimization of a Rapid Viability Assay for Mycobacterium avium subsp. paratuberculosis by Using alamarBlue. Appl. Environ. Microbiol. 2009, 75, 7870–7872. [Google Scholar] [CrossRef] [PubMed]
- Feldman, W.H.; Hinshaw, H.C. Streptomycin: A Valuable Anti-tuberculosis Agent. Br. Med. J. 1948, 1, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Bard, J.D.; Lee, F. Why Can’t We Just Use PCR? The Role of Genotypic versus Phenotypic Testing for Antimicrobial Resistance Testing. Clin. Microbiol. Newsl. 2018, 40, 87–95. [Google Scholar] [CrossRef]
- Rolain, J.M.; Mallet, M.N.; Fournier, P.E.; Raoult, D. Real-time PCR for universal antibiotic susceptibility testing. J. Antimicrob. Chemother. 2004, 54, 538–541. [Google Scholar] [CrossRef]
- Rashed, R.; Valcheva, R.; Dieleman, L.A. Manipulation of Gut Microbiota as a Key Target for Crohn’s Disease. Front. Med. 2022, 9, 887044. [Google Scholar] [CrossRef] [PubMed]
- Mirsepasi-Lauridsen, H.C.; Vallance, B.A.; Krogfelt, K.A.; Petersen, A.M. Escherichia coli Pathobionts Associated with Inflammatory Bowel Disease. Clin. Microbiol. Rev. 2019, 32, e00060-18. [Google Scholar] [CrossRef] [PubMed]
- Sears, C.L. The toxins of Bacteroides fragilis. Toxicon 2001, 39, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- Adamji, M.; Day, A.S. An overview of the role of exclusive enteral nutrition for complicated Crohn’s disease. Intest. Res. 2019, 17, 171–176. [Google Scholar] [CrossRef]
- Logan, M.; Clark, C.M.; Ijaz, U.Z.; Gervais, L.; Duncan, H.; Garrick, V.; Curtis, L.; Buchanan, E.; Cardigan, T.; Armstrong, L.; et al. The reduction of faecal calprotectin during exclusive enteral nutrition is lost rapidly after food re-introduction. Aliment. Pharmacol. Ther. 2019, 50, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Heuschkel, R. Synergy Between Immunosuppressive Therapy and Enteral Nutrition in the Management of Childhood Crohn’s Disease. J. Parenter. Enter. Nutr. 2005, 29, S160–S165. [Google Scholar] [CrossRef]
- Tokmak, S. P012 Efficacy of Exclusive Enteral Nutrition and Weekly Adalimumab Combination in Crohn’s Patients with Fibroinflammatory Stenosis. Am. J. Gastroenterol. 2021, 116 (Suppl. S1), S3. [Google Scholar] [CrossRef]
- Mckirdy, S.; Russell, R.K.; Svolos, V.; Gkikas, K.; Logan, M.; Hansen, R.; Gerasimidis, K. The Impact of Compliance during Exclusive Enteral Nutrition on Faecal Calprotectin in Children with Crohn Disease. J. Pediatr. Gastroenterol. Nutr. 2022, 74, 801. [Google Scholar] [CrossRef]
- Nikolaus, S.; Schulte, B.; Al-Massad, N.; Thieme, F.; Schulte, D.M.; Bethge, J.; Rehman, A.; Tran, F.; Aden, K.; Häsler, R.; et al. Increased Tryptophan Metabolism Is Associated with Activity of Inflammatory Bowel Diseases. Gastroenterology 2017, 153, 1504–1516.e2. [Google Scholar] [CrossRef]
- Pradhan, S.; Weiss, A.A. Probiotic Properties of Escherichia coli Nissle in Human Intestinal Organoids. mBio 2020, 11, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Click, R.E. Successful treatment of asymptomatic or clinically terminal bovine Mycobacterium avium subspecies paratuberculosis infection (Johne’s disease) with the bacterium Dietzia used as a probiotic alone or in combination with dexamethasone. Virulence 2011, 2, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Prantera, C. Probiotics for Crohn’s disease: What have we learned? Gut 2006, 55, 757–759. [Google Scholar] [CrossRef] [PubMed]
- Vakadaris, G.; Stefanis, C.; Giorgi, E.; Brouvalis, M.; Voidarou, C.; Kourkoutas, Y.; Tsigalou, C.; Bezirtzoglou, E. The Role of Probiotics in Inducing and Maintaining Remission in Crohn’s Disease and Ulcerative Colitis: A Systematic Review of the Literature. Biomedicines 2023, 11, 494. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Landman, C.; Seksik, P.; Berard, L.; Montil, M.; Nion-Larmurier, I.; Bourrier, A.; Le Gall, G.; Lalande, V.; De Rougemont, A.; et al. Fecal microbiota transplantation to maintain remission in Crohn’s disease: A pilot randomized controlled study. Microbiome 2020, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Sarrabayrouse, G.; Landolfi, S.; Pozuelo, M.; Willamil, J.; Varela, E.; Clark, A.; Campos, D.; Herrera, C.; Santiago, A.; Machiels, K.; et al. Mucosal microbial load in Crohn’s disease: A potential predictor of response to faecal microbiota transplantation. EBioMedicine 2020, 51, 102611. [Google Scholar] [CrossRef] [PubMed]
- Boicean, A.; Birlutiu, V.; Ichim, C.; Anderco, P.; Birsan, S. Fecal Microbiota Transplantation in Inflammatory Bowel Disease. Biomedicines 2023, 11, 1016. [Google Scholar] [CrossRef] [PubMed]
- Kao, D.H.; Roach, B.; Walter, J.; Lobenberg, R.; Wong, K. A51 effect of lyophilized sterile fecal filtrate vs lyophilized donor stool on recurrent clostridium difficile infection (rcdi): Preliminary results from a randomized, double-blind pilot study. J. Can. Assoc. Gastroenterol. 2019, 2, 101–102. [Google Scholar] [CrossRef]
- Ott, S.J.; Waetzig, G.H.; Rehman, A.; Moltzau-Anderson, J.; Bharti, R.; Grasis, J.A.; Cassidy, L.; Tholey, A.; Fickenscher, H.; Seegert, D.; et al. Efficacy of Sterile Fecal Filtrate Transfer for Treating Patients with Clostridium difficile Infection. Gastroenterology 2017, 152, 799–811.e7. [Google Scholar] [CrossRef]
- Agrawal, G.; Clancy, A.; Huynh, R.; Borody, T. Profound remission in Crohn’s disease requiring no further treatment for 3–23 years: A case series. Gut Pathog. 2020, 12, 16. [Google Scholar] [CrossRef]
- Hodgson, H.J.F. Review: Gut sterilization in inflammatory bowel disease. Aliment. Pharmacol. Ther. 1987, 1, 359–366. [Google Scholar] [CrossRef]
- Croswell, A.; Amir, E.; Teggatz, P.; Barman, M.; Salzman, N.H. Prolonged Impact of Antibiotics on Intestinal Microbial Ecology and Susceptibility to Enteric Salmonella Infection. Infect. Immun. 2009, 77, 2741–2753. [Google Scholar] [CrossRef]
- Ramirez, J.; Guarner, F.; Bustos Fernandez, L.; Maruy, A.; Sdepanian, V.L.; Cohen, H. Antibiotics as Major Disruptors of Gut Microbiota. Front. Cell. Infect. Microbiol. 2020, 10, 572912. [Google Scholar] [CrossRef]
- Singh, P.; Alm, E.J.; Kelley, J.M.; Cheng, V.; Smith, M.; Kassam, Z.; Nee, J.; Iturrino, J.; Lembo, A. Effect of antibiotic pretreatment on bacterial engraftment after Fecal Microbiota Transplant (FMT) in IBS-D. Gut Microbes 2022, 14, 2020067. [Google Scholar] [CrossRef]
- Hassouneh, R.; Bajaj, J.S. Gut Microbiota Modulation and Fecal Transplantation: An Overview on Innovative Strategies for Hepatic Encephalopathy Treatment. J. Clin. Med. 2021, 10, 330. [Google Scholar] [CrossRef]
- Schwartz, D.J.; Langdon, A.; Sun, X.; Langendorf, C.; Berthé, F.; Grais, R.F.; Trehan, I.; Isanaka, S.; Dantas, G. Effect of amoxicillin on the gut microbiome of children with severe acute malnutrition in Madarounfa, Niger: A retrospective metagenomic analysis of a placebo-controlled trial. Lancet Microbe 2023, 4, e931–e942. [Google Scholar] [CrossRef] [PubMed]
- Brives, C.; Pourraz, J. Phage therapy as a potential solution in the fight against AMR: Obstacles and possible futures. Palgrave Commun. 2020, 6, 100. [Google Scholar] [CrossRef]
- Claessen, D.; Errington, J. Cell Wall Deficiency as a Coping Strategy for Stress. Trends Microbiol. 2019, 27, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Ongenae, V.; Briegel, A.; Claessen, D. Cell wall deficiency as an escape mechanism from phage infection. Open Biol. 2021, 11, 210199. [Google Scholar] [CrossRef] [PubMed]
- Hermon-Taylor, J. Mycobacterium avium subspecies paratuberculosis, Crohn’s disease and the Doomsday scenario. Gut Pathog. 2009, 1, 15. [Google Scholar] [CrossRef] [PubMed]
- Orujyan, D.; Narinyan, W.; Rangarajan, S.; Rangchaikul, P.; Prasad, C.; Saviola, B.; Venketaraman, V. Protective Efficacy of BCG Vaccine against Mycobacterium leprae and Non-Tuberculous Mycobacterial Infections. Vaccines 2022, 10, 390. [Google Scholar] [CrossRef]
- El-Matary, W.; Yap, J.; Deora, V.; Singh, H. Bacillus Calmette Guerin (BCG) Vaccine for Inducing and Maintaining Remission in Crohn’s Disease: A Systematic Review. J. Clin. Gastroenterol. Hepatol. 2018, 2, 6. [Google Scholar] [CrossRef]
- Parrish, N.; Vadlamudi, A.; Goldberg, N. Anaerobic adaptation of Mycobacterium avium subspecies paratuberculosis in vitro: Similarities to M. tuberculosis and differential susceptibility to antibiotics. Gut Pathog. 2017, 9, 34. [Google Scholar] [CrossRef]
- Henderson, P.; Stevens, C. The Role of Autophagy in Crohn’s Disease. Cells 2012, 1, 492–519. [Google Scholar] [CrossRef]
- Santana, P.T.; Rosas, S.L.B.; Ribeiro, B.E.; Marinho, Y.; de Souza, H.S.P. Dysbiosis in Inflammatory Bowel Disease: Pathogenic Role and Potential Therapeutic Targets. Int. J. Mol. Sci. 2022, 23, 3464. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.P.; Proctor, S.D.; Willing, B.P. Koch’s postulates, microbial dysbiosis and inflammatory bowel disease. Clin. Microbiol. Infect. 2016, 22, 594–599. [Google Scholar] [CrossRef]
- Abdellrazeq, G.S.; Elnaggar, M.M.; Bannantine, J.P.; Schneider, D.A.; Souza, C.D.; Hwang, J.; Mahmoud, A.H.A.; Hulubei, V.; Fry, L.M.; Park, K.-T.; et al. A peptide-based vaccine for Mycobacterium avium subspecies paratuberculosis. Vaccine 2019, 37, 2783–2790. [Google Scholar] [CrossRef]
- Liu, Y.; Liao, F. Vaccination therapy for inflammatory bowel disease. Hum. Vaccin. Immunother. 2023, 19, 2259418. [Google Scholar] [CrossRef]
- Edwards, B.D.; Brode, S.K.; Mehrabi, M.; Marras, T.K. Time to Positive Culture Detection Predicts Mycobacterium avium Pulmonary Disease Severity and Treatment Initiation. Ann. Am. Thorac. Soc. 2022, 19, 925–932. [Google Scholar] [CrossRef]
- Zurba, Y.; Gros, B.; Shehab, M. Exploring the Pipeline of Novel Therapies for Inflammatory Bowel Disease; State of the Art Review. Biomedicines 2023, 11, 747. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aitken, J.M.; Aitken, J.E.; Agrawal, G. Mycobacterium avium ssp. paratuberculosis and Crohn’s Disease—Diagnostic Microbiological Investigations Can Inform New Therapeutic Approaches. Antibiotics 2024, 13, 158. https://doi.org/10.3390/antibiotics13020158
Aitken JM, Aitken JE, Agrawal G. Mycobacterium avium ssp. paratuberculosis and Crohn’s Disease—Diagnostic Microbiological Investigations Can Inform New Therapeutic Approaches. Antibiotics. 2024; 13(2):158. https://doi.org/10.3390/antibiotics13020158
Chicago/Turabian StyleAitken, John M., Jack E. Aitken, and Gaurav Agrawal. 2024. "Mycobacterium avium ssp. paratuberculosis and Crohn’s Disease—Diagnostic Microbiological Investigations Can Inform New Therapeutic Approaches" Antibiotics 13, no. 2: 158. https://doi.org/10.3390/antibiotics13020158
APA StyleAitken, J. M., Aitken, J. E., & Agrawal, G. (2024). Mycobacterium avium ssp. paratuberculosis and Crohn’s Disease—Diagnostic Microbiological Investigations Can Inform New Therapeutic Approaches. Antibiotics, 13(2), 158. https://doi.org/10.3390/antibiotics13020158