In Vitro Analysis of the Effects of ITER-Like Tungsten Nanoparticles: Cytotoxicity and Epigenotoxicity in BEAS-2B Cells

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Particles Synthesis and Suspensions Preparation
2.3. Cellular Model
2.4. Cytotoxicity Assessment
([(number binucleated cells) + (2 × number multinucleated cells)]/total control cells) × 100.
2.5. Genotoxicity Studies: Micronucleus and Alkaline Comet Assay
2.6. Oxidative Stress Evaluation
2.7. Epigenotoxic Effects of W-NPs
2.8. Statistical Analysis
3. Results
3.1. ITER-Like Plasma and Laser W-NPs Physico-Chemical Characterization
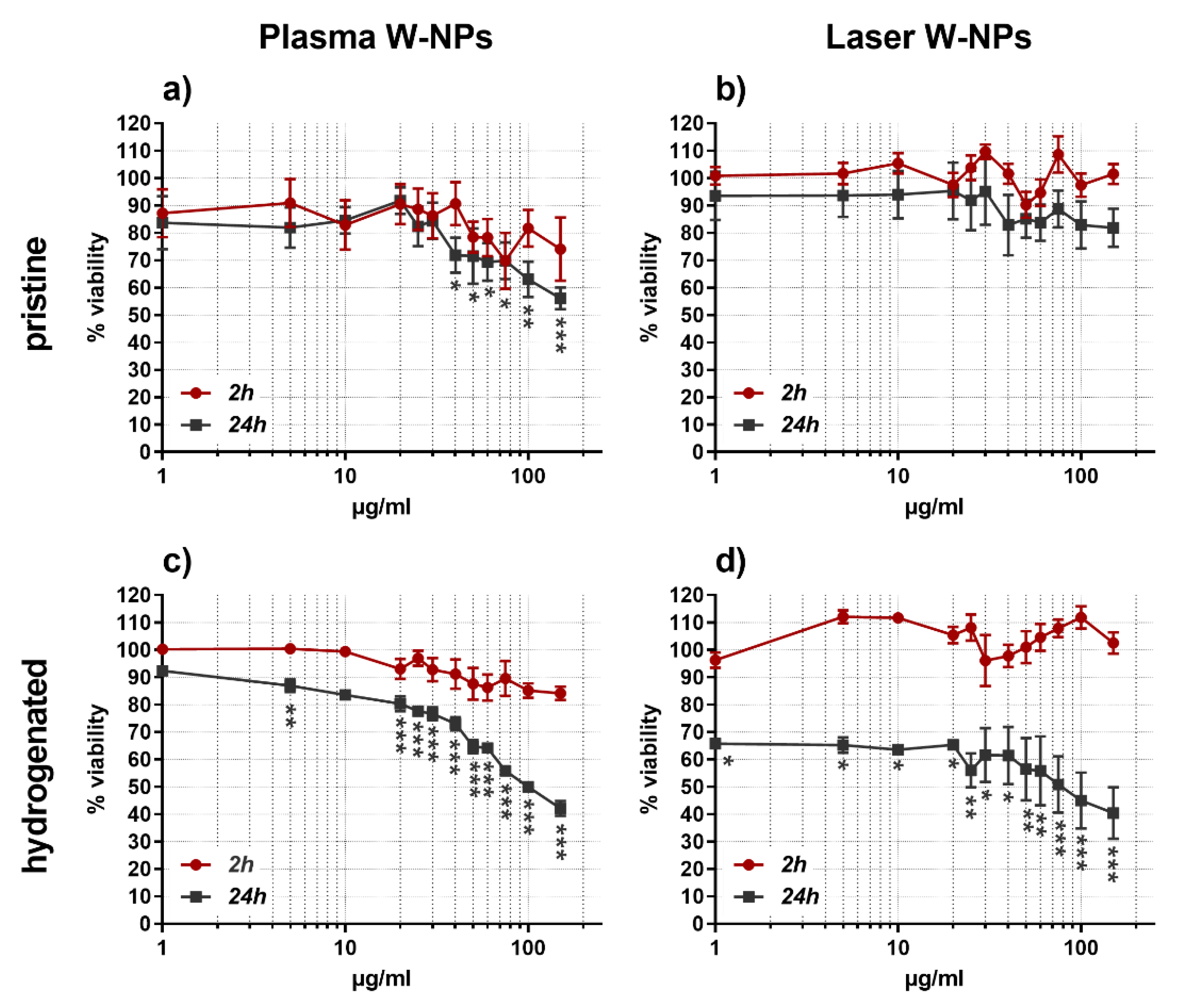
3.2. Cytotoxic Effects
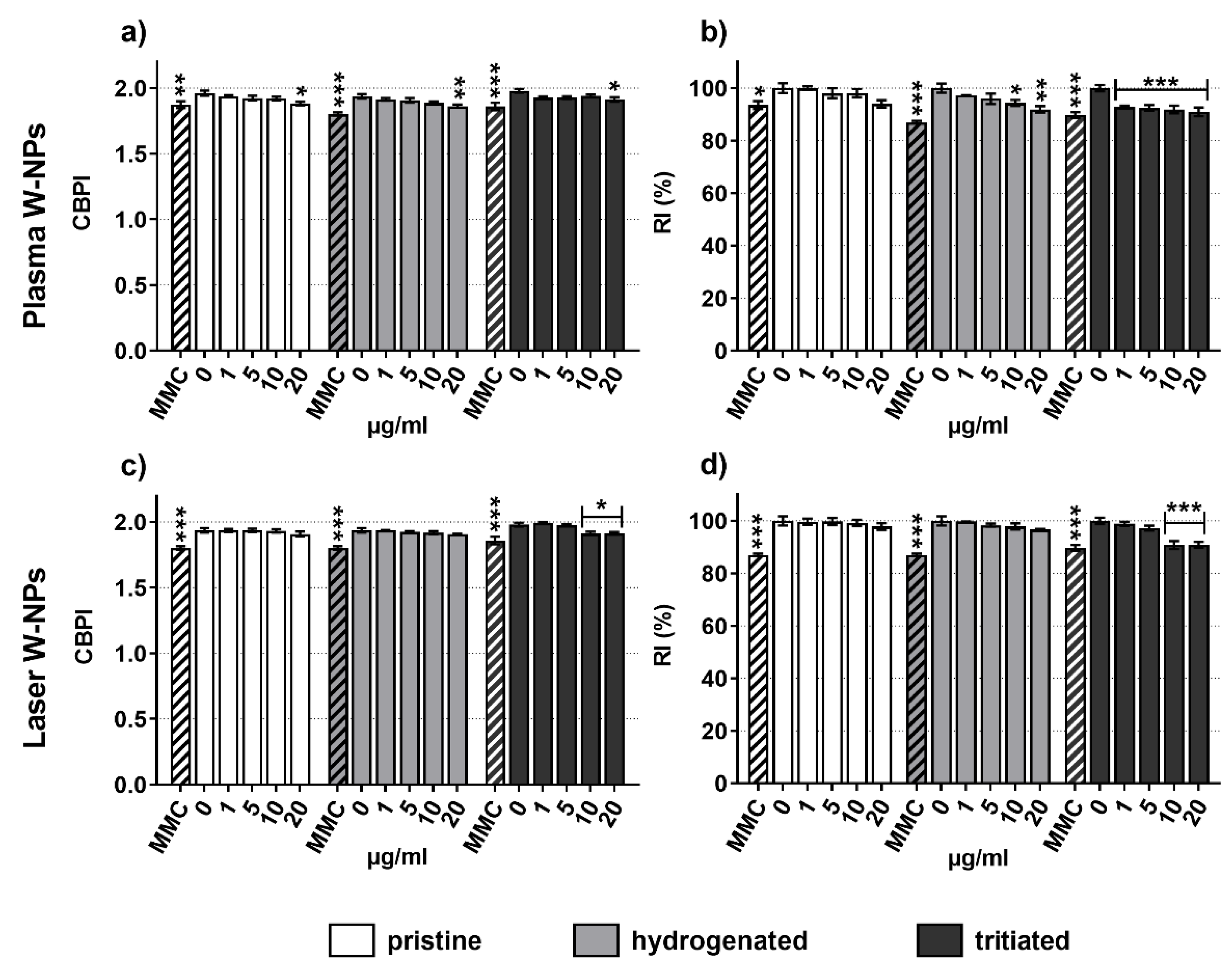
3.3. Effects on Cytostasis and Cellular Replication
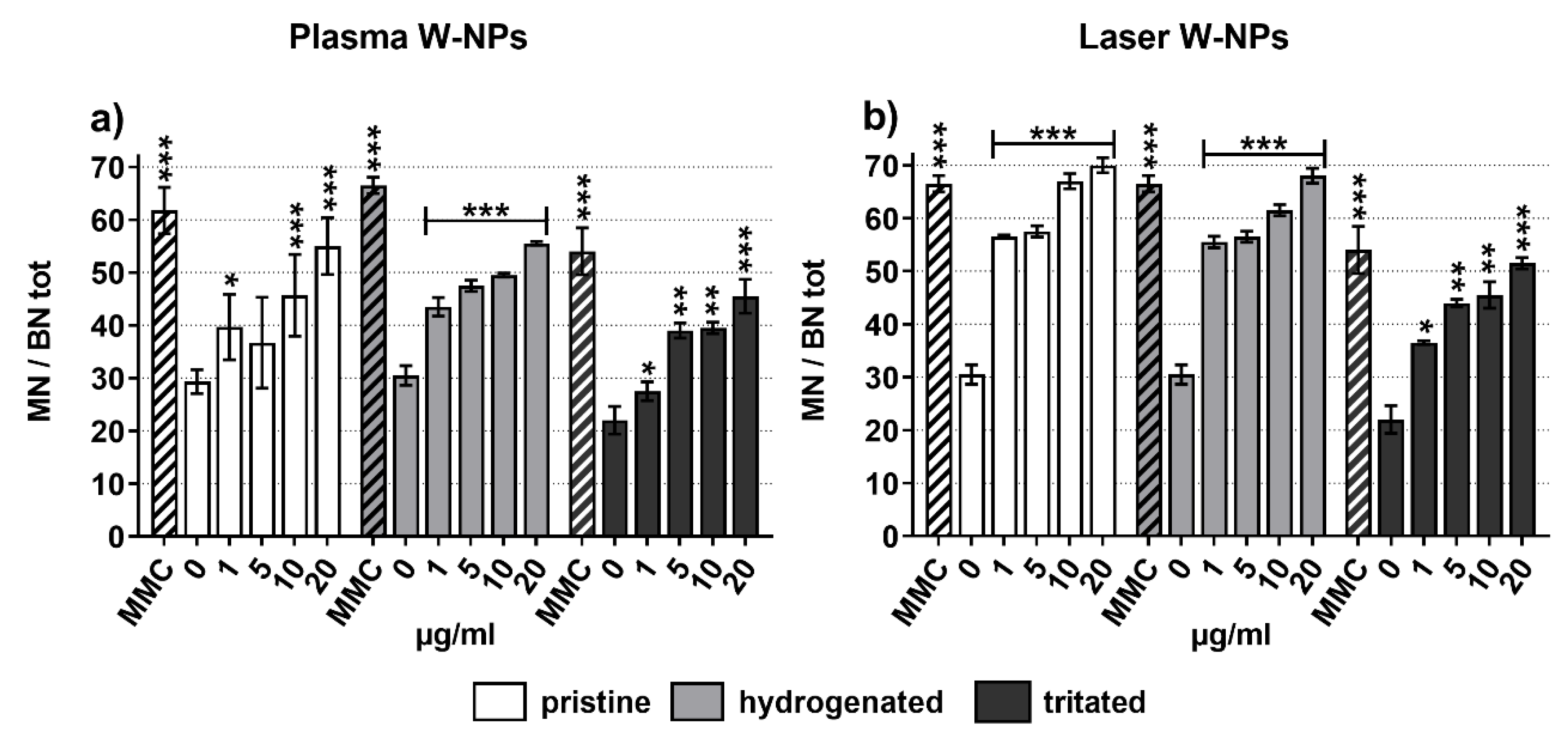
3.4. Micronuclei Formation
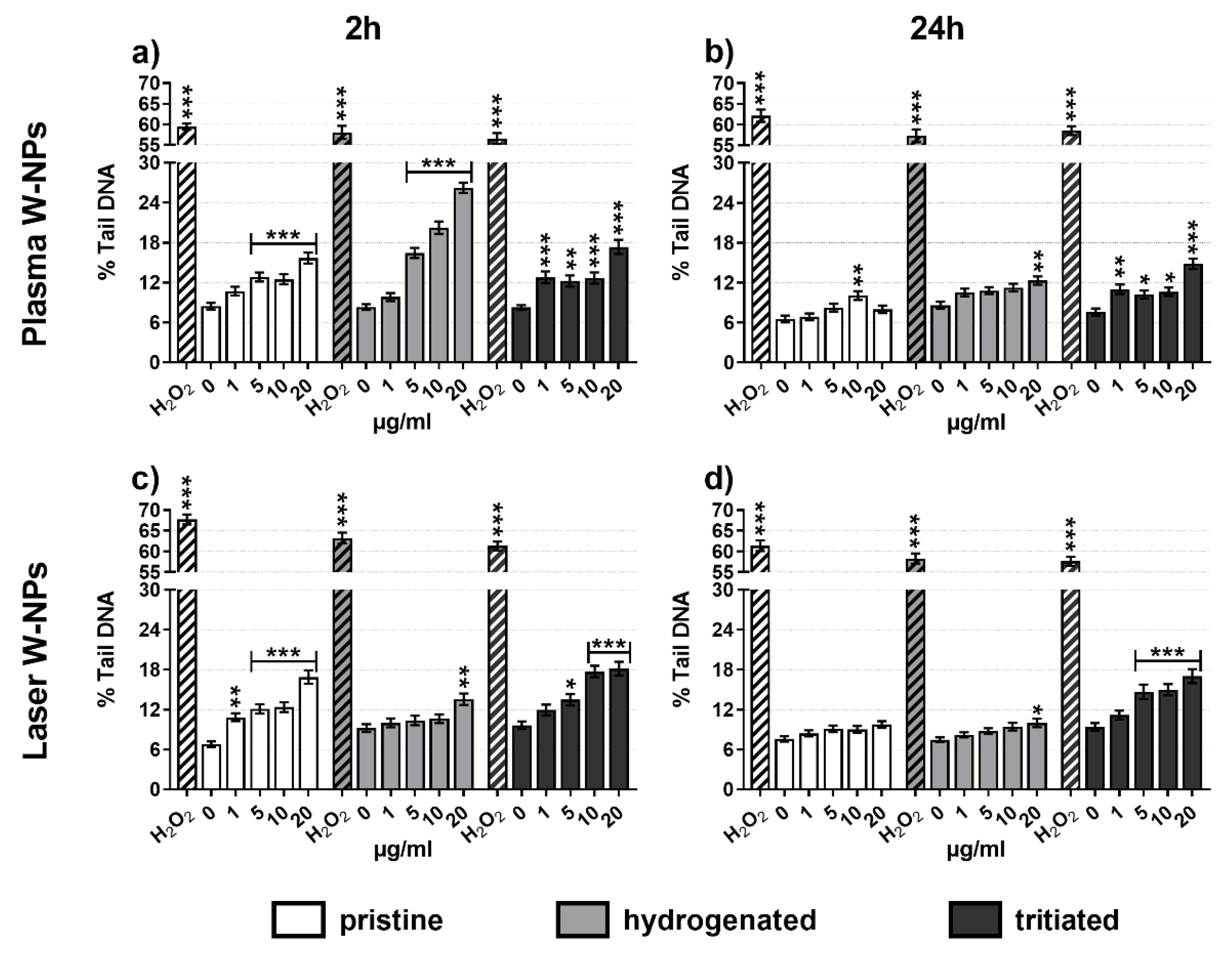
3.5. DNA Damage Quantification
3.6. Oxidative Stress
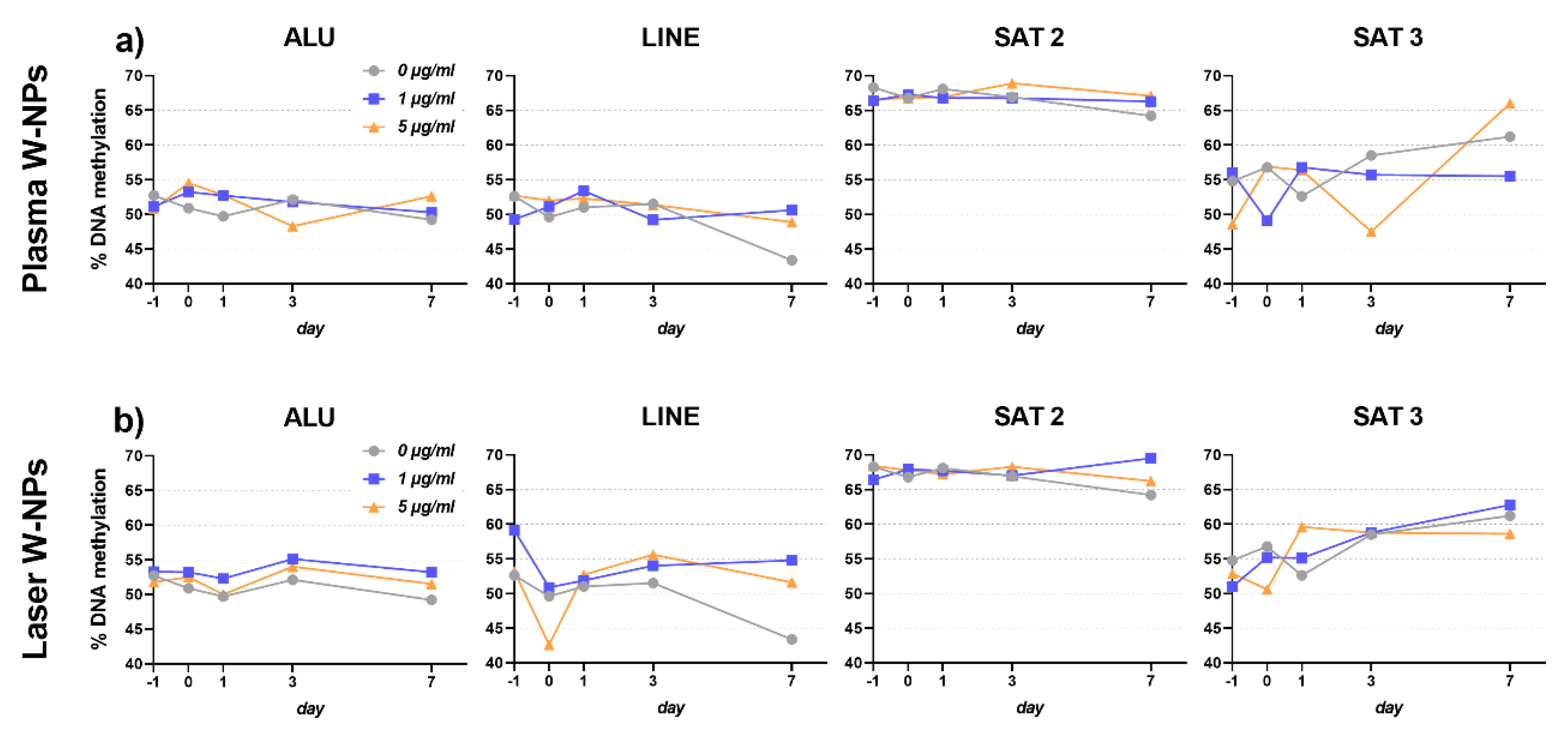
3.7. DNA Methylation Changes Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Davis, J.W.; Barabash, V.R.; Makhankov, A.; Plöchl, L.; Slattery, K.T. Assessment of tungsten for use in the ITER plasma facing components. J. Nucl. Mater. 1998, 258–263, 308–312. [Google Scholar] [CrossRef]
- Huang, S.-H.; Chen, C.-W.; Kuo, Y.-M.; Lai, C.-Y.; McKay, R.; Chen, C.-C. Factors Affecting Filter Penetration and Quality Factor of Particulate Respirators. Aerosol Air Qual. Res. 2013, 13, 162–171. [Google Scholar]
- Osterburg, A.R.; Robinson, C.T.; Schwemberger, S.; Mokashi, V.; Stockelman, M.; Babcock, G.F. Sodium tungstate (Na2WO4) exposure increases apoptosis in human peripheral blood lymphocytes. J. Immunotoxicol. 2010, 7, 174–182. [Google Scholar] [PubMed]
- Laulicht, F.; Brocato, J.; Cartularo, L.; Vaughan, J.; Wu, F.; Kluz, T.; Sun, H.; Oksuz, B.A.; Shen, S.; Peana, M.; et al. Tungsten-induced carcinogenesis in human bronchial epithelial cells. Toxicol. Appl. Pharmacol. 2015, 288, 33–39. [Google Scholar] [PubMed]
- Chinde, S.; Poornachandra, Y.; Panyala, A.; Kumari, S.I.; Yerramsetty, S.; Adicherla, H.; Grover, P. Comparative study of cyto-and genotoxic potential with mechanistic insights of tungsten oxide nano- and microparticles in lung carcinoma cells: Genotoxic potential of tungsten oxide nanoparticles in A549 cells. J. Appl. Toxicol. 2018, 38, 896–913. [Google Scholar]
- WHO International Agency for Research on Cancer. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; Cobalt in Hard Metals and Cobalt Sulfate, Gallium Arsenide, Indium Phosphide and Vanadium Pentoxide; World Health Organization: Geneva, Switzerland, 2006; Volume 86. [Google Scholar]
- Armstead, A.L.; Arena, C.B.; Li, B. Exploring the potential role of tungsten carbide cobalt (WC-Co) nanoparticle internalization in observed toxicity toward lung epithelial cells in vitro. Toxicol. Appl. Pharmacol. 2014, 278, 1–8. [Google Scholar] [CrossRef]
- Bastian, S.; Busch, W.; Kühnel, D.; Springer, A.; Meißner, T.; Holke, R.; Scholz, S.; Iwe, M.; Pompe, W.; Gelinsky, M.; et al. Toxicity of Tungsten Carbide and Cobalt-Doped Tungsten Carbide Nanoparticles in Mammalian Cells in vitro. Environ. Health Perspect. 2009, 117, 530–536. [Google Scholar] [PubMed]
- Moche, H.; Chevalier, D.; Barois, N.; Lorge, E.; Claude, N.; Nesslany, F. Tungsten Carbide-Cobalt as a Nanoparticulate Reference Positive Control in In Vitro Genotoxicity Assays. Toxicol. Sci. 2014, 137, 125–134. [Google Scholar] [PubMed]
- Paget, V.; Moche, H.; Kortulewski, T.; Grall, R.; Irbah, L.; Nesslany, F.; Chevillard, S. Human Cell Line-Dependent WC-Co Nanoparticle Cytotoxicity and Genotoxicity: A Key Role of ROS Production. Toxicol. Sci. 2015, 143, 385–397. [Google Scholar]
- Lison, D.; Carbonnelle, P.; Mollo, L.; Lauwerys, R.; Fubini, B. Physicochemical Mechanism of the Interaction between Cobalt Metal and Carbide Particles to Generate Toxic Activated Oxygen Species. Chem. Res. Toxicol. 1995, 8, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Machado, B.; Suro, R.M.; Garza, K.M.; Murr, L. Comparative microstructures and cytotoxicity assays for ballistic aerosols composed of micrometals and nanometals: Respiratory health implications. Int. J. Nanomed. 2011, 6, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Courcot, E.; Leclerc, J.; Lafitte, J.-J.; Mensier, E.; Jaillard, S.; Gosset, P.; Shirali, P.; Pottier, N.; Broly, F.; Lo-Guidice, J.-M. Xenobiotic metabolism and disposition in human lung cell models: Comparison with in vivo expression profiles. Drug Metab. Dispos. Biol. Fate Chem. 2012, 40, 1953–1965. [Google Scholar]
- Forbes, B. Human airway epithelial cell lines for in vitro drug transport and metabolism studies. Pharm. Sci. Technol. Today 2000, 3, 18–27. [Google Scholar] [CrossRef]
- Bernard, E.; Delaporte, P.; Jambon, F.; Rousseau, B.; Grisolia, C.; Chaudanson, D.; Nitsche, S. Tungsten dust in fusion tokamaks: Relevant dust laser production, characterization and behaviour under tritium loading. Phys. Scr. 2016, T167, 014071. [Google Scholar]
- Bernard, E.; Jambon, F.; Georges, I.; Sanles Sobrido, M.; Rose, J.; Herlin-Boime, N.; Miserque, F.; Beaunier, P.; Vrel, D.; Dine, S.; et al. Design of model tokamak particles for future toxicity studies: Morphology and physical characterization. Fusion Eng. Des. 2019, 145, 60–65. [Google Scholar]
- Sanles Sobrido, M.; Bernard, E.; Angeletti, B.; Malard, V.; Georges, I.; Chaurand, P.; Uboldi, C.; Orsière, T.; Dine, S.; Vrel, D.; et al. Oxidative transformation of Tungsten (W) nanoparticles released in aqueous and biological media in case of Tokamak (nuclear fusion) Lost of Vacuum Accident (LOVA). Environ. Sci. Nano 2019, in press. [Google Scholar]
- Acsente, T.; Negrea, R.F.; Nistor, L.C.; Logofatu, C.; Matei, E.; Birjega, R.; Grisolia, C.; Dinescu, G. Synthesis of flower-like tungsten nanoparticles by magnetron sputtering combined with gas aggregation. Eur. Phys. J. D 2015, 69. [Google Scholar]
- Vatry, A.; Habib, M.N.; Delaporte, P.; Sentis, M.; Grojo, D.; Grisolia, C.; Rosanvallon, S. Experimental investigation on laser removal of carbon and tungsten particles. Appl. Surf. Sci. 2009, 255, 5569–5573. [Google Scholar] [CrossRef]
- Uboldi, C.; Orsière, T.; Darolles, C.; Aloin, V.; Tassistro, V.; George, I.; Malard, V. Poorly soluble cobalt oxide particles trigger genotoxicity via multiple pathways. Part. Fibre Toxicol. 2015, 13, 5. [Google Scholar]
- Fenech, M. Cytokinesis-block micronucleus cytome assay. Nat. Protoc. 2007, 2, 1084–1104. [Google Scholar] [PubMed]
- Gonzalez, L.; Sanderson, B.J.S.; Kirsch-Volders, M. Adaptations of the in vitro MN assay for the genotoxicity assessment of nanomaterials. Mutagenesis 2011, 26, 185–191. [Google Scholar] [PubMed]
- OECD. Test No. 487: In Vitro Mammalian Cell Micronucleus Test; OECD Publishing: Paris, France, 2006. [Google Scholar]
- Dion, C.; Roche, S.; Laberthonnière, C.; Broucqsault, N.; Mariot, V.; Xue, S.; Gurzau, A.D.; Nowak, A.; Gordon, C.T.; Gaillard, M.-C.; et al. SMCHD1 is involved in de novo methylation of the DUX4-encoding D4Z4 macrosatellite. Nucleic Acids Res. 2019, 47, 2822–2839. [Google Scholar] [PubMed]
- Turkez, H.; Cakmak, B.; Celik, K. Evaluation of the Potential In Vivo Genotoxicity of Tungsten (VI) Oxide Nanopowder for Human Health. Key Eng. Mater. 2013, 543, 89–92. [Google Scholar]
- Turkez, H.; Sonmez, E.; Turkez, O.; Mokhtar, Y.I.; Stefano, A.D.; Turgut, G. The Risk Evaluation of Tungsten Oxide Nanoparticles in Cultured Rat Liver Cells for Its Safe Applications in Nanotechnology. Braz. Arch. Biol. Technol. 2014, 57, 532–541. [Google Scholar]
- Verma, R.; Xu, X.; Jaiswal, M.K.; Olsen, C.; Mears, D.; Caretti, G.; Galdzicki, Z. In vitro profiling of epigenetic modifications underlying heavy metal toxicity of tungsten-alloy and its components. Toxicol. Appl. Pharmacol. 2011, 253, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Laulicht-Glick, F.; Wu, F.; Zhang, X.; Jordan, A.; Brocato, J.; Kluz, T.; Sun, H.; Costa, M. Tungsten exposure causes a selective loss of histone demethylase protein. Mol. Carcinog. 2017, 56, 1778–1788. [Google Scholar] [PubMed]
- Leggett, R.W. A model of the distribution and retention of tungsten in the human body. Sci. Total Environ. 1997, 206, 147–165. [Google Scholar] [CrossRef]
- OSHA. Occupational Safety and Health Standards for Shipyard Employment. Air Contaminants. Code of Federal Regulations 29 CFR 1915.1000; Occupational Safety and Health Administration: Washington, DC, USA, 2005.
- OSHA. Safety and Health Regulations for Construction. Gases, Vapors, Fumes, Dusts, and Mists. Code of Federal Regulations 29 CFR 1926.55; Occupational Safety and Health Administration: Washington, DC, USA, 2005.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day −1 | Cells collected before exposure (generation F0) |
| Day 0 | Cells collected at the time of their exposure to plasma and laser ITER-like W-NPs (generation F0) |
| Day 1 | Cells collected at the end of the 24 h exposure (generation F0) |
| Day 3 | Cells collected 2 days after the end of the exposure (generation F0) |
| Day 7 | Day 3 cells passaged and collected at day 6 after the end of the exposure (generation F1) |
| Plasma W-NPs | ||
| Synthesis | Magnetron Sputtering | |
| Electron microscopy | Scanning electron microscopy | 100–200 nm mean size |
| Transmission electron microscopy | Inhomogeneous shape: star-like, squared and round | |
| Crystalline structure | Fourier transform pattern | Beta-phase W metal |
| XRD | Beta-phase W metal | |
| Density | Helium pycnometer | 14.32 g/cm3 |
| Surface area | BET | 4 m2/g |
| W/WOx ratio (in mass) | Helium pycnometer | 90% W metal, 10% WO3 |
| XAS | 90% W metal, 10% WO3 | |
| Laser W-NPs | ||
| Synthesis | High energy laser ablation | |
| Electron microscopy | Transmission electron microscopy | 60–80 nm mean size |
| Crystalline structure | XRD | Alpha-phase W metal |
| Density | Helium pycnometer | 8.27 g/cm3 |
| Surface area | BET | 43.5 m2/g |
| W/WOx ratio (in mass) | Helium pycnometer | 22% W metal, 78% WO3 |
| XAS | 18% W metal, 32% WO2, 50% WO3 | |
| t = 0 h 100 µg/mL | Plasma-Derived W-NPs | Laser-Derived W-NPs | ||||
|---|---|---|---|---|---|---|
| Mean Size (nm) | PDI * | Z Pot (mV) | Mean Size (nm) | PDI | Z Pot (mV) | |
| TRIS buffer | ||||||
| Pristine | 122 | 0.016 | −44 | 314 | 0.290 | −24 |
| Hydrogenated | 107 | 0.090 | −39 | 265 | 0.330 | −32 |
| Tritiated | 126 | 0.120 | −31 | 284 | 0.301 | −27 |
| LHC-9 culture medium | ||||||
| Pristine | 103 | 0.116 | −8 | 162 | 0.400 | −7 |
| Hydrogenated | 122 | 0.250 | −10 | 155 | 0.350 | −11 |
| Tritiated | 135 | 0.210 | −13 | 174 | 0.500 | −10 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uboldi, C.; Sanles Sobrido, M.; Bernard, E.; Tassistro, V.; Herlin-Boime, N.; Vrel, D.; Garcia-Argote, S.; Roche, S.; Magdinier, F.; Dinescu, G.; et al. In Vitro Analysis of the Effects of ITER-Like Tungsten Nanoparticles: Cytotoxicity and Epigenotoxicity in BEAS-2B Cells. Nanomaterials 2019, 9, 1233. https://doi.org/10.3390/nano9091233
Uboldi C, Sanles Sobrido M, Bernard E, Tassistro V, Herlin-Boime N, Vrel D, Garcia-Argote S, Roche S, Magdinier F, Dinescu G, et al. In Vitro Analysis of the Effects of ITER-Like Tungsten Nanoparticles: Cytotoxicity and Epigenotoxicity in BEAS-2B Cells. Nanomaterials. 2019; 9(9):1233. https://doi.org/10.3390/nano9091233
Chicago/Turabian StyleUboldi, Chiara, Marcos Sanles Sobrido, Elodie Bernard, Virginie Tassistro, Nathalie Herlin-Boime, Dominique Vrel, Sébastien Garcia-Argote, Stéphane Roche, Fréderique Magdinier, Gheorghe Dinescu, and et al. 2019. "In Vitro Analysis of the Effects of ITER-Like Tungsten Nanoparticles: Cytotoxicity and Epigenotoxicity in BEAS-2B Cells" Nanomaterials 9, no. 9: 1233. https://doi.org/10.3390/nano9091233
APA StyleUboldi, C., Sanles Sobrido, M., Bernard, E., Tassistro, V., Herlin-Boime, N., Vrel, D., Garcia-Argote, S., Roche, S., Magdinier, F., Dinescu, G., Malard, V., Lebaron-Jacobs, L., Rose, J., Rousseau, B., Delaporte, P., Grisolia, C., & Orsière, T. (2019). In Vitro Analysis of the Effects of ITER-Like Tungsten Nanoparticles: Cytotoxicity and Epigenotoxicity in BEAS-2B Cells. Nanomaterials, 9(9), 1233. https://doi.org/10.3390/nano9091233