Liposome-Mediated Inhibition of Inflammation by Hydroxycitrate

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Liposome Preparation and Characterization
2.3. Cell Culture and Treatments
2.4. Cytotoxicity Assay
2.5. ROS, NO, and PGE2 Detection
2.6. Cellular Uptake of HCA by HPLC–Tandem Mass Spectrometry
2.7. Statistical Analysis
3. Results
3.1. Liposomal Formulation
3.2. Effect of HCA-Loaded Liposomes on Cell Count
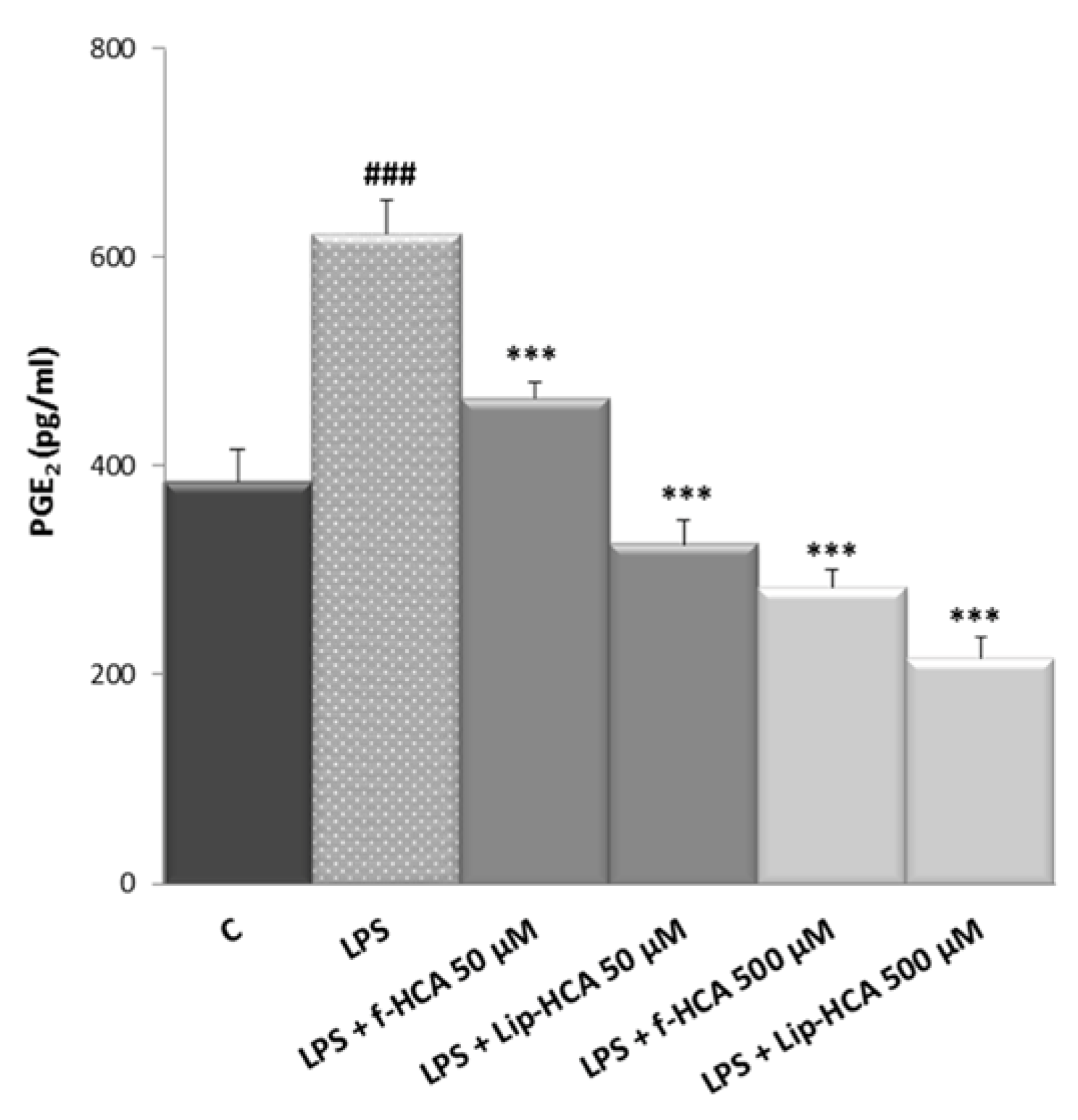
3.3. Effect of HCA-Loaded Liposomes on PGE2 Production
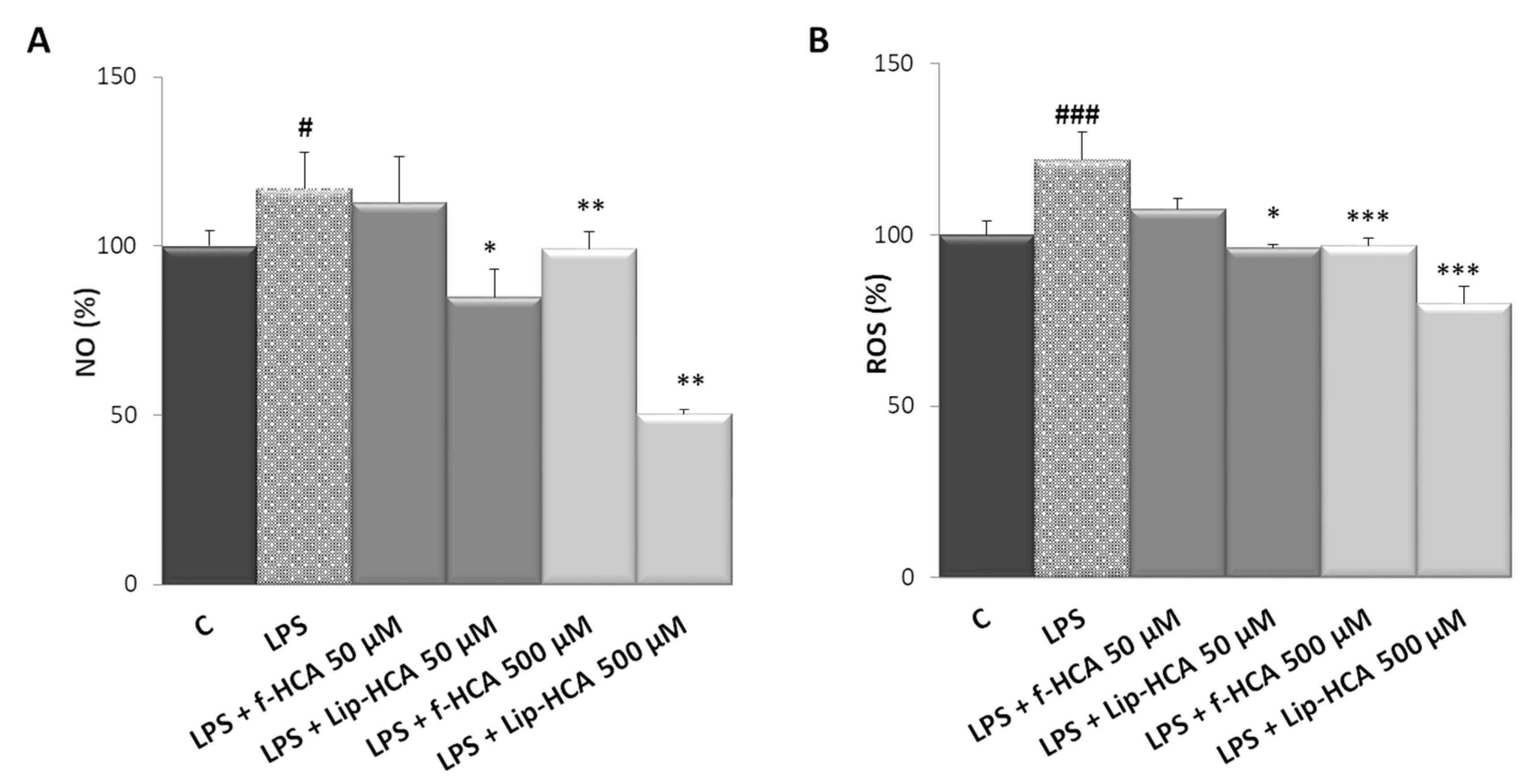
3.4. Effect of HCA-Loaded Liposomes on NO and ROS Production
3.5. Intracellular Uptake of HCA
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- O’Neill, L.A.; Hardie, D.G. Metabolism of inflammation limited by AMPK and pseudo-starvation. Nature 2013, 493, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Todisco, S.; Convertini, P.; Iacobazzi, V.; Infantino, V. TCA Cycle Rewiring as Emerging Metabolic Signature of Hepatocellular Carcinoma. Cancers 2019, 12, 68. [Google Scholar] [CrossRef] [Green Version]
- Ryan, D.G.; O’Neill, L.A.J. Krebs cycle rewired for macrophage and dendritic cell effector functions. FEBS Lett. 2017, 591, 2992–3006. [Google Scholar] [CrossRef] [Green Version]
- Iacobazzi, V.; Infantino, V. Citrate—New functions for an old metabolite. Biol. Chem. 2014, 395, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.T.; Yang, C.M. Role of NADPH oxidase/ROS in pro-inflammatory mediators-induced airway and pulmonary diseases. Biochem. Pharmacol. 2012, 84, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Lauterbach, M.A.; Hanke, J.E.; Serefidou, M.; Mangan, M.S.J.; Kolbe, C.C.; Hess, T.; Rothe, M.; Kaiser, R.; Hoss, F.; Gehlen, J.; et al. Toll-like Receptor Signaling Rewires Macrophage Metabolism and Promotes Histone Acetylation via ATP-Citrate Lyase. Immunity 2019, 51, 997–1011.e7. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Pillinger, M.H.; Abramson, S.B. Prostaglandin E2 synthesis and secretion: The role of PGE2 synthases. Clin. Immunol. 2006, 119, 229–240. [Google Scholar] [CrossRef]
- Convertini, P.; Menga, A.; Andria, G.; Scala, I.; Santarsiero, A.; Castiglione Morelli, M.A.; Iacobazzi, V.; Infantino, V. The contribution of the citrate pathway to oxidative stress in Down syndrome. Immunology 2016, 149, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Santarsiero, A.; Leccese, P.; Convertini, P.; Padula, A.; Abriola, P.; D’Angelo, S.; Bisaccia, F.; Infantino, V. New Insights into Behcet’s Syndrome Metabolic Reprogramming: Citrate Pathway Dysregulation. Mediat. Inflamm. 2018, 2018, 1419352. [Google Scholar] [CrossRef]
- Infantino, V.; Convertini, P.; Cucci, L.; Panaro, M.A.; Di Noia, M.A.; Calvello, R.; Palmieri, F.; Iacobazzi, V. The mitochondrial citrate carrier: A new player in inflammation. Biochem. J. 2011, 438, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Infantino, V.; Iacobazzi, V.; Menga, A.; Avantaggiati, M.L.; Palmieri, F. A key role of the mitochondrial citrate carrier (SLC25A1) in TNFalpha- and IFNgamma-triggered inflammation. Biochim. Biophys. Acta 2014, 1839, 1217–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Infantino, V.; Iacobazzi, V.; Palmieri, F.; Menga, A. ATP-citrate lyase is essential for macrophage inflammatory response. Biochem. Biophys. Res. Commun. 2013, 440, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Infantino, V.; Pierri, C.L.; Iacobazzi, V. Metabolic Routes in Inflammation: The Citrate Pathway and its Potential as Therapeutic Target. Curr. Med. Chem. 2019, 26, 7104–7116. [Google Scholar] [CrossRef]
- Abramson, H.N. The lipogenesis pathway as a cancer target. J. Med. Chem. 2011, 54, 5615–5638. [Google Scholar] [CrossRef] [PubMed]
- Semwal, R.B.; Semwal, D.K.; Vermaak, I.; Viljoen, A. A comprehensive scientific overview of Garcinia cambogia. Fitoterapia 2015, 102, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Caddeo, C.; Nacher, A.; Vassallo, A.; Armentano, M.F.; Pons, R.; Fernandez-Busquets, X.; Carbone, C.; Valenti, D.; Fadda, A.M.; Manconi, M. Effect of quercetin and resveratrol co-incorporated in liposomes against inflammatory/oxidative response associated with skin cancer. Int. J. Pharm. 2016, 513, 153–163. [Google Scholar] [CrossRef]
- Pham, A.; Bortolazzo, A.; White, J.B. Rapid dimerization of quercetin through an oxidative mechanism in the presence of serum albumin decreases its ability to induce cytotoxicity in MDA-MB-231 cells. Biochem. Biophys. Res. Commun. 2012, 427, 415–420. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chirollo, C.; Vassallo, A.; Dal Piaz, F.; Lamagna, B.; Tortora, G.; Neglia, G.; De Tommasi, N.; Severino, L. Investigation of the Persistence of Penicillin G and Dihydrostreptomycin Residues in Milk of Lactating Buffaloes (Bubalus bubalis) Using Ultra-High-Performance Liquid Chromatography and Tandem Mass Spectrometry. J. Agric. Food Chem. 2018, 66, 6388–6393. [Google Scholar] [CrossRef]
- Pandey, R.C.P.; Kumar, B.; Srivastva, B.; Aravind, A.; Rameshkumard, S.K.B. Simultaneous determination of multi-class bioactive constituents for quality assessment of Garcinia species using UHPLC–QqQLIT–MS/MS. Ind. Crops Prod. 2015, 77, 12. [Google Scholar] [CrossRef]
- Babior, B.M. NADPH oxidase. Curr. Opin. Immunol. 2004, 16, 42–47. [Google Scholar] [CrossRef]
- Shara, M.; Ohia, S.E.; Yasmin, T.; Zardetto-Smith, A.; Kincaid, A.; Bagchi, M.; Chatterjee, A.; Bagchi, D.; Stohs, S.J. Dose- and time-dependent effects of a novel (−)-hydroxycitric acid extract on body weight, hepatic and testicular lipid peroxidation, DNA fragmentation and histopathological data over a period of 90 days. Mol. Cell. Biochem. 2003, 254, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.M.; Cullis, P.R. Liposomal drug delivery systems: From concept to clinical applications. Adv. Drug Deliv. Rev. 2013, 65, 36–48. [Google Scholar] [CrossRef]
- Bozzuto, G.; Molinari, A. Liposomes as nanomedical devices. Int. J. Nanomed. 2015, 10, 975–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caddeo, C.; Manconi, M.; Sinico, C.; Valenti, D.; Celia, C.; Monduzzi, M.; Fadda, A.M. Penetration Enhancer-Containing Vesicles: Does the Penetration Enhancer Structure Affect Topical Drug Delivery? Curr. Drug Targets 2015, 16, 1438–1447. [Google Scholar] [CrossRef]
- Caddeo, C.; Pucci, L.; Gabriele, M.; Carbone, C.; Fernandez-Busquets, X.; Valenti, D.; Pons, R.; Vassallo, A.; Fadda, A.M.; Manconi, M. Stability, biocompatibility and antioxidant activity of PEG-modified liposomes containing resveratrol. Int. J. Pharm. 2018, 538, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Sercombe, L.; Veerati, T.; Moheimani, F.; Wu, S.Y.; Sood, A.K.; Hua, S. Advances and Challenges of Liposome Assisted Drug Delivery. Front. Pharmacol. 2015, 6, 286. [Google Scholar] [CrossRef] [Green Version]
- Ezhilarasi, P.N.; Muthukumar, S.P.; Anandharamakrishnan, C. Solid lipid nanoparticle enhances bioavailability of hydroxycitric acid compared to a microparticle delivery system. RSC Adv. 2016, 6, 10. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Mean Diameter | PI § | ZP | |
|---|---|---|---|
| (nm ± SD) | (mV ± SD) | ||
| Empty Liposomes | 63 ± 4.4 | 0.24 | −49 ± 13.8 |
| HCA Liposomes | 65 ± 9.1 | 0.26 | −59 ± 6.3 |
| Cell Number (% ± SD) | |||
|---|---|---|---|
| 0 µM | 50 µM | 500 µM | |
| f-HCA | 100 ± 3.6 | 103.5 ± 3.1 | 98.7 ± 2.5 |
| Lip-HCA | 100 ± 5.3 | 118.8 ± 2.4 | 92.3 ± 2.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vassallo, A.; Santoro, V.; Pappalardo, I.; Santarsiero, A.; Convertini, P.; De Luca, M.; Martelli, G.; Infantino, V.; Caddeo, C. Liposome-Mediated Inhibition of Inflammation by Hydroxycitrate. Nanomaterials 2020, 10, 2080. https://doi.org/10.3390/nano10102080
Vassallo A, Santoro V, Pappalardo I, Santarsiero A, Convertini P, De Luca M, Martelli G, Infantino V, Caddeo C. Liposome-Mediated Inhibition of Inflammation by Hydroxycitrate. Nanomaterials. 2020; 10(10):2080. https://doi.org/10.3390/nano10102080
Chicago/Turabian StyleVassallo, Antonio, Valentina Santoro, Ilaria Pappalardo, Anna Santarsiero, Paolo Convertini, Maria De Luca, Giuseppe Martelli, Vittoria Infantino, and Carla Caddeo. 2020. "Liposome-Mediated Inhibition of Inflammation by Hydroxycitrate" Nanomaterials 10, no. 10: 2080. https://doi.org/10.3390/nano10102080
APA StyleVassallo, A., Santoro, V., Pappalardo, I., Santarsiero, A., Convertini, P., De Luca, M., Martelli, G., Infantino, V., & Caddeo, C. (2020). Liposome-Mediated Inhibition of Inflammation by Hydroxycitrate. Nanomaterials, 10(10), 2080. https://doi.org/10.3390/nano10102080