
Synergistic Effects of Capric Acid and Colistin against Colistin-Susceptible and Colistin-Resistant Enterobacterales

 ,
,
Abstract
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Oliveira, D.; Forde, B.M.; Kidd, T.J.; Harris, P.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef]
- Du, H.; Chen, L.; Tang, Y.W.; Kreiswirth, B.N. Emergence of the mcr-1 colistin resistance gene in carbapenem-resistant Enterobacteriaceae. Lancet Infect. Dis. 2016, 16, 287–288. [Google Scholar] [CrossRef]
- Dortet, L.; Cuzon, G.; Ponties, V.; Nordmann, P. Trends in carbapenemase-producing Enterobacteriaceae, France, 2012 to 2014. Eurosurveillance 2017, 22, 30461. [Google Scholar] [CrossRef]
- Schuelter-Trevisol, F.; Schmitt, G.J.; Araujo, J.M.; Souza, L.B.; Nazario, J.G.; Januario, R.L.; Mello, R.S.; Trevisol, D.J. New Delhi metallo-beta-lactamase-1-producing Acinetobacter spp. infection: Report of a survivor. Rev. Soc. Bras. Med. Trop. 2016, 49, 130–134. [Google Scholar] [CrossRef]
- Livermore, D.M.; Warner, M.; Mushtaq, S.; Doumith, M.; Zhang, J.; Woodford, N. What remains against carbapenem-resistant Enterobacteriaceae? Evaluation of chloramphenicol, ciprofloxacin, colistin, fosfomycin, minocycline, nitrofurantoin, temocillin and tigecycline. Int. J. Antimicrob. Agents 2011, 37, 415–419. [Google Scholar] [CrossRef]
- Abdul, R.N.; Cheah, S.E.; Johnson, M.D.; Yu, H.; Sidjabat, H.E.; Boyce, J.; Butler, M.S.; Cooper, M.A.; Fu, J.; Paterson, D.L.; et al. Synergistic killing of NDM-producing MDR Klebsiella pneumoniae by two ‘old’ antibiotics-polymyxin B and chloramphenicol. J. Antimicrob. Chemother. 2015, 70, 2589–2597. [Google Scholar] [CrossRef]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Nang, S.C.; Li, J.; Velkov, T. The rise and spread of mcr plasmid-mediated polymyxin resistance. Crit. Rev. Microbiol. 2019, 45, 131–161. [Google Scholar] [CrossRef]
- Hussein, N.H.; Al-Kadmy, I.; Taha, B.M.; Hussein, J.D. Mobilized colistin resistance (mcr) genes from 1 to 10: A comprehensive review. Mol. Biol. Rep. 2021, 48, 2897–2907. [Google Scholar] [CrossRef]
- Macnair, C.R.; Stokes, J.M.; Carfrae, L.A.; Fiebig-Comyn, A.A.; Coombes, B.K.; Mulvey, M.R.; Brown, E.D. Overcoming mcr-1 mediated colistin resistance with colistin in combination with other antibiotics. Nat. Commun. 2018, 9, 458. [Google Scholar] [CrossRef]
- Hussein, M.; Schneider-Futschik, E.K.; Paulin, O.; Allobawi, R.; Crawford, S.; Zhou, Q.T.; Hanif, A.; Baker, M.; Zhu, Y.; Li, J.; et al. Effective Strategy Targeting Polymyxin-Resistant Gram-Negative Pathogens: Polymyxin B in Combination with the Selective Serotonin Reuptake Inhibitor Sertraline. ACS Infect. Dis. 2020, 6, 1436–1450. [Google Scholar] [CrossRef]
- Falagas, M.E.; Voulgaris, G.L.; Tryfinopoulou, K.; Giakkoupi, P.; Kyriakidou, M.; Vatopoulos, A.; Coates, A.; Hu, Y. Synergistic activity of colistin with azidothymidine against colistin-resistant Klebsiella pneumoniae clinical isolates collected from inpatients in Greek hospitals. Int. J. Antimicrob. Agents 2019, 53, 855–858. [Google Scholar] [CrossRef]
- Zhou, Y.F.; Liu, P.; Dai, S.H.; Sun, J.; Liu, Y.H.; Liao, X.P. Activity of Tigecycline or Colistin in Combination with Zidovudine against Escherichia coli Harboring tet(X) and mcr-1. Antimicrob. Agents Chemother. 2020, 65, e01172-20. [Google Scholar] [CrossRef]
- Martinez-Guitian, M.; Vazquez-Ucha, J.C.; Odingo, J.; Parish, T.; Poza, M.; Waite, R.D.; Bou, G.; Wareham, D.W.; Beceiro, A. Synergy between Colistin and the Signal Peptidase Inhibitor MD3 Is Dependent on the Mechanism of Colistin Resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 4375–4379. [Google Scholar] [CrossRef]
- Chung, J.H.; Bhat, A.; Kim, C.J.; Yong, D.; Ryu, C.M. Combination therapy with polymyxin B and netropsin against clinical isolates of multidrug-resistant Acinetobacter baumannii. Sci. Rep. 2016, 6, 28168. [Google Scholar] [CrossRef]
- Dai, C.; Wang, Y.; Sharma, G.; Shen, J.; Velkov, T.; Xiao, X. Polymyxins-Curcumin Combination Antimicrobial Therapy: Safety Implications and Efficacy for Infection Treatment. Antioxidants 2020, 9, 506. [Google Scholar] [CrossRef]
- Feng, X.; Liu, S.; Wang, Y.; Zhang, Y.; Sun, L.; Li, H.; Wang, C.; Liu, Y.; Cao, B. Synergistic Activity of Colistin Combined With Auranofin Against Colistin-Resistant Gram-Negative Bacteria. Front. Microbiol. 2021, 12, 676414. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef]
- McGaw, L.J.; Jäger, A.K.; Van Staden, J.; Houghton, P.J. Antibacterial effects of fatty acids and related compounds from plants. South Afr. J. Bot. 2002, 68, 417–423. [Google Scholar] [CrossRef]
- Li, L.; Chen, H.; Liu, Y.; Xu, S.; Wu, M.; Liu, Z.; Qi, C.; Zhang, G.; Li, J.; Huang, X. Synergistic effect of linezolid with fosfomycin against Staphylococcus aureus in vitro and in an experimental Galleria mellonella model. J. Microbiol. Immunol. Infect. 2020, 53, 731–738. [Google Scholar] [CrossRef]
- Huang, W.C.; Tsai, T.H.; Chuang, L.T.; Li, Y.Y.; Zouboulis, C.C.; Tsai, P.J. Anti-bacterial and anti-inflammatory properties of capric acid against Propionibacterium acnes: A comparative study with lauric acid. J. Dermatol. Sci. 2014, 73, 232–240. [Google Scholar] [CrossRef]
- Murzyn, A.; Krasowska, A.; Stefanowicz, P.; Dziadkowiec, D.; Bukaszewicz, M. Capric acid secreted by S. boulardii inhibits C. albicans filamentous growth, adhesion and biofilm formation. PLoS ONE 2010, 5, e12050. [Google Scholar] [CrossRef]
- Vidaillac, C.; Benichou, L.; Duval, R.E. In vitro synergy of colistin combinations against colistin-resistant Acinetobacter baumannii, Pseudomonas aeruginosa, and Klebsiella pneumoniae isolates. Antimicrob. Agents Chemother. 2012, 56, 4856–4861. [Google Scholar] [CrossRef]
- Betts, J.W.; Sharili, A.S.; La Ragione, R.M.; Wareham, D.W. In Vitro Antibacterial Activity of Curcumin-Polymyxin B Combinations against Multidrug-Resistant Bacteria Associated with Traumatic Wound Infections. J. Nat. Prod. 2016, 79, 1702–1706. [Google Scholar] [CrossRef]
- Pillai, S.K.; Moellering, R.C.; Eliopoulos, G.M. Antimicrobial combinations. In Antibiotics in Laboratory Medicine, 5th ed.; Lorian, V., Ed.; The Lippincott Williams & Wilkins Co.: Philadelphia, PA, USA, 2005; pp. 365–440. [Google Scholar]
- Bergen, P.J.; Smith, N.M.; Bedard, T.B.; Bulman, Z.P.; Cha, R.; Tsuji, B.T. Rational Combinations of Polymyxins with Other Antibiotics. Adv. Exp. Med. Biol. 2019, 1145, 251–288. [Google Scholar] [CrossRef]
- Suchodolski, J.; Derkacz, D.; Bernat, P.; Krasowska, A. Capric acid secreted by Saccharomyces boulardii influences the susceptibility of Candida albicans to fluconazole and amphotericin B. Sci. Rep. 2021, 11, 6519. [Google Scholar] [CrossRef]


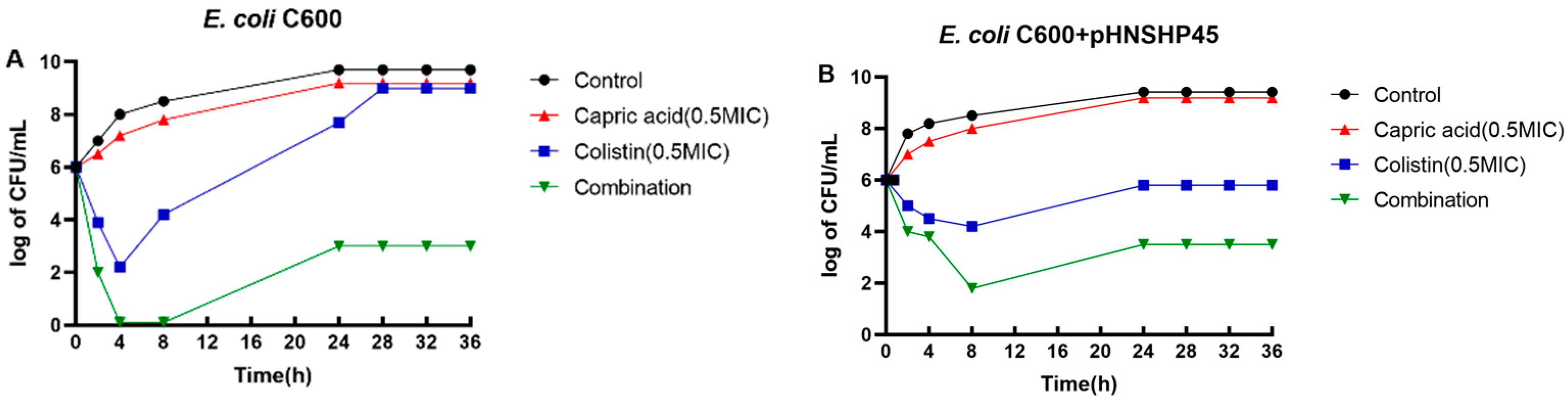{kind=link}
| Strain | Origin | PCR for mcr-1 a | MIC (mg/L) | FICI b | |||
|---|---|---|---|---|---|---|---|
| Single Drug | Combination | ||||||
| Colistin | Capric Acid | Colistin | Capric Acid | ||||
| K. pneumoniae P11 | Human | - | 1 | >3200 | 0.13 | 400 | 0.26 |
| K. pneumoniae P11+ pHNSHP45 | Transformant | + | 8 | >3200 | 0.5 | 200 | 0.13 |
| K. pneumoniae 117 | Pig | + | 16 | >3200 | 2 | 400 | 0.26 |
| K. pneumoniae 281 | Pig | + | 16 | >3200 | 1 | 200 | 0.13 |
| K. pneumoniae SDQ8C53R c | Chicken | - | >512 | 3200 | 1 | 50 | 0.02 |
| K. pneumoniae HNJ9C285 c | Chicken | - | >512 | 3200 | 4 | 200 | 0.07 |
| K. pneumoniae HNJ9C245 c | Chicken | - | >512 | 3200 | 2 | 100 | 0.04 |
| E. coli C600 | - | - | 0.5 | >3200 | 0.125 | 200 | 0.26 |
| E. coli C600 + pHNSHP45 | Transformant | + | 8 | >3200 | 0.5 | 200 | 0.13 |
| E. coli 2D-8 d | Pig | - | 8 | >3200 | 0.5 | 200 | 0.13 |
| E. coli SHP7 | Pig | + | 8 | >3200 | 2 | 200 | 0.31 |
| E. coli SHP8 | Pig | + | 8 | >3200 | 1 | 200 | 0.19 |
| E. coli SHP50 | Pig | + | 8 | >3200 | 0.5 | 200 | 0.13 |
| E. coli GDF36 | Fish | + | 8 | 3200 | 1 | 800 | 0.38 |
| Salmonella SA316 | Pig | + | 8 | >3200 | 1 | 400 | 0.26 |
| Salmonella SH271 | Pig | + | 4 | 3200 | 1 | 50 | 0.27 |
| Salmonella SH138 | Pig | + | 8 | >3200 | 1 | 400 | 0.26 |
| Salmonella SH47 | Pig | - | 1 | >3200 | 0.25 | 800 | 0.51 |
| Salmonella SH17 | Pig | + | 4 | 3200 | 1 | 50 | 0.27 |
| P. aeruginosa 10104 | CVCC f | - | 1 | >3200 | 1 | >3200 | 2.00 |
| P. aeruginosa 10104 (R) e | Induced | - | 8 | >3200 | 1 | 400 | 0.26 |
| Strain | Antibiotic | ||||||
|---|---|---|---|---|---|---|---|
| Colistin | Ciprofloxacin | Cefotaxime | Neomycin | Tetracycline | Ampicillin | ||
| E. coli 2D-8 | MIC a | 8 | 0.5 | 0.06 | 256 | 32 | 128 |
| FICI b | 0.13 | 2 | 2 | 1 | 1 | 1 | |
| E. coli SHP50 | MIC a | 8 | 32 | 0.5 | 0.25 | 128 | 8 |
| FICI b | 0.19 | 1 | 2 | 1 | 1 | 1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.-Y.; Qin, Z.-H.; Yue, H.-Y.; Bergen, P.J.; Deng, L.-M.; He, W.-Y.; Zeng, Z.-L.; Peng, X.-F.; Liu, J.-H. Synergistic Effects of Capric Acid and Colistin against Colistin-Susceptible and Colistin-Resistant Enterobacterales. Antibiotics 2023, 12, 36. https://doi.org/10.3390/antibiotics12010036
Liu Y-Y, Qin Z-H, Yue H-Y, Bergen PJ, Deng L-M, He W-Y, Zeng Z-L, Peng X-F, Liu J-H. Synergistic Effects of Capric Acid and Colistin against Colistin-Susceptible and Colistin-Resistant Enterobacterales. Antibiotics. 2023; 12(1):36. https://doi.org/10.3390/antibiotics12010036
Chicago/Turabian StyleLiu, Yi-Yun, Zong-Hua Qin, Hui-Ying Yue, Phillip J. Bergen, Li-Min Deng, Wan-Yun He, Zhen-Ling Zeng, Xian-Feng Peng, and Jian-Hua Liu. 2023. "Synergistic Effects of Capric Acid and Colistin against Colistin-Susceptible and Colistin-Resistant Enterobacterales" Antibiotics 12, no. 1: 36. https://doi.org/10.3390/antibiotics12010036
APA StyleLiu, Y.-Y., Qin, Z.-H., Yue, H.-Y., Bergen, P. J., Deng, L.-M., He, W.-Y., Zeng, Z.-L., Peng, X.-F., & Liu, J.-H. (2023). Synergistic Effects of Capric Acid and Colistin against Colistin-Susceptible and Colistin-Resistant Enterobacterales. Antibiotics, 12(1), 36. https://doi.org/10.3390/antibiotics12010036