
Asymmetric Lipid Transfer between Zwitterionic Vesicles by Nanoviscosity Measurements

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials Used
2.2. Lipid Vesicle Preparation
2.3. Dynamic Light Scattering and Zeta Potential Measurements
2.4. Quartz Crystal Microbalance with Dissipation Monitoring
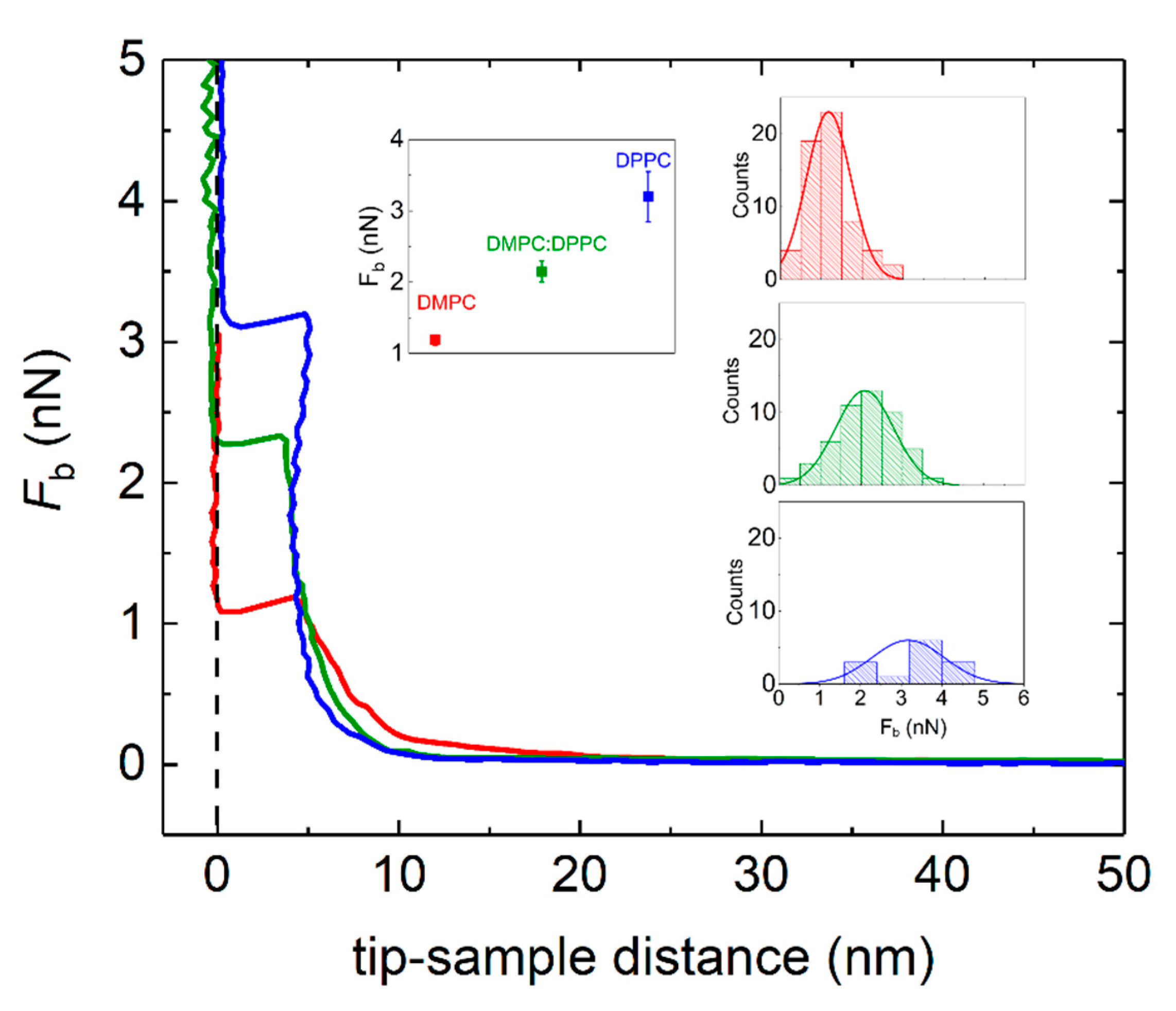
2.5. Atomic Force Microscopy (AFM)
2.6. Contact Angle Measurements
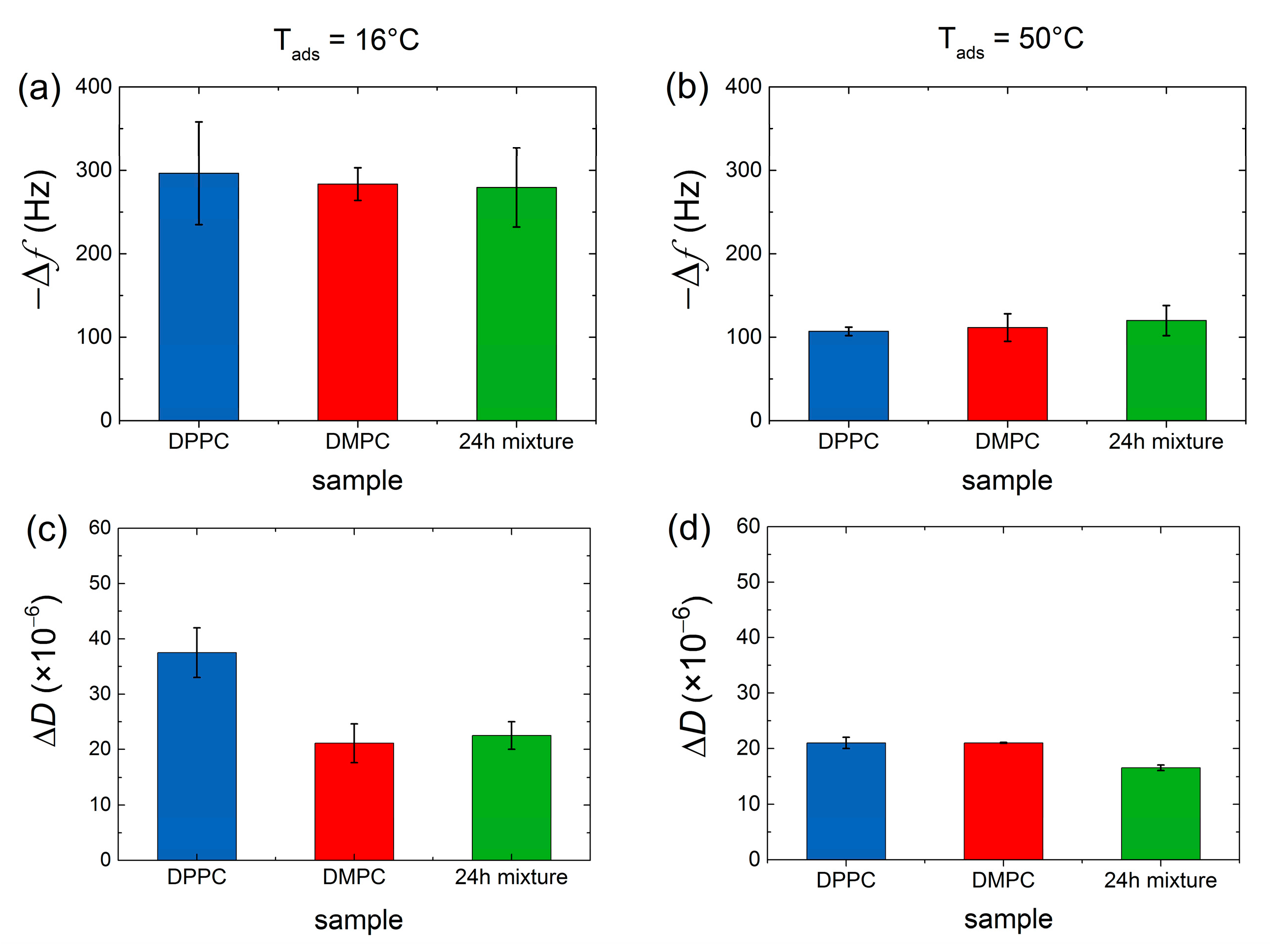
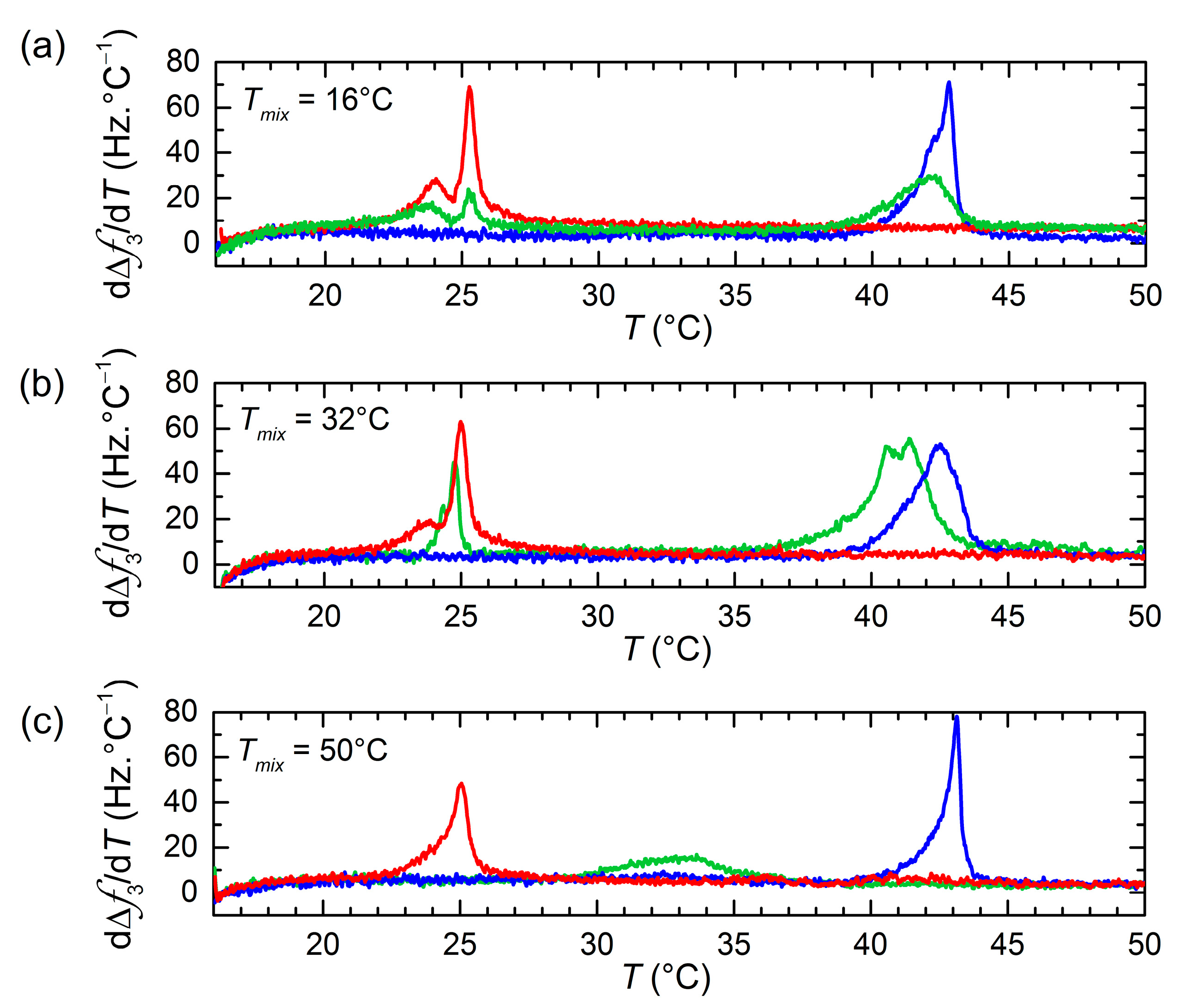
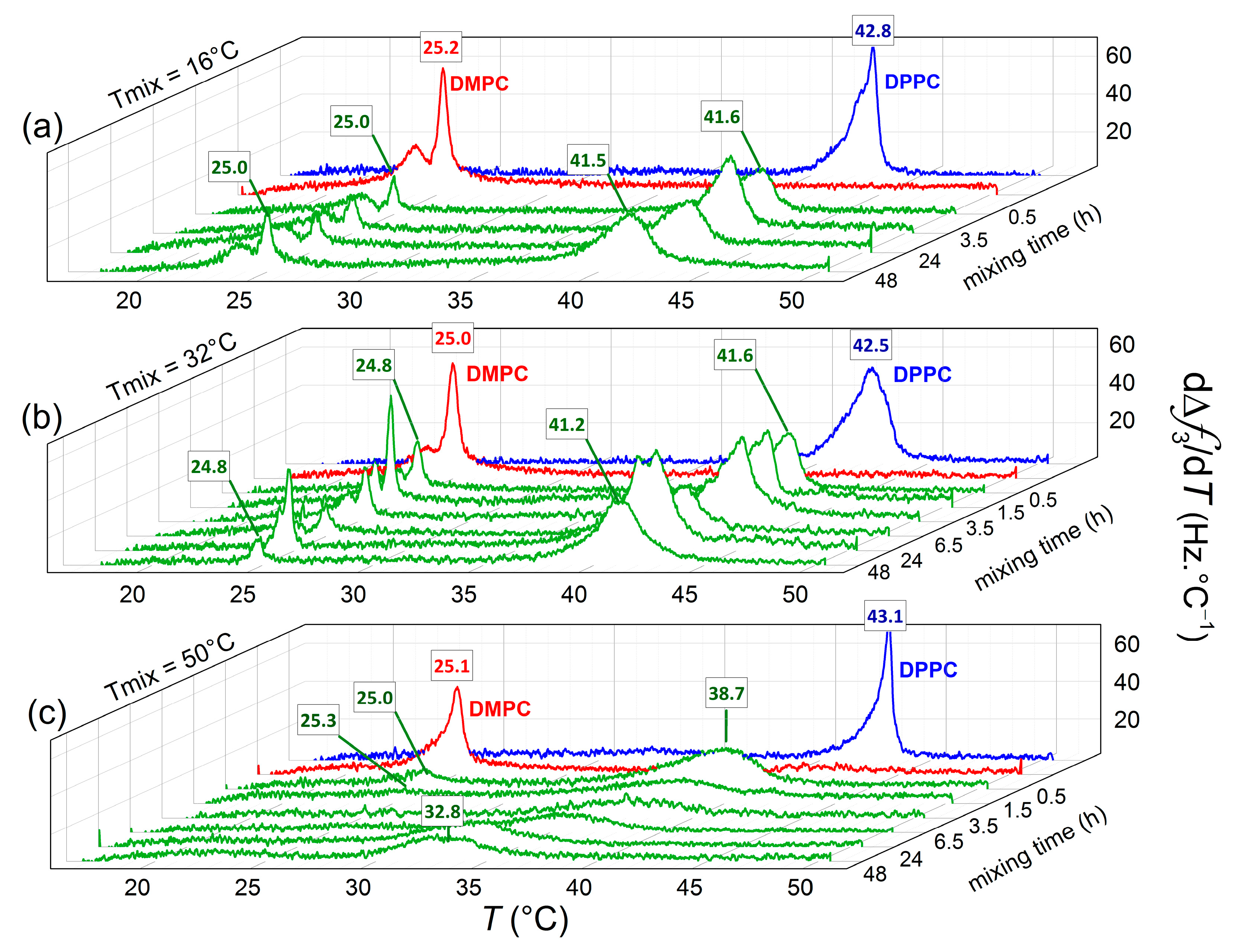
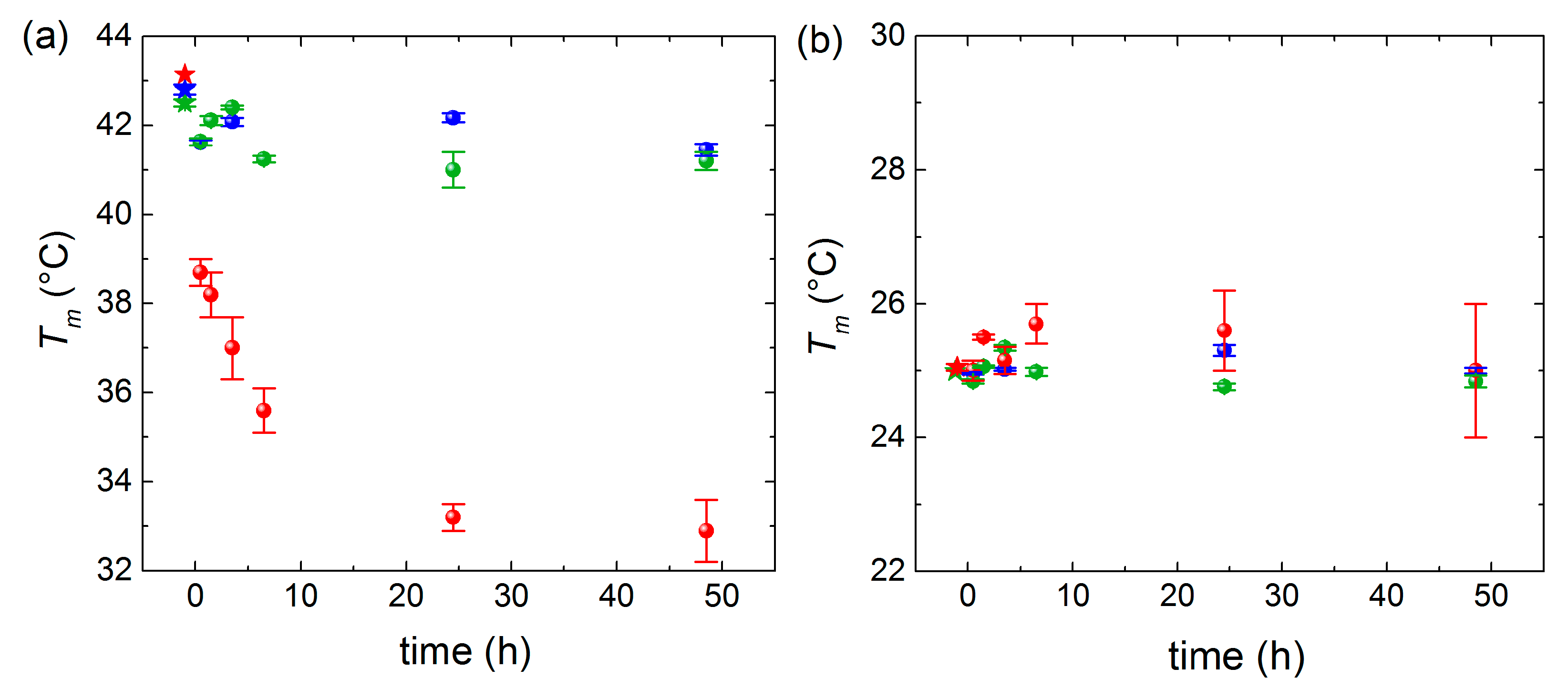
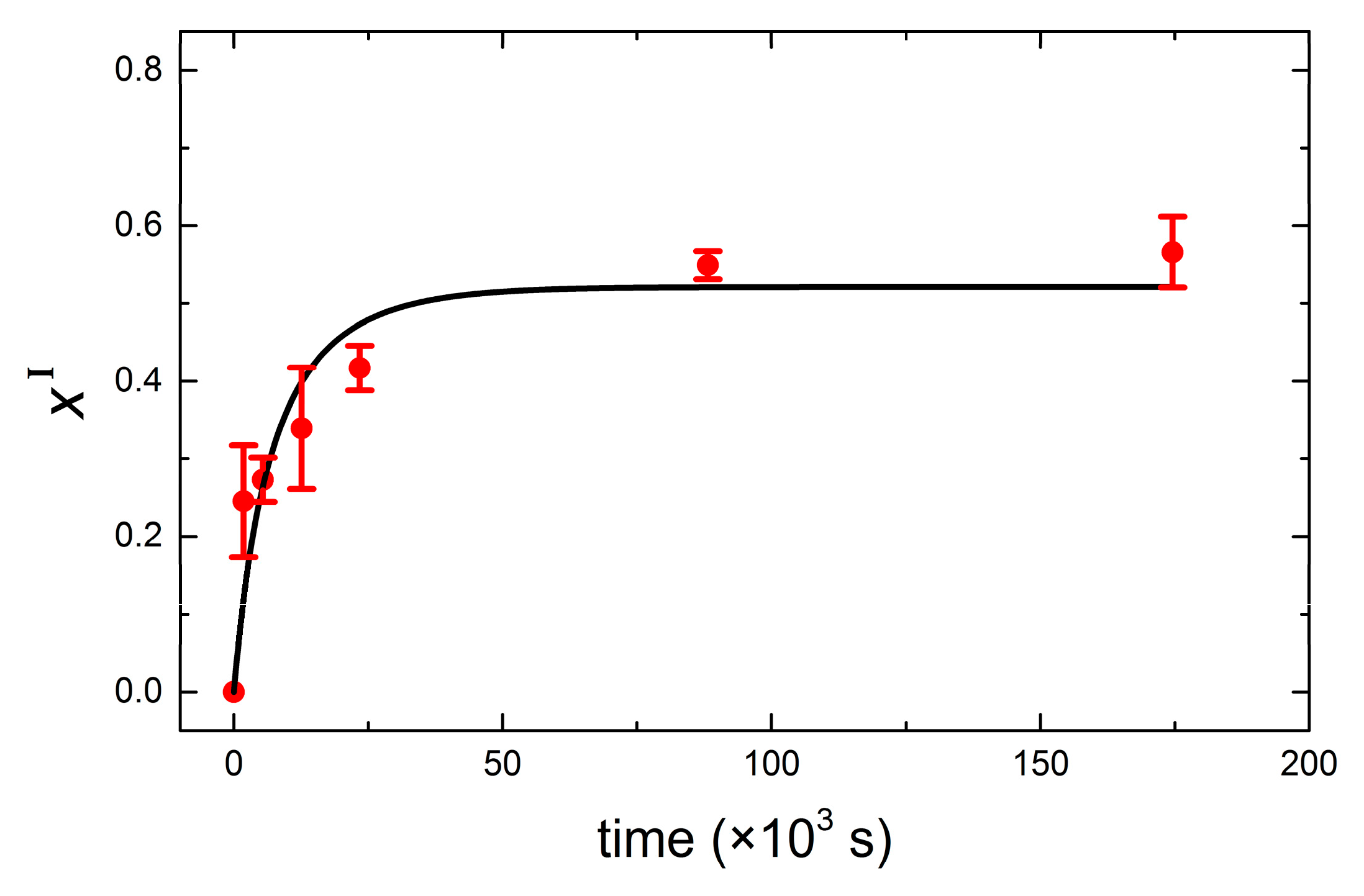
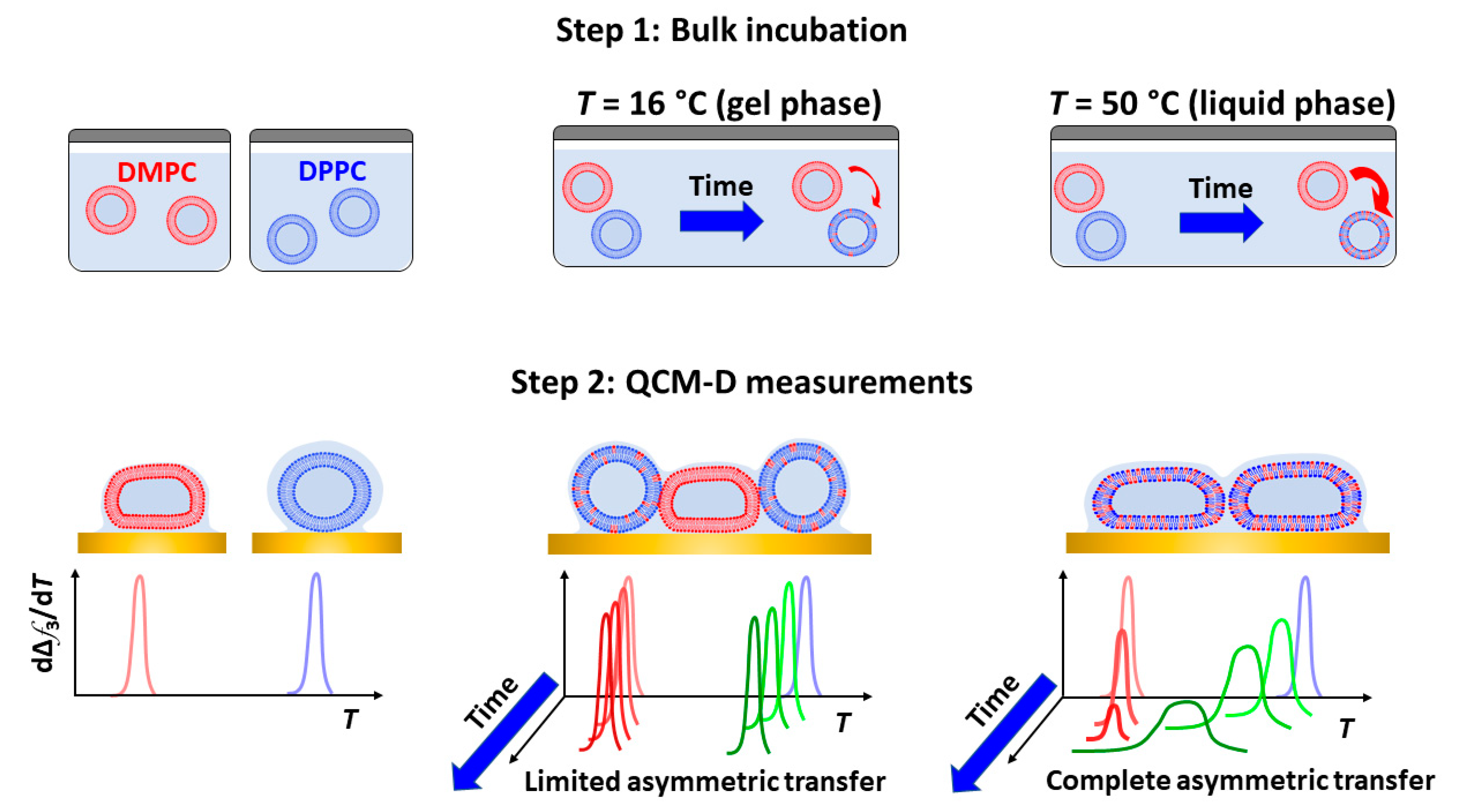
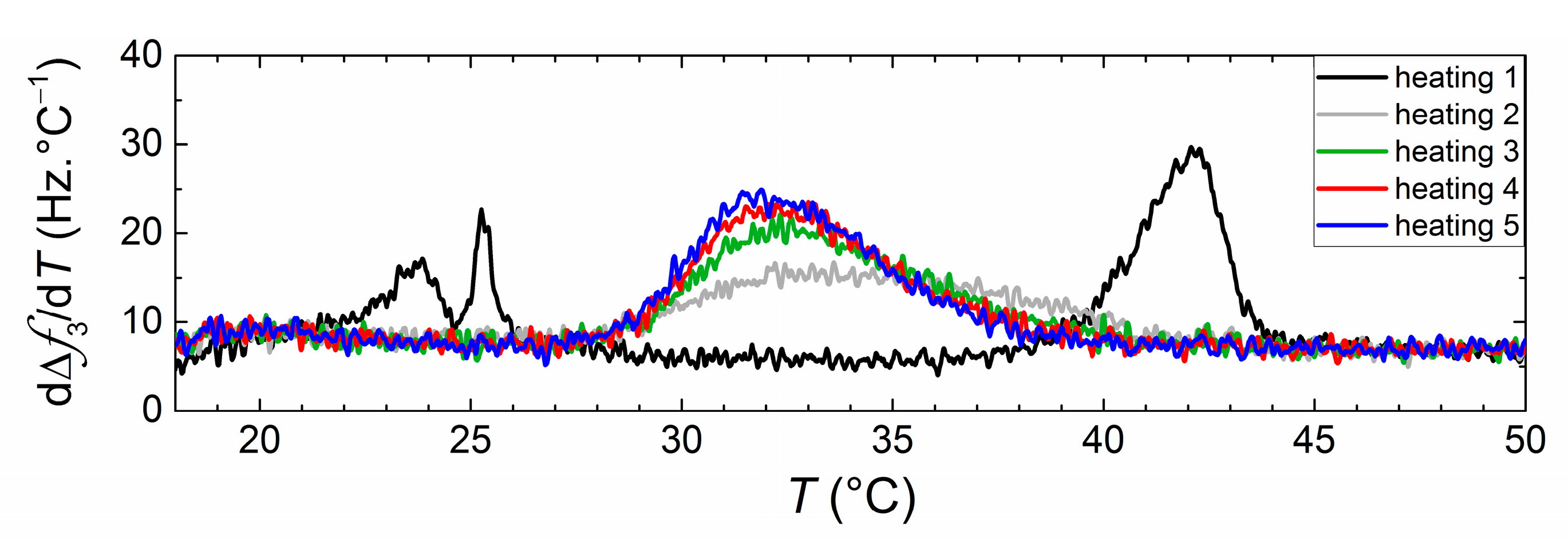
3. Results and Discussion
QCM-D Results
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Toulmay, A.; Prinz, W.A. Lipid transfer and signaling at organelle contact sites: The tip of the iceberg. Curr. Opin. Cell Biol. 2011, 23, 458–463. [Google Scholar] [CrossRef]
- Mazzon, M.; Mercer, J. Lipid interactions during virus entry and infection. Cell. Microbiol. 2014, 16, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Postila, P.A.; Róg, T. A Perspective: Active Role of Lipids in Neurotransmitter Dynamics. Mol. Neurobiol. 2020, 57, 910–925. [Google Scholar] [CrossRef]
- Stefan, C.J.; Trimble, W.S.; Grinstein, S.; Drin, G.; Reinisch, K.; De Camilli, P.; Cohen, S.; Valm, A.M.; Lippincott-Schwartz, J.; Levine, T.P.; et al. Membrane dynamics and organelle biogenesis—lipid pipelines and vesicular carriers. BMC Biol. 2017, 15, 102. [Google Scholar] [CrossRef]
- Reinisch, K.M.; Prinz, W.A. Mechanisms of nonvesicular lipid transport. J. Cell Biol. 2021, 220, 202012058. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.F. Spontaneous lipid transfer between organized lipid assemblies. Biochim. et Biophys. Acta (BBA) - Rev. Biomembr. 1992, 1113, 375–389. [Google Scholar] [CrossRef]
- Richens, J.L.; Tyler, A.I.I.; Barriga, H.M.G.; Bramble, J.P.; Law, R.V.; Brooks, N.J.; Seddon, J.M.; Ces, O.; O’Shea, P. Spontaneous charged lipid transfer between lipid vesicles. Sci. Rep. 2017, 7, 12606. [Google Scholar] [CrossRef] [PubMed]
- Golan, D.E.; Furlong, S.T.; Brown, C.S.; Caulfield, J.P. Monopalmitoylphosphatidylcholine incorporation into human erythrocyte ghost membranes causes protein and lipid immobilization and cholesterol depletion. Biochemistry 1988, 27, 2661–2667. [Google Scholar] [CrossRef] [PubMed]
- Staggers, J.E.; Hernell, O.; Stafford, R.J.; Carey, M.C. Physical-chemical behavior of dietary and biliary lipids during intestinal digestion and absorption. 1. Phase behavior and aggregation states of model lipid systems patterned after aqueous duodenal contents of healthy adult human beings. Biochemistry 1990, 29, 2028–2040. [Google Scholar] [CrossRef]
- Somerharju, P. Is Spontaneous Translocation of Polar Lipids Between Cellular Organelles Negligible? Lipid Insights 2015, 8s1, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.W.; Pagano, R.E. Kinetics of soluble lipid monomer diffusion between vesicles. Biochemistry 1981, 20, 2783–2789. [Google Scholar] [CrossRef]
- Zhu, T.; Jiang, Z.; Ma, Y. Lipid exchange between membranes: Effects of membrane surface charge, composition, and curvature. Colloids Surfaces B: Biointerfaces 2012, 97, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Lei, G.; Macdonald, R.C. Lipid Bilayer Vesicle Fusion: Intermediates Captured by High-Speed Microfluorescence Spectroscopy. Biophys. J. 2003, 85, 1585–1599. [Google Scholar] [CrossRef]
- Elvington, S.M.; Nichols, J.W. Spontaneous, intervesicular transfer rates of fluorescent, acyl chain-labeled phosphatidylcholine analogs. Biochim. et Biophys. Acta (BBA) - Biomembr. 2007, 1768, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Lira, R.B.; Robinson, T.; Dimova, R.; Riske, K.A. Highly Efficient Protein-free Membrane Fusion: A Giant Vesicle Study. Biophys. J. 2019, 116, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Kunze, A.; Sjövall, P.; Kasemo, B.; Svedhem, S. In Situ Preparation and Modification of Supported Lipid Layers by Lipid Transfer from Vesicles Studied by QCM-D and TOF-SIMS. J. Am. Chem. Soc. 2009, 131, 2450–2451. [Google Scholar] [CrossRef]
- Margheri, G.; D’Agostino, R.; Becucci, L.; Guidelli, R.; Tiribilli, B.; Del Rosso, M. Surface plasmon resonance as detection tool for lipids lateral mobility in biomimetic membranes. Biomed. Opt. Express 2012, 3, 3119–3126. [Google Scholar] [CrossRef] [PubMed]
- Reviakine, I.; Johannsmann, D.; Richter, R.P. Hearing What You Cannot See and Visualizing What You Hear: Interpreting Quartz Crystal Microbalance Data from Solvated Interfaces. Anal. Chem. 2011, 83, 8838–8848. [Google Scholar] [CrossRef] [PubMed]
- Wikström, A.; Svedhem, S.; Sivignon, M.; Kasemo, B. Real-Time QCM-D Monitoring of Electrostatically Driven Lipid Transfer between Two Lipid Bilayer Membranes. J. Phys. Chem. B 2008, 112, 14069–14074. [Google Scholar] [CrossRef] [PubMed]
- Kunze, A.; Svedhem, S.; Kasemo, B. Lipid Transfer between Charged Supported Lipid Bilayers and Oppositely Charged Vesicles. Langmuir 2009, 25, 5146–5158. [Google Scholar] [CrossRef]
- Tabaei, S.R.; Gillissen, J.J.J.; Vafaei, S.; Groves, J.T.; Cho, N.-J. Size-dependent, stochastic nature of lipid exchange between nano-vesicles and model membranes. Nanoscale 2016, 8, 13513–13520. [Google Scholar] [CrossRef]
- Ohlsson, G.; Tigerström, A.; Höök, F.; Kasemo, B. Phase transitions in adsorbed lipid vesicles measured using a quartz crystal microbalance with dissipation monitoring. Soft Matter 2011, 7, 10749–10755. [Google Scholar] [CrossRef]
- Neupane, S.; De Smet, Y.; Renner, F.U.; Losada-Pérez, P. Quartz Crystal Microbalance With Dissipation Monitoring: A Versatile Tool to Monitor Phase Transitions in Biomimetic Membranes. Front. Mater. 2018, 5, 46. [Google Scholar] [CrossRef]
- Peschel, A.; Langhoff, A.; Uhl, E.; Dathathreyan, A.; Haindl, S.; Johannsmann, D.; Reviakine, I. Lipid phase behavior studied with a quartz crystal microbalance: A technique for biophysical studies with applications in screening. J. Chem. Phys. 2016, 145, 204904. [Google Scholar] [CrossRef]
- Pramanik, S.K.; Seneca, S.; Ethirajan, A.; Neupane, S.; Renner, F.U.; Losada-Pérez, P. Ionic strength dependent vesicle adsorption and phase behavior of anionic phospholipids on a gold substrate. Biointerphases 2016, 11, 019006. [Google Scholar] [CrossRef] [PubMed]
- Hasan, I.Y.; Mechler, A. Cholesterol Rich Domains Identified in Unilamellar Supported Biomimetic Membranes via Nano-Viscosity Measurements. Anal. Chem. 2016, 88, 5037–5041. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Enoki, T.A.; Henriques, V.B.; Lamy, M.T. Light scattering on the structural characterization of DMPG vesicles along the bilayer anomalous phase transition. Chem. Phys. Lipids 2012, 165, 826–837. [Google Scholar] [CrossRef]
- Lipowsky, R.; Seifert, U. Adhesion of Vesicles and Membranes. Mol. Cryst. Liq. Cryst. 1991, 202, 17–25. [Google Scholar] [CrossRef]
- Florin, E.-L.; Rief, M.; Lehmann, H.; Ludwig, M.; Dornmair, C.; Moy, V.; Gaub, H. Sensing specific molecular interactions with the atomic force microscope. Biosens. Bioelectron. 1995, 10, 895–901. [Google Scholar] [CrossRef]
- Bibissidis, N.; Betlem, K.; Cordoyiannis, G.; Bonhorst, F.P.-V.; Goole, J.; Raval, J.; Daniel, M.; Góźdź, W.; Iglič, A.; Losada-Pérez, P. Correlation between adhesion strength and phase behaviour in solid-supported lipid membranes. J. Mol. Liq. 2020, 320, 114492. [Google Scholar] [CrossRef]
- Lind, T.K.; Cárdenas, M. Understanding the formation of supported lipid bilayers via vesicle fusion—A case that exemplifies the need for the complementary method approach (Review). Biointerphases 2016, 11, 020801. [Google Scholar] [CrossRef]
- Lee, C.-H.; Lin, W.-C.; Wang, J. All-optical measurements of the bending rigidity of lipid-vesicle membranes across structural phase transitions. Phys. Rev. E 2001, 64, 020901. [Google Scholar] [CrossRef]
- Losada-Pérez, P.; Mertens, N.; De Medio-Vasconcelos, B.; Slenders, E.; Leys, J.; Peeters, M.; Van Grinsven, B.; Gruber, J.; Glorieux, C.; Pfeiffer, H.; et al. Phase Transitions of Binary Lipid Mixtures: A Combined Study by Adiabatic Scanning Calorimetry and Quartz Crystal Microbalance with Dissipation Monitoring. Adv. Condens. Matter Phys. 2015, 2015, 1–14. [Google Scholar] [CrossRef]
- Leys, J.; Losada-Pérez, P.; Slenders, E.; Glorieux, C.; Thoen, J. Investigation of the melting behavior of the reference materials biphenyl and phenyl salicylate by a new type adiabatic scanning calorimeter. Thermochim. Acta 2014, 582, 68–76. [Google Scholar] [CrossRef]
- Ebel, H.; Grabitz, P.; Heimburg, T. Enthalpy and Volume Changes in Lipid Membranes. I. The Proportionality of Heat and Volume Changes in the Lipid Melting Transition and Its Implication for the Elastic Constants. J. Phys. Chem. B 2001, 105, 7353–7360. [Google Scholar] [CrossRef]
- Betlem, K.; Cordoyiannis, G.; Losada-Pérez, P. Supported Lipid Membranes at the Au-Buffer Interface by Solvent Exchange: The Effect of Initial Solvent Concentration. Phys. Status Solidi (a) 2020, 217, 1900837. [Google Scholar] [CrossRef]
- Hasan, I.Y.; Mechler, A. Nanoviscosity Measurements Revealing Domain Formation in Biomimetic Membranes. Anal. Chem. 2017, 89, 1855–1862. [Google Scholar] [CrossRef]
- Thilo, L. Kinetics of phospholipid exchange between bilayer membranes. Biochim. Biophys. Acta (BBA) - Biomembr. 1977, 469, 326–334. [Google Scholar] [CrossRef]
- Bayerl, T.M.; Schmidt, C.F.; Sackmann, E. Kinetics of symmetric and asymmetric phospholipid transfer between small sonicated vesicles studied by high-sensitivity differential scanning calorimetry, NMR, electron microscopy, and dynamic light scattering. Biochemistry 1988, 27, 6078–6085. [Google Scholar] [CrossRef]
- Rogers, J.R.; Espinoza-García, G.; Geissler, P.L. Membrane Hydrophobicity Determines the Activation Free Energy of Passive Lipid Transport. BioRxiv 2021. Available online: https://www.biorxiv.org/content/10.1101/2021.03.17.435885v1 (accessed on 29 March 2021). [CrossRef]
- Nakano, M.; Fukuda, M.; Kudo, T.; Endo, H.; Handa, T. Determination of Interbilayer and Transbilayer Lipid Transfers by Time-Resolved Small-Angle Neutron Scattering. Phys. Rev. Lett. 2007, 98, 238101. [Google Scholar] [CrossRef]
- Neupane, S.; Betlem, K.; Renner, F.U.; Losada-Pérez, P. Solvent-Assisted Lipid Bilayer Formation on Au Surfaces: Effect of Lipid Concentration on Solid-Supported Membrane Formation. Phys. Status solidi (a) 2020. [Google Scholar] [CrossRef]
- Li, M.; Chen, M.; Sheepwash, E.; Brosseau, C.L.; Li, H.; Pettinger, B.; Gruler, H.; Lipkowski, J. AFM Studies of Solid-Supported Lipid Bilayers Formed at a Au(111) Electrode Surface Using Vesicle Fusion and a Combination of Langmuir−Blodgett and Langmuir−Schaefer Techniques. Langmuir 2008, 24, 10313–10323. [Google Scholar] [CrossRef] [PubMed]
- Dufrêne, Y.F.; Barger, W.R.; Green, A.J.-B.D.; Lee, G.U. Nanometer-Scale Surface Properties of Mixed Phospholipid Monolayers and Bilayers. Langmuir 1997, 13, 4779–4784. [Google Scholar] [CrossRef]
- Redondo-Morata, L.; Giannotti, M.I.; Sanz, F. Influence of Cholesterol on the Phase Transition of Lipid Bilayers: A Temperature-Controlled Force Spectroscopy Study. Langmuir 2012, 28, 12851–12860. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid | Temperature (°C) | Mean Diameter (nm) | PI |
|---|---|---|---|
| DPPC | 16 | 119 ± 6 | 0.12 ± 0.06 |
| DMPC | 16 | 118 ± 9 | 0.05 ± 0.01 |
| DPPC | 32 | 110 ± 3 | 0.08 ±0.01 |
| DMPC | 32 | 120 ± 3 | 0.08 ± 0.01 |
| DPPC | 50 | 130 ± 5 | 0.07 ± 0.02 |
| DMPC | 50 | 126 ± 7 | 0.09 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bar, L.; Cordoyiannis, G.; Neupane, S.; Goole, J.; Grosfils, P.; Losada-Pérez, P. Asymmetric Lipid Transfer between Zwitterionic Vesicles by Nanoviscosity Measurements. Nanomaterials 2021, 11, 1087. https://doi.org/10.3390/nano11051087
Bar L, Cordoyiannis G, Neupane S, Goole J, Grosfils P, Losada-Pérez P. Asymmetric Lipid Transfer between Zwitterionic Vesicles by Nanoviscosity Measurements. Nanomaterials. 2021; 11(5):1087. https://doi.org/10.3390/nano11051087
Chicago/Turabian StyleBar, Laure, George Cordoyiannis, Shova Neupane, Jonathan Goole, Patrick Grosfils, and Patricia Losada-Pérez. 2021. "Asymmetric Lipid Transfer between Zwitterionic Vesicles by Nanoviscosity Measurements" Nanomaterials 11, no. 5: 1087. https://doi.org/10.3390/nano11051087
APA StyleBar, L., Cordoyiannis, G., Neupane, S., Goole, J., Grosfils, P., & Losada-Pérez, P. (2021). Asymmetric Lipid Transfer between Zwitterionic Vesicles by Nanoviscosity Measurements. Nanomaterials, 11(5), 1087. https://doi.org/10.3390/nano11051087