
Potentiation of Antibiotic Action and Efflux Pump Inhibitory Effect on Staphylococcus aureus Strains by Solasodine

 , , , , ,
, , , , , 
Abstract
1. Introduction
2. Results
2.1. Antibacterial Activity
2.2. Antibiotic Activity Modifier Action
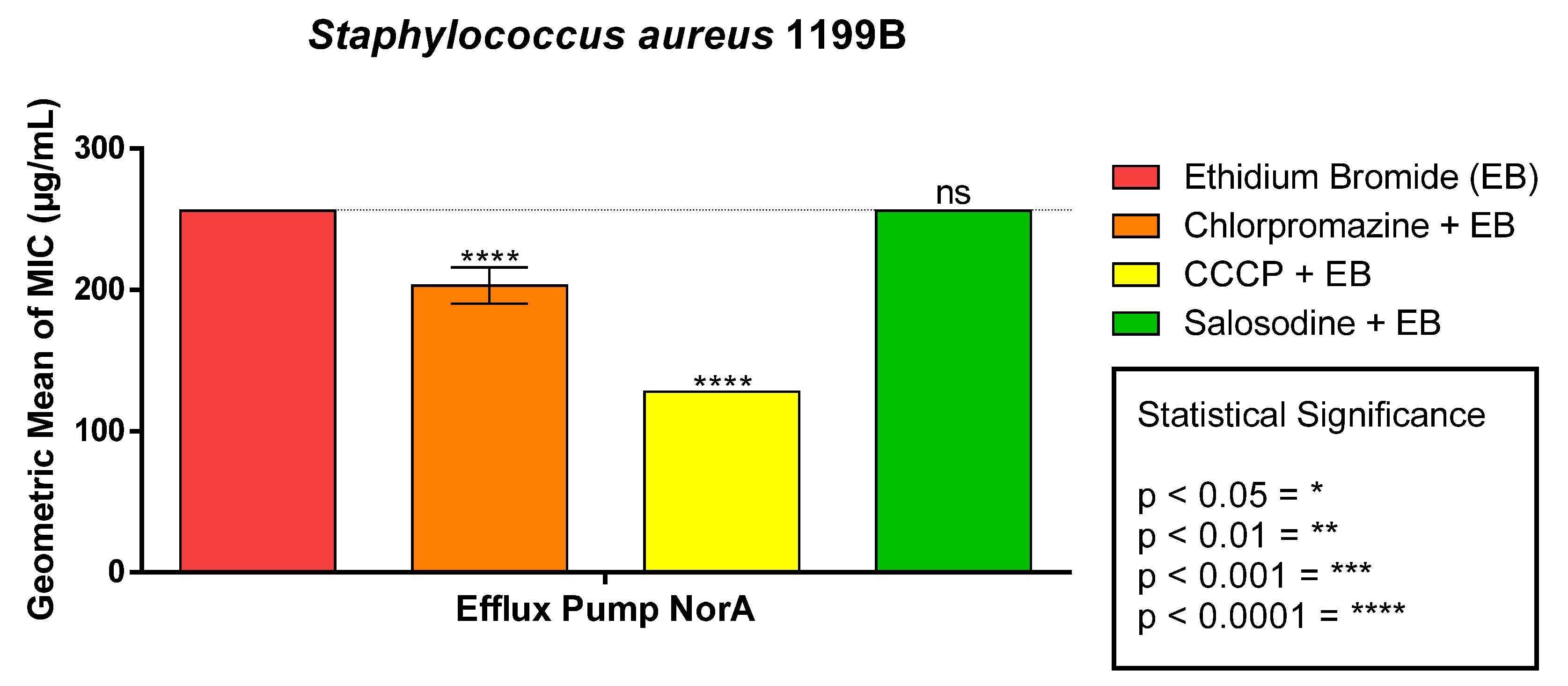
2.3. Efflux Pump Inhibitory Effect Nora E Mepa
3. Discussion
4. Materials and Methods
4.1. Substances, Antibiotics and Reagents
4.2. Bacterial Strains
4.3. Determination of the Minimum Inhibitory Concentration (MIC)
4.4. Evaluation of Modifying Antibiotic Activity Action
4.5. Evaluation of Efflux Pump Inhibitory Effect Nora E Mepa
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yu, H.; Han, X.; Quiñones Pérez, D. La humanidad enfrenta un desastre: La resistencia antimicrobiana. Rev. Habanera Cienc. Médicas 2021, 20, e3850. [Google Scholar]
- Brito, G.B.; Trevisan, M. O uso indevido de antibióticos e o eminente risco de resistência bacteriana. Rev. Artigos. Com. 2021, 30, e7902. [Google Scholar]
- Alves, J.W.S. Antibióticos e mecanismos de resistência bacteriana: Uma questão de saúde pública. Rev. Multidiscip. Saúde 2021, 2, 38. [Google Scholar]
- Bhardwaj, K.A.; Mohanty, P. Bacterial efflux pumps involved in multidrug resistance and their inhibitors: Rejuvinating the antimicrobial chemotherapy. Recent Pat. Antiinfect. Drug Discov. 2012, 7, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Schindler, B.D.; Jacinto, P.; Kaatz, G.W. Inhibition of drug efflux pumps in Staphylococcus aureus: Current status of potentiating existing antibiotics. Future Microbiol. 2013, 8, 491–507. [Google Scholar] [CrossRef] [PubMed]
- Kaatz, G.W.; Mcaleese, F.; Seo, S.M. Multidrug resistance in Staphylococcus aureus due to overexpression of a novel multidrug and toxin extrusion (MATE) transport protein. Antimicrob. Agents Chemother. 2005, 49, 1857–1864. [Google Scholar] [CrossRef]
- Bhattarai, K.; Baral, B. Next-generation microbial drugs developed from microbiome’s natural products. Adv. Genet. 2021, 108, 341–382. [Google Scholar]
- Coutinho, H.D.M.; Brito, S.M.O.; Leite, N.F.; Vandesmet, V.C.S.; Oliveira, M.T.A.; Martins, G.M.A.B.; Silva, A.R.P.; Costa, M.S. Avaliação comparativa da modulação de antibióticos, frente às cepas bacterianas de Escherichia coli, Staphylococcus aureus. Rev. Cienc. Salud 2015, 13, 345–354. [Google Scholar] [CrossRef][Green Version]
- Tintino, S.R.; Oliveira-Tintino, C.D.; Campina, F.F.; Limaverde, P.W.; Pereira, P.S.; Siqueira-Junior, J.P.; Coutinho, H.D.M.; Quintans-Júnior, L.J.; Silva, T.G.; Leal-Balbino, T.C.; et al. Vitamin K enhances the effect of antibiotics inhibiting the efflux pumps of Staphylococcus aureus strains. Med. Chem. Res. 2018, 27, 261–267. [Google Scholar] [CrossRef]
- Jastrzebska, I.; Morzycki, J.M. Some observations on solasodine reactivity. Steroids 2017, 127, 13–17. [Google Scholar] [CrossRef]
- Kumar, R.; Khan, M.I.; Prasad, M. Solasodine: A Perspective on their roles in Health and Disease. Res. J. Pharm. Technol. 2019, 12, 2571–2576. [Google Scholar] [CrossRef]
- Qu, M.; Xeu, P.; Zhang, Q.; Lu, T.; Liu, K.; Hu, B.; Pang, J.; Xiao, Q.; Xu, T.; Wang, Q.; et al. Pharmacokinetics, oral bioavailability and metabolic analysis of solasodine in mice by dried blood spotLC-MS/MS and UHPLC-Q-Exactive MS. J. Pharm. Biom. Anal. 2022, 210, 114–542. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.E.F.; Oliveira, N.C.C.; Ferrer, M.E.A.; Ferrer, M.T. Estudio de las hojas y frutos de Solanum monachophyllum Dunal.(jurubebina) como una fuente de compuestos esteroidales. Rev. Cuba. Plantas Med. 2022, 26, 968–982. [Google Scholar]
- Pendharkar, G.B.; Benerjee, T.; Patil, S.; Dalal, K.S.; Chaudhari, B.L. Biotransformation of Industrially Important Steroid Drug Precursors. In Industrial Microbiology and Biotechnology; Springer: Singapore, 2022; pp. 307–333. [Google Scholar]
- Chauhan, K.; Sheth, N.; Ranpariya, V.; Parmar, S. Anticonvulsant activity of solasodine isolated from Solanum sisymbriifolium fruits in rodents. Pharm. Biol. 2011, 49, 194–199. [Google Scholar] [CrossRef]
- Pandurangan, A.; Khosa, R.L.; Hemalatha, S. Anti-inflammatory activity of an alkaloid from Solanum trilobatum on acute and chronic inflammation models. Nat. Prod. Res. 2011, 25, 1132–1141. [Google Scholar] [CrossRef]
- Sharma, T.; Airao, V.; Panara, N.; Vaishnav, D.; Ranpariya, V.; Sheth, N.; Parmar, S. Solasodine protects rat brain against ischemia/reperfusion injury through its antioxidant activity. Eur. J. Pharmacol. 2014, 15, 40–46. [Google Scholar] [CrossRef]
- Ashal, H.A. Biosynthesis and biological activities of in vitro derived solasodine glycoalkaloids from Solanum laciniatum. Nat. Prod. J. 2017, 7, 199–207. [Google Scholar]
- Li, Y.; Chang, W.; Zhang, M.; Lou, H. Natural Product Solasodine3-O-ß-glucopyranoside inhibits the virulence factors of Candida albicans. FEMS Yeast Res. 2015, 15, fov060. [Google Scholar] [CrossRef][Green Version]
- Chang, W.; Li, Y.; Zhang, M.; Zheng, S.; Li, Y.; Lou, H. Solasodine-3-O-β-d-glucopyranoside kills Candida albicans by disrupting the intracellular vacuole. Food Chem. Toxicol. 2017, 106, 139–146. [Google Scholar] [CrossRef]
- Hameed, A.; Ijaz, S.; Mohammad, I.S.; Muhammad, K.S.; Akhtar, N.; Khan, H.M.S. Aglycone solanidine and solasodine derivatives: A natural approach towards cancer. Biomed. Pharmacother. 2017, 94, 446–457. [Google Scholar] [CrossRef]
- Zhuang, Y.W.; Wu, C.; Zhou, J.Y.; Chen, X.; Wu, J.; Jiang, S.; Peng, H.Y.; Zou, X.; Liu, J.Y.; Wu, D.P.; et al. Solasodine inhibits human colorectal cancer cells through suppression of the AKT/glycogen synthase kinase-3β/β-catenin pathway. Câncer Sci. 2017, 108, 2248–2264. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Li, Z.; Wu, L.; Lin, F.; Shao, J.; Ma, X.; Yao, Y.; Zhuang, W.; Wang, Y. Solasodine, Isolated from Solanum sisymbriifolium Fruits, has a Potent Anti-Tumor Activity Against Pancreatic Cancer. Drug Des. Devel. Ther. 2021, 13, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Almazinia, M.A.I.; Abbas, H.G.; Abdul-Amer, A. Antibacterial activity of the solasodine of Solanum nigrum against bacterial isolates from the wounds. Basrah J. Vet. Res. 2009, 8, 137–147. [Google Scholar]
- Niño, J.; Correa, Y.M.; Mosqueram, O.M. Biological activities of steroidal alkaloids isolated from Solanum leucocarpum. Pharm. Biol. 2009, 47, 255–259. [Google Scholar] [CrossRef]
- Bibon, M.B. Antibacterial in-vitro evaluation of phenotypically screened solasodine from Solanum nigrum Linn. against enterohemorrhagic Escherichia coli (0157:H7). Int. J. Sci. Res. Multidiscip. Stud. 2021, 7, 43–48. [Google Scholar]
- Tortora, G.J.; Funke, B.R.; Case, C.L. Microbiologia, 10th ed.; Artmed: Porto Alegre, Brazil, 2012; pp. 1–939. [Google Scholar]
- Soltani, R.; Fazeli, H.; Najafi, R.B.; Jelokhanian, A. Evaluation of the synergistic effect of tomatidine with several antibiotics against standard and clinical isolates of Staphylococcus aureus, Enterococcus faecalis, Pseudomonas aeruginosa and Escherichia coli. Iran. J. Pharm. Res. 2017, 16, 290–296. [Google Scholar]
- Siriyong, T.; Voravuthikunchai, S.P.; Coote, P.J. Steroidal alkaloids and conessine from the medicinal plant Holarrhena antidysenterica restore antibiotic efficacy in a Galleria mellonella model of multidrug-resistant Pseudomonas aeruginosa infection. BMC Comp. Altern. Med. 2018, 18, 285. [Google Scholar] [CrossRef]
- Casciaro, B.; Mangiardi, L.; Cappiello, F.; Romeo, I.; Loffredo, M.R.; Iazzetti, A.; Calcaterra, A.; Goggiamani, A.; Ghirga, F.; Mangoni, M.L.; et al. Naturally-Occurring Alkaloids of Plant Origin as Potential Antimicrobials against Antibiotic-Resistant Infections. Molecules 2020, 25, 3619. [Google Scholar] [CrossRef]
- Yan, Y.; Li, X.; Zhang, C.; Lv, L.; Gao, B.; Li, M. Research Progress on Antibacterial Activities and Mechanisms of Natural Alkaloids: A Review. Antibiotics 2021, 10, 318. [Google Scholar] [CrossRef]
- Braquehais, I.D.; Vasconcelos, F.R.; Ribeiro, A.R.C.; Da Silva, A.R.A.; Franca, M.G.A.; De Lima, D.R.; De Paiva, C.F.; Guedes, M.I.F.; Magalhães, F.E.A. Toxicological, antibacterial, and phytochemical preliminary study of the ethanolic extract of Jatropha mollissima (Pohl) Baill (pinhão-bravo, Euphorbiaceae) leaves, collected in Tauá, Ceará, Northeastern Brazil. Rev. Bras. De Plantas Med. 2016, 18, 582–587. [Google Scholar] [CrossRef]
- Araújo, L.L.C.; Azevedo, F.H.C. Study of the beta-lactamase enzyme and its relationship with antibiotic resistance. Res. Soc. Dev. 2020, 9, e663974594. [Google Scholar] [CrossRef]
- Guliu, C.; Green, S.I.; Min, L.; Clark, J.R.; Salazar, C.K.; Terwilliger, A.L.; Kaplan, H.B.; Trautner, R.F.; Ramig, R.F.; Maresso, A.w. Phage-antibiotic synergy is driven by a unique combination of antibacterial mechanism of action and stoichiometry. MBio 2020, 11, e01462-20. [Google Scholar]
- Shrestha, P.; Ni, J.; Wong, T.Y. Synergistic and antagonistic interactions of triclosan with various antibiotics in bacteria. J. Environ. Sci. Health Toxicol. Carcinog. 2020, 38, 187–203. [Google Scholar] [CrossRef]
- Coêlho, M.L.; Ferreira, J.H.L.; Siqueira-Júnior, J.P.; Kaatz, W.G.; Barreto, H.M.; Cavalcante, A.A.C.M. Inhibition of the NorA multi-drug transporter by oxygenated monoterpenes. Microb. Pathog. 2016, 99, 173–177. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement. In CLSI Document M100-S25; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015; Volume 35. [Google Scholar]
- Coutinho, H.D.M.; Falcão-Silva, V.S.; Gonçalves, F.G. Pulmonary bacterial pathogens in cystic fibrosis patients and antibiotic therapy: A tool for the health workers. Int. Arch. Med. 2008, 1, 24. [Google Scholar] [CrossRef]
- Tintino, S.R.; Oliveira-Tintino, C.D.; Campina, F.F.; Silva, R.L.P.; Costa, M.S.; Menezes, I.R.A.; Calixto-Júnior, J.T.; Siqueira-Junior, J.P.; Coutinho, H.D.M.; Leal-Balbino, T.C.; et al. Evaluation of the tannic acid inhibitory effect against the NorA efflux pump of Staphylococcus aureus. Microb. Pathog. 2016, 97, 9–13. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | S.A 10 | P.A 24 | E.C 06 | S.A K2068 | S.A 1199B | S.A ATCC 25923 | P.A ATCC 9027 | E.C ATCC 25922 |
|---|---|---|---|---|---|---|---|---|
| Solasodine | ≥1024 | ≥1024 | ≥1024 | ≥1024 | 512 | 512 | 512 | 512 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, A.R.P.; Costa, M.d.S.; Araújo, N.J.S.; de Freitas, T.S.; de Almeida, R.S.; Barbosa Filho, J.M.; Tavares, J.F.; de Souza, E.O.; de Farias, P.A.M.; Pinheiro, J.C.A.; et al. Potentiation of Antibiotic Action and Efflux Pump Inhibitory Effect on Staphylococcus aureus Strains by Solasodine. Antibiotics 2022, 11, 1309. https://doi.org/10.3390/antibiotics11101309
da Silva ARP, Costa MdS, Araújo NJS, de Freitas TS, de Almeida RS, Barbosa Filho JM, Tavares JF, de Souza EO, de Farias PAM, Pinheiro JCA, et al. Potentiation of Antibiotic Action and Efflux Pump Inhibitory Effect on Staphylococcus aureus Strains by Solasodine. Antibiotics. 2022; 11(10):1309. https://doi.org/10.3390/antibiotics11101309
Chicago/Turabian Styleda Silva, Ana Raquel Pereira, Maria do Socorro Costa, Nara Juliana Santos Araújo, Thiago Sampaio de Freitas, Ray Silva de Almeida, José Maria Barbosa Filho, Josean Fechine Tavares, Erlânio Oliveira de Souza, Pablo Antonio Maia de Farias, Jacqueline Cosmo Andrade Pinheiro, and et al. 2022. "Potentiation of Antibiotic Action and Efflux Pump Inhibitory Effect on Staphylococcus aureus Strains by Solasodine" Antibiotics 11, no. 10: 1309. https://doi.org/10.3390/antibiotics11101309
APA Styleda Silva, A. R. P., Costa, M. d. S., Araújo, N. J. S., de Freitas, T. S., de Almeida, R. S., Barbosa Filho, J. M., Tavares, J. F., de Souza, E. O., de Farias, P. A. M., Pinheiro, J. C. A., & Coutinho, H. D. M. (2022). Potentiation of Antibiotic Action and Efflux Pump Inhibitory Effect on Staphylococcus aureus Strains by Solasodine. Antibiotics, 11(10), 1309. https://doi.org/10.3390/antibiotics11101309