Strategies for Improving Small-Molecule Biosensors in Bacteria

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
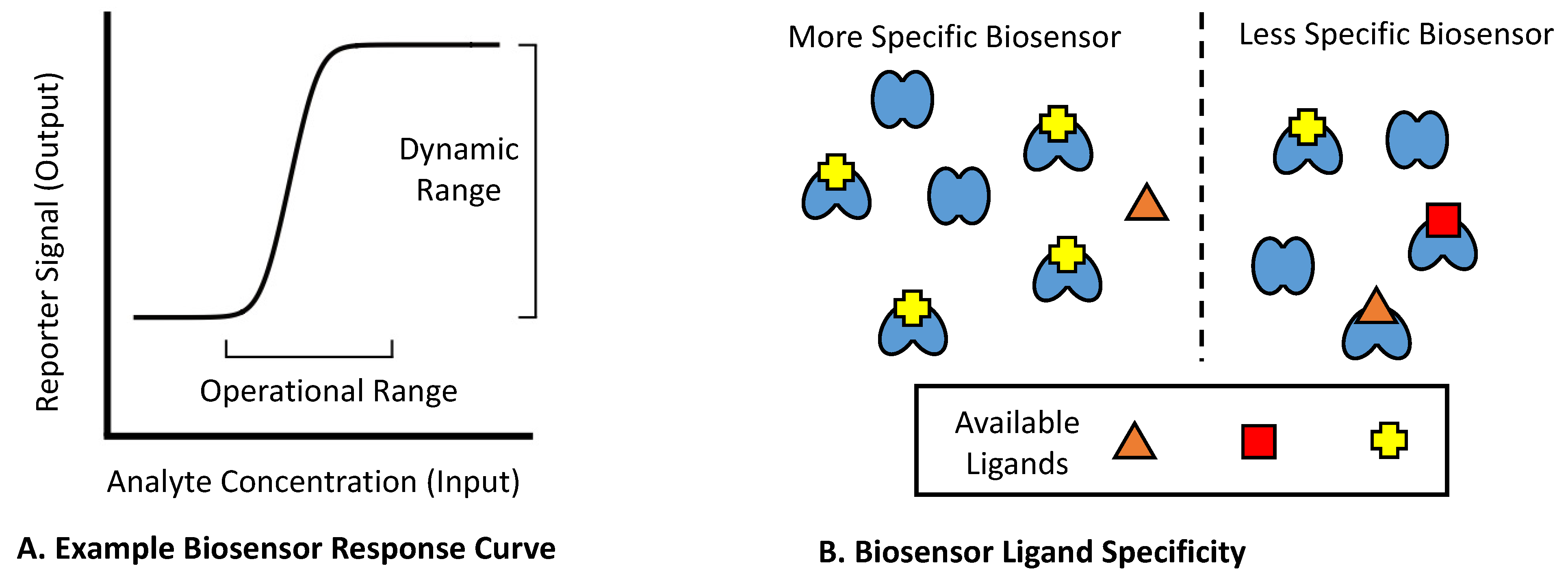
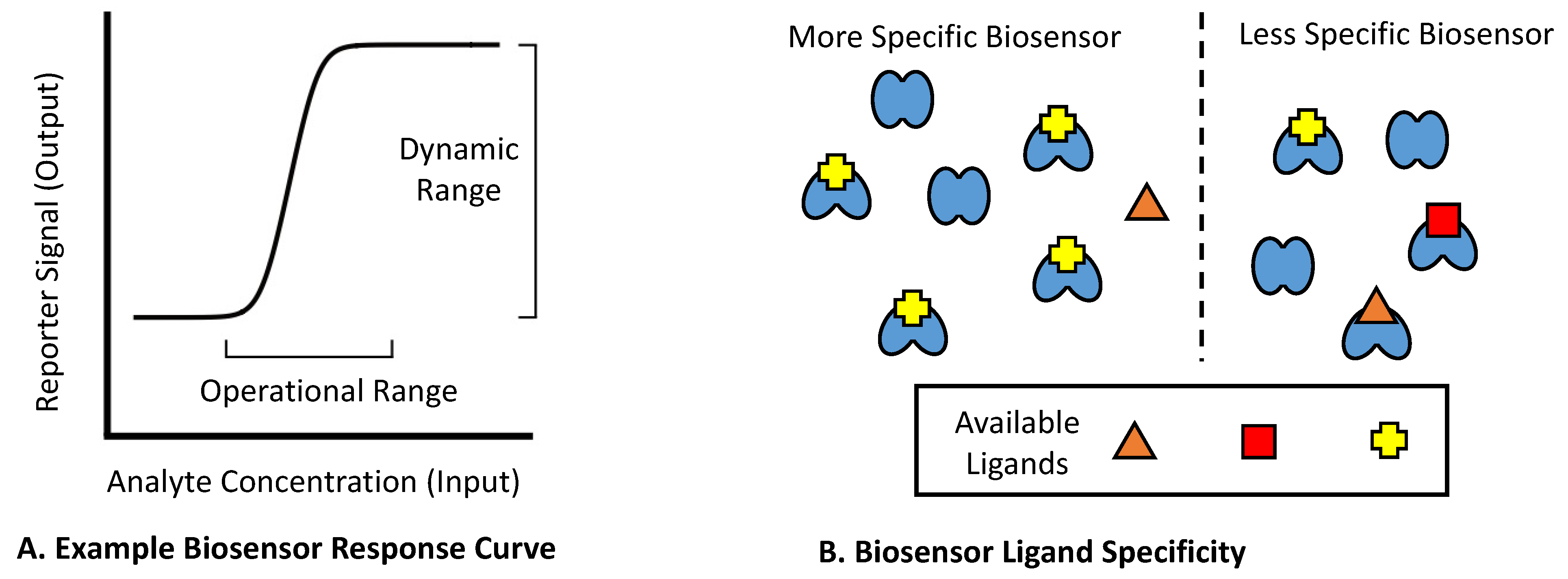
2. Mechanistic Classes of Biosensors within Bacterial Cells
2.1. Known Mechanisms of Bacterial Biosensors
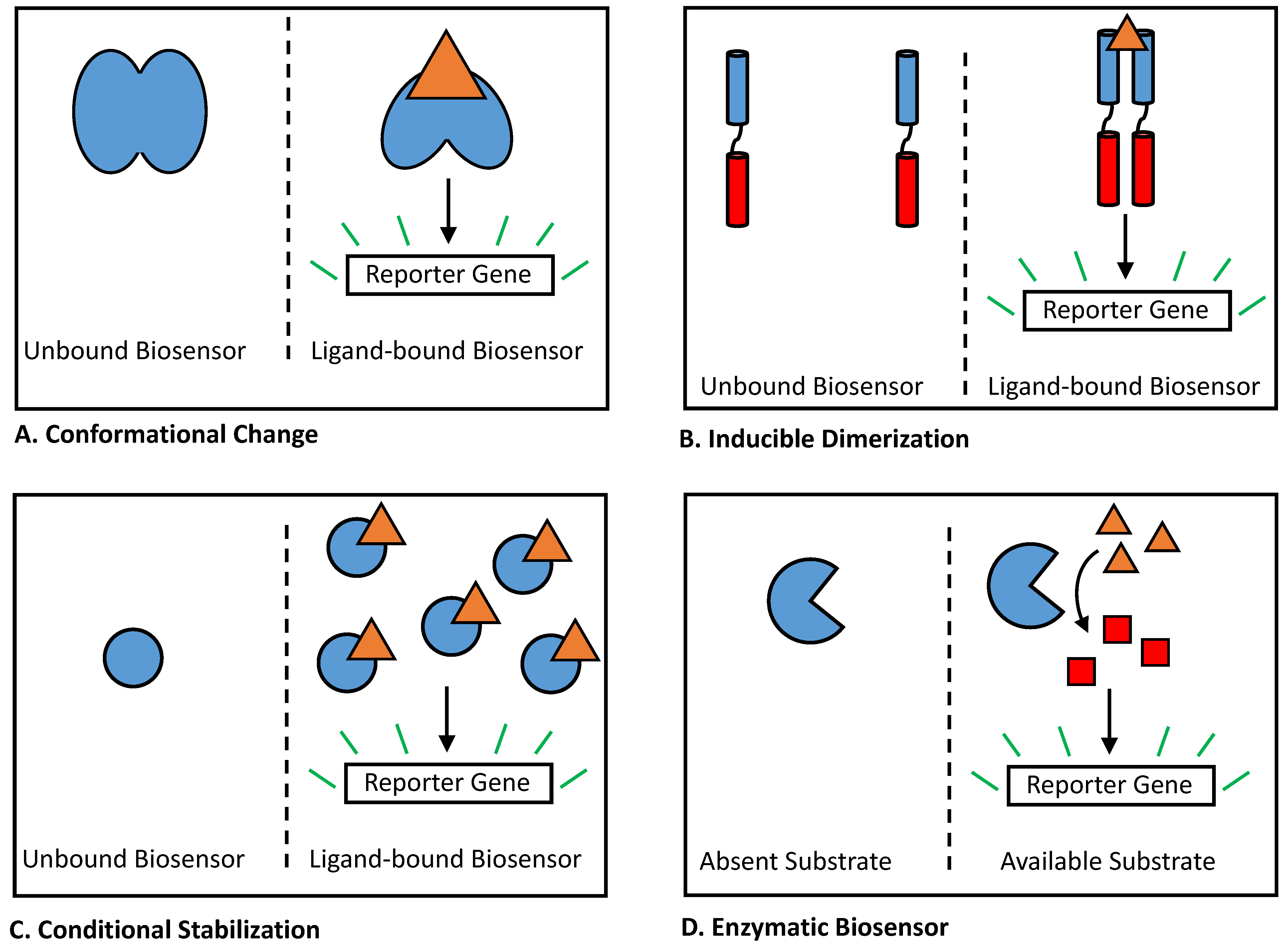
2.2. Biosensors Reliant on Conformational Change
2.3. Biosensors Utilizing Inducible Dimerization
2.4. Conditionally Stabilized Biosensors
2.5. Enzymatic Biosensors
3. Methods for Improving Bacterial Biosensor Properties
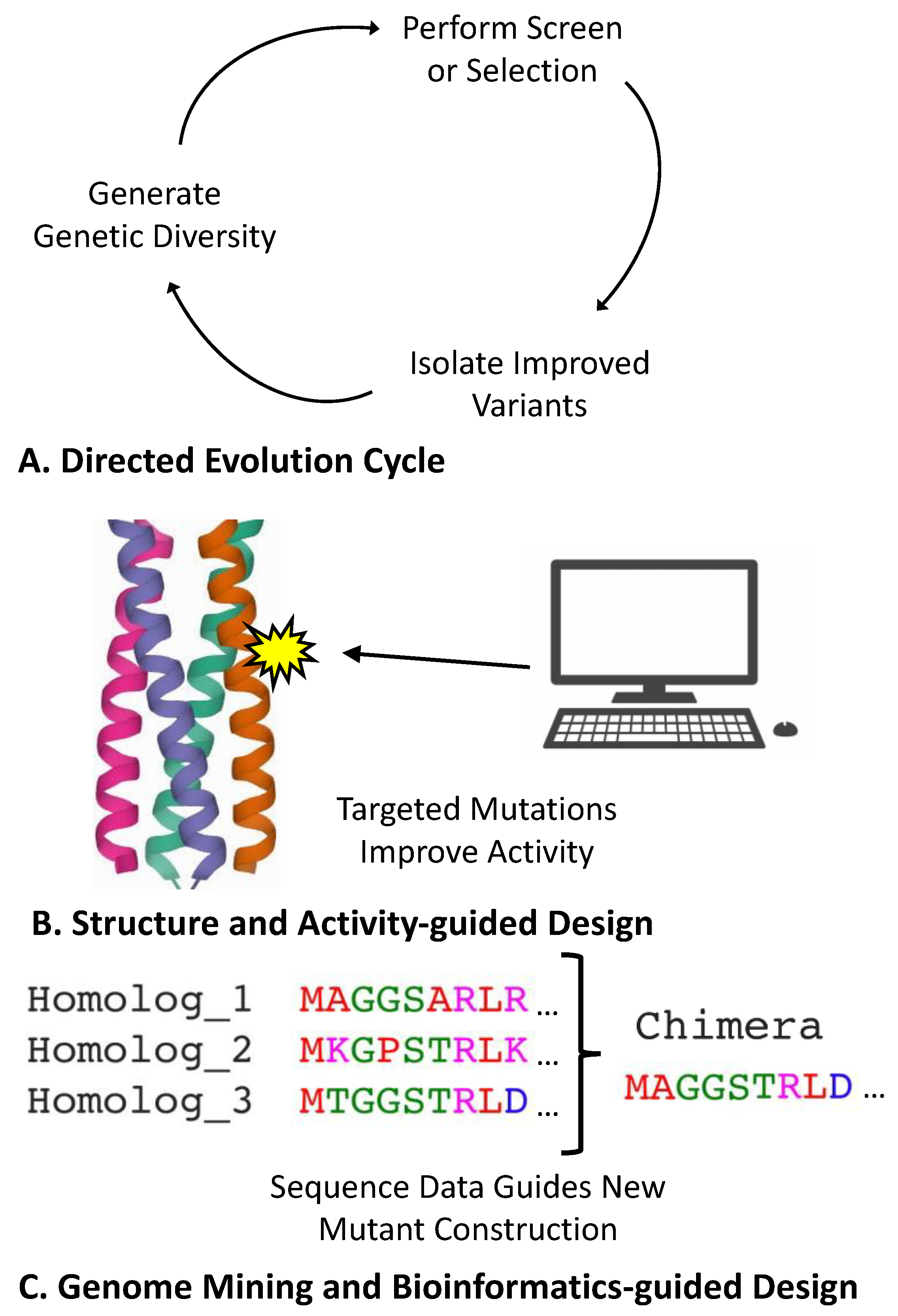
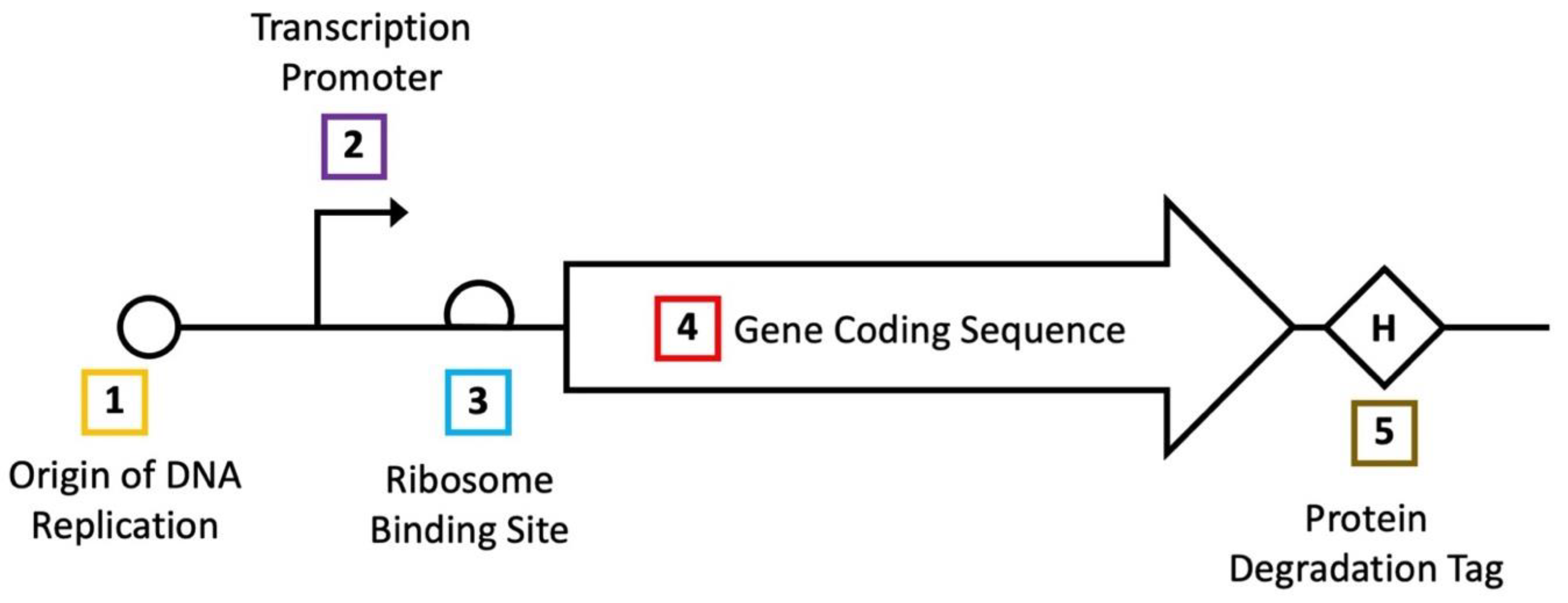
3.1. Direct Engineering of Biosensor Genes
3.2. Optimization of Gene Expression
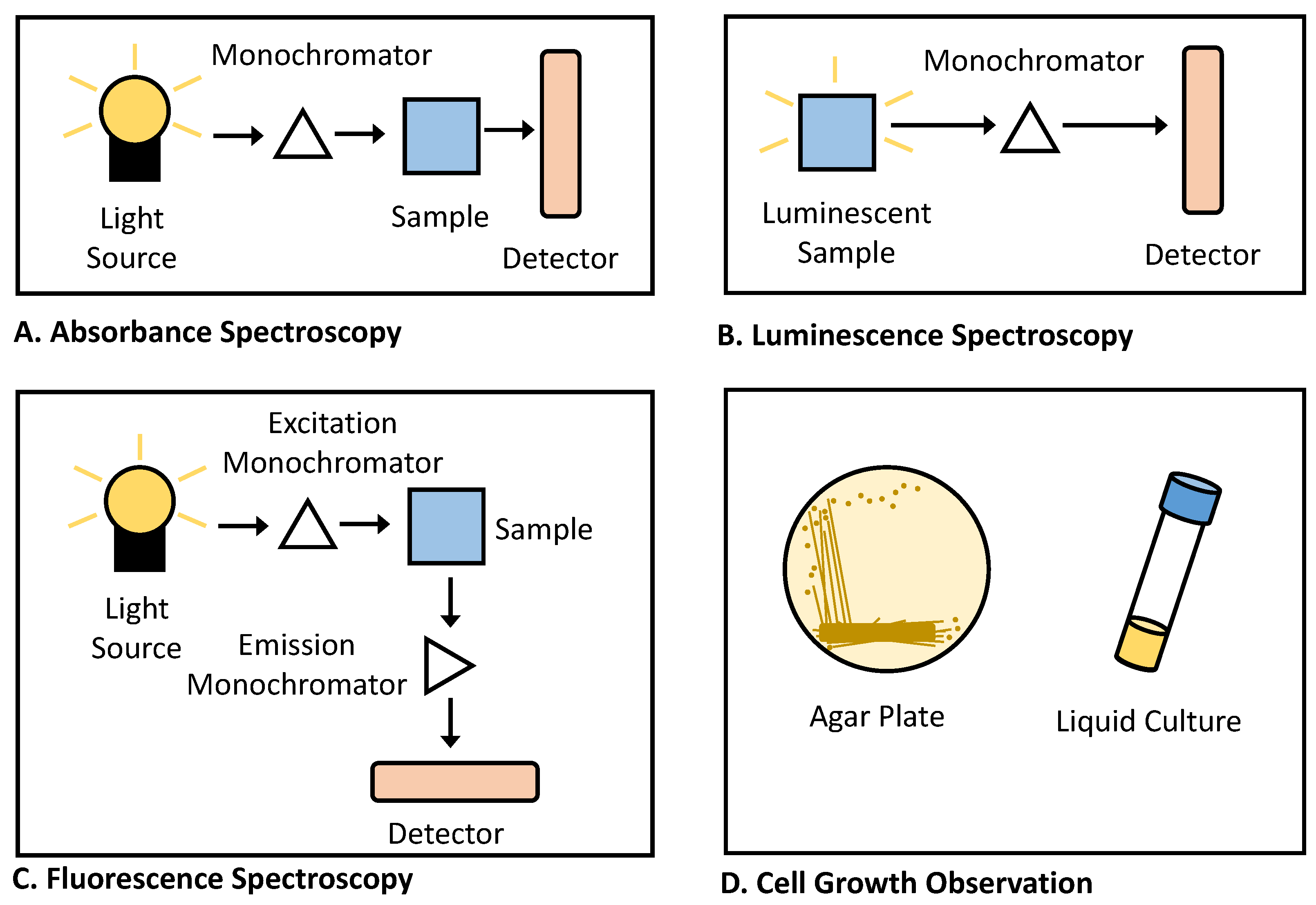
3.3. Selection of Reporter System
3.4. Incorporation of Additional Genetic Modules
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Jacob, F.; Monod, J. Genetic regulatory mechanisms in the synthesis of proteins. J. Mol. Biol. 1961, 3, 318–356. [Google Scholar] [CrossRef]
- Lewis, M. A tale of two repressors—A historical perspective. J. Mol. Biol. 2011, 409, 14. [Google Scholar] [CrossRef]
- Lewis, M. The lac repressor. C. R. Biol. 2005, 328, 521–548. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, A.C.; Paulsen, I.T.; Williams, T.C. Blueprints for Biosensors: Design, Limitations, and Applications. Genes 2018, 9, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigneshvar, S.; Sudhakumari, C.C.; Senthilkumaran, B.; Prakash, H. Recent Advances in Biosensor Technology for Potential Applications—An Overview. Front. Bioeng. Biotechnol. 2016, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Clark, L.C., Jr.; Lyons, C. Electrode systems for continuous monitoring in cardiovascular surgery. Ann. N. Y. Acad. Sci. 1962, 102, 29–45. [Google Scholar] [CrossRef]
- Liu, D.; Xiao, Y.; Evans, B.S.; Zhang, F. Negative feedback regulation of fatty acid production based on a malonyl-CoA sensor-actuator. ACS Synth. Biol. 2015, 4, 132–140. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Wang, M. Design, Optimization and Application of Small Molecule Biosensor in Metabolic Engineering. Front. Microbiol. 2017, 8, 2012. [Google Scholar] [CrossRef] [Green Version]
- Kasey, C.M.; Zerrad, M.; Li, Y.; Cropp, T.A.; Williams, G.J. Development of Transcription Factor-Based Designer Macrolide Biosensors for Metabolic Engineering and Synthetic Biology. ACS Synth. Biol. 2018, 7, 227–239. [Google Scholar] [CrossRef]
- Schallmey, M.; Frunzke, J.; Eggeling, L.; Marienhagen, J. Looking for the pick of the bunch: High-throughput screening of producing microorganisms with biosensors. Curr. Opin. Biotechnol. 2014, 26, 148–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Din, M.O.; Danino, T.; Prindle, A.; Skalak, M.; Selimkhanov, J.; Allen, K.; Julio, E.; Atolia, E.; Tsimring, L.S.; Bhatia, S.N.; et al. Synchronized cycles of bacterial lysis for in vivo delivery. Nature 2016, 536, 81–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, T.; Harimoto, T.; Kepecs, B.; Gray, K.; Coker, C.; Hou, N.; Pu, K.; Azad, T.; Nolasco, A.; Pavlicova, M.; et al. Enhancing the tropism of bacteria via genetically programmed biosensors. Nat. Biomed. Eng. 2021, 6, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Riglar, D.; Giessen, T.; Baym, M.; Kerns, S.J.; Niederhuber, M.J.; Bronson, R.T.; Kotula, J.W.; Gerber, G.K.; Way, J.C.; Silver, P.A. Engineered bacteria can function in the mammalian gut long-term as live diagnostics of inflammation. Nat. Biotechnol. 2017, 35, 653–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Bram, E.; Weiss, R. Genetically Programmable Pathogen Sense and Destroy. ACS Synth. Biol. 2013, 2, 715–723. [Google Scholar] [CrossRef]
- Hwang, I.Y.; Tan, M.H.; Koh, E.; Ho, C.L.; Poh, C.L.; Chang, M.W. Reprogramming Microbes to Be Pathogen-Seeking Killers. ACS Synth. Biol. 2014, 3, 228–237. [Google Scholar] [CrossRef]
- Saeidi, N.; Wong, A.; Lo, T.; Nguyen, H.X.; Ling, H.; Leong, S.S.J.; Poh, C.L.; Chang, M.W. Engineering microbes to sense and eradicate Pseudomonas aeruginosa, a human pathogen. Mol. Syst. Biol. 2011, 7, 521. [Google Scholar] [CrossRef]
- Gui, Q.; Lawson, T.; Shan, S.; Yan, L.; Liu, Y. The Application of Whole Cell-Based Biosensors for Use in Environmental Analysis and in Medical Diagnostics. Sensors 2017, 17, 1623. [Google Scholar] [CrossRef] [Green Version]
- Charbonneau, M.R.; Isabella, V.M.; Li, N.; Kurtz, C.B. Developing a new class of engineered live bacterial therapeutics to treat human diseases. Nat. Commun. 2020, 11, 1738. [Google Scholar] [CrossRef] [Green Version]
- Ejeian, F.; Etedali, P.; Mansouri-Tehrani, H.A.; Soozanipour, A.; Low, Z.X.; Asadnia, M.; Taheri-Kafrani, A.; Razmjou, A. Biosensors for wastewater monitoring: A review. Biosens. Bioelectron. 2018, 118, 66–79. [Google Scholar] [CrossRef]
- Mahbub, K.; Krishnan, K.; Naidu, R.; Megharaj, M. Development of a whole cell biosensor for the detection of inorganic mercury. Environ. Technol. Innov. 2017, 8, 64–70. [Google Scholar] [CrossRef]
- Bereza-Malcolm, L.; Aracic, S.; Franks, A.E. Development and Application of a Synthetically-Derived Lead Biosensor Construct for Use in Gram-Negative Bacteria. Sensors 2016, 16, 2174. [Google Scholar] [CrossRef] [Green Version]
- Willardson, B.M.; Wilkins, J.; Rand, T.A.; Schupp, J.M.; Hill, K.K.; Keim, P.; Jackson, P.J. Development and Testing of a Bacterial Biosensor for Toluene-Based Environmental Contaminants. Appl. Environ. Microbiol. 1998, 64, 1006–1012. [Google Scholar] [CrossRef] [Green Version]
- Tecon, R.; Beggah, S.; Czechowska, K.; Sentchilo, V.; Chronopoulou, P.-M.; McGenity, T.; van der Meer, J.R. Development of a Multistrain Bacterial Bioreporter Platform for the Monitoring of Hydrocarbon Contaminants in Marine Environments. Environ. Sci. Technol. 2010, 44, 1049–1055. [Google Scholar] [CrossRef]
- Jia, X.; Zhang, J.; Sun, W.; He, W.; Jiang, H.; Chen, D.; Murchie, A.I.H. Riboswitch control of aminoglycoside antibiotic resistance. Cell 2013, 152, 68–81. [Google Scholar] [CrossRef] [Green Version]
- DeFraia, C.T.; Schmelz, E.A.; Mou, Z. A rapid biosensor-based method for quantification of free and glucose-conjugated salicylic acid. Plant Methods 2008, 4, 28. [Google Scholar] [CrossRef] [Green Version]
- Bick, M.J.; Greisen, P.J.; Morey, K.J.; Antunes, M.S.; La, D.; Sankaran, B.; Reymond, L.; Johnsson, K.; Medford, J.I.; Baker, D. Computational design of environmental sensors for the potent opioid fentanyl. eLife 2017, 6, e28909. [Google Scholar] [CrossRef]
- Sonntag, C.K.; Flachbart, L.K.; Maass, C.; Vogt, M.; Marienhagen, J. A unified design allows fine-tuning of biosensor parameters and application across bacterial species. Metab. Eng. Commun. 2020, 11, e00150. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.K.; Taylor, N.D.; Church, G.M. Biosensor-based engineering of biosynthetic pathways. Curr. Opin. Biotechnol. 2016, 42, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Doshi, A.; Chowdhury, R.; Wang, Y.; Maranas, C.D.; Cirino, P.C. Engineering sensitivity and specificity of AraC-based biosensors responsive to triacetic acid lactone and orsellinic acid. Protein Eng. Des. Sel. 2020, 33, gzaa027. [Google Scholar] [CrossRef]
- Reshetilov, A.N.; Iliasov, P.V.; Fesay, A.P.; Ivashchenko, G.V.; Taranova, L.A.; Winther-Nielsen, M.; Emnéus, J. Evaluation of substrate specificity of biosensor models based on strains degrading polycyclic aromatic compounds. Appl. Biochem. Microbiol. 2005, 41, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Mannan, A.A.; Liu, D.; Zhang, F.; Oyarzún, D.A. Fundamental Design Principles for Transcription-Factor-Based Metabolite Biosensors. ACS Synth. Biol. 2017, 6, 1851–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, J.-G.; Miao, Y.-Q.; Zhang, Q.-J. Impedimetric biosensors. J. Biosci. Bioeng. 2004, 97, 219–226. [Google Scholar] [CrossRef]
- Davis, G. Electrochemical techniques for the development of amperometric biosensors. Biosensors 1985, 1, 161–178. [Google Scholar] [CrossRef]
- Ciminelli, C.; Dell’Olio, F.; Conteduca, D.; Armenise, M.N. Integrated Photonic and Plasmonic Resonant Devices for Label-Free Biosensing and Trapping at the Nanoscale. Phys. Status Solidi 2019, 216, 1800561. [Google Scholar] [CrossRef]
- Park, M.-K.; Park, J.W.; Wikle, H.; Chin, B.A. Evaluation of phage-based magnetoelastic biosensors for direct detection of Salmonella Typhimurium on spinach leaves. Sens. Actuators B Chem. 2013, 176, 1134–1140. [Google Scholar] [CrossRef]
- Mannoor, M.S.; James, T.; Ivanov, D.V.; Beadling, L.; Braunlin, W. Nanogap Dielectric Spectroscopy for Aptamer-Based Protein Detection. Biophys. J. 2010, 98, 724–732. [Google Scholar] [CrossRef] [Green Version]
- Conteduca, D.; Brunetti, G.; Dell’Olio, F.; Armenise, M.N.; Krauss, T.F.; Ciminelli, C. Monitoring of individual bacteria using electro-photonic traps. Biomed. Opt. Express 2019, 10, 3463–3471. [Google Scholar] [CrossRef]
- Dante, S.; Duval, D.; Sepulveda, B.; González-Guerrero, A.B.; Sendra, J.R.; Lechuga, L.M. All-optical phase modulation for integrated interferometric biosensors. Opt. Express 2012, 20, 7195–7205. [Google Scholar] [CrossRef]
- Sepúlveda, B.; Del Rio, J.S.; Moreno, M.; Blanco, F.J.; Mayora, K.; Domínguez, C.; Lechuga, L.M. Optical biosensor microsystems based on the integration of highly sensitive Mach–Zehnder interferometer devices. J. Opt. A Pure Appl. Opt. 2006, 8, S561. [Google Scholar] [CrossRef]
- Liu, Q.; Tu, X.; Kim, K.W.; Kee, J.S.; Shin, Y.; Han, K.; Yoon, Y.-J.; Lo, G.-Q.; Park, M.K. Highly sensitive Mach–Zehnder interferometer biosensor based on silicon nitride slot waveguide. Sens. Actuators B Chem. 2013, 188, 681–688. [Google Scholar] [CrossRef]
- Ravcheev, D.A.; Khoroshkin, M.S.; Laikova, O.N.; Tsoy, O.V.; Sernova, N.V.; Petrova, S.A.; Rakhmaninova, A.B.; Novichkov, B.S.; Rodionov, D.A. Comparative genomics and evolution of regulons of the LacI-family transcription factors. Front. Microbiol. 2014, 5, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, J.L.; Bueno, M.M.; Molina-Henares, A.J.; Terán, W.; Watanabe, K.; Zhang, X.; Gallegos, M.-T.; Brennan, R.; Tobes, R. The TetR Family of Transcriptional Repressors. Microbiol. Mol. Biol. Rev. 2005, 69, 326–356. [Google Scholar] [CrossRef] [Green Version]
- Suvorova, I.A.; Korostelev, Y.D.; Gelfand, M.S. GntR Family of Bacterial Transcription Factors and Their DNA Binding Motifs: Structure, Positioning and Co-Evolution. PLoS ONE 2015, 10, e0132618. [Google Scholar] [CrossRef]
- Suvorova, I.A.; Gelfand, M.S. Comparative Analysis of the IclR-Family of Bacterial Transcription Factors and Their DNA-Binding Motifs: Structure, Positioning, Co-Evolution, Regulon Content. Front. Microbiol. 2021, 12, 1479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jensen, M.K.; Keasling, J.D. Development of biosensors and their application in metabolic engineering. Curr. Opin. Chem. Biol. 2015, 28, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; He, F.; Yu, Y.; Wang, Y. Application of FRET Biosensors in Mechanobiology and Mechanopharmacological Screening. Front. Bioeng. Biotechnol. 2020, 8, 1299. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.-M.; Fu, T.-F.; Lin, C.-S.; Chiu, T.-Y.; Huang, C.-C.; Huang, H.-Y.; Chung, M.-W.; Lin, Y.-S.; Manurung, R.V.; Nguyen, P.N.N.; et al. High-performance FRET biosensors for single-cell and in vivo lead detection. Biosens. Bioelectron. 2020, 168, 112571. [Google Scholar] [CrossRef]
- Otten, J.; Tenhaef, N.; Jansen, R.P.; Döbber, J.; Jungbluth, L.; Noack, S.; Oldiges, M.; Wiechert, W.; Pohl, M. A FRET-based biosensor for the quantification of glucose in culture supernatants of mL scale microbial cultivations. Microb. Cell Factories 2019, 18, 143. [Google Scholar] [CrossRef] [Green Version]
- Martín, A.S.; Ceballo, S.; Baeza-Lehnert, F.; Lerchundi, R.; Valdebenito, R.; Contreras-Baeza, Y.; Alegría, K.; Barros, L.F. Imaging Mitochondrial Flux in Single Cells with a FRET Sensor for Pyruvate. PLoS ONE 2014, 9, e85780. [Google Scholar] [CrossRef] [Green Version]
- Juárez, J.F.; Lecube-Azpeitia, B.; Brown, S.L.; Johnston, C.D.; Church, G.M. Biosensor libraries harness large classes of binding domains for construction of allosteric transcriptional regulators. Nat. Commun. 2018, 9, 3101. [Google Scholar] [CrossRef] [Green Version]
- Mandal, M.; Breaker, R. Gene regulation by riboswitches. Nat. Rev. Mol. Cell Biol. 2004, 5, 451–463. [Google Scholar] [CrossRef]
- Serganov, A.; Nudler, E. A Decade of Riboswitches. Cell 2013, 152, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Tucker, B.J.; Breaker, R.R. Riboswitches as versatile gene control elements. Curr. Opin. Struct. Biol. 2005, 15, 342–348. [Google Scholar] [CrossRef]
- Breaker, R.R. Riboswitches and Translation Control. Cold Spring Harb. Perspect. Biol. 2018, 10, a032797. [Google Scholar] [CrossRef]
- Zhong, G.; Wang, H.; Bailey, C.C.; Gao, G.; Farzan, M. Rational design of aptazyme riboswitches for efficient control of gene expression in mammalian cells. eLife 2016, 5, e18858. [Google Scholar] [CrossRef]
- Walter, J.-G.; Stahl, F. Aptazymes: Expanding the Specificity of Natural Catalytic Nucleic Acids by Application of In Vitro Selected Oligonucleotides. Mol. Biominer. 2019, 170, 107–119. [Google Scholar] [CrossRef]
- Capra, E.J.; Laub, M.T. Evolution of Two-Component Signal Transduction Systems. Annu. Rev. Microbiol. 2012, 66, 325–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazar, J.T.; Tabor, J.J. Bacterial two-component systems as sensors for synthetic biology applications. Curr. Opin. Syst. Biol. 2021, 28, 100398. [Google Scholar] [CrossRef]
- Gao, R.; Mack, T.R.; Stock, A.M. Bacterial response regulators: Versatile regulatory strategies from common domains. Trends Biochem. Sci. 2007, 32, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-J.; Mayonove, P.; Zavala, A.; De Visch, A.; Minard, P.; Cohen-Gonsaud, M.; Bonnet, J. A Modular Receptor Platform to Expand the Sensing Repertoire of Bacteria. ACS Synth. Biol. 2018, 7, 166–175. [Google Scholar] [CrossRef]
- Chang, H.-J.; Zúñiga, A.; Conejero, I.; Voyvodic, P.L.; Gracy, J.; Fajardo-Ruiz, E.; Cohen-Gonsaud, M.; Cambray, G.; Pageaux, G.-P.; Meszaros, M.; et al. Programmable receptors enable bacterial biosensors to detect pathological biomarkers in clinical samples. Nat. Commun. 2021, 12, 5216. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Yan, M.; Yang, S. Split aptamers and their applications in sandwich aptasensors. TrAC Trends Anal. Chem. 2016, 80, 581–593. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, Q.; Zhang, Y.; Shi, H.-C.; Nguyen, K.T.; Zhou, X. Unconventional Split Aptamers Cleaved at Functionally Essential Sites Preserve Biorecognition Capability. Anal. Chem. 2019, 91, 15811–15817. [Google Scholar] [CrossRef]
- Welz, R.; Schmidt, C.; Müller, S. Spermine Supports Catalysis of Hairpin Ribozyme Variants to Differing Extents. Biochem. Biophys. Res. Commun. 2001, 283, 648–654. [Google Scholar] [CrossRef]
- Hasegawa, S.; Gowrishankar, G.; Rao, J. Detection of mRNA in Mammalian Cells with a Split Ribozyme Reporter. ChemBioChem 2006, 7, 925–928. [Google Scholar] [CrossRef]
- Wu, Y.; Huang, J.; Yang, X.; Yang, Y.; Quan, K.; Xie, N.; Li, J.; Ma, C.; Wang, K. Gold Nanoparticle Loaded Split-DNAzyme Probe for Amplified miRNA Detection in Living Cells. Anal. Chem. 2017, 89, 8377–8383. [Google Scholar] [CrossRef]
- Huang, J.; He, Y.; Yang, X.; Wang, K.; Quan, K.; Lin, X. Split aptazyme-based catalytic molecular beacons for amplified detection of adenosine. Analyst 2014, 139, 2994–2997. [Google Scholar] [CrossRef]
- Ausländer, S.; Fuchs, D.; Hürlemann, S.; Ausländer, D.; Fussenegger, M. Engineering a ribozyme cleavage-induced split fluorescent aptamer complementation assay. Nucleic Acids Res. 2016, 44, e94. [Google Scholar] [CrossRef] [Green Version]
- Debiais, M.; Lelievre, A.; Smietana, M.; Müller, S. Splitting aptamers and nucleic acid enzymes for the development of advanced biosensors. Nucleic Acids Res. 2020, 48, 3400–3422. [Google Scholar] [CrossRef] [PubMed]
- Schuster, M.; Greenberg, E.P. LuxR-Type Proteins in Pseudomonas aeruginosa Quorum Sensing: Distinct Mechanisms with Global Implications. Chem. Commun. Among Bact. 2014, 191, 131–144. [Google Scholar] [CrossRef]
- Bottomley, M.J.; Muraglia, E.; Bazzo, R.; Carfì, A. Molecular Insights into Quorum Sensing in the Human Pathogen Pseudomonas aeruginosa from the Structure of the Virulence Regulator LasR Bound to Its Autoinducer. J. Biol. Chem. 2007, 282, 13592–13600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCready, A.R.; Paczkowski, J.E.; Henke, B.R.; Bassler, B.L. Structural determinants driving homoserine lactone ligand selection in the Pseudomonas aeruginosa LasR quorum-sensing receptor. Proc. Natl. Acad. Sci. USA 2019, 116, 245–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Winans, S.C. The quorum-sensing transcriptional regulator TraR requires its cognate signaling ligand for protein folding, protease resistance, and dimerization. Proc. Natl. Acad. Sci. USA 2001, 98, 1507–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandsen, B.M.; Mattheisen, J.M.; Noel, T.; Fields, S. A Biosensor Strategy for E. coli Based on Ligand-Dependent Stabilization. ACS Synth. Biol. 2018, 7, 1990–1999. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Jester, B.W.; Tinberg, C.E.; Mandell, D.J.; Antunes, M.S.; Chari, R.; Morey, K.J.; Rios, X.; Medford, J.I.; Church, G.M.; et al. A general strategy to construct small molecule biosensors in eukaryotes. eLife 2015, 4, e10606. [Google Scholar] [CrossRef]
- Brunoud, G.; Wells, D.M.; Oliva, M.; Larrieu, A.; Mirabet, V.; Burrow, A.H.; Beeckman, T.; Kepinski, S.; Traas, J.; Bennett, M.J.; et al. A novel sensor to map auxin response and distribution at high spatio-temporal resolution. Nature 2012, 482, 103–106. [Google Scholar] [CrossRef]
- Libis, V.; Delépine, B.; Faulon, J.-L. Expanding Biosensing Abilities through Computer-Aided Design of Metabolic Pathways. ACS Synth. Biol. 2016, 5, 1076–1085. [Google Scholar] [CrossRef]
- Ho, J.C.H.; Pawar, S.V.; Hallam, S.J.; Yadav, V.G. An Improved Whole-Cell Biosensor for the Discovery of Lignin-Transforming Enzymes in Functional Metagenomic Screens. ACS Synth. Biol. 2018, 7, 392–398. [Google Scholar] [CrossRef]
- Aslan, S.; Noor, E.; Vaquerizo, S.B.; Lindner, S.N.; Bar-Even, A. Design and engineering of E. coli metabolic sensor strains with a wide sensitivity range for glycerate. Metab. Eng. 2020, 57, 96–109. [Google Scholar] [CrossRef]
- Adolfsen, K.J.; Callihan, I.; Monahan, C.E.; Greisen, P.J.; Spoonamore, J.; Momin, M.; Fitch, L.E.; Castillo, M.J.; Weng, L.; Renaud, L.; et al. Improvement of a synthetic live bacterial therapeutic for phenylketonuria with biosensor-enabled enzyme engineering. Nat. Commun. 2021, 12, 6215. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Sung, B.H.; Oh, S.-H.; Kwon, K.K.; Lee, H.; Kim, H.; Lee, D.-H.; Yeom, S.-J.; Lee, S.-G. C1 Compound Biosensors: Design, Functional Study, and Applications. Int. J. Mol. Sci. 2019, 20, 2253. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Li, Q.; Wang, Y.; Zhou, W.; Guo, Y.; Chen, J.; Zheng, P.; Sun, J.; Ma, Y. Isoleucyl-tRNA synthetase mutant based whole-cell biosensor for high-throughput selection of isoleucine overproducers. Biosens. Bioelectron. 2021, 172, 112783. [Google Scholar] [CrossRef]
- Ho, J.M.L.; Miller, C.A.; Smith, K.A.; Mattia, J.R.; Bennett, M.R. Improved pyrrolysine biosynthesis through phage assisted non-continuous directed evolution of the complete pathway. Nat. Commun. 2021, 12, 3914. [Google Scholar] [CrossRef]
- Fan, C.; Xiong, H.; Reynolds, N.M.; Söll, D. Rationally evolving tRNAPyl for efficient incorporation of noncanonical amino acids. Nucleic Acids Res. 2015, 43, e156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryson, D.I.; Fan, C.; Guo, L.T.; Miller, C.; Söll, D.; Liu, D.R. Continuous directed evolution of aminoacyl-tRNA synthetases. Nat. Chem. Biol. 2017, 13, 1253–1260. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Miller, C.; Guo, L.T.; Ho, J.M.; Bryson, D.I.; Wang, Y.S.; Liu, D.R.; Söll, D. Crystal structures reveal an elusive functional domain of pyrrolysyl-tRNA synthetase. Nat. Chem. Biol. 2017, 13, 1261. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.T.; Wang, Y.S.; Nakamura, A.; Eiler, D.; Kavran, J.M.; Wong, M.; Kiessling, L.L.; Steitz, T.A.; O’Donoghue, P.; Söll, D. Polyspecific pyrrolysyl-tRNA synthetases from directed evolution. Proc. Natl. Acad. Sci. USA 2014, 111, 16724–16729. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Liu, J.; Ao, H.; Yue, S.; Peng, C. Targeting gut microbiota for precision medicine: Focusing on the efficacy and toxicity of drugs. Theranostics 2020, 10, 11278–11301. [Google Scholar] [CrossRef]
- Koppel, N.; Rekdal, V.M.; Balskus, E.P. Chemical transformation of xenobiotics by the human gut microbiota. Science 2017, 356, 6344. [Google Scholar] [CrossRef]
- Rubini, R.; Mayer, C. Addicting Escherichia coli to New-to-Nature Reactions. ACS Chem. Biol. 2020, 15, 3093–3098. [Google Scholar] [CrossRef]
- Wang, Y.; Xue, P.; Cao, M.; Yu, T.; Lane, S.T.; Zhao, H. Directed Evolution: Methodologies and Applications. Chem. Rev. 2021, 121, 12384–12444. [Google Scholar] [CrossRef]
- Packer, M.S.; Liu, D.R. Methods for the directed evolution of proteins. Nat. Rev. Genet. 2015, 16, 379–394. [Google Scholar] [CrossRef]
- Tang, S.-Y.; Fazelinia, H.; Cirino, P.C. AraC Regulatory Protein Mutants with Altered Effector Specificity. J. Am. Chem. Soc. 2008, 130, 5267–5271. [Google Scholar] [CrossRef]
- Taylor, N.D.; Garruss, A.S.; Moretti, R.; Chan, S.; Arbing, M.A.; Cascio, D.; Rogers, J.K.; Isaacs, F.J.; Kosuri, S.; Baker, D.; et al. Engineering an allosteric transcription factor to respond to new ligands. Nat. Methods 2016, 13, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Guedez, A.; Sherman, M.; Ryu, Y. Dual genetic selection of the theophylline riboswitch with altered aptamer specificity for caffeine. Biochem. Biophys. Res. Commun. 2021, 579, 105–109. [Google Scholar] [CrossRef]
- Snoek, T.; Chaberski, E.K.; Ambri, F.; Kol, S.; Bjørn, S.P.; Pang, B.; Barajas, J.F.; Welner, D.H.; Jensen, M.K.; Keasling, J.D. Evolution-guided engineering of small-molecule biosensors. Nucleic Acids Res. 2020, 48, e3. [Google Scholar] [CrossRef] [PubMed]
- Machado, L.F.M.; Currin, A.; Dixon, N. Directed evolution of the PcaV allosteric transcription factor to generate a biosensor for aromatic aldehydes. J. Biol. Eng. 2019, 13, 91. [Google Scholar] [CrossRef] [Green Version]
- D’Ambrosio, V.; Pramanik, S.; Goroncy, K.; Jakočiūnas, T.; Schönauer, D.; Davari, M.D.; Schwaneberg, U.; Keasling, J.D.; Jensen, M.K. Directed evolution of VanR biosensor specificity in yeast. Biotechnol. Notes 2020, 1, 9–15. [Google Scholar] [CrossRef]
- Hallberg, Z.F.; Su, Y.; Kitto, R.Z.; Hammond, M.C. Engineering and In Vivo Applications of Riboswitches. Annu. Rev. Biochem. 2017, 86, 515–539. [Google Scholar] [CrossRef]
- Page, K.; Shaffer, J.; Lin, S.; Zhang, M.; Liu, J.M. Engineering Riboswitches in Vivo Using Dual Genetic Selection and Fluorescence-Activated Cell Sorting. ACS Synth. Biol. 2018, 7, 2000–2006. [Google Scholar] [CrossRef] [Green Version]
- Muranaka, N.; Sharma, V.; Nomura, Y.; Yokobayashi, Y. An efficient platform for genetic selection and screening of gene switches in Escherichia coli. Nucleic Acids Res. 2009, 37, e39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhlman, B.; Bradley, P. Advances in protein structure prediction and design. Nat. Rev. Mol. Cell Biol. 2019, 20, 681–697. [Google Scholar] [CrossRef]
- Tiwari, M.K.; Singh, R.; Singh, R.K.; Kim, I.-W.; Lee, J.-K. Computational Approaches for Rational Design of Proteins with Novel Functionalities. Comput. Struct. Biotechnol. J. 2012, 2, e201204002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korendovych, I.V. Rational and Semirational Protein Design. Methods Mol. Biol. 2018, 1685, 15–23. [Google Scholar]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, K.W.; Lemmon, G.H.; Deluca, S.L.; Sheehan, J.H.; Meiler, J. Practically useful: What the R osetta protein modeling suite can do for you. Biochemistry 2010, 49, 2987–2998. [Google Scholar] [CrossRef]
- Rouhani, M.; Khodabakhsh, F.; Norouzian, D.; Cohan, R.A.; Valizadeh, V. Molecular dynamics simulation for rational protein engineering: Present and future prospectus. J. Mol. Graph. Model. 2018, 84, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Looger, L.L.; Dwyer, M.A.; Smith, J.J.; Hellinga, H.W. Computational design of receptor and sensor proteins with novel functions. Nature 2003, 423, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Jester, B.W.; Tinberg, C.E.; Rich, M.S.; Baker, D.; Fields, S. Engineered Biosensors from Dimeric Ligand-Binding Domains. ACS Synth. Biol. 2018, 7, 2457–2467. [Google Scholar] [CrossRef]
- Della Corte, D.; van Beek, H.L.; Syberg, F.; Schallmey, M.; Tobola, F.; Cormann, K.U.; Schlicker, C.; Baumann, P.T.; Krumbach, K.; Sokolowsky, S.; et al. Engineering and application of a biosensor with focused ligand specificity. Nat. Commun. 2020, 11, 4851. [Google Scholar] [CrossRef]
- Ike, K.; Arasawa, Y.; Koizumi, S.; Mihashi, S.; Kawai-Noma, S.; Saito, K.; Umeno, D. Evolutionary Design of Choline-Inducible and -Repressible T7-Based Induction Systems. ACS Synth. Biol. 2015, 4, 1352–1360. [Google Scholar] [CrossRef] [PubMed]
- Scholz, O.; Henßler, E.-M.; Bail, J.; Schubert, P.; Bogdanska-Urbaniak, J.; Sopp, S.; Reich, M.; Wisshak, S.; Köstner, M.; Bertram, R.; et al. Activity reversal of Tet repressor caused by single amino acid exchanges. Mol. Microbiol. 2004, 53, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; de Rond, T.; Moore, B.S. Mining genomes to illuminate the specialized chemistry of life. Nat. Rev. Genet. 2021, 22, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Ziemert, N.; Alanjary, M.; Weber, T. The evolution of genome mining in microbes—A review. Nat. Prod. Rep. 2016, 33, 988–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Zhao, H.; Ang, E.-L. A New Biosensor for Stilbenes and a Cannabinoid Enabled by Genome Mining of a Transcriptional Regulator. ACS Synth. Biol. 2020, 9, 698–705. [Google Scholar] [CrossRef]
- Hanko, E.K.R.; Paiva, A.C.; Jonczyk, M.; Abbott, M.; Minton, N.P.; Malys, N. A genome-wide approach for identification and characterisation of metabolite-inducible systems. Nat. Commun. 2020, 11, 1213. [Google Scholar] [CrossRef] [Green Version]
- Grazon, C.; Baer, R.C.; Kuzmanović, U.; Nguyen, T.; Chen, M.; Zamani, M.; Chern, M.; Aquino, P.M.; Zhang, X.; Lecommandoux, S.; et al. A progesterone biosensor derived from microbial screening. Nat. Commun. 2020, 11, 1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidl, S.R.; Ekness, F.; Sofjan, K.; Daeffler, K.N.-M.; Brink, K.; Landry, B.P.; Gerhardt, K.P.; Dyulgyarov, N.; Sheth, R.U.; Tabor, J.J. Rewiring bacterial two-component systems by modular DNA-binding domain swapping. Nat. Chem. Biol. 2019, 15, 690–698. [Google Scholar] [CrossRef]
- Skerker, J.M.; Perchuk, B.S.; Siryaporn, A.; Lubin, E.A.; Ashenberg, O.; Goulian, M.; Laub, M.T. Rewiring the Specificity of Two-Component Signal Transduction Systems. Cell 2008, 133, 1043–1054. [Google Scholar] [CrossRef] [Green Version]
- Tungtur, S.; Schwingen, K.M.; Riepe, J.J.; Weeramange, C.J.; Swint-Kruse, L. Homolog comparisons further reconcile in vitro and in vivo correlations of protein activities by revealing over-looked physiological factors. Protein Sci. 2019, 28, 1806–1818. [Google Scholar] [CrossRef] [PubMed]
- del Solar, G.; Giraldo, R.; Ruiz-Echevarría, M.J.; Espinosa, M.; Díaz-Orejas, R. Replication and Control of Circular Bacterial Plasmids. Microbiol. Mol. Biol. Rev. 1998, 62, 434–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, M.; Sedaghatian, N.; Barajas, J.F.; Wehrs, M.; Bailey, C.; Kaplan, N.; Hillson, N.J.; Mukhopadhyay, A.; Keasling, J.D. Isolation and characterization of novel mutations in the pSC101 origin that increase copy number. Sci. Rep. 2018, 8, 1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin-Chao, S.; Chen, W.-T.; Wong, T.-T. High copy number of the pUC plasmid results from a Rom/Rop-suppressible point mutation in RNA II. Mol. Microbiol. 1992, 6, 3385–3393. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ho, J.; Shis, D.L.; Gupta, C.; Long, J.; Wagner, D.S.; Ott, W.; Josić, K.; Bennett, M.R. Tuning the dynamic range of bacterial promoters regulated by ligand-inducible transcription factors. Nat. Commun. 2018, 9, 64. [Google Scholar] [CrossRef]
- Davis, J.; Rubin, A.J.; Sauer, R.T. Design, construction and characterization of a set of insulated bacterial promoters. Nucleic Acids Res. 2010, 39, 1131–1141. [Google Scholar] [CrossRef] [Green Version]
- Ringquist, S.; Shinedling, S.; Barrick, D.; Green, L.; Binkley, J.; Stormo, G.D.; Gold, L. Translation initiation in Escherichia coli: Sequences within the ribosome-binding site. Mol. Microbiol. 1992, 6, 1219–1229. [Google Scholar] [CrossRef]
- Reis, A.; Salis, H.M. An Automated Model Test System for Systematic Development and Improvement of Gene Expression Models. ACS Synth. Biol. 2020, 9, 3145–3156. [Google Scholar] [CrossRef]
- Kudla, G.; Murray, A.W.; Tollervey, D.; Plotkin, J.B. Coding-Sequence Determinants of Gene Expression in Escherichia coli. Science 2009, 324, 255–258. [Google Scholar] [CrossRef] [Green Version]
- Butzin, N.C.; Mather, W.H. Crosstalk between Diverse Synthetic Protein Degradation Tags inEscherichia coli. ACS Synth. Biol. 2018, 7, 54–62. [Google Scholar] [CrossRef]
- Cameron, D.E.; Collins, J.J. Tunable protein degradation in bacteria. Nat. Biotechnol. 2014, 32, 1276–1281. [Google Scholar] [CrossRef]
- Sekar, K.; Gentile, A.M.; Bostick, J.W.; Tyo, K.E.J. N-Terminal-Based Targeted, Inducible Protein Degradation in Escherichia coli. PLoS ONE 2016, 11, e0149746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, J.K.; Guzman, C.; Taylor, N.D.; Raman, S.; Anderson, K.; Church, G.M. Synthetic biosensors for precise gene control and real-time monitoring of metabolites. Nucleic Acids Res. 2015, 43, 7648–7660. [Google Scholar] [CrossRef]
- Meyer, A.J.; Segall-Shapiro, T.H.; Glassey, E.; Zhang, J.; Voigt, C.A. Escherichia coli “Marionette” strains with 12 highly optimized small-molecule sensors. Nat. Chem. Biol. 2018, 15, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Watstein, D.M.; McNerney, M.P.; Styczynski, M.P. Precise metabolic engineering of carotenoid biosynthesis in Escherichia coli towards a low-cost biosensor. Metab. Eng. 2015, 31, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grilly, C.; Stricker, J.; Pang, W.L.; Bennett, M.R.; Hasty, J. A synthetic gene network for tuning protein degradation in Saccharomyces cerevisiae. Mol. Syst. Biol. 2007, 3, 127. [Google Scholar] [CrossRef] [PubMed]
- Rajasabhai, R.; Go, M.K.; Shan, Y.W. Characterisation of the bacterial biosensor GMG in E-coli BL21 (DE3). Eng. Biol. 2019, 3, 46–56. [Google Scholar] [CrossRef]
- Raman, S.; Rogers, J.K.; Taylor, N.D.; Church, G. Evolution-guided optimization of biosynthetic pathways. Proc. Natl. Acad. Sci. USA 2014, 111, 17803–17808. [Google Scholar] [CrossRef] [Green Version]
- Chalfie, M.; Tu, Y.; Euskirchen, G.; Ward, W.W.; Prasher, D.C. Green fluorescent protein as a marker for gene expression. Science 1994, 263, 802–805. [Google Scholar] [CrossRef] [Green Version]
- Schlechter, R.O.; Jun, H.; Bernach, M.; Oso, S.; Boyd, E.; Muñoz-Lintz, D.A.; Dobson, R.C.J.; Remus, D.M.; Remus-Emsermann, M.N.P. Chromatic Bacteria—A Broad Host-Range Plasmid and Chromosomal Insertion Toolbox for Fluorescent Protein Expression in Bacteria. Front. Microbiol. 2018, 9, 3052. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Park, S.Y.; Lee, S.Y. Production of Rainbow Colorants by Metabolically Engineered Escherichia coli. Adv. Sci. 2021, 8, 2100743. [Google Scholar] [CrossRef]
- Fernandez-Rodriguez, J.; Moser, F.; Song, M.; Voigt, C.A. Engineering RGB color vision into Escherichia coli. Nat. Chem. Biol. 2017, 13, 706–708. [Google Scholar] [CrossRef] [PubMed]
- Fleiss, A.; Sarkisyan, K.S. A brief review of bioluminescent systems. Curr. Genet. 2019, 65, 877–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, C.-W.; Magnuson, T. A Novel Selection Marker for Efficient DNA Cloning and Recombineering in E. coli. PLoS ONE 2013, 8, e57075. [Google Scholar] [CrossRef]
- Peubez, I.; Chaudet, N.; Mignon, C.; Hild, G.; Husson, S.; Courtois, V.; De Luca, K.; Speck, D.; Sodoyer, R. Antibiotic-free selection in E. coli: New considerations for optimal design and improved production. Microb. Cell Factories 2010, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Bird, L.J.; Kundu, B.B.; Tschirhart, T.; Corts, A.D.; Su, L.; Gralnick, J.A.; Ajo-Franklin, C.M.; Glaven, S.M. Engineering Wired Life: Synthetic Biology for Electroactive Bacteria. ACS Synth. Biol. 2021, 10, 2808–2823. [Google Scholar] [CrossRef] [PubMed]
- Fulk, E.M.; Huh, D.; Atkinson, J.T.; Lie, M.; Masiello, C.; Silberg, J.J. A Split Methyl Halide Transferase AND Gate That Reports by Synthesizing an Indicator Gas. ACS Synth. Biol. 2020, 9, 3104–3113. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Liu, D.R. Rewritable multi-event analog recording in bacterial and mammalian cells. Science 2018, 360, eaap8992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzadfard, F.; Lu, T.K. Genomically encoded analog memory with precise in vivo DNA writing in living cell populations. Science 2014, 346, 1256272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smale, S.T. β-Galactosidase Assay; Cold Spring Harbor Protocols: 2010; CSHL Press: Cold Spring Harbor, NY, USA, 2010; pdb.prot5423. [Google Scholar]
- Smale, S.T. Luciferase Assay; Cold Spring Harbor Protocols: 2010; CSHL Press: Cold Spring Harbor, NY, USA, 2010; pdb.prot5421. [Google Scholar]
- Lopreside, A.; Wan, X.; Michelini, E.; Roda, A.; Wang, B. Comprehensive Profiling of Diverse Genetic Reporters with Application to Whole-Cell and Cell-Free Biosensors. Anal. Chem. 2019, 91, 15284–15292. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.E.; Kalimuthu, S.; Ahn, B.-C. In Vivo Cell Tracking with Bioluminescence Imaging. Nucl. Med. Mol. Imaging 2015, 49, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Shcherbakova, D.M.; Stepanenko, O.V.; Turoverov, K.K.; Verkhusha, V.V. Near-Infrared Fluorescent Proteins: Multiplexing and Optogenetics across Scales. Trends Biotechnol. 2018, 36, 1230–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, N.; Wu, Y.; Chen, W.; Huang, Q.; Cai, P. Whole-Cell Microbial Bioreporter for Soil Contaminants Detection. Front. Bioeng. Biotechnol. 2021, 9, 79. [Google Scholar] [CrossRef]
- Streett, H.E.; Kalis, K.M.; Papoutsakis, E.T. A Strongly Fluorescing Anaerobic Reporter and Protein-Tagging System for Clostridium Organisms Based on the Fluorescence-Activating and Absorption-Shifting Tag Protein (FAST). Appl. Environ. Microbiol. 2019, 85, e00622-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wójcik, M.; Telzerow, A.; Quax, W.J.; Boersma, Y.L. High-Throughput Screening in Protein Engineering: Recent Advances and Future Perspectives. Int. J. Mol. Sci. 2015, 16, 24918–24945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsien, R.Y. The green fluorescent protein. Annu. Rev. Biochem. 1998, 67, 509–544. [Google Scholar] [CrossRef]
- Pédelacq, J.-D.; Cabantous, S.; Tran, T.; Terwilliger, T.; Waldo, G.S. Engineering and characterization of a superfolder green fluorescent protein. Nat. Biotechnol. 2005, 24, 79–88. [Google Scholar] [CrossRef]
- Scott, D.J.; Gunn, N.; Yong, K.J.; Wimmer, V.; Veldhuis, N.; Challis, L.M.; Haidar, M.; Petrou, S.; Bathgate, R.A.D.; Griffin, M.D.W. A Novel Ultra-Stable, Monomeric Green Fluorescent Protein for Direct Volumetric Imaging of Whole Organs Using CLARITY. Sci. Rep. 2018, 8, 667. [Google Scholar] [CrossRef]
- Valbuena, F.M.; Fitzgerald, I.; Strack, R.L.; Andruska, N.; Smith, L.; Glick, B.S. A Photostable Monomeric Superfolder GFP; Traffic: Copenhagen, Denmark, 2020; Volume 21, p. 534. [Google Scholar]
- Kaku, T.; Sugiura, K.; Entani, T.; Osabe, K.; Nagai, T. Enhanced brightness of bacterial luciferase by bioluminescence resonance energy transfer. Sci. Rep. 2021, 11, 14994. [Google Scholar] [CrossRef]
- Gregor, C.; Gwosch, K.; Sahl, S.J.; Hell, S.W. Strongly enhanced bacterial bioluminescence with the ilux operon for single-cell imaging. Proc. Natl. Acad. Sci. USA 2018, 115, 962–967. [Google Scholar] [CrossRef] [Green Version]
- Juers, D.H.; Matthews, B.W.; Huber, R.E. LacZ β-galactosidase: Structure and function of an enzyme of historical and molecular biological importance. Protein Sci. A Publ. Protein Soc. 2012, 21, 1792. [Google Scholar] [CrossRef] [Green Version]
- Shoemaker, G.K.; Juers, D.H.; Coombs, J.M.L.; Matthews, B.W.; Craig, D.B. Crystallization of β-Galactosidase Does Not Reduce the Range of Activity of Individual Molecules. Biochemistry 2003, 42, 1707–1710. [Google Scholar] [CrossRef] [PubMed]
- Kuchimaru, T.; Iwano, S.; Kiyama, M.; Mitsumata, S.; Kadonosono, T.; Niwa, H.; Maki, S.; Kizaka-Kondoh, S. A luciferin analogue generating near-infrared bioluminescence achieves highly sensitive deep-tissue imaging. Nat. Commun. 2016, 7, 11856. [Google Scholar] [CrossRef] [Green Version]
- Johnston, C.W.; Badran, A.H.; Collins, J.J. Continuous bioactivity-dependent evolution of an antibiotic biosynthetic pathway. Nat. Commun. 2020, 11, 4202. [Google Scholar] [CrossRef]
- Miller, C.A.; Ho, J.M.; Parks, S.E.; Bennett, M.R. Macrolide Biosensor Optimization through Cellular Substrate Sequestration. ACS Synth. Biol. 2021, 10, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Barahona, M.; Buck, M. Engineering modular and tunable genetic amplifiers for scaling transcriptional signals in cascaded gene networks. Nucleic Acids Res. 2014, 42, 9484–9492. [Google Scholar] [CrossRef]
- Garenne, D.; Thompson, S.; Brisson, A.; Khakimzhan, A.; Noireaux, V. The all-E. coliTXTL toolbox 3.0: New capabilities of a cell-free synthetic biology platform. Synth. Biol. 2021, 6, ysab017. [Google Scholar] [CrossRef] [PubMed]
- Juhas, M.; Ajioka, J.W. T7 RNA polymerase-driven inducible cell lysis for DNA transfer from Escherichia coli to Bacillus subtilis. Microb. Biotechnol. 2017, 10, 1797. [Google Scholar] [CrossRef]
- Wan, X.; Volpetti, F.; Petrova, E.; French, C.; Maerkl, S.J.; Wang, B. Cascaded amplifying circuits enable ultrasensitive cellular sensors for toxic metals. Nat. Chem. Biol. 2019, 15, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Hu, J.H.; Liu, D.R. Aptazyme-embedded guide RNAs enable ligand-responsive genome editing and transcriptional activation. Nat. Commun. 2017, 8, 15939. [Google Scholar] [CrossRef] [Green Version]
- Wieland, M.; Hartig, J.S. Improved Aptazyme Design and In Vivo Screening Enable Riboswitching in Bacteria. Angew. Chem. Int. Ed. 2008, 47, 2604–2607. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.M.L.; Miller, C.A.; Parks, S.E.; Mattia, J.R.; Bennett, M.R. A suppressor tRNA-mediated feedforward loop eliminates leaky gene expression in bacteria. Nucleic Acids Res. 2021, 49, e25. [Google Scholar] [CrossRef] [PubMed]
- Greco, F.V.; Pandi, A.; Erb, T.J.; Grierson, C.S.; Gorochowski, T.E. Harnessing the central dogma for stringent multi-level control of gene expression. Nat. Commun. 2021, 12, 1738. [Google Scholar] [CrossRef]
- Fernandez-Rodriguez, J.; Voigt, C.A. Post-translational control of genetic circuits using Potyvirus proteases. Nucleic Acids Res. 2016, 44, 6493–6502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tas, H.; Grozinger, L.; Stoof, R.; de Lorenzo, V.; Goñi-Moreno, Á. Contextual dependencies expand the re-usability of genetic inverters. Nat. Commun. 2021, 12, 355. [Google Scholar] [CrossRef] [PubMed]
- Lou, C.; Stanton, B.C.; Chen, Y.-J.; Munsky, B.; Voigt, C.A. Ribozyme-based insulator parts buffer synthetic circuits from genetic context. Nat. Biotechnol. 2012, 30, 1137–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shis, D.L.; Bennett, M.R. Library of synthetic transcriptional AND gates built with split T7 RNA polymerase mutants. Proc. Natl. Acad. Sci. USA 2013, 110, 5028–5033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Kitney, R.I.; Joly, N.; Buck, M. Engineering modular and orthogonal genetic logic gates for robust digital-like synthetic biology. Nat. Commun. 2011, 2, 508. [Google Scholar] [CrossRef] [Green Version]
- Brophy, J.A.; Voigt, C.A. Principles of genetic circuit design. Nat. Methods 2014, 11, 508–520. [Google Scholar] [CrossRef] [Green Version]
- Wan, X.; Saltepe, B.; Yu, L.; Wang, B. Programming living sensors for environment, health and biomanufacturing. Microb. Biotechnol. 2021, 14, 2334–2342. [Google Scholar] [CrossRef]
- Morrison, M.S.; Podracky, C.J.; Liu, D.R. The developing toolkit of continuous directed evolution. Nat. Chem. Biol. 2020, 16, 610–619. [Google Scholar] [CrossRef]
- Pan, X.; Kortemme, T. Recent advances in de novo protein design: Principles, methods, and applications. J. Biol. Chem. 2021, 296, 100558. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Iqbal, H.M. Microbial-derived biosensors for monitoring environmental contaminants: Recent advances and future outlook. Process. Saf. Environ. Prot. 2019, 124, 8–17. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, C.A.; Ho, J.M.L.; Bennett, M.R. Strategies for Improving Small-Molecule Biosensors in Bacteria. Biosensors 2022, 12, 64. https://doi.org/10.3390/bios12020064
Miller CA, Ho JML, Bennett MR. Strategies for Improving Small-Molecule Biosensors in Bacteria. Biosensors. 2022; 12(2):64. https://doi.org/10.3390/bios12020064
Chicago/Turabian StyleMiller, Corwin A., Joanne M. L. Ho, and Matthew R. Bennett. 2022. "Strategies for Improving Small-Molecule Biosensors in Bacteria" Biosensors 12, no. 2: 64. https://doi.org/10.3390/bios12020064
APA StyleMiller, C. A., Ho, J. M. L., & Bennett, M. R. (2022). Strategies for Improving Small-Molecule Biosensors in Bacteria. Biosensors, 12(2), 64. https://doi.org/10.3390/bios12020064