
Outbreak of Pseudomonas aeruginosa High-Risk Clone ST309 Serotype O11 Featuring blaPER-1 and qnrVC6
 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Isolates
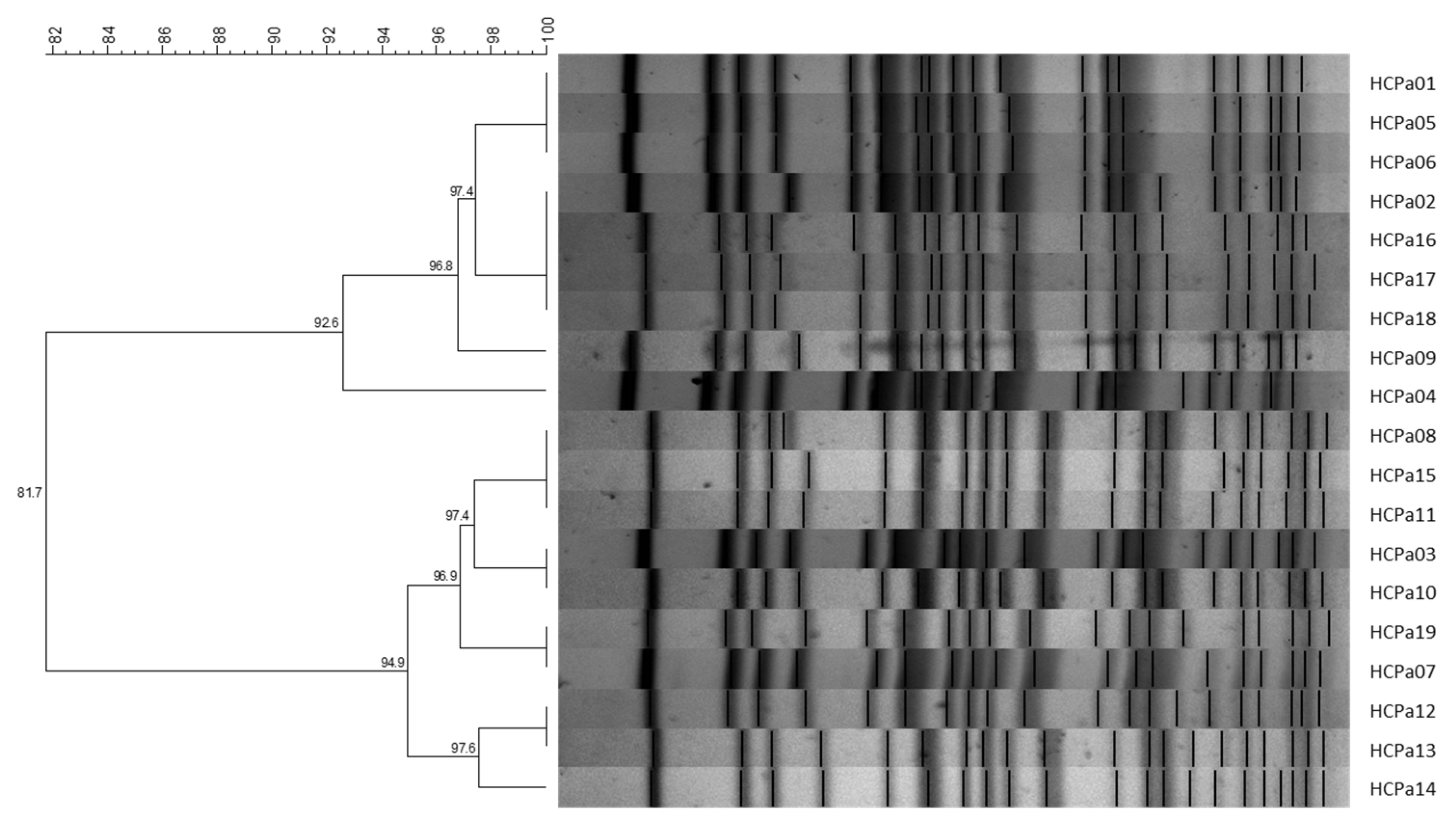
2.2. Pulsed-Field gel Electrophoresis
2.3. Susceptibility Profiles and Mechanisms of Antibiotic Resistance
2.4. Effect of Antibiotics Combined with Rifampicin
2.5. Biofilm Characterization
2.6. Genetic Features
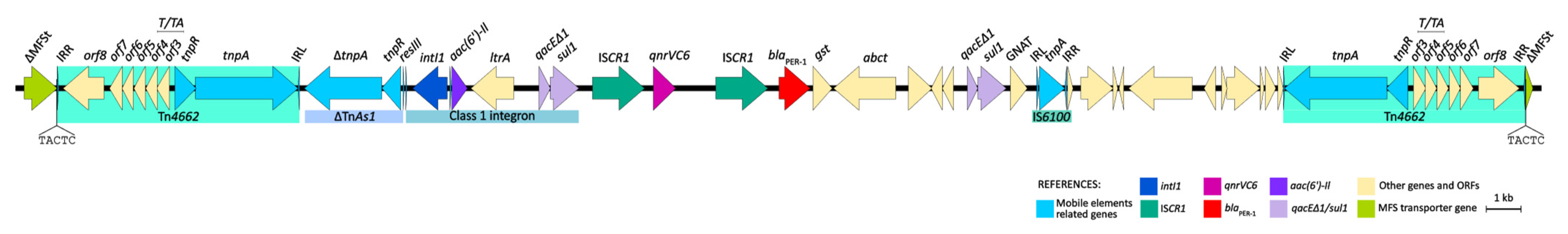
2.7. qnrVC6 and blaPER-1 Genetic Environment
3. Discussion
4. Materials and Methods
4.1. Strains, Identification, and Antibiotic Susceptibility Testing
4.2. Resistance Mechanisms Detection
4.3. Pulse-Field Gel Electrophoresis
4.4. Susceptibility to Combination of Antibiotics
4.5. Biofilm Formation and Antibiotic Susceptibility of Mature Biofilm
4.6. Short- and Long-Read Genome Sequencing
4.7. Sequence Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; The Review on Antimicrobial Resistance Chaired by Jim O’Neill; The Government of the United Kingdom: London, UK, 2016.
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Ikuta, K.S.; Swetschinski, L.R.; Aguilar, G.R.; Sharara, F.; Mestrovic, T.; Gray, A.P.; Weaver, N.D.; Wool, E.E.; Han, C.; Hayoon, A.G.; et al. Global Mortality Associated with 33 Bacterial Pathogens in 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 2221–2248. [Google Scholar] [CrossRef] [PubMed]
- Jian, J.; Yu, P.; Zheng-Li, C.; Hao, L.; Ze-Jing, W.; Shao-Shuo, Y.; Yu, S.; Guang-Yi, W.; Shi-Hui, Z.; Bing, M.; et al. Epidemiological Retrospective Analysis in Major Burn Patients: Single Centre Medical Records from 2009 to 2019. Updates Surg. 2022, 74, 1453–1459. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Xiao, W.; Zhou, C.; Pu, Q.; Deng, X.; Lan, L.; Liang, H.; Song, X.; Wu, M. Pseudomonas aeruginosa: Pathogenesis, Virulence Factors, Antibiotic Resistance, Interaction with Host, Technology Advances and Emerging Therapeutics. Signal Transduct. Target. Ther. 2022, 7, 199. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Silva, V.; López, M.; Rojo-Bezares, B.; Carvalho, J.A.; Castro, A.P.; Sáenz, Y.; Igrejas, G.; Poeta, P. Antimicrobial Resistance, Genetic Lineages, and Biofilm Formation in Pseudomonas aeruginosa Isolated from Human Infections: An Emerging One Health Concern. Antibiotics 2023, 12, 1248. [Google Scholar] [CrossRef] [PubMed]
- del Barrio-Tofiño, E.; López-Causapé, C.; Oliver, A. Pseudomonas aeruginosa Epidemic High-Risk Clones and Their Association with Horizontally-Acquired β-Lactamases: 2020 Update. Int. J. Antimicrob. Agents 2020, 56, 106196. [Google Scholar] [CrossRef]
- Fonseca, É.L.; Morgado, S.M.; Caldart, R.V.; Freitas, F.; Vicente, A.C.P. Emergence of a VIM-2-Producing Extensively Drug-Resistant (XDR) Pseudomonas aeruginosa ST309 in South America: A Comparative Genomic Analysis. Int. J. Antimicrob. Agents 2022, 59, 106507. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Tran, T.T.; Rios, R.; Hanson, B.; Shropshire, W.C.; Sun, Z.; Diaz, L.; Dinh, A.Q.; Wanger, A.; Ostrosky-Zeichner, L.; et al. Extensively Drug-Resistant Pseudomonas aeruginosa ST309 Harboring Tandem Guiana Extended Spectrum β-Lactamase Enzymes: A Newly Emerging Threat in the United States. Open Forum Infect. Dis. 2019, 6, ofz273. [Google Scholar] [CrossRef]
- Morales-Espinosa, R.; Delgado, G.; Espinosa, L.F.; Isselo, D.; Méndez, J.L.; Rodriguez, C.; Miranda, G.; Cravioto, A. Fingerprint Analysis and Identification of Strains ST309 as a Potential High Risk Clone in a Pseudomonas aeruginosa Population Isolated from Children with Bacteremia in Mexico City. Front. Microbiol. 2017, 8, 313. [Google Scholar] [CrossRef]
- Tamma, P.D.; Aitken, S.L.; Bonomo, R.A.; Mathers, A.J.; van Duin, D.; Clancy, C.J. Infectious Diseases Society of America 2022 Guidance on the Treatment of Extended-Spectrum β-Lactamase Producing Enterobacterales (ESBL-E), Carbapenem-Resistant Enterobacterales (CRE), and Pseudomonas aeruginosa with Difficult-to-Treat Resistance (DTR-P. aeruginosa). Clin. Infect. Dis. 2022, 75, 187–212. [Google Scholar] [CrossRef]
- Botelho, J.; Grosso, F.; Peixe, L. Antibiotic Resistance in Pseudomonas aeruginosa—Mechanisms, Epidemiology and Evolution. Drug Resist. Updat. 2019, 44, 100640. [Google Scholar] [CrossRef] [PubMed]
- Richard, E.; Darracq, B.; Loot, C.; Mazel, D. Unbridled Integrons: A Matter of Host Factors. Cells 2022, 11, 925. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R. Analysis of Antibiotic Resistance Regions in Gram-Negative Bacteria. FEMS Microbiol. Rev. 2011, 35, 820–855. [Google Scholar] [CrossRef] [PubMed]
- Papa-Ezdra, R.; Bado, I.; Cordeiro, N.; Gutierrez, C.; Hitateguy, P.; Seija, V.; Vignoli, R. VIM-2-Producing Pseudomonas Spp. in Uruguay: Sequence Types, Pulsotypes, and Class 1 Integrons Including New Variable Regions Featuring blaVIM-2 and blaGES-7. Antimicrob. Agents Chemother. 2016, 60, 5620–5622. [Google Scholar] [CrossRef] [PubMed]
- Papa-Ezdra, R.; Cordeiro, N.F.; Outeda, M.; Garcia-Fulgueiras, V.; Araújo, L.; Seija, V.; Ayala, J.A.; Bado, I.; Vignoli, R. Novel Resistance Regions Carrying TnaphA6, blaVIM-2, and blaPER-1, Embedded in an ISPa40-Derived Transposon from Two Multi-Resistant Pseudomonas aeruginosa Clinical Isolates. Antibiotics 2023, 12, 304. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Diener, I.; Zamorano, L.; Peña, C.; Ocampo-Sosa, A.; Cabot, G.; Gómez-Zorrilla, S.; Almirante, B.; Aguilar, M.; Granados, A.; Calbo, E.; et al. Weighting the Impact of Virulence on the Outcome of Pseudomonas aeruginosa Bloodstream Infections. Clin. Microbiol. Infect. 2020, 26, 351–357. [Google Scholar] [CrossRef]
- Fournier, D.; Carrière, R.; Bour, M.; Grisot, E.; Triponney, P.; Muller, C.; Lemoine, J.; Jeannot, K.; Plésiat, P. Mechanisms of Resistance to Ceftolozane/Tazobactam in Pseudomonas aeruginosa: Results of the GERPA Multicenter Study. Antimicrob. Agents Chemother. 2021, 65, e01117-20. [Google Scholar] [CrossRef]
- Torrens, G.; Van Der Schalk, T.E.; Cortes-Lara, S.; Timbermont, L.; Del Barrio-Tofiño, E.; Xavier, B.B.; Zamorano, L.; Lammens, C.; Ali, O.; Ruzin, A.; et al. Susceptibility Profiles and Resistance Genomics of Pseudomonas aeruginosa Isolates from European ICUs Participating in the ASPIRE-ICU Trial. J. Antimicrob. Chemother. 2022, 77, 1862–1872. [Google Scholar] [CrossRef]
- Ortiz De La Rosa, J.M.; Nordmann, P.; Poirel, L. ESBLs and Resistance to Ceftazidime/Avibactam and Ceftolozane/Tazobactam Combinations in Escherichia coli and Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2019, 74, 1934–1939. [Google Scholar] [CrossRef]
- Polse, R.F.; Khalid, H.M.; Mero, W.M.S. Distribution of blaOXA-10, blaPER-1, and blaSHV Genes in ESBL-Producing Pseudomonas aeruginosa Strains Isolated from Burn Patients. Sci. Rep. 2023, 13, 18402. [Google Scholar] [CrossRef] [PubMed]
- Babouee Flury, B.; Bösch, A.; Gisler, V.; Egli, A.; Seiffert, S.N.; Nolte, O.; Findlay, J. Multifactorial Resistance Mechanisms Associated with Resistance to Ceftazidime-Avibactam in Clinical Pseudomonas aeruginosa Isolates from Switzerland. Front. Cell. Infect. Microbiol. 2023, 13, 1098944. [Google Scholar] [CrossRef]
- Ruiz, J. Transferable Mechanisms of Quinolone Resistance from 1998 Onward. Clin. Microbiol. Rev. 2019, 32, e00007-19. [Google Scholar] [CrossRef] [PubMed]
- Hernández-García, M.; García-Castillo, M.; García-Fernández, S.; López-Mendoza, D.; Díaz-Regañón, J.; Romano, J.; Pássaro, L.; Paixão, L.; Cantón, R. Presence of Chromosomal CrpP-like Genes Is Not Always Associated with Ciprofloxacin Resistance in Pseudomonas aeruginosa Clinical Isolates Recovered in ICU Patients from Portugal and Spain. Microorganisms 2021, 9, 388. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, S.M.; Tolmasky, E.M. Aminoglycoside Modifing Enzymes. Drug Resist. Updat. 2011, 13, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Mulet, X.; Cabot, G.; Ocampo-Sosa, A.A.; Domínguez, M.A.; Zamorano, L.; Juan, C.; Tubau, F.; Rodríguez, C.; Moyà, B.; Peña, C.; et al. Biological Markers of Pseudomonas aeruginosa Epidemic High-Risk Clones. Antimicrob. Agents Chemother. 2013, 57, 5527–5535. [Google Scholar] [CrossRef] [PubMed]
- Zimmerli, W.; Sendi, P. Role of Rifampin against Staphylococcal Biofilm Infections In Vitro, in Animal Models, and in Orthopedic-Device-Related Infections. Antimicrob. Agents Chemother. 2019, 63, e01746-18. [Google Scholar] [CrossRef]
- Hu, Y.-F.; Liu, C.-P.; Wang, N.-Y.; Shih, S.-C. In Vitro Antibacterial Activity of Rifampicin in Combination with Imipenem, Meropenem and Doripenem against Multidrug-Resistant Clinical Isolates of Pseudomonas aeruginosa. BMC Infect. Dis. 2016, 16, 444. [Google Scholar] [CrossRef]
- Mikalauskas, A.; Parkins, M.D.; Poole, K. Rifampicin Potentiation of Aminoglycoside Activity against Cystic Fibrosis Isolates of Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2017, 72, 3349–3352. [Google Scholar] [CrossRef]
- Nicolas, E.; Lambin, M.; Dandoy, D.; Galloy, C.; Nguyen, N.; Oger, C.A.; Hallet, B. The Tn3-Family of Replicative Transposons. Microbiol. Spectr. 2015, 3, MDNA3-0060-2014. [Google Scholar] [CrossRef]
- Martak, D.; Gbaguidi-Haore, H.; Meunier, A.; Valot, B.; Conzelmann, N.; Eib, M.; Autenrieth, I.B.; Slekovec, C.; Tacconelli, E.; Bertrand, X.; et al. High Prevalence of Pseudomonas aeruginosa Carriage in Residents of French and German Long-Term Care Facilities. Clin. Microbiol. Infect. 2022, 28, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Abdouchakour, F.; Grau, D.; Aujoulat, F.; Mournetas, P.; Parer, S.; Gibert, P.; Valcarcel, J.; Jumas-bilak, E. Leads to Successive Waves of Contamination of Water in Dental Care. Appl. Environ. Microbiol. 2015, 81, 7509–7524. [Google Scholar] [CrossRef]
- Chilam, J.; Argimon, S.; Limas, M.; Masim, M.; Gayeta, J.; Lagrada, M.; Olorosa, A.; Cohen, V.; Hernandez, L.; Jeffrey, B.; et al. Genomic Surveillance of Pseudomonas aeruginosa in the Philippines, 2013–2014. West. Pacific Surveill. Response J. 2021, 12, 4–18. [Google Scholar] [CrossRef]
- Nasrin, S.; Hegerle, N.; Sen, S.; Nkeze, J.; Sen, S.; Permala-Booth, J.; Choi, M.; Sinclair, J.; Tapia, M.D.; Johnson, J.K.; et al. Distribution of Serotypes and Antibiotic Resistance of Invasive Pseudomonas aeruginosa in a Multi-Country Collection. BMC Microbiol. 2022, 22, 13. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Eggimann, P.; Luyt, C.E.; Wolff, M.; Tamm, M.; François, B.; Mercier, E.; Garbino, J.; Laterre, P.F.; Koch, H.; et al. Pseudomonas aeruginosa Serotypes in Nosocomial Pneumonia: Prevalence and Clinical Outcomes. Crit. Care 2014, 18, R17. [Google Scholar] [CrossRef] [PubMed]
- The Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022. [Google Scholar]
- Pagani, L.; Mantengoli, E.; Migliavacca, R.; Nucleo, E.; Pollini, S.; Spalla, M.; Romero, E.; Rossolini, G.M.; Daturi, R. Multifocal Detection of Multidrug-Resistant Pseudomonas aeruginosa Producing the PER-1 Extended-Spectrum β -Lactamase in Northern Italy. J. Clin. Microbiol. 2004, 42, 2523–2529. [Google Scholar] [CrossRef] [PubMed]
- Papa-Ezdra, R.; Bado, I.; Caiata, L.; Vignoli, R.; Seija, V. First Report of Pseudomonas aeruginosa Co-Harbouring blaVIM-2 and blaPER-1 in Latin America. J. Glob. Antimicrob. Resist. 2018, 15, 121–122. [Google Scholar] [CrossRef]
- Liu, M.; Wong, M.H.Y.; Chen, S. Molecular Characterisation of a Multidrug Resistance Conjugative Plasmid from Vibrio parahaemolyticus. Int. J. Antimicrob. Agents 2013, 42, 575–579. [Google Scholar] [CrossRef]
- PulseNet International Standard Operating Procedure for PulseNet PFGE of Escherichia coli O157:H7, Escherichia coli Non-O157 (STEC), Salmonella Serotypes, Shigella sonnei and Shigella flexneri. Code PNL05. Available online: https://pulsenetinternational.org/assets/PulseNet/uploads/pfge/PNL05_Ec-Sal-ShigPFGEprotocol.pdf (accessed on 1 November 2023).
- Singh, A.; Goering, R.V.; Simjee, S.; Foley, S.L.; Zervos, M.J. Application of Molecular Techniques to the Study of Hospital Infection. Clin. Microbiol. Rev. 2006, 19, 512–530. [Google Scholar] [CrossRef]
- Synergism Testing: Broth Microdilution Checkerboard and Broth Macrodilution Methods. In Clinical Microbiology Procedures Handbook; ASM Press: Washington, DC, USA, 2016; pp. 5.16.1–5.16.23.
- González, M.J.; Robino, L.; Iribarnegaray, V.; Zunino, P.; Scavone, P. Effect of Different Antibiotics on Biofilm Produced by Uropathogenic Escherichia coli Isolated from Children with Urinary Tract Infection. Pathog. Dis. 2017, 75, ftx053. [Google Scholar] [CrossRef]
- Villegas, N.A.; Baronetti, J.; Albesa, I.; Polifroni, R.; Parma, A.; Etcheverría, A.; Becerra, M.; Padola, N.; Paraje, M. Relevance of Biofilms in the Pathogenesis of Shiga-Toxin-Producing Escherichia coli Infection. Sci. World J. 2013, 2013, 607258. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the Reference Gene Catalog Facilitate Examination of the Genomic Links among Antimicrobial Resistance, Stress Response, and Virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef] [PubMed]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of Microbial Genomes Using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, 206–214. [Google Scholar] [CrossRef]
- Carver, T.; Berriman, M.; Tivey, A.; Patel, C.; Böhme, U.; Barrell, B.G.; Parkhill, J.; Rajandream, M.A. Artemis and ACT: Viewing, Annotating and Comparing Sequences Stored in a Relational Database. Bioinformatics 2008, 24, 2672–2676. [Google Scholar] [CrossRef]
- Tansirichaiya, S.; Rahman, M.A.; Roberts, A.P. The Transposon Registry. Mob. DNA 2019, 10, 40. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient ID/Strain ‡ | Date of 1st Isolate (dd/mm/yyyy) | Sample/s (in Chronological Order) | Genes (PCR) | Minimum Inhibitory Concentration (mg/L) | CZA (E-Test) | C/T (E-Test) | FDC (DD) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PTZ | CAZ | FEP | IPM | MEM | GM | AK | CIP | |||||||
| HCPa01 | 11/8/2021 | Blood culture, catheter tip | blaPER-1/qnrVC | 32 (I) | ≥64 (R) | ≥32 (R) | ≥16 (R) | 8 (R) | 4 (S) | ≥64 (R) | 0.25 (S) | 16 (R) | >256 (R) | S |
| HCPa02 | 27/8/2021 | Respiratory secretions, blood culture, bronchoalveolar lavage | blaPER-1/qnrVC | 32 (I) | ≥64 (R) | ≥32 (R) | 8 (R) | ≤0.25 (S) | 4 (S) | ≥64 (R) | 0.5 (S) | 24 (R) | >256 (R) | S |
| HCPa03 | 29/8/2021 | Respiratory secretions, blood culture | blaPER-1/qnrVC | 32 (I) | ≥64 (R) | ≥32 (R) | 8 (R) | 1 (S) | 4 (S) | ≥64 (R) | 1 (R) | 24 (R) | >256 (R) | S |
| HCPa04 | 30/8/2021 | Blood culture | blaPER-1/qnrVC | 8 (S) | ≥64 (R) | ≥32 (R) | 2 (S) | 0.5 (S) | 4 (S) | ≥64 (R) | 0.5 (S) | 8 (S) | >256 (R) | S |
| HCPa05 | 3/10/2021 | Respiratory secretions, blood culture, catheter tip | blaPER-1 | 16 (S) | ≥64 (R) | ≥32 (R) | ≥16 (R) | 0.5 (S) | 4 (S) | ≥64 (R) | 0.25 (S) | 16 (R) | >256 (R) | S |
| HCPa06 | 12/10/2021 | Respiratory secretions | blaPER-1/qnrVC | 16 (S) | ≥64 (R) | ≥32 (R) | 1 (S) | ≤0.25 (S) | 4 (S) | ≥64 (R) | 0.5 (S) | 16 (R) | >256 (R) | S |
| HCPa07 | 20/10/2021 | Respiratory secretions | blaPER-1/qnrVC | 8 (S) | ≥64 (R) | ≥32 (R) | 2 (S) | 1 (S) | 8 (I) | ≥64 (R) | 0.25 (S) | 8 (S) | 128 (R) | S |
| HCPa08 | 10/12/2021 | Respiratory secretions | blaPER-1 | 16 (S) | ≥64 (R) | ≥32 (R) | 2 (S) | 0.5 (S) | 4 (S) | ≥64 (R) | 0.25 (S) | 8 (S) | >256 (R) | S |
| HCPa09 | 9/1/2022 | Respiratory secretions | blaPER-1 | 32 (I) | ≥64 (R) | ≥32 (R) | 2 (S) | ≤0.25 (S) | 4 (S) | ≥64 (R) | 1 (R) | 16 (R) | >256 (R) | S |
| HCPa10 | 11/1/2022 | Blood culture, respiratory secretions | blaPER-1/qnrVC | 16 (S) | ≥64 (R) | ≥32 (R) | 2 (S) | 0.5 (S) | 8 (I) | ≥64 (R) | 0.25 (S) | 16 (R) | >256 (R) | S |
| HCPa11 | 21/1/2022 | Respiratory secretions | blaPER-1 | 16 (S) | ≥64 (R) | ≥32 (R) | 2 (S) | 0.5 (S) | 4 (S) | ≥64 (R) | 0.25 (S) | 16 (R) | 128 (R) | S |
| HCPa12 | 21/1/2022 | Urine, blood culture, respiratory secretions | blaPER-1 | 16 (S) | ≥64 (R) | ≥32 (R) | 2 (S) | 2 (S) | 4 (S) | ≥64 (R) | ≤0.06 (S) | 16 (R) | >256 (R) | S |
| HCPa13 | 24/1/2022 | Surgical wound | blaPER-1 | 16 (S) | ≥64 (R) | ≥32 (R) | 2 (S) | ≤0.25 (S) | 4 (S) | ≥64 (R) | ≤0.06 (S) | 32 (R) | >256 (R) | S |
| HCPa14 | 12/2/2022 | Respiratory secretions | blaPER-1 | 32 (I) | ≥64 (R) | ≥32 (R) | 1 (S) | ≤0.25 (S) | ≥16 (R) | ≥64 (R) | ≤0.06 (S) | 16 (R) | >256 (R) | S |
| HCPa15 | 13/3/2022 | Respiratory secretions, blood culture | blaPER-1/qnrVC | 16 (S) | ≥64 (R) | ≥32 (R) | 2 (S) | 1 (S) | 4 (S) | ≥64 (R) | 0.25 (S) | 8 (S) | >256 (R) | S |
| HCPa16 | 17/3/2022 | Bronchoalveolar lavage | blaPER-1/qnrVC | 16 (S) | ≥64 (R) | ≥32 (R) | 2 (S) | ≤0.25 (S) | 4 (S) | ≥64 (R) | 0.25 (S) | 16 (R) | >256 (R) | S |
| HCPa17 | 27/3/2022 | Respiratory secretions, surgical wound, bronchoalveolar lavage, urine | blaPER-1 | 16 (S) | ≥64 (R) | ≥32 (R) | 2 (S) | 1 (S) | 4 (S) | ≥64 (R) | 0.25 (S) | 16 (R) | >256 (R) | S |
| HCPa18 | 7/4/2022 | Respiratory secretions | blaPER-1/qnrVC | 16 (S) | ≥64 (R) | ≥32 (R) | 2 (S) | ≤0.25 (S) | 8 (I) | ≥64 (R) | 0.125 (S) | 32 (R) | >256 (R) | S |
| HCPa19 | 17/7/2022 | Respiratory secretions, catheter tip | blaPER-1/qnrVC | 16 (S) | ≥64 (R) | ≥32 (R) | 2 (S) | ≤0.25 (S) | 4 (S) | ≥64 (R) | ≥4 (R) | 8 (S) | 48 (R) | S |
| MIC † | HCPa01 | HCPa12 | MIC | HCPa01 | HCPa12 | MIC | HCPa01 | HCPa12 | MIC | HCPa01 | HCPa12 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RDS | 16 | 16 | RDS | 16 | 16 | RDS | 16 | 16 | RDS | 16 | 16 | ||
| MEMS | 8 | 2 | GMS | 4 | 4 | AKS | 64 | 64 | CIPS | 0.25 | 0.06 | ||
| RDC | 4 | 2 | 4 | RDC | 4 | 4 | RDC | 4 | 4 | 0.5 | RDC | 8 | 8 |
| MEMC | 2 | 0.5 | 0.06 | GMC | 0.5 | 0.5 | AKC | 8 | 8 | 16 | CIPC | 0.06 | 0.03 |
| FICI § | 0.5 | 0.375 | 0.28 | FICI | 0.375 | 0.375 | FICI | 0.375 | 0.375 | 0.28 | FICI | 0.74 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papa-Ezdra, R.; Outeda, M.; Cordeiro, N.F.; Araújo, L.; Gadea, P.; Garcia-Fulgueiras, V.; Seija, V.; Bado, I.; Vignoli, R. Outbreak of Pseudomonas aeruginosa High-Risk Clone ST309 Serotype O11 Featuring blaPER-1 and qnrVC6. Antibiotics 2024, 13, 159. https://doi.org/10.3390/antibiotics13020159
Papa-Ezdra R, Outeda M, Cordeiro NF, Araújo L, Gadea P, Garcia-Fulgueiras V, Seija V, Bado I, Vignoli R. Outbreak of Pseudomonas aeruginosa High-Risk Clone ST309 Serotype O11 Featuring blaPER-1 and qnrVC6. Antibiotics. 2024; 13(2):159. https://doi.org/10.3390/antibiotics13020159
Chicago/Turabian StylePapa-Ezdra, Romina, Matilde Outeda, Nicolás F. Cordeiro, Lucía Araújo, Pilar Gadea, Virginia Garcia-Fulgueiras, Verónica Seija, Inés Bado, and Rafael Vignoli. 2024. "Outbreak of Pseudomonas aeruginosa High-Risk Clone ST309 Serotype O11 Featuring blaPER-1 and qnrVC6" Antibiotics 13, no. 2: 159. https://doi.org/10.3390/antibiotics13020159
APA StylePapa-Ezdra, R., Outeda, M., Cordeiro, N. F., Araújo, L., Gadea, P., Garcia-Fulgueiras, V., Seija, V., Bado, I., & Vignoli, R. (2024). Outbreak of Pseudomonas aeruginosa High-Risk Clone ST309 Serotype O11 Featuring blaPER-1 and qnrVC6. Antibiotics, 13(2), 159. https://doi.org/10.3390/antibiotics13020159