
Contributions of Organic and Mineral Matter to Vertical Accretion in Tidal Wetlands across a Chesapeake Bay Subestuary



Abstract
:1. Introduction
2. Materials and Methods
3. Results
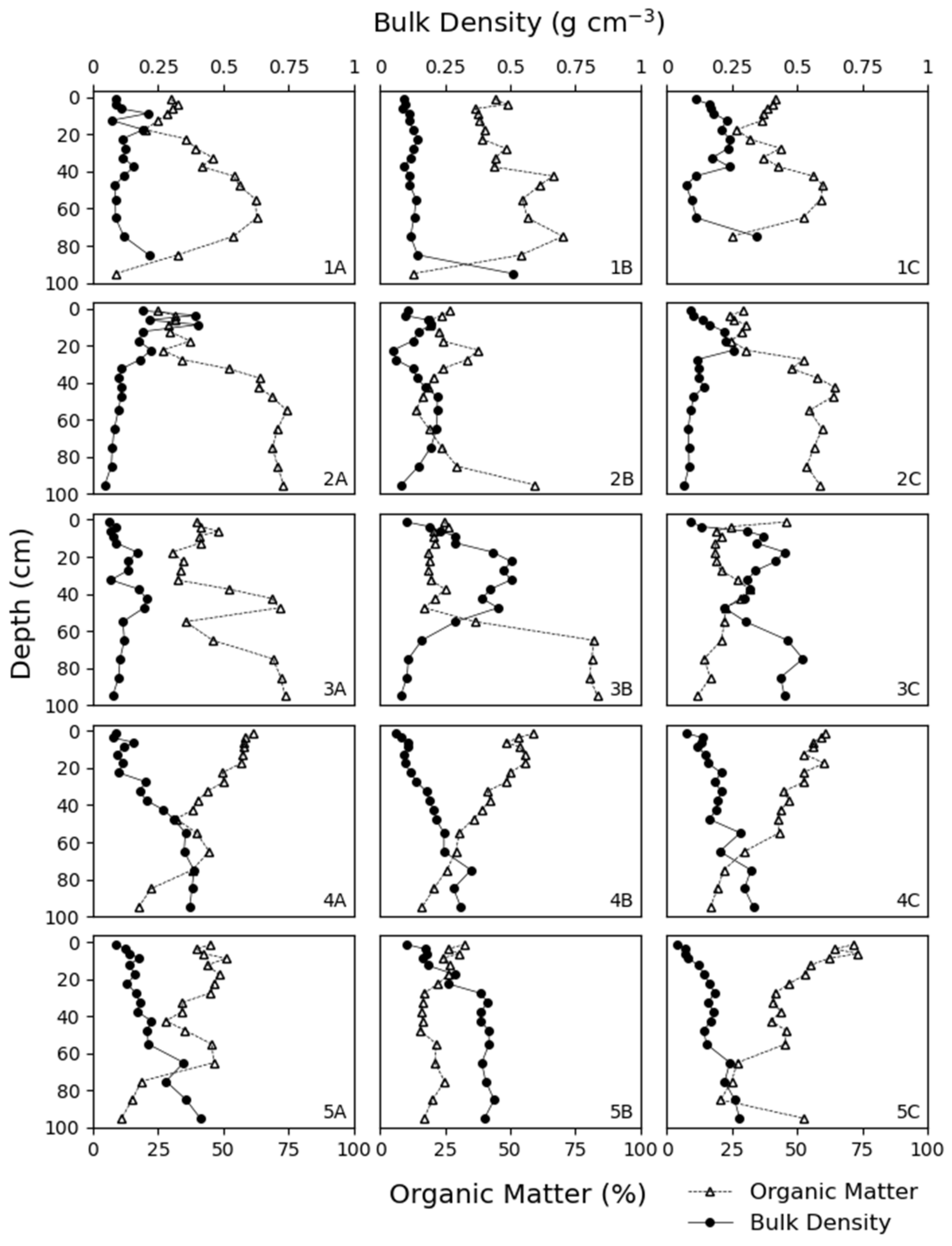
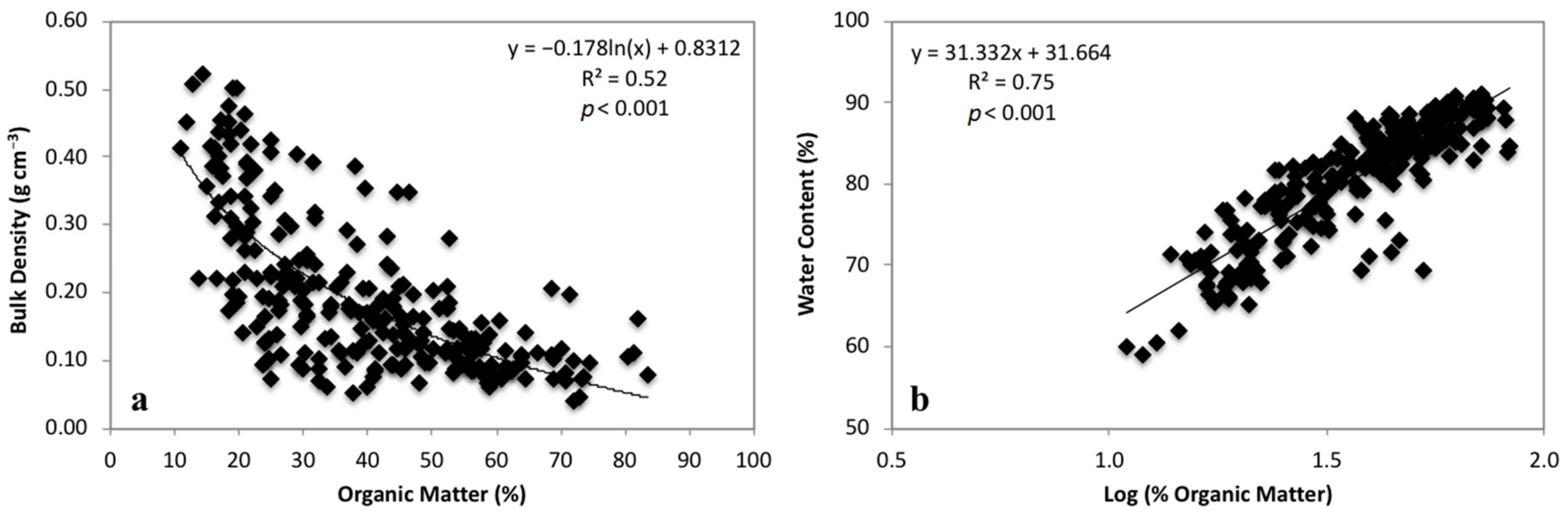
3.1. Soil Characteristics
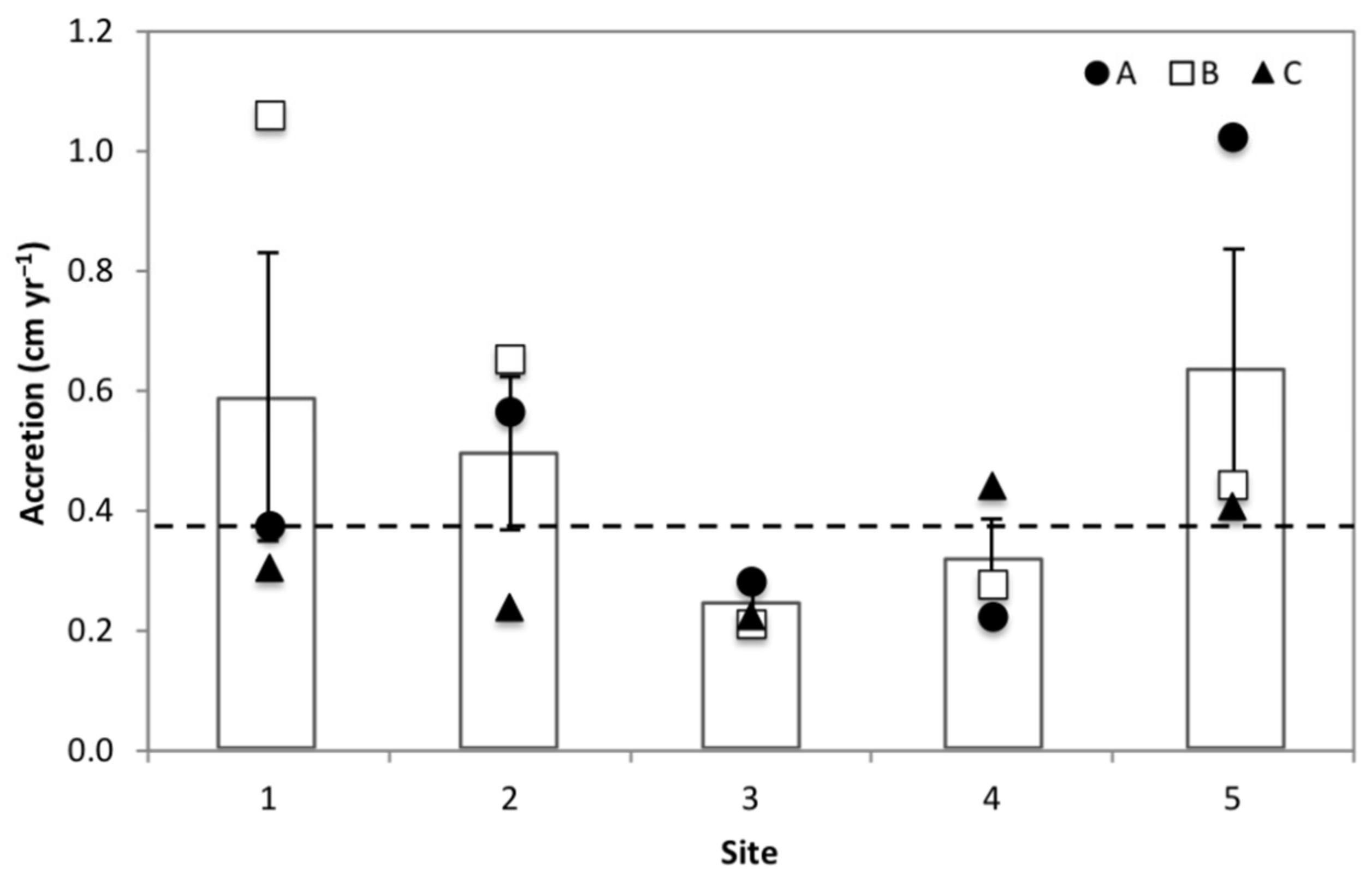
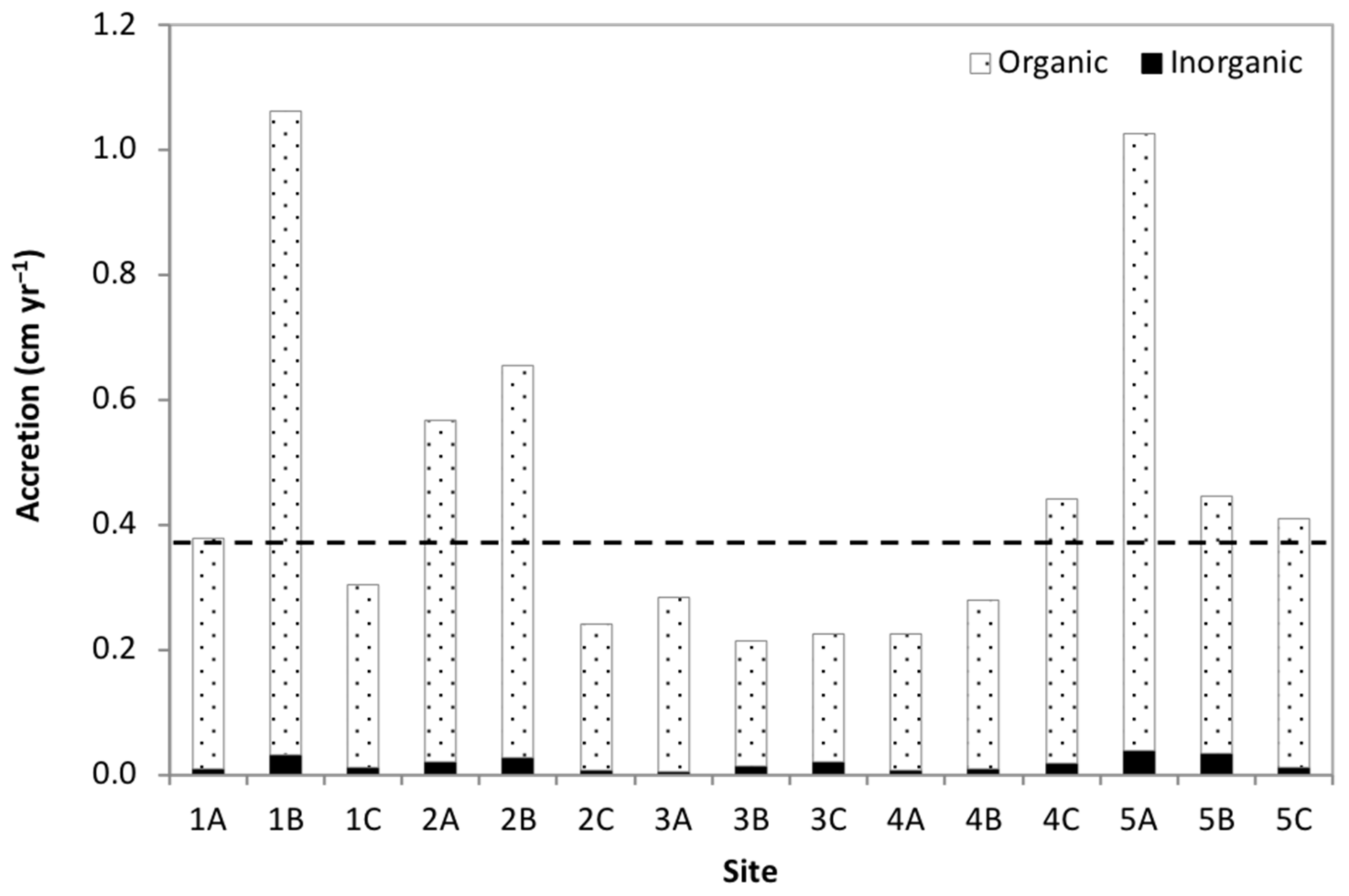
3.2. Accretion Rates
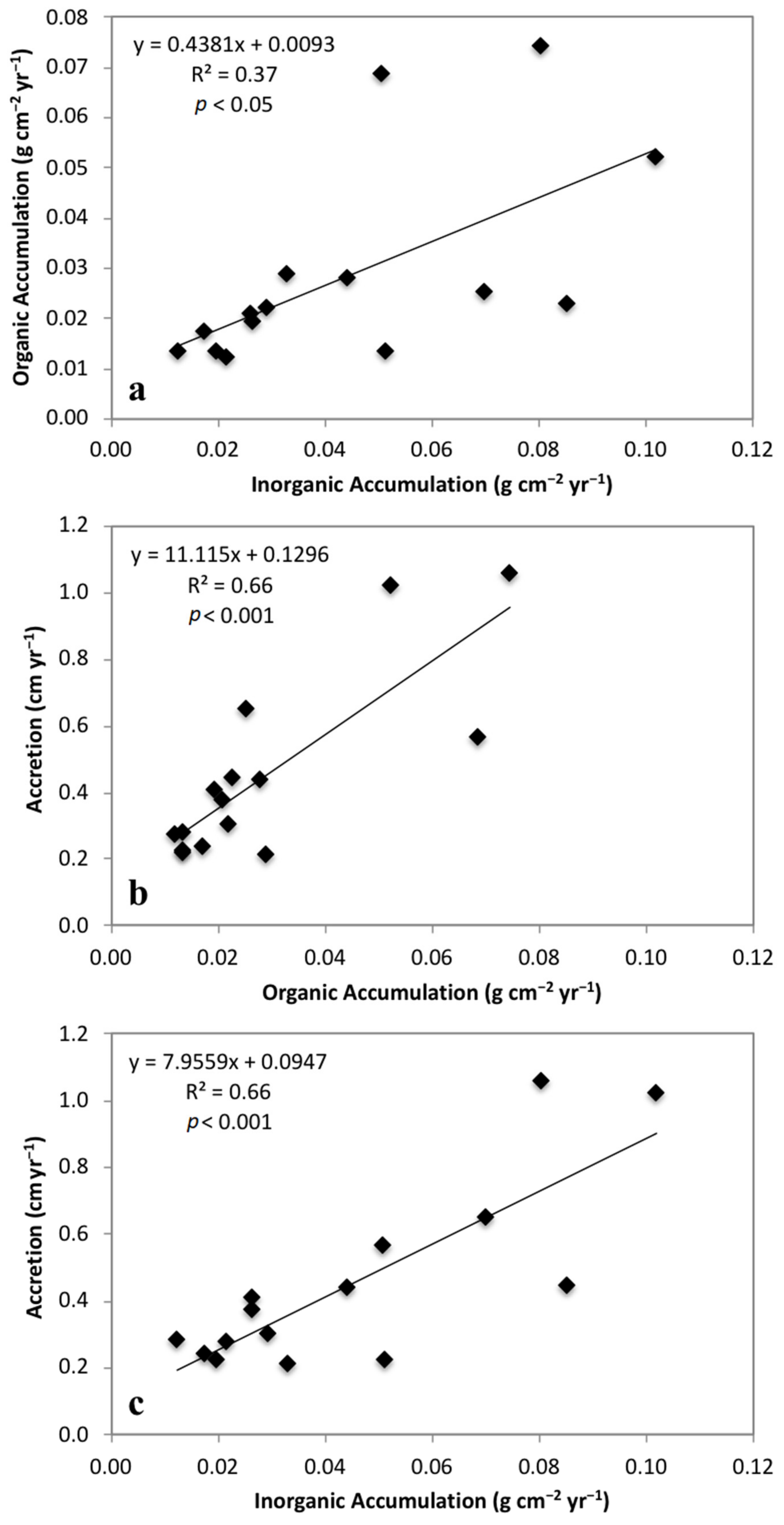
3.3. Mass Accumulation Rates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2014: Synthesis Report; Pachauri, R.K., Meyer, L., Eds.; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Hay, C.C.; Morrow, E.; Kopp, R.E.; Mitrovica, J.X. Probabilistic Reanalysis of Twentieth-Century Sea-Level Rise. Nature 2015, 517, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Nerem, R.S.; Beckley, B.D.; Fasullo, J.T.; Hamlington, B.D.; Masters, D.; Mitchum, G.T. Climate-Change-Driven Accelerated Sea-Level Rise Detected in the Altimeter Era. Proc. Natl. Acad. Sci. USA 2018, 115, 2022–2025. [Google Scholar] [CrossRef] [Green Version]
- WCRP Global Sea Level Budget Group. Global Sea-Level Budget 1993–Present. Earth Syst. Sci. Data 2018, 10, 1551–1590. [Google Scholar] [CrossRef] [Green Version]
- Sallenger, A.H.; Doran, K.S.; Howd, P.A. Hotspot of Accelerated Sea-Level Rise on the Atlantic Coast of North America. Nat. Clim. Chang. 2012, 2, 884–888. [Google Scholar] [CrossRef]
- Boon, J.D.; Mitchell, M. Nonlinear Change in Sea Level Observed at North American Tide Stations. J. Coast. Res. 2015, 31, 1295–1305. [Google Scholar] [CrossRef]
- Boon, J.D.; Brubaker, J.M.; Forrest, D.R. Chesapeake bay land subsidence and sea level change: An evaluation of past and present trends and future outlook. In Applied Marine Science and Ocean Engineering 2010; Special Report No. 425; Virginia Institute of Marine Science, William and Mary: Williamsburg, VA, USA, 2010. [Google Scholar] [CrossRef]
- Reed, D.J. The Response of Coastal Marshes to Sea-Level Rise: Survival or Submergence? Earth Surf. Process. Landf. 1995, 20, 39–48. [Google Scholar] [CrossRef]
- Morris, J.T.; Sundareshwar, P.V.; Nietch, C.T.; Kjerfve, B.; Cahoon, D.R. Responses of Coastal Wetlands to Rising Sea Level. Ecology 2002, 83, 2869–2877. [Google Scholar] [CrossRef]
- Cahoon, D.R.; Hensel, P.F.; Spencer, T.; Reed, D.J.; McKee, K.L.; Saintilan, N. Coastal Wetland Vulnerability to Relative Sea-Level Rise: Wetland Elevation Trends and Process Controls. In Wetlands and Natural Resource Management; Verhoeven, J.T.A., Beltman, B., Bobbink, R., Whigham, D.F., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 271–292. ISBN 978-3-540-33187-2. [Google Scholar]
- Neubauer, S.C. Contributions of Mineral and Organic Components to Tidal Freshwater Marsh Accretion. Estuar. Coast. Shelf Sci. 2008, 78, 78–88. [Google Scholar] [CrossRef]
- Bricker-Urso, S.; Nixon, S.W.; Cochran, J.K.; Hirschberg, D.J.; Hunt, C. Accretion and Sediment Accumulation in Rhode Island Salt Marshes. Estuaries 1989, 12, 300–317. [Google Scholar] [CrossRef]
- Morris, J.T.; Barber, D.C.; Callaway, J.C.; Chambers, R.; Hagen, S.C.; Hopkinson, C.S.; Johnson, B.J.; Megonigal, P.; Neubauer, S.C.; Troxler, T.; et al. Contributions of Organic and Inorganic Matter to Sediment Volume and Accretion in Tidal Wetlands at Steady State. Earth’s Future 2016, 4, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Neubauer, S.C.; Anderson, I.C.; Constantine, J.A.; Kuehl, S.A. Sediment Deposition and Accretion in a Mid-Atlantic (U.S.A.) Tidal Freshwater Marsh. Estuar. Coast. Shelf Sci. 2002, 54, 713–727. [Google Scholar] [CrossRef] [Green Version]
- Callaway, J.C.; Borgnis, E.L.; Turner, R.E.; Milan, C.S. Carbon Sequestration and Sediment Accretion in San Francisco Bay Tidal Wetlands. Estuaries Coasts 2012, 35, 1163–1181. [Google Scholar] [CrossRef]
- Butzeck, C.; Eschenbach, A.; Gröngröft, A.; Hansen, K.; Nolte, S.; Jensen, K. Sediment Deposition and Accretion Rates in Tidal Marshes Are Highly Variable Along Estuarine Salinity and Flooding Gradients. Estuaries Coasts 2014, 434–450. [Google Scholar] [CrossRef]
- Turner, R.E.; Swenson, E.M.; Milan, C.S. Organic and Inorganic Contributions to Vertical Accretion in Salt Marsh Sediments. In Concepts and Controversies in Tidal Marsh Ecology; Weinstein, M.P., Kreeger, D.A., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 583–595. ISBN 978-0-306-47534-4. [Google Scholar]
- Carey, J.C.; Moran, S.B.; Kelly, R.P.; Kolker, A.S.; Fulweiler, R.W. The Declining Role of Organic Matter in New England Salt Marshes. Estuaries Coasts 2017, 40, 626–639. [Google Scholar] [CrossRef]
- McCaffrey, R.J.; Thomson, J. A record of accumulation of sediment and trace metals in a Connecticut, USA salt marsh. In Advances in Geophysics; Academic Press: Cambridge, MA, USA, 1980; Volume 22, pp. 165–229. ISBN 0120188228. [Google Scholar]
- Nyman, J.A.; Walters, R.J.; Delaune, R.D.; Patrick, W.H. Marsh Vertical Accretion via Vegetative Growth. Estuar. Coast. Shelf Sci. 2006, 69, 370–380. [Google Scholar] [CrossRef]
- Weston, N.B. Declining Sediments and Rising Seas: An Unfortunate Convergence for Tidal Wetlands. Estuaries Coasts 2014, 37, 1–23. [Google Scholar] [CrossRef]
- Boyd, B.M.; Sommerfield, C.K.; Elsey-Quirk, T. Hydrogeomorphic Influences on Salt Marsh Sediment Accumulation and Accretion in Two Estuaries of the U.S. Mid-Atlantic Coast. Mar. Geol. 2017, 383, 132–145. [Google Scholar] [CrossRef] [Green Version]
- Kearney, M.S.; Ward, L.G. Accretion Rates in Brackish Marshes of a Chesapeake Bay Estuarine Tributary. Geo-Mar. Lett. 1986, 6, 41–49. [Google Scholar] [CrossRef]
- Orson, R.A.; Simpson, R.L.; Good, R.E. Rates of Sediment Accumulation in a Tidal Freshwater Marsh. J. Sediment. Res. 1990, 60, 859–869. [Google Scholar]
- Craft, C. Freshwater Input Structures Soil Properties, Vertical Accretion, and Nutrient Accumulation of Georgia and U.S. Tidal Marshes. Limnol. Oceanogr. 2007, 52, 1220–1230. [Google Scholar] [CrossRef]
- Kearney, M.S.; Turner, R.E. Microtidal Marshes: Can These Widespread and Fragile Marshes Survive Increasing Climate-Sea Level Variability and Human Action? J. Coast. Res. 2016, 319, 686–699. [Google Scholar] [CrossRef]
- Stevenson, J.C.; Ward, L.G.; Kearney, M.S. Sediment Transport and Trapping in Marsh Systems: Implications of Tidal Flux Studies. Mar. Geol. 1988, 80, 37–59. [Google Scholar] [CrossRef]
- Chesapeake Bay Foundation. Nanticoke River Watershed: Natural and Cultural Resources Atlas; Chesapeake Bay Foundation: Annapolis, MD, USA, 1996. [Google Scholar]
- Kearney, M.S.; Grace, R.E.; Stevenson, J.C. Marsh Loss in Nanticoke Estuary, Chesapeake Bay. Geogr. Rev. 1988, 78, 205–220. [Google Scholar] [CrossRef]
- Beckett, L.H.; Baldwin, A.H.; Kearney, M.S. Tidal Marshes across a Chesapeake Bay Subestuary Are Not Keeping up with Sea-Level Rise. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.D.; Bleil, D.F. Condition of Nontidal Wetlands in the Nanticoke River Watershed, Maryland and Delaware; Delaware Department of Natural Resources and Environmental Control, Watershed Assessment Section: Dover, DE, USA, 2008. [Google Scholar]
- The Nature Conservancy. Nanticoke River Bioreserve Strategic Plan; The Nature Conservancy: Bethesda, MD, USA, 1998. [Google Scholar]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H. Methods of Soil Analysis Part 3—Chemical Methods; SSSA Book Series; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996; ISBN 978-0-89118-866-7. [Google Scholar]
- Armentano, T.V.; Woodwell, G.M. Sedimentation Rates in a Long Island Marsh Determined by 210Pb Dating. Limnol. Oceanogr. 1975, 20, 452–456. [Google Scholar] [CrossRef]
- Flynn, W.W. The Determination of Low Levels of Polonium-210 in Environmental Materials. Anal. Chim. Acta 1968, 43, 221–227. [Google Scholar] [CrossRef]
- Robbins, J.A.; Edgington, D.N.; Kemp, A.L.W. Comparative 210Pb,137Cs, and Pollen Geochronologies of Sediments from Lakes Ontario and Erie. Quat. Res. 1978, 10, 256–278. [Google Scholar] [CrossRef] [Green Version]
- DeLaune, R.D.; Baumann, R.H.; Gosselink, J.G. Relationships among Vertical Accretion, Coastal Submergence, and Erosion in a Louisiana Gulf Coast Marsh. J. Sediment. Petrol. 1983, 53, 147–157. [Google Scholar] [CrossRef]
- Noe, G.B.; Hupp, C.R.; Bernhardt, C.E.; Krauss, K.W. Contemporary Deposition and Long-Term Accumulation of Sediment and Nutrients by Tidal Freshwater Forested Wetlands Impacted by Sea Level Rise. Estuaries Coasts 2016, 39, 1006–1019. [Google Scholar] [CrossRef]
- Temmerman, S.; Bouma, T.J.; Govers, G.; Lauwaet, D. Flow Paths of Water and Sediment in a Tidal Marsh: Relations with Marsh Developmental Stage and Tidal Inundation Height. Estuaries 2005, 28, 338–352. [Google Scholar] [CrossRef]
- Fagherazzi, S.; Kirwan, M.L.; Mudd, S.M.; Guntenspergen, G.R.; Temmerman, S.; D’Alpaos, A.; van de Koppel, J.; Rybczyk, J.M.; Reyes, E.; Craft, C.; et al. Numerical Models of Salt Marsh Evolution: Ecological, Geomorphic, and Climatic Factors. Rev. Geophys. 2012, 50, RG1002. [Google Scholar] [CrossRef]
- Gleason, M.L.; Elmer, D.A.; Pien, N.C.; Fisher, J.S. Effects of Stem Density upon Sediment Retention by Salt Marsh Cord Grass, Spartina Alterniflora Loisel. Estuaries 1979, 2, 271. [Google Scholar] [CrossRef]
- DeLaune, R.D.; Reddy, C.N.; Patrick, W.H. Accumulation Metals through Accretion of Plant Nutrients Sedimentation and Heavy Processes and in a Louisiana. Estuaries 1981, 4, 328–334. [Google Scholar] [CrossRef]
- Barbier, E.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The Value of Estuarine and Coastal Ecosystem Services. Ecol. Monogr. 2011, 81, 43. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Salinity Regime | Average Salinity (ppt) | Distance from Mouth (km) | Dominant Vegetation |
|---|---|---|---|---|
| 1 | Mesohaline | 10 | 12 | Juncus roemerianus, Spartina alterniflora, Distichlis spicata, Spartina patens, Bolboschoenus robustus |
| 2 | Mesohaline | 5 | 25 | Spartina alterniflora, Spartina patens, Spartina cynosuroides, Distichlis spicata, Bolboschoenus robustus |
| 3 | Oligohaline | 2 | 40 | Peltandra virginica, Mikania scandens, Bidens laevis, Typha sp., Polygonum punctatum |
| 4 | Oligohaline | 1 | 50 | Polygonum arifolium, Impatiens capensis, Bolboschoenus fluviatilis, Peltandra virginica, Pilea pumila |
| 5 | Tidal Freshwater | 0.1 | 60 | Peltandra virginica, Polygonum arifolium, Zizania aquatica, Impatiens capensis, Nuphar lutea |
| Sub- Site | Accretion (cm/yr) | Bulk Density (g cm−3) | Organic Matter (%) | Accumulation Rates (g cm−2 y−1) | Percent of Accretion | |||
|---|---|---|---|---|---|---|---|---|
| Org | Inorg | Org | Inorg | W/P | ||||
| 1A | 0.3772 | 0.1242 | 44.49 | 0.0210 | 0.0262 | 5.07 | 2.68 | 92.26 |
| 1B | 1.0611 | 0.1618 | 48.11 | 0.0745 | 0.0803 | 6.38 | 2.91 | 90.71 |
| 1C | 0.3045 | 0.1835 | 42.95 | 0.0221 | 0.0293 | 6.59 | 3.70 | 89.71 |
| 2A | 0.5674 | 0.1265 | 57.49 | 0.0686 | 0.0507 | 11.00 | 3.44 | 85.57 |
| 2B | 0.6537 | 0.1539 | 26.59 | 0.0253 | 0.0699 | 3.52 | 4.12 | 92.36 |
| 2C | 0.2397 | 0.1195 | 49.62 | 0.0172 | 0.0175 | 6.53 | 2.81 | 90.66 |
| 3A | 0.2833 | 0.1170 | 52.07 | 0.0134 | 0.0124 | 4.31 | 1.68 | 94.01 |
| 3B | 0.2133 | 0.2683 | 46.67 | 0.0289 | 0.0330 | 12.32 | 5.96 | 81.72 |
| 3C | 0.2240 | 0.3753 | 20.68 | 0.0134 | 0.0512 | 5.42 | 8.80 | 85.78 |
| 4A | 0.2259 | 0.2687 | 40.51 | 0.0135 | 0.0198 | 5.44 | 3.38 | 91.18 |
| 4B | 0.2781 | 0.2142 | 35.99 | 0.0121 | 0.0215 | 3.95 | 2.97 | 93.08 |
| 4C | 0.4415 | 0.2285 | 38.73 | 0.0279 | 0.0441 | 5.74 | 3.84 | 90.43 |
| 5A | 1.0252 | 0.2431 | 33.86 | 0.0521 | 0.1019 | 4.62 | 3.82 | 91.56 |
| 5B | 0.4445 | 0.3571 | 21.10 | 0.0228 | 0.0852 | 4.66 | 7.37 | 87.97 |
| 5C | 0.4081 | 0.1857 | 42.26 | 0.0193 | 0.0264 | 4.31 | 2.49 | 93.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allen, J.R.; Cornwell, J.C.; Baldwin, A.H. Contributions of Organic and Mineral Matter to Vertical Accretion in Tidal Wetlands across a Chesapeake Bay Subestuary. J. Mar. Sci. Eng. 2021, 9, 751. https://doi.org/10.3390/jmse9070751
Allen JR, Cornwell JC, Baldwin AH. Contributions of Organic and Mineral Matter to Vertical Accretion in Tidal Wetlands across a Chesapeake Bay Subestuary. Journal of Marine Science and Engineering. 2021; 9(7):751. https://doi.org/10.3390/jmse9070751
Chicago/Turabian StyleAllen, Jenny R., Jeffrey C. Cornwell, and Andrew H. Baldwin. 2021. "Contributions of Organic and Mineral Matter to Vertical Accretion in Tidal Wetlands across a Chesapeake Bay Subestuary" Journal of Marine Science and Engineering 9, no. 7: 751. https://doi.org/10.3390/jmse9070751
APA StyleAllen, J. R., Cornwell, J. C., & Baldwin, A. H. (2021). Contributions of Organic and Mineral Matter to Vertical Accretion in Tidal Wetlands across a Chesapeake Bay Subestuary. Journal of Marine Science and Engineering, 9(7), 751. https://doi.org/10.3390/jmse9070751