A Recombinase-Based Genetic Circuit for Heavy Metal Monitoring


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Growth Media
2.2. Sensor Plasmid Assembly
2.3. Fluorescence Measurement and Data Analysis
2.4. Statistical Analysis
3. Results and Discussion
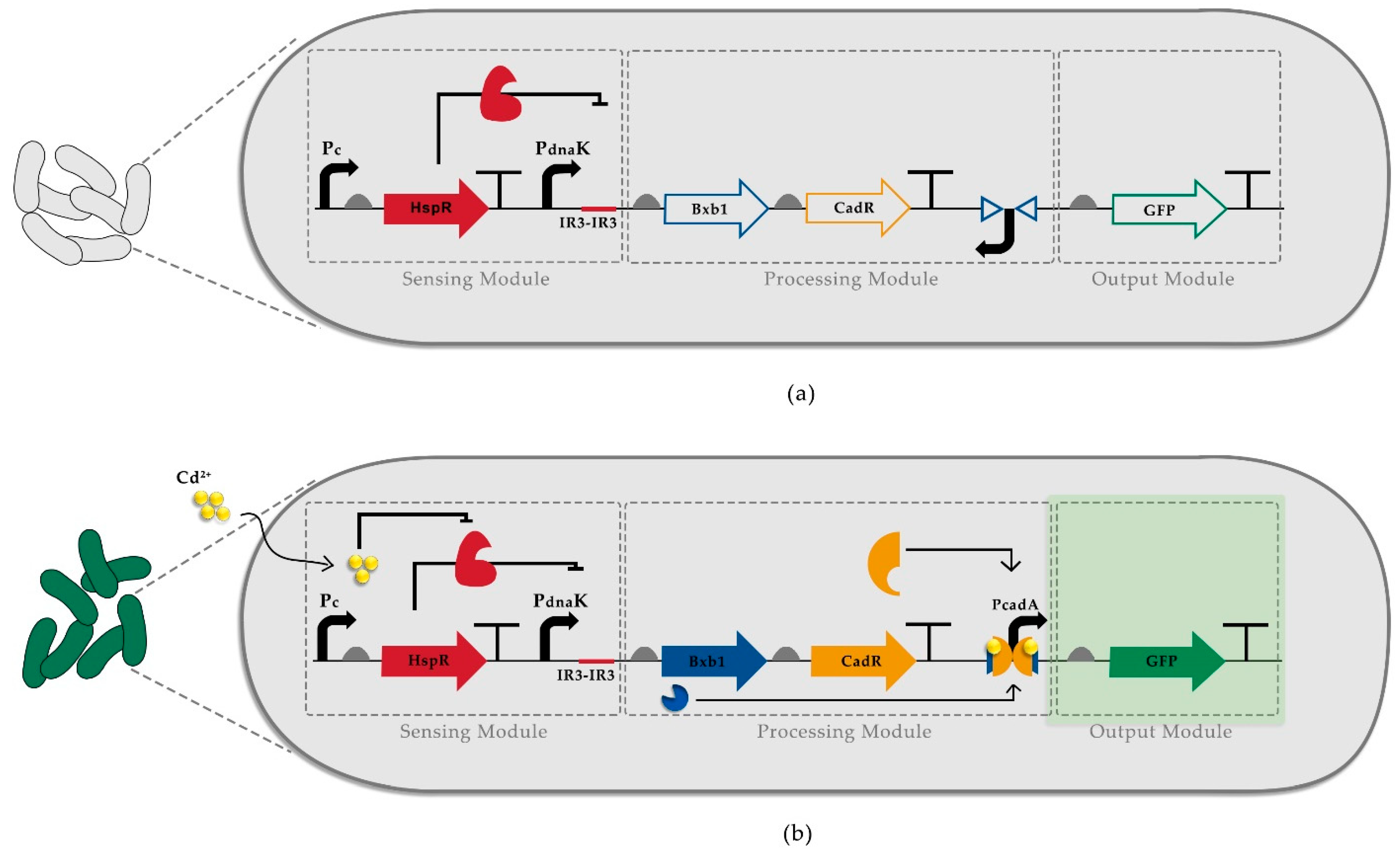
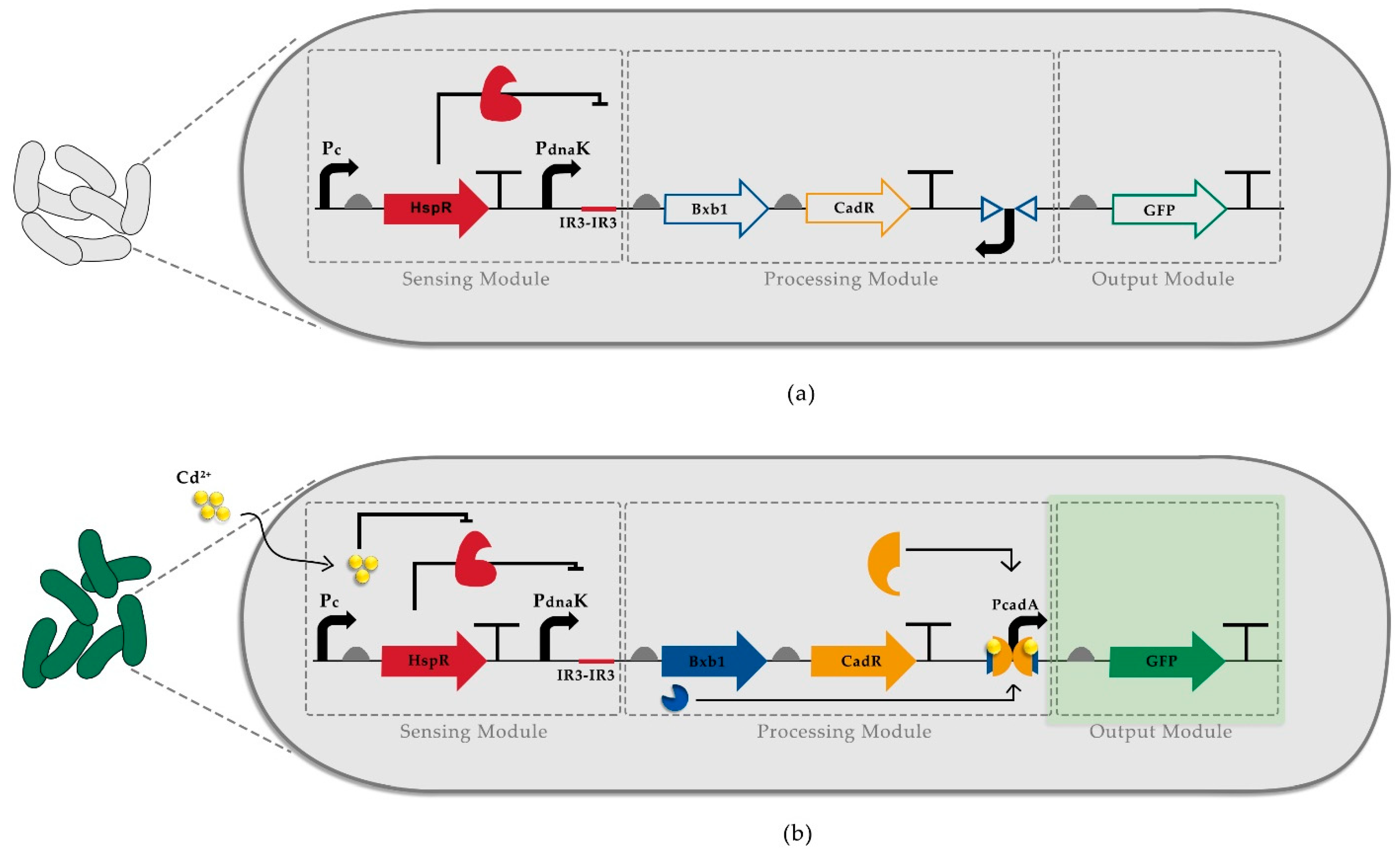
3.1. Construction of Recombinase-Based Cadmium Detecting Sensor
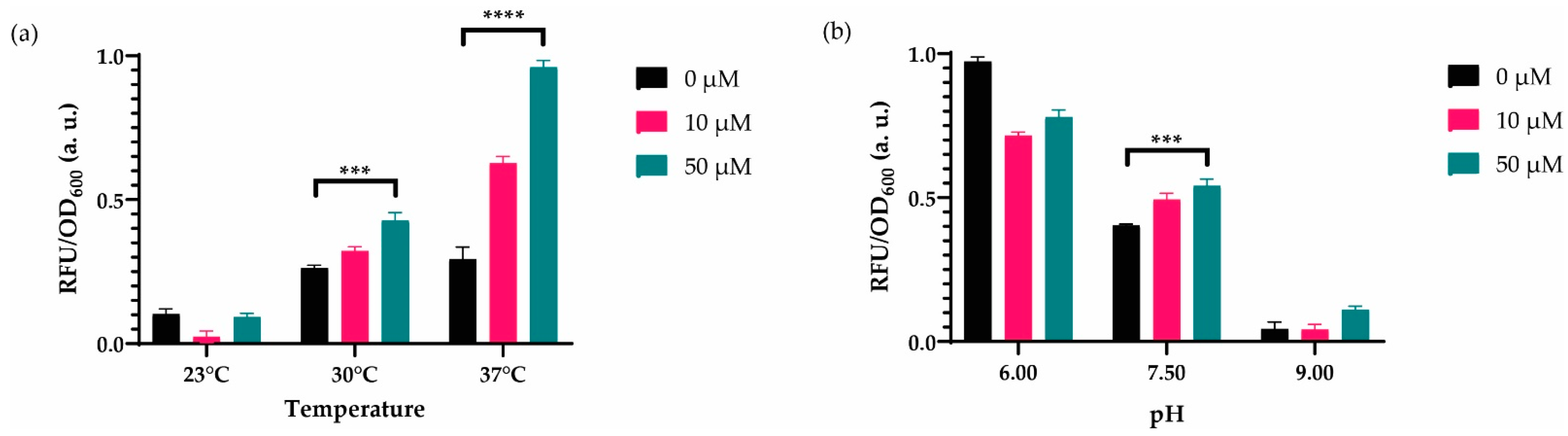
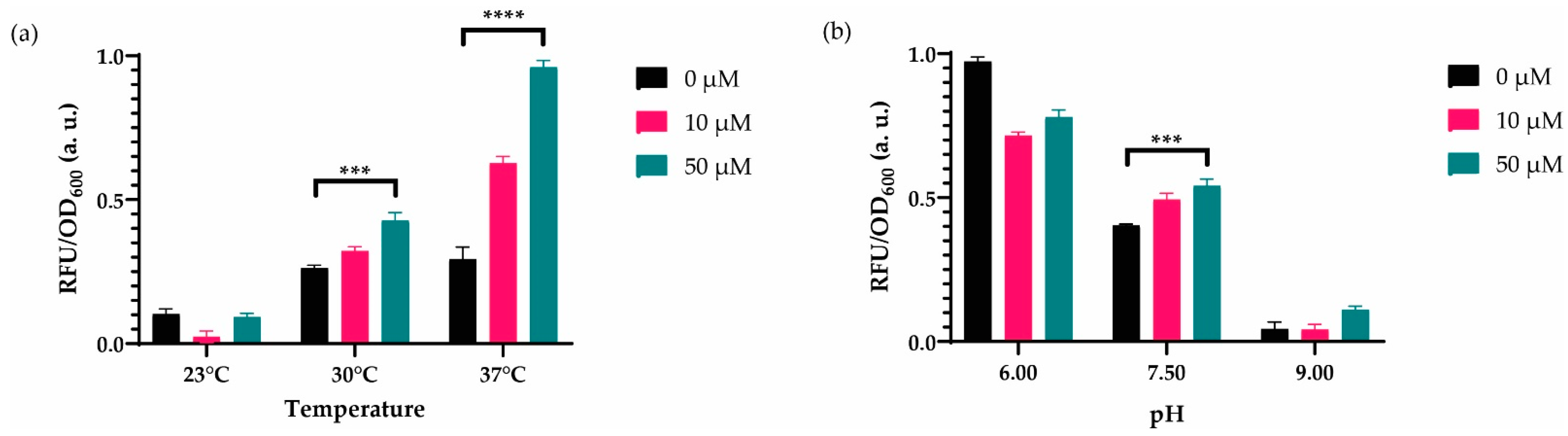
3.2. Characterization of the System to Optimal Working Conditions
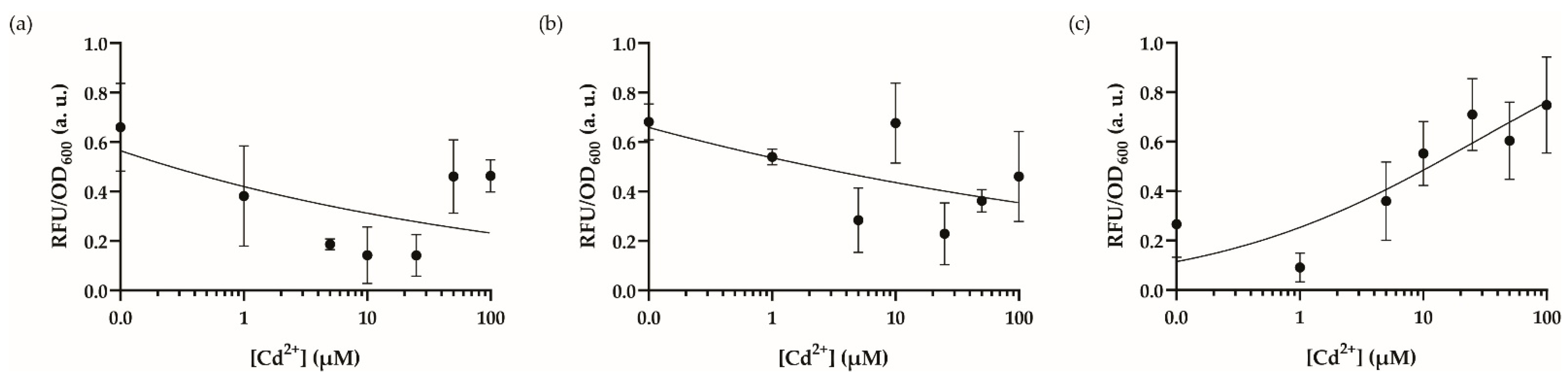
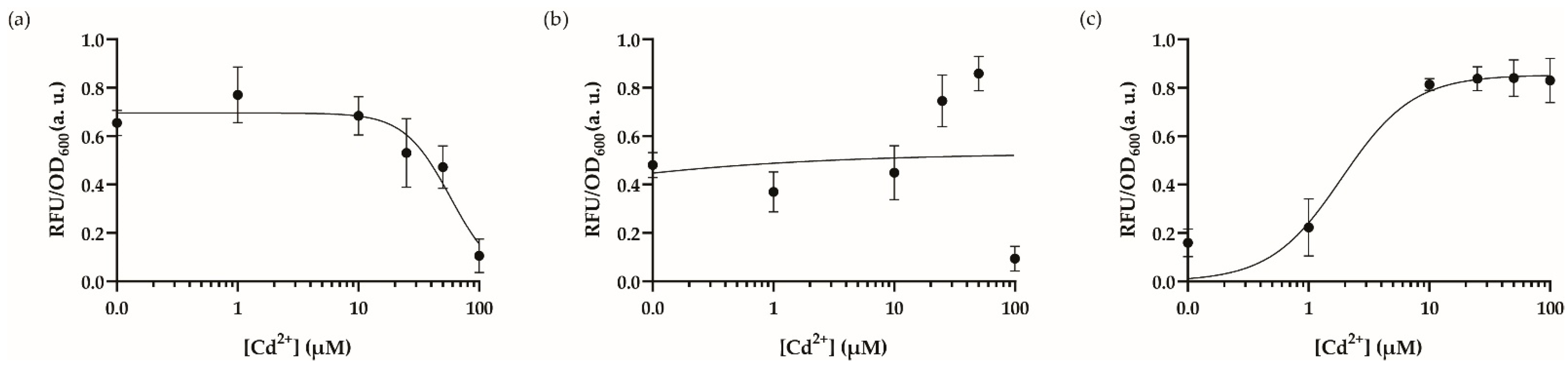
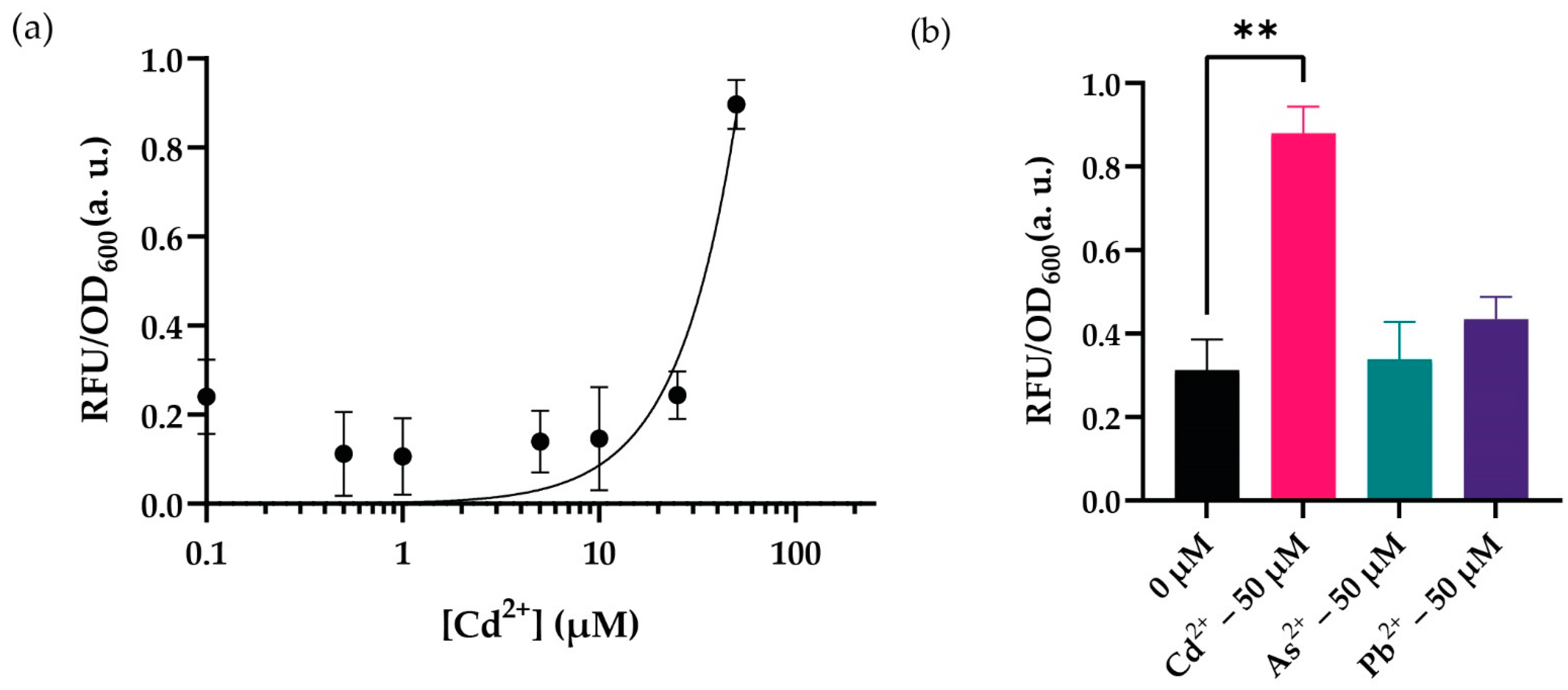
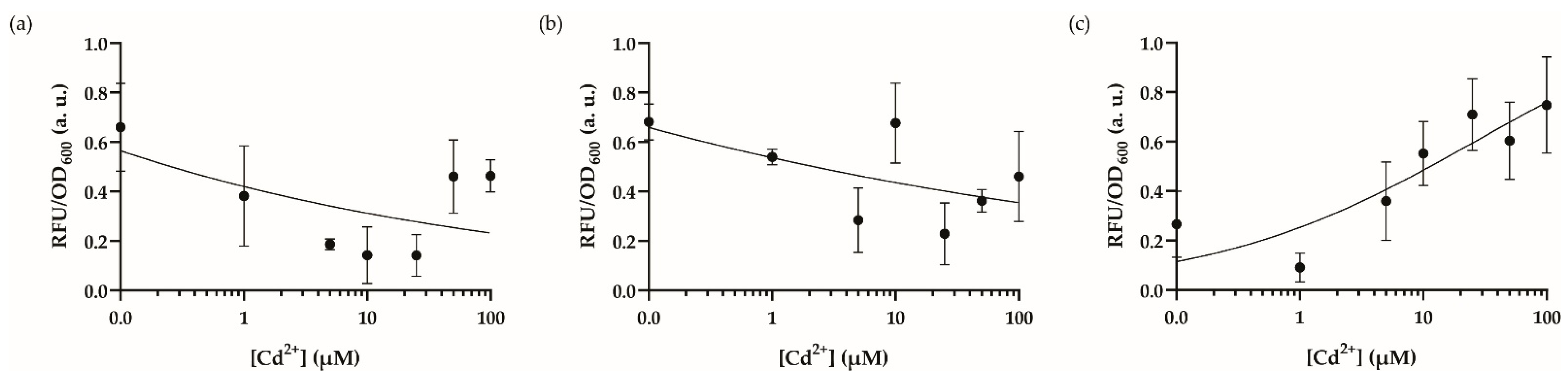
3.3. Optimal Dynamic Range and Cross-Reactivity Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| WCBs | Whole-cell biosensors |
| HSR | Heat shock response |
| ICP-MS | Inductively coupled plasma mass spectrometry |
| Cd(II), Cd2+ | Cadmium ion |
| As(II), As2+ | Arsenic ion |
| Pb(II), Pb2+ | Lead ion |
| Hg(II) | Mercury ion |
| Zn(II) | Zinc ion |
| LB | Lysogeny Broth |
| OD600 | Optical density (600 nm) |
| SEM | Standard error mean |
| RFU | Relative Fluorescence Unit |
| GFP | Green Fluorescent Protein |
| Pc | Constitutively active promoter |
| MOPS | Sodium;3-(N-morpholino)propanesulfonic acid |
| HMM | Heavy Metal MOPS media |
| CadR | Cadmium-binding transcription factor of CadA |
| PcadA | Promoter of CadA |
| HspR | Heat shock protein transcriptional repressor |
| PdnaK | Promoter of dnaK, cognitive promoter of HspR |
| IR3 | HspR binding motif |
| Bxb1 | Bxb1 serine recombinase |
References
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on hu-mans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef]
- Baran, A.; Tarnawski, M. Assessment of heavy metals mobility and toxicity in contaminated sediments by sequential extraction and a battery of bioassays. Ecotoxicology 2015, 24, 1279–1293. [Google Scholar] [CrossRef] [Green Version]
- Egger, A.E.; Grabmann, G.; Gollmann-Tepeköylü, C.; Pechriggl, E.J.; Artner, C.; Türkcan, A.; Hartinger, C.G.; Fritsch, H.; Keppler, B.K.; Brenner, E.; et al. Chemical imaging and assessment of cadmium distribution in the human body. Metallomics 2019, 11, 2010–2019. [Google Scholar] [CrossRef] [Green Version]
- Olmedo, P.; Pla, A.; Hernández, A.F.; López-Guarnido, O.; Rodrigo, L.; Gil, F. Validation of a method to quantify chromium, cadmium, manganese, nickel and lead in human whole blood, urine, saliva and hair samples by electrothermal atomic absorption spectrometry. Anal. Chim. Acta 2010, 659, 60–67. [Google Scholar] [CrossRef]
- Mehana, E.S.E.; Khafaga, A.F.; Elblehi, S.S.; Abd El-Hack, M.E.; Naiel, M.A.E.; Bin-Jumah, M.; Othman, S.I.; Allam, A.A. Biomonitoring of Heavy Metal Pollution Using Acanthocephalans Parasite in Ecosystem: An Updated Overview. Animals 2020, 10, 811. [Google Scholar] [CrossRef]
- Tauriainen, S.; Karp, M.; Chang, W.; Virta, M. Luminescent bacterial sensor for cadmium and lead. Biosens. Bioelectron. 1998, 13, 931–938. [Google Scholar] [CrossRef]
- Futra, D.; Heng, L.Y.; Surif, S.; Ahmad, A.; Ling, T.L. Microencapsulated Aliivibrio fischeri in Alginate Microspheres for Monitoring Heavy Metal Toxicity in Environmental Waters. Sensors 2014, 14, 23248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacKen, A.; Giltrap, M.; Ryall, K.; Foley, B.; McGovern, E.; McHugh, B.; Davoren, M. A test battery approach to the ecotoxicological evaluation of cadmium and copper employing a battery of marine bioassays. Ecotoxicology 2009, 18, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Fulladosa, E.; Murat, J.C.; Villaescusa, I. Study on the toxicity of binary equitoxic mixtures of metals using the lumi-nescent bacteria Vibrio fischeri as a biological target. Chemosphere 2005, 58, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Jeong, H.; Lee, S.J. Synthetic biology for microbial heavy metal biosensors. Anal. Bioanal. Chem. 2017, 410, 1191–1203. [Google Scholar] [CrossRef]
- Wang, B.; Buck, M. Customizing cell signaling using engineered genetic logic circuits. Trends Microbiol. 2012, 20, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Barahona, M.; Buck, M. A modular cell-based biosensor using engineered genetic logic circuits to detect and integrate multiple environmental signals. Biosens. Bioelectron. 2013, 40, 368–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saltepe, B.; Kehribar, E.Ş.; Su Yirmibeşoǧlu, S.S.; Şafak Şeker, U.Ö. Cellular Biosensors with Engineered Genetic Cir-cuits. ACS Sensors 2017, 3, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Cerminati, S.; Soncini, F.C.; Checa, S.K. A sensitive whole-cell biosensor for the simultaneous detection of a broad-spectrum of toxic heavy metal ions. Chem. Commun. 2015, 51, 5917–5920. [Google Scholar] [CrossRef]
- Jung, J.K.; Alam, K.K.; Verosloff, M.S.; Capdevila, D.A.; Desmau, M.; Clauer, P.R.; Lee, J.W.; Nguyen, P.Q.; Pastén, P.A.; Matiasek, S.J.; et al. Cell-free biosensors for rapid detection of water contaminants. Nat. Biotechnol. 2020, 38, 1451–1459. [Google Scholar] [CrossRef]
- Wan, X.; Volpetti, F.; Petrova, E.; French, C.; Maerkl, S.J.; Wang, B. Cascaded amplifying circuits enable ultrasensitive cellular sensors for toxic metals. Nat. Chem. Biol. 2019, 15, 540–548. [Google Scholar] [CrossRef]
- Rodrigues, J.L.; Rodrigues, L.R. Potential Applications of the Escherichia coli Heat Shock Response in Synthetic Biol-ogy. Trends Biotechnol. 2018, 36, 186–198. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, S.J.; Burmølle, M.; Hansen, L.H. Making bio-sense of toxicity: New developments in whole-cell biosensors. Curr. Opin. Biotechnol. 2016, 17, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Mahr, R.; Frunzke, J. Transcription factor-based biosensors in biotechnology: Current state and future prospects. Appl. Microbiol. Biotechnol. 2016, 100, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Checa, S.K.; Zurbriggen, M.D.; Soncini, F.C. Bacterial signaling systems as platforms for rational design of new gen-erations of biosensors. Curr. Opin. Biotechnol. 2012, 23, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Siuti, P.; Yazbek, J.; Lu, T.K. Engineering genetic circuits that compute and remember. Nat. Protoc. 2014, 9, 1292–1300. [Google Scholar] [CrossRef] [PubMed]
- Roquet, N.; Soleimany, A.P.; Ferris, A.C.; Aaronson, S.; Lu, T.K. Synthetic recombinase-based State machines in liv-ing cells. Science 2016, 353, aad8559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, X.; Ho, T.Y.H.; Wang, B. Engineering Prokaryote Synthetic Biology Biosensors. Handb. Cell Biosens. 2022, 283–318. [Google Scholar] [CrossRef]
- Kotula, J.W.; Kerns, S.J.; Shaket, L.A.; Siraj, L.; Collins, J.J.; Way, J.C.; Silver, P.A. Programmable bacteria detect and record an environmental signal in the mammalian gut. Proc. Natl. Acad. Sci. USA 2014, 111, 4838–4843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saltepe, B.; Bozkurt, E.U.; Güngen, M.A.; Çiçek, A.E.; Şeker, U.Ö.Ş. Genetic circuits combined with machine learning provides fast responding living sensors. Biosens. Bioelectron. 2021, 178, 113028. [Google Scholar] [CrossRef]
- Saltepe, B.; Bozkurt, E.U.; Haclosmanoǧlu, N.; Şeker, U.Ö.Ş. Genetic Circuits to Detect Nanomaterial Triggered Tox-icity through Engineered Heat Shock Response Mechanism. ACS Synth. Biol. 2019, 8, 2404–2417. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.W.; Glickmann, E.; Cooksey, D.A. Chromosomal locus for cadmium resistance in Pseudomonas putida con-sisting of a cadmium-transporting ATPase and a MerR family response regulator. Appl. Environ. Microbiol. 2001, 67, 1437–1444. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, P.; Pannunzio, N.R.; Hatfull, G.F.; Gottesman, M. Synapsis in phage Bxb1 integration: Selection mechanism for the correct pair of recombination sites. J. Mol. Biol. 2005, 349, 331–348. [Google Scholar] [CrossRef]
- Neidhardt, F.C.; Bloch, P.L.; Smith1, D.F. Culture Medium for Enterobacteria. J. Bacteriol. 1974, 119, 736–747. [Google Scholar] [CrossRef] [Green Version]
- LaRossa, R.A.; Smulski, D.R.; van Dyk, T.K. Interaction of lead nitrate and cadmium chloride with Escherichia coli K-12 and Salmonella typhimurium global regulatory mutants. J. Ind. Microbiol. 1995, 14, 252–258. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Jäättelä, M. Heat shock proteins as cellular lifeguards. Ann. Med. 1999, 31, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Berepiki, A.; Kent, R.; MacHado, L.F.M.; Dixon, N. Development of High-Performance Whole Cell Biosensors Aided by Statistical Modeling. ACS Synth. Biol. 2020, 9, 576–589. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, A.; Hirakawa, H.; Hirata, T.; Nishino, K.; Yamaguchi, A. Growth Phase-Dependent Expression of Drug Exporters in Escherichia coli and Its Contribution to Drug Tolerance. J. Bacteriol. 2006, 188, 5693. [Google Scholar] [CrossRef] [Green Version]
- Tao, H.C.; Peng, Z.W.; Li, P.S.; Yu, T.A.; Su, J. Optimizing cadmium and mercury specificity of CadR-based E. coli biosensors by redesign of CadR. Biotechnol. Lett. 2013, 35, 1253–1258. [Google Scholar] [CrossRef]
- Smith, M.C.A.; Till, R.; Smith, M.C.M. Switching the polarity of a bacteriophage integration system. Mol. Microbiol. 2004, 51, 1719–1728. [Google Scholar] [CrossRef]
- Wang, X.; Tang, B.; Ye, Y.; Mao, Y.; Lei, X.; Zhao, G.; Ding, X. Bxb1 integrase serves as a highly efficient DNA recombinase in rapid metabolite pathway assembly. Acta Biochim. Et Biophys. Sin. 2017, 49, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Brown, N.L.; Stoyanov, J.V.; Kidd, S.P.; Hobman, J.L. The MerR family of transcriptional regulators. FEMS Microbiol. Rev. 2003, 27, 145–163. [Google Scholar] [CrossRef] [Green Version]
- Bereza-Malcolm, L.; Aracic, S.; Kannan, R.; Mann, G.; Franks, A.E. Functional characterization of Gram-negative bacteria from different genera as multiplex cadmium biosensors. Biosens. Bioelectron. 2017, 94, 380–387. [Google Scholar] [CrossRef]
- Hakkila, K.M.; Nikander, P.A.; Junttila, S.M.; Lamminmäki, U.J.; Virta, M.P. Cd-Specific Mutants of Mercury-Sensing Regulatory Protein MerR, Generated by Directed Evolution. Appl. Environ. Microbiol. 2011, 77, 6215. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Hu, S.; Jia, X. Highly Sensitive Whole-Cell Biosensor for Cadmium Detection Based on a Negative Feed-back Circuit. Front. Bioeng. Biotechnol. 2021, 9, 1251. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Barahona, M.; Buck, M. Engineering modular and tunable genetic amplifiers for scaling transcriptional signals in cascaded gene networks. Nucleic Acids Res. 2014, 42, 9484–9492. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akboğa, D.; Saltepe, B.; Bozkurt, E.U.; Şeker, U.Ö.Ş. A Recombinase-Based Genetic Circuit for Heavy Metal Monitoring. Biosensors 2022, 12, 122. https://doi.org/10.3390/bios12020122
Akboğa D, Saltepe B, Bozkurt EU, Şeker UÖŞ. A Recombinase-Based Genetic Circuit for Heavy Metal Monitoring. Biosensors. 2022; 12(2):122. https://doi.org/10.3390/bios12020122
Chicago/Turabian StyleAkboğa, Doğuş, Behide Saltepe, Eray Ulaş Bozkurt, and Urartu Özgür Şafak Şeker. 2022. "A Recombinase-Based Genetic Circuit for Heavy Metal Monitoring" Biosensors 12, no. 2: 122. https://doi.org/10.3390/bios12020122
APA StyleAkboğa, D., Saltepe, B., Bozkurt, E. U., & Şeker, U. Ö. Ş. (2022). A Recombinase-Based Genetic Circuit for Heavy Metal Monitoring. Biosensors, 12(2), 122. https://doi.org/10.3390/bios12020122