
Nitrogen Fixation in Subtropical Seagrass Sediments: Seasonal Patterns in Activity in Santa Rosa Sound, Florida, USA


Abstract
:1. Introduction
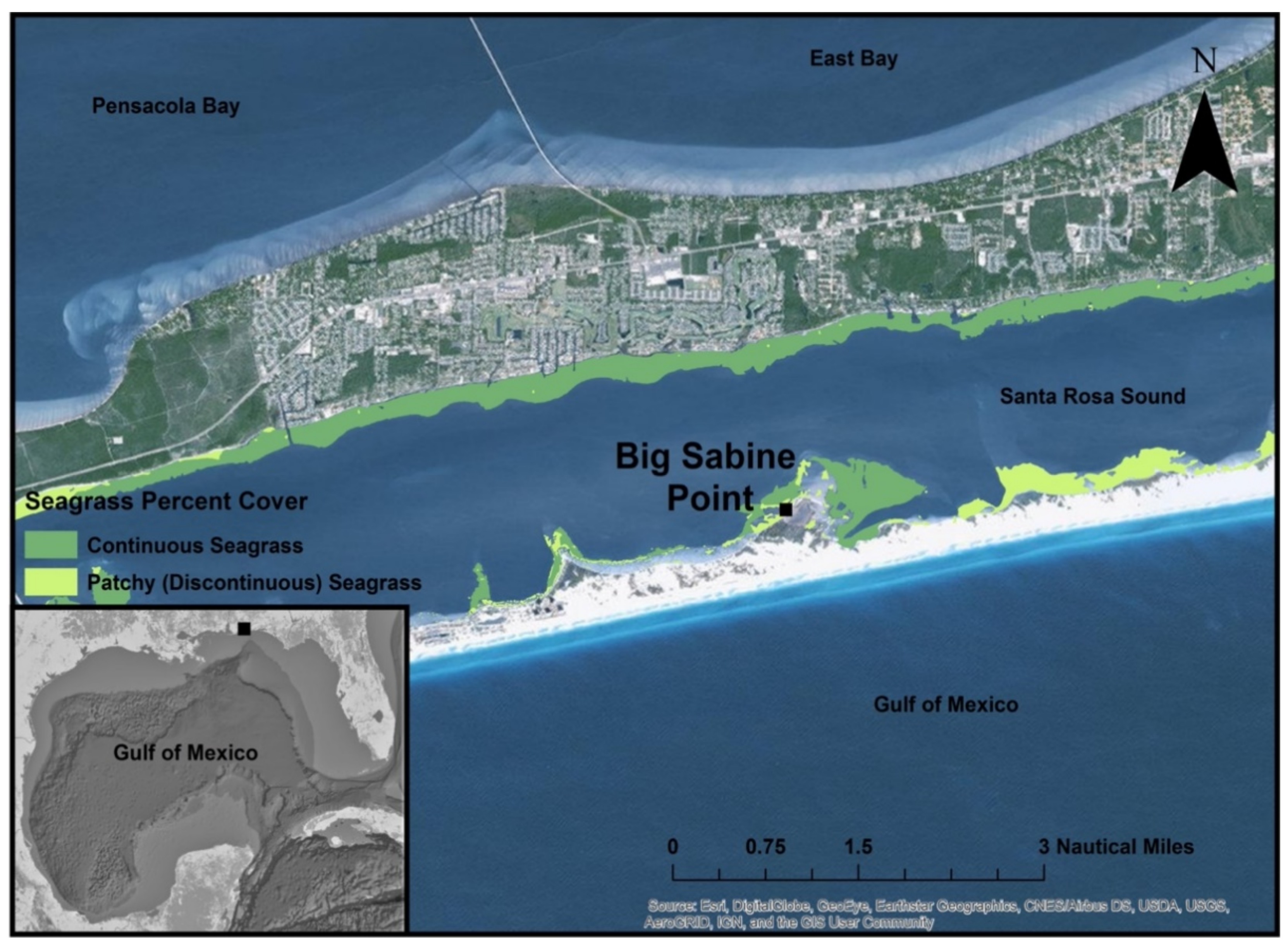
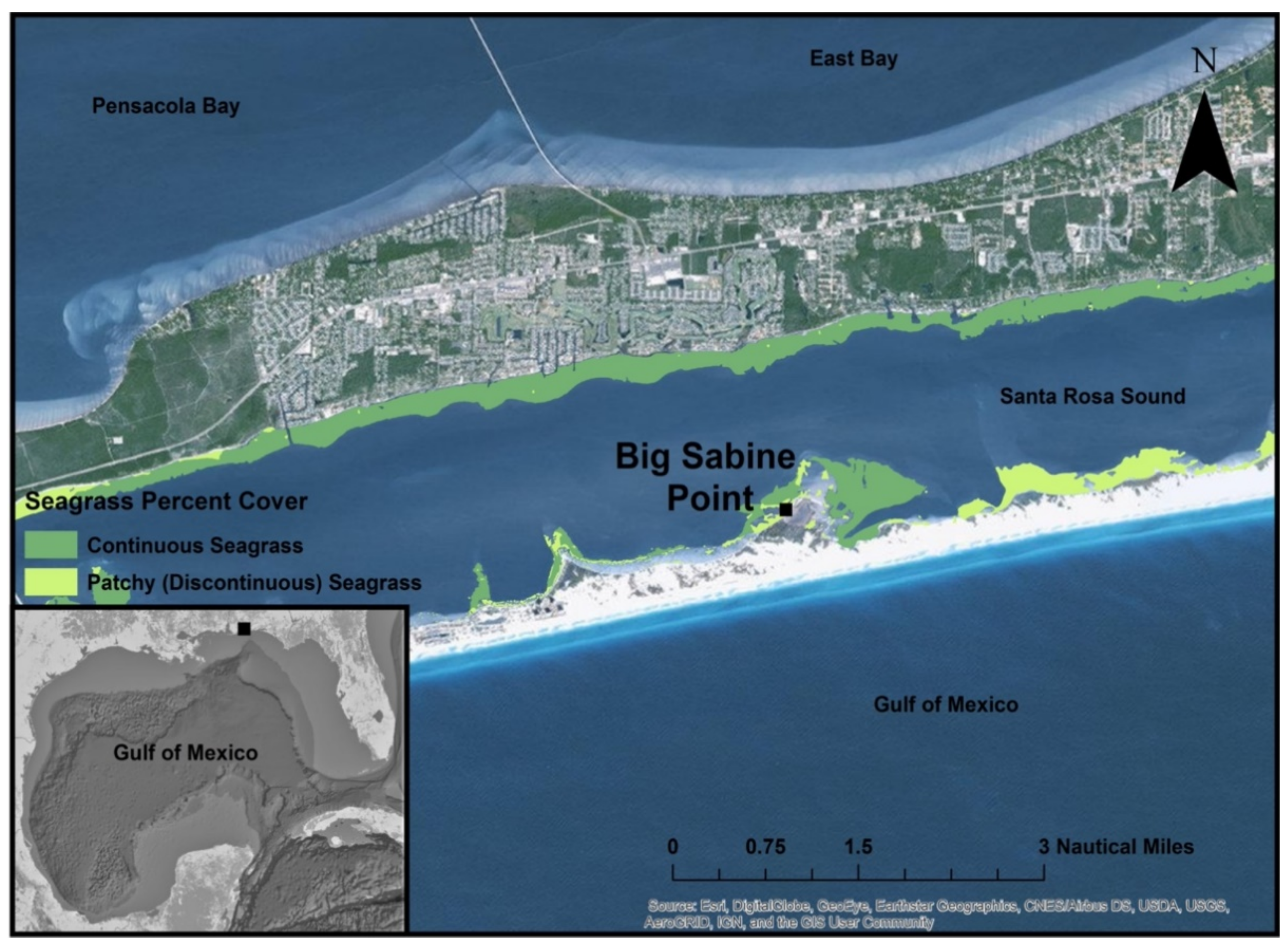
2. Materials and Methods
3. Results
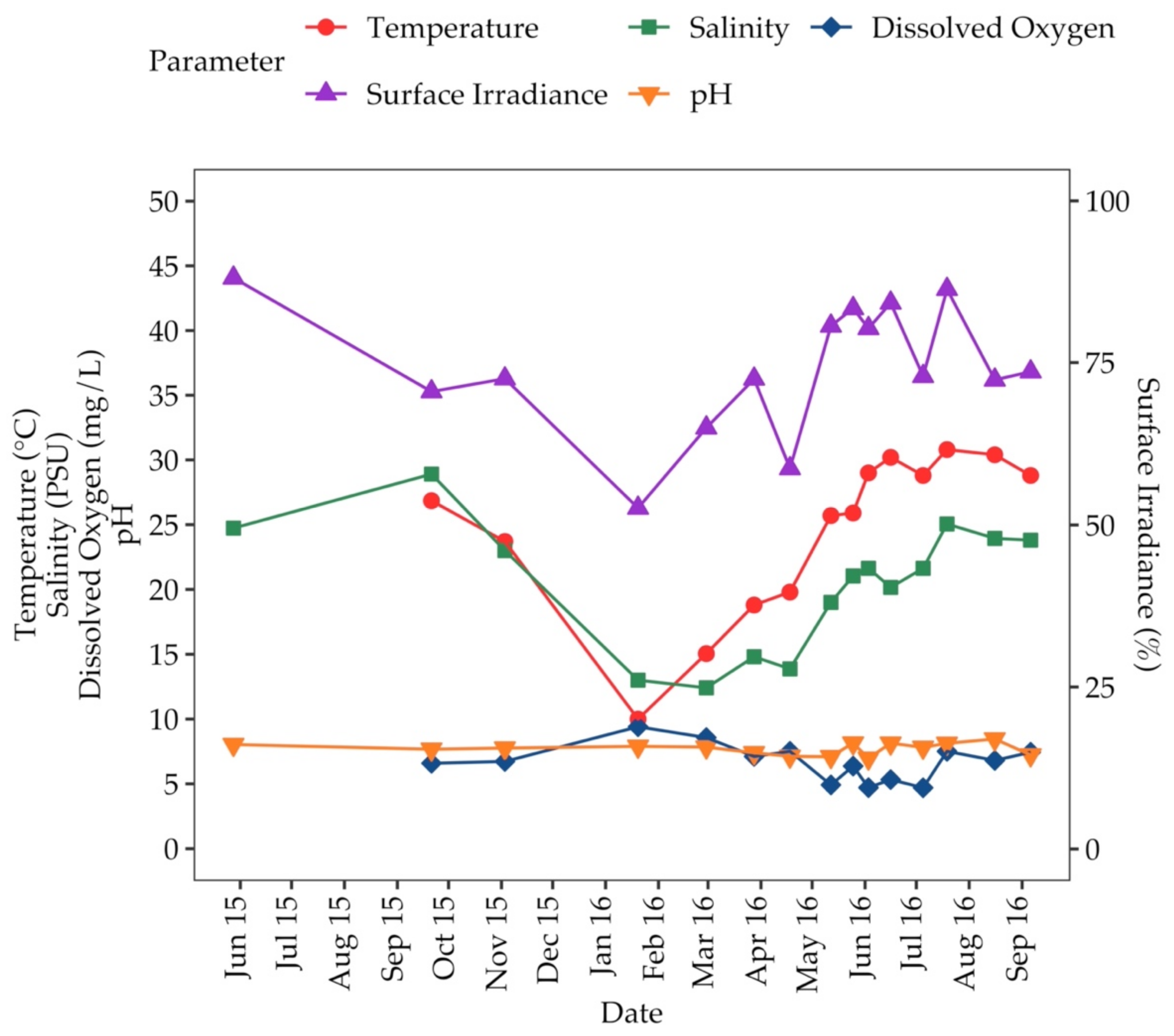
3.1. Environmental Parameters
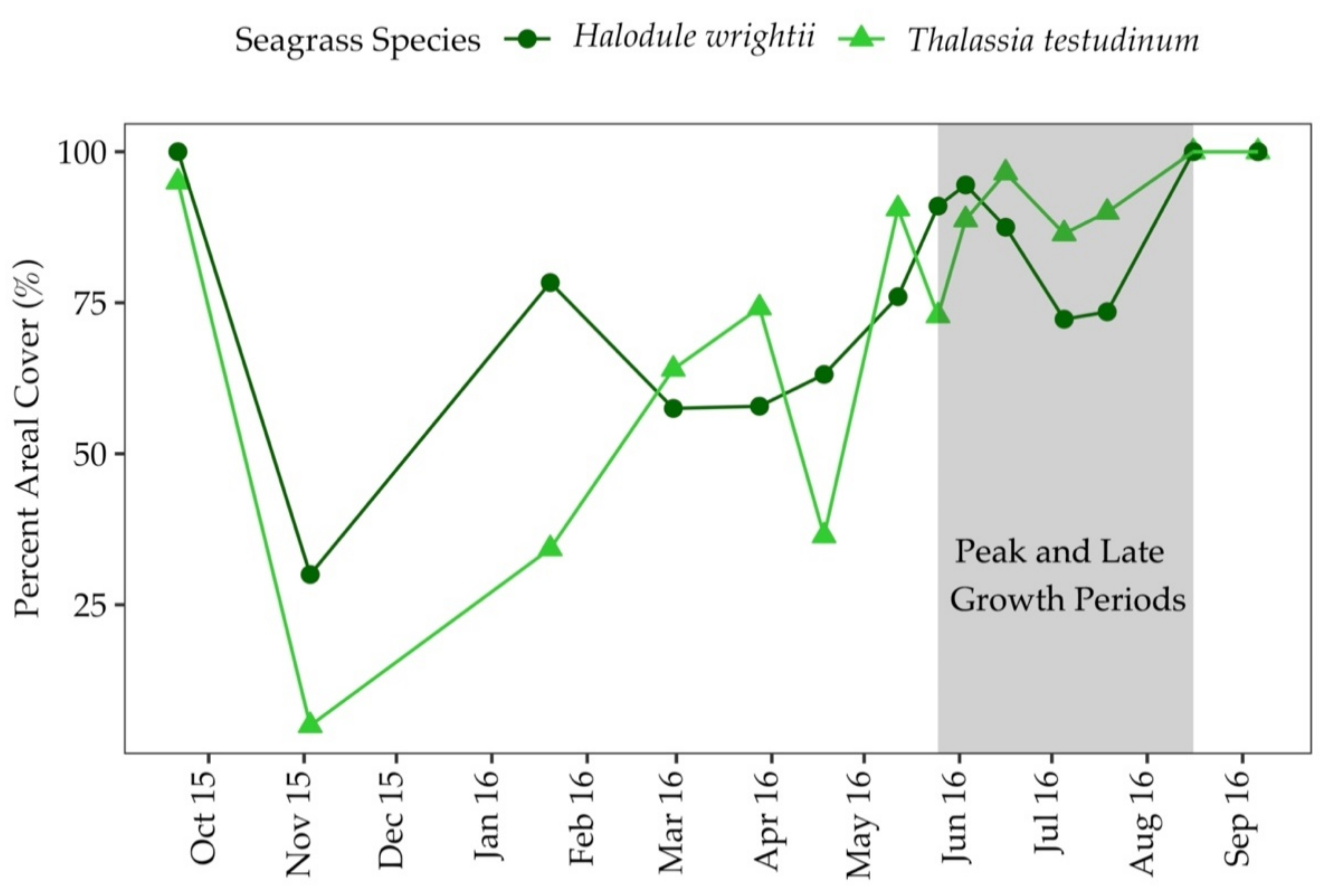
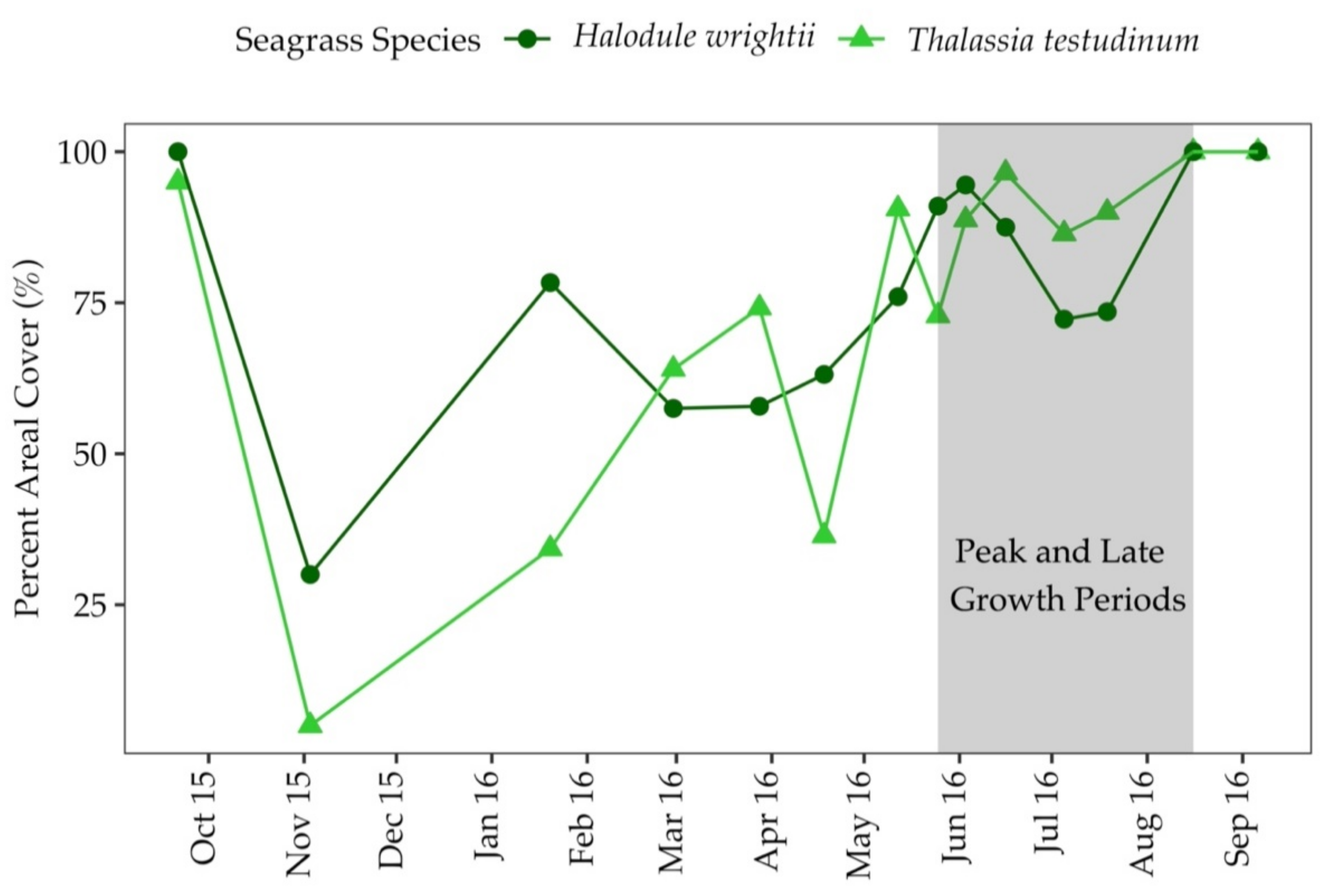
3.2. Percent Areal Cover and Sediment Characteristics
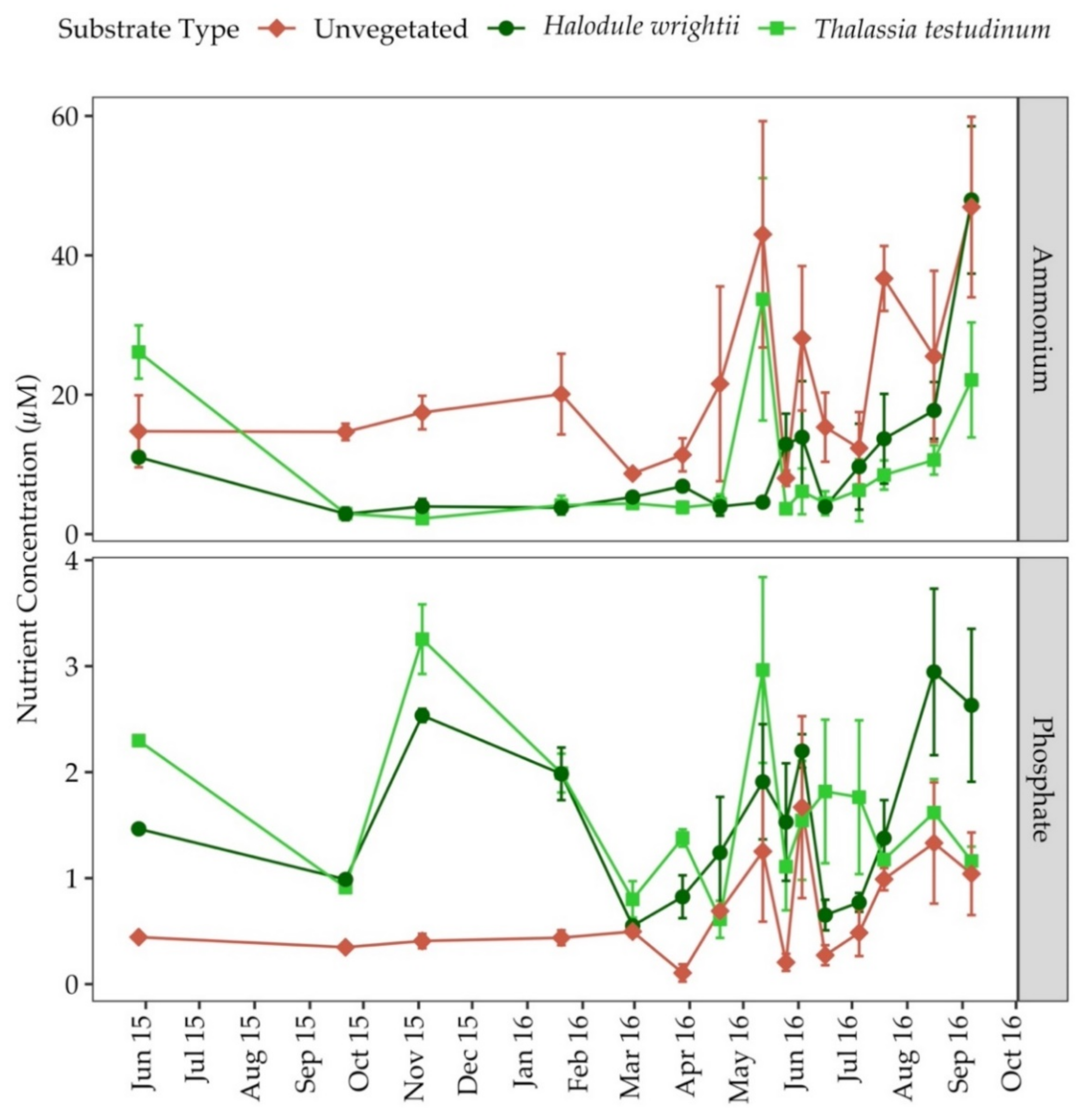
3.3. Water Column Chlorophyll a and Nutrients
3.4. Porewater Nutrients
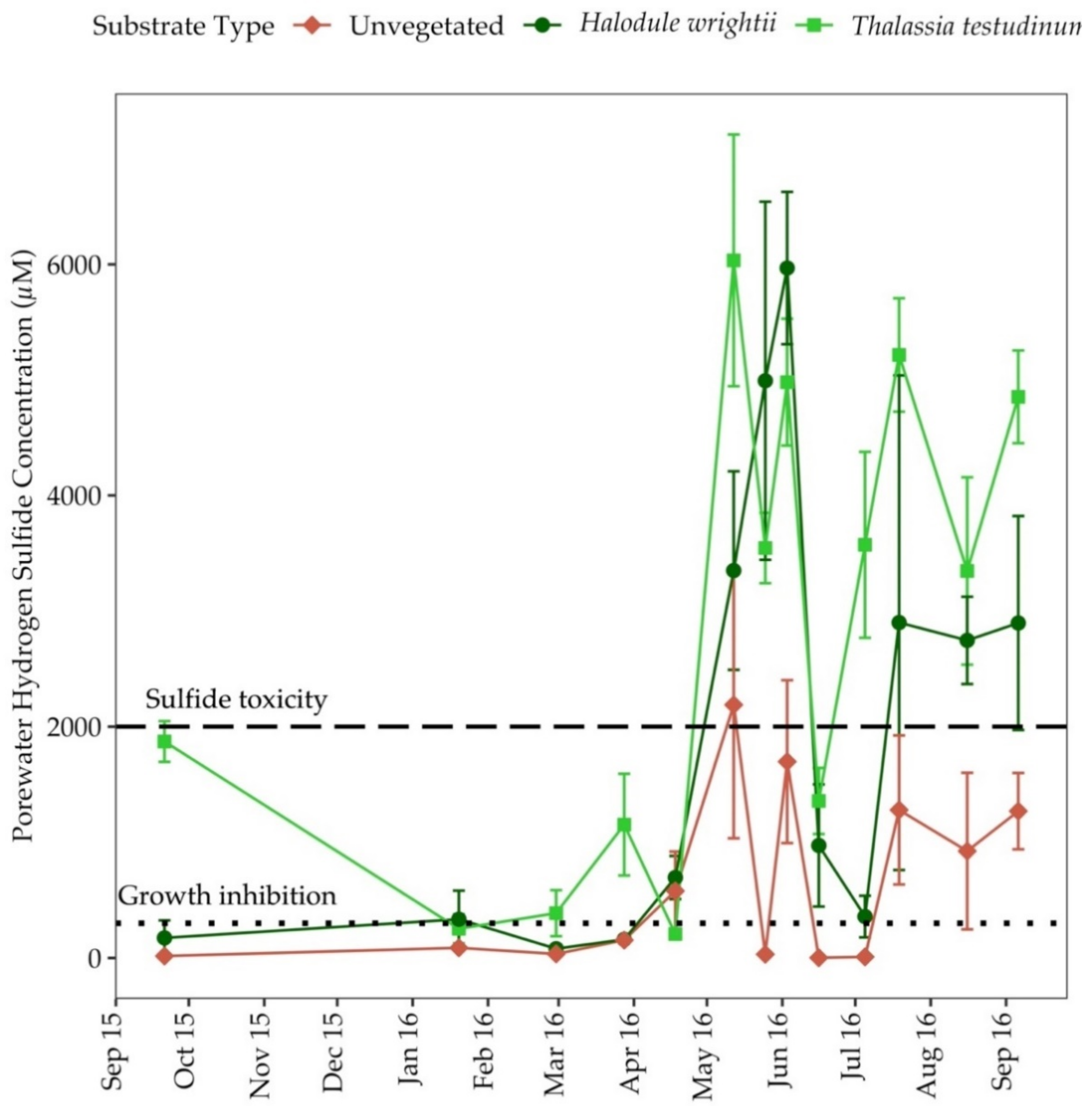
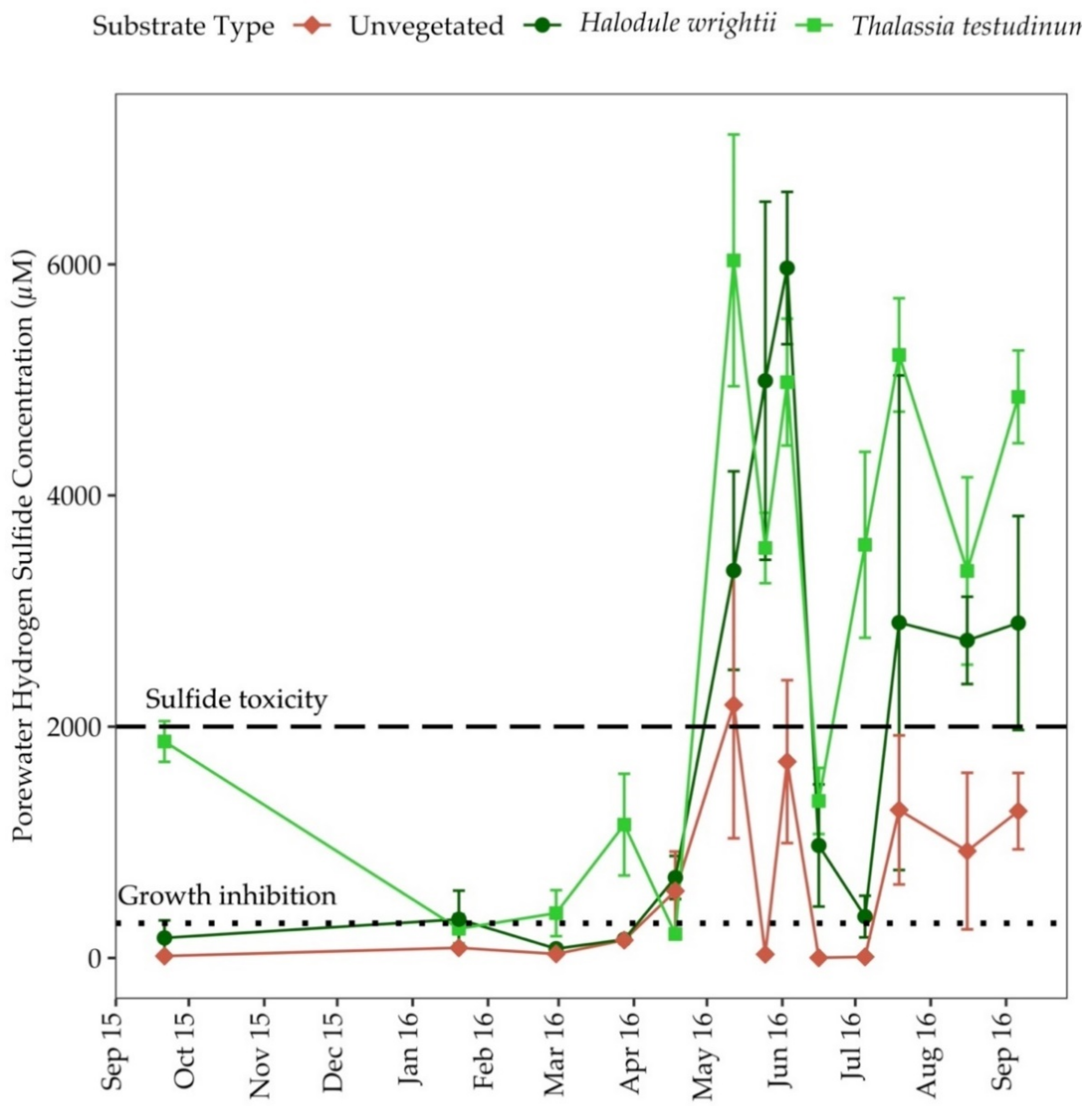
3.5. Porewater Sulfide (H2S)
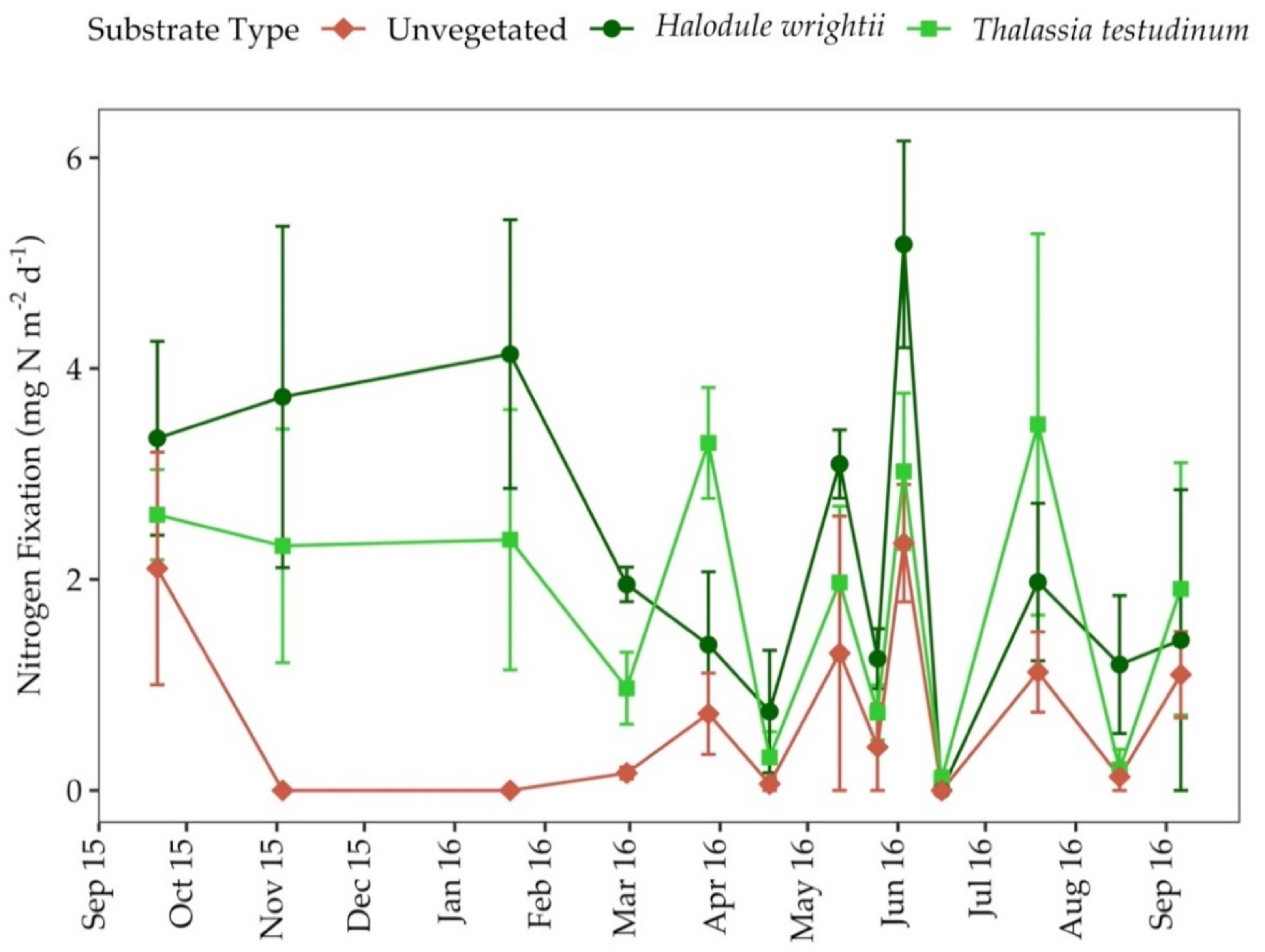
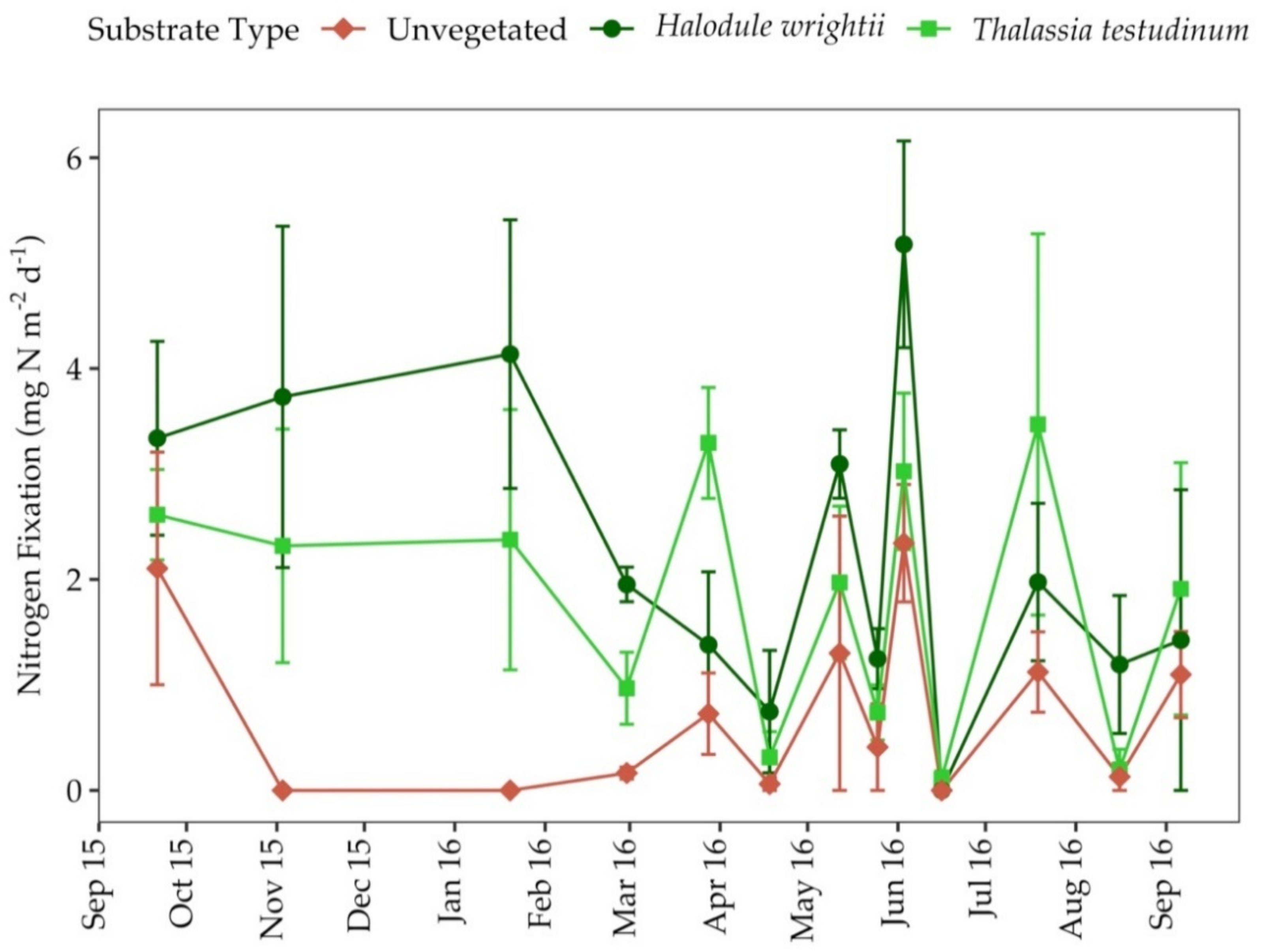
3.6. Nitrogen Fixation Rates
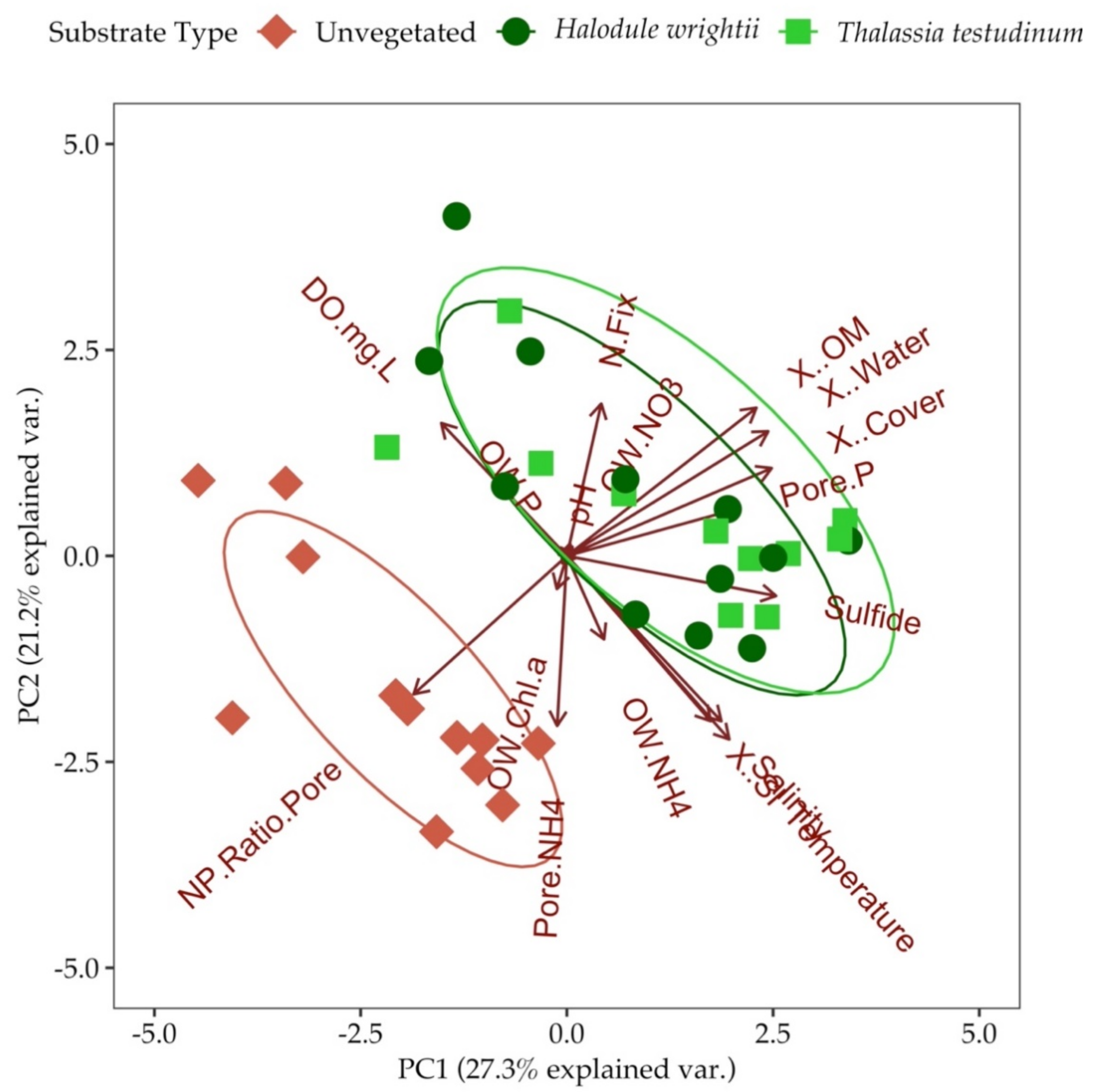
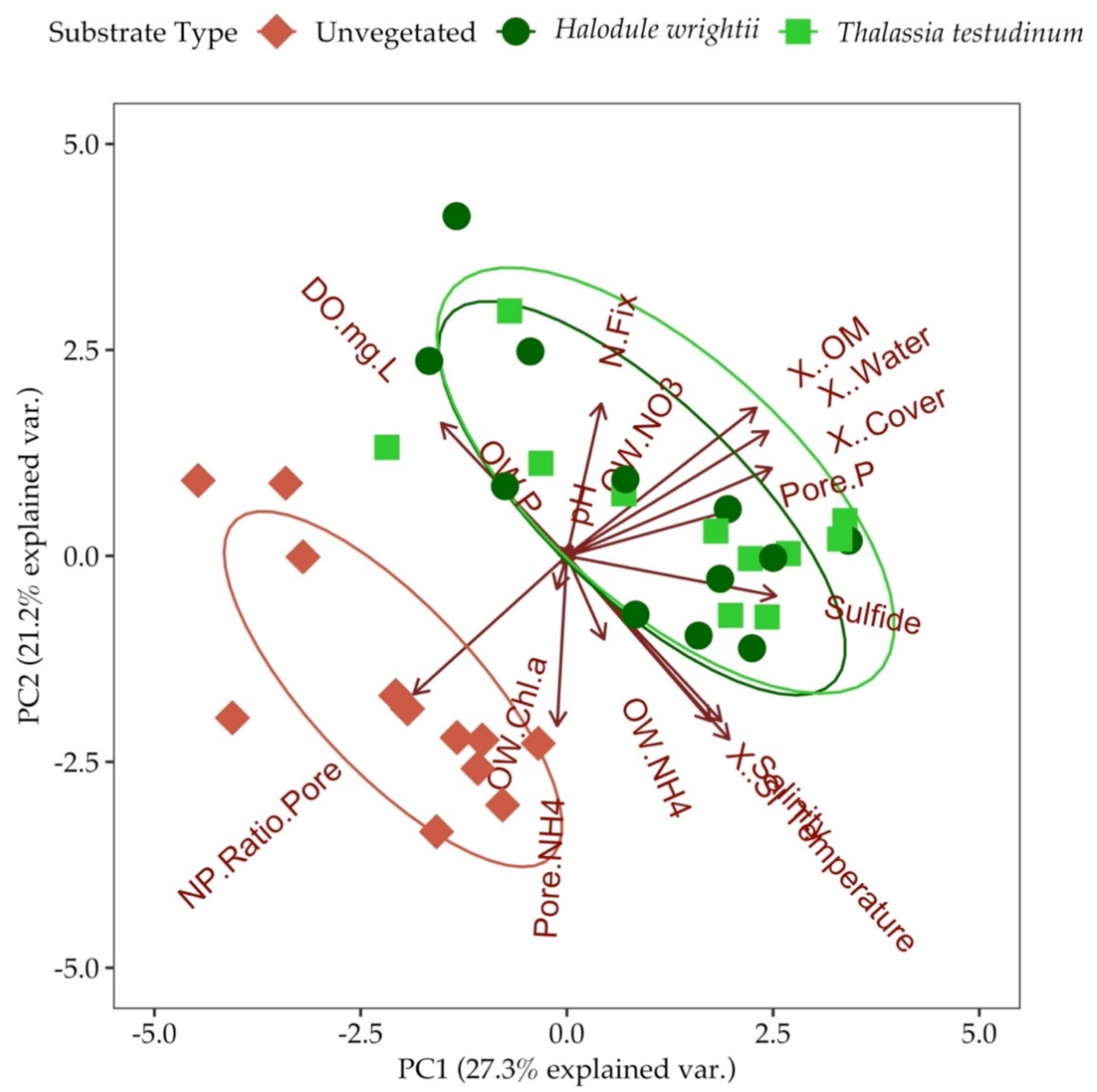
3.7. Principal Component Analysis (PCA) and Analysis of Similarities (ANOSIM)
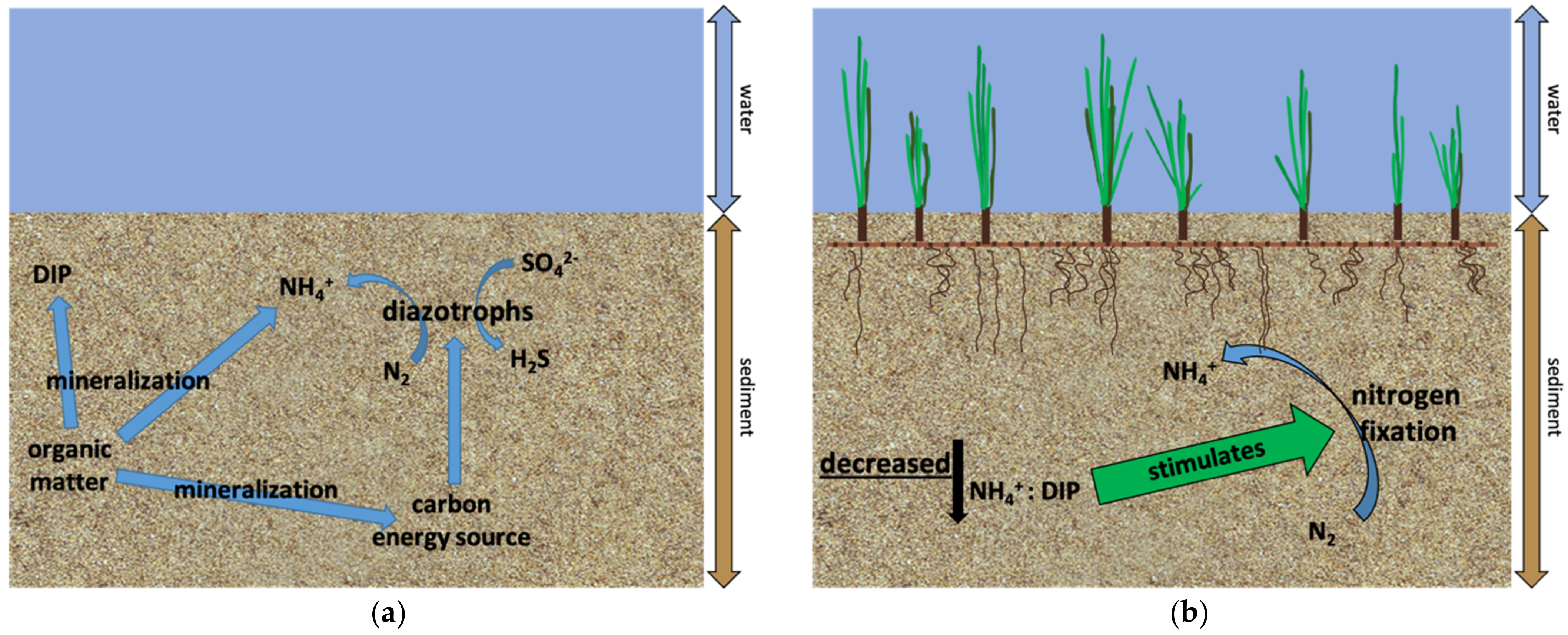
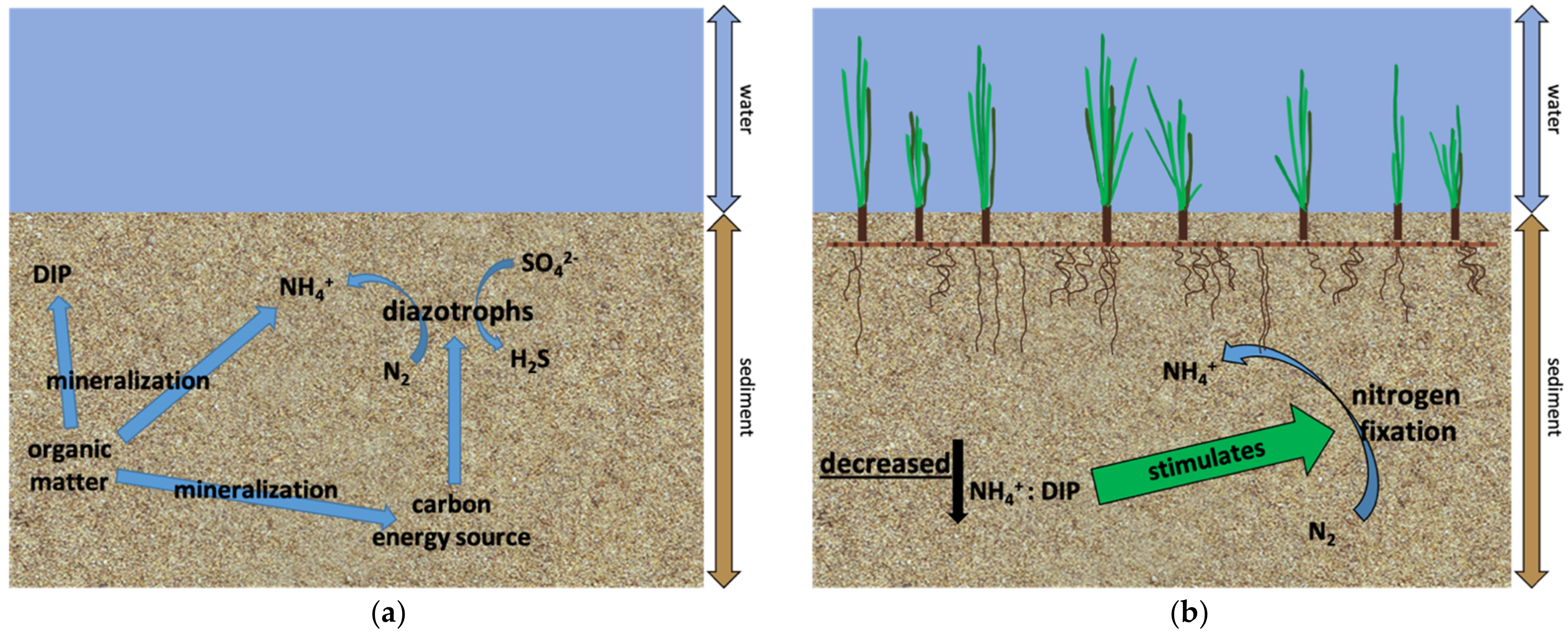
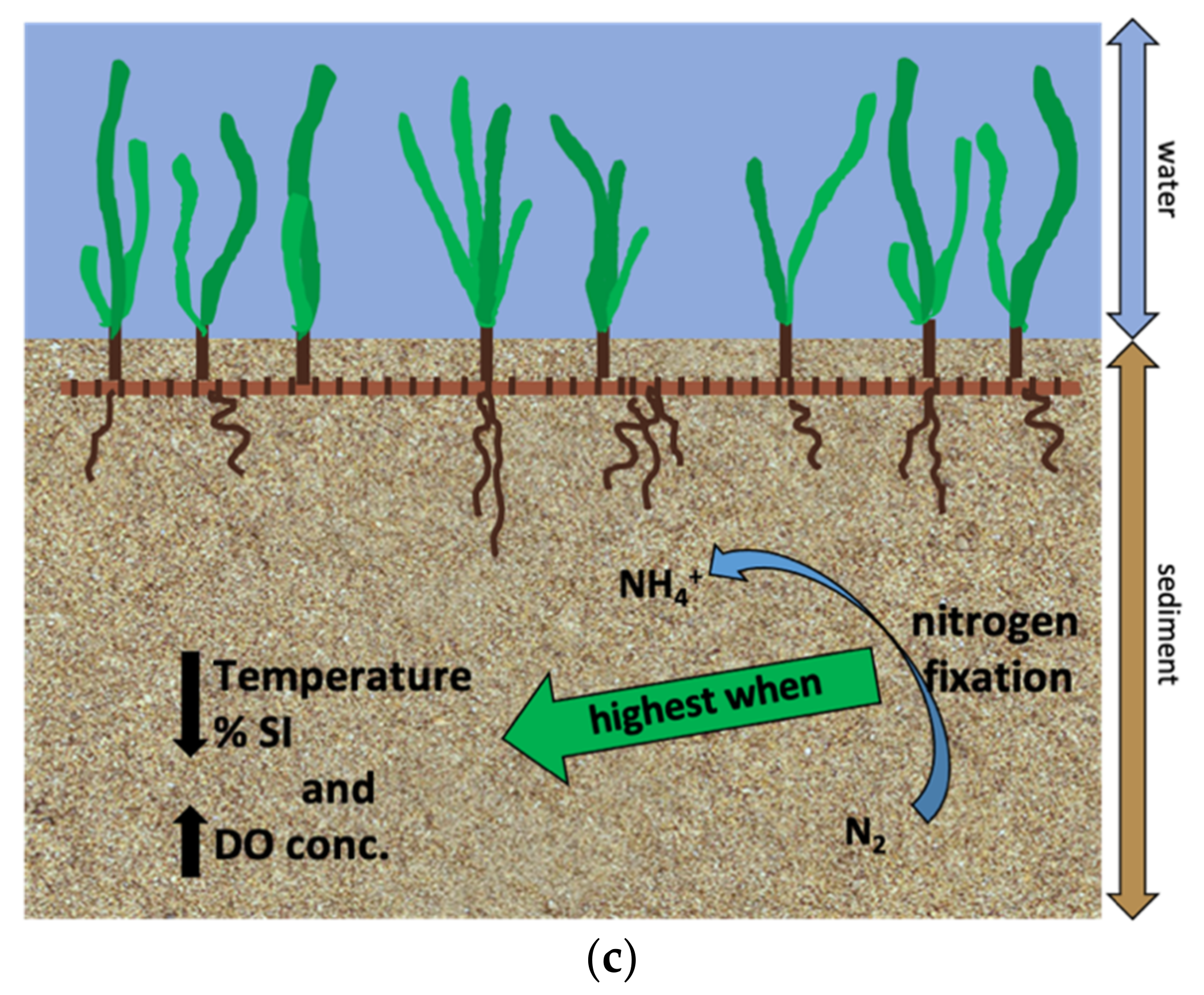
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thayer, G.W.; Wolfe, D.A.; Williams, R.B. The impact of man on seagrass systems. Am. Sci. 1975, 63, 288–296. [Google Scholar]
- Short, F.T.; Wylle-Echeverria, S. Natural and human-induced disturbances of seagrasses. Environ. Conserv. 1996, 23, 17–27. [Google Scholar] [CrossRef]
- Hemminga, M.A.; Duarte, C.M. Seagrass Ecology; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Orth, R.J.; Carruthers, J.B.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L., Jr.; Hughes, A.R.; Kendrick, G.A.; Kenworthy, W.J.; Olyarnik, S.; et al. A global crisis for seagrass ecosystems. Bioscience 2006, 56, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Lamb, J.B.; Van De Water, J.A.J.M.; Bourne, D.G.; Altier, C.; Hein, M.Y.; Fiorenza, E.A.; Abu, N.; Jompa, J.; Harvell, C.D. Seagrass ecosystems reduce exposure to bacterial pathogens of humans, fishes, and invertebrates. Science 2017, 355, 731–733. [Google Scholar] [CrossRef]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.B.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L., Jr.; Hughes, A.R.; et al. Accelerating loss of seagrass across the globe threatens coastal ecosystems. PNAS 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iverson, R.L.; Bittaker, H.F. Seagrass distribution and abundance in Eastern Gulf of Mexico Coastal Waters. Estuar. Coast. Shelf Sci. 1986, 22, 577–602. [Google Scholar] [CrossRef]
- Devereaux, R. Seagrass Rhizosphere Microbial Communities. In Interactions between Macro- and Microorganisms in Marine Sediments; Kristensen, E., Haese, R.F., Kostka, J.E., Eds.; AGU: Washington, DC, USA, 2005; Volume 60, pp. 199–216. [Google Scholar]
- Greve, T.M.; Binzer, T. Which factors regulate seagrass growth and distribution? In European Seagrass: An Introduction to Monitoring and Management; Borum, J., Duarte, C.M., Krause-Jensen, D., Greve, T.M., Eds.; EU Project Monitoring and Managing of European Seagrass (M&MS): Copenhagen, Denmark, 2004; pp. 19–23. [Google Scholar]
- Welsh, D.T. Nitrogen fixation in seagrass meadows: Regulation, plant-bacteria interactions and significance to primary production. Limnol. Oceanogr. 2000, 3, 58–71. [Google Scholar] [CrossRef]
- Bray, C.M. Nitrogen Metabolism in Plants; Longman Inc.: New York, NY, USA, 1983. [Google Scholar]
- Müller, A.; Newton, W.E. Nitrogen Fixation: The Chemical-Biochemical-Genetic Interface; Plenum Press: New York, NY, USA, 1983. [Google Scholar]
- Howard, J.B.; Rees, D.C. Structural Basis of Biological Nitrogen Fixation. Chem. Rev. 1996, 96, 2965–2982. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, R.C.; Smith, R.D.; Alberte, R.S. Is growth of eelgrass nitrogen limited? A numerical simulation of the effects of light and nutrients on the growth dynamics of Zostera marina. Mar. Ecol. Prog. Ser. 1987, 41, 167–176. [Google Scholar] [CrossRef]
- Lee, K.S.; Dunton, K.H. Inorganic nitrogen acquisition in the seagrass Thalassia testudinum: Development of a whole-plant nitrogen budget. Limnol. Oceanogr. 1999, 44, 1204–1215. [Google Scholar] [CrossRef] [Green Version]
- O’Donohue, M.J.; Moriarty, D.J.W.; Rae, I.C.M. Nitrogen fixation in sediments and the rhizosphere of the seagrass Zostera capricorni. Microb. Ecol. 1991, 22, 53–64. [Google Scholar] [CrossRef]
- Welsh, D.T.; Bourguès, S.; De Wit, R.; Herbert, R.A. Seasonal variations in nitrogen-fixation (acetylene reduction) and sulphate-reduction rates in the rhizosphere of Zostera noltii: Nitrogen fixation by sulphate-reducing bacteria. Mar. Biol. 1996, 125, 619–628. [Google Scholar] [CrossRef]
- McGlathery, K.J.; Risgaard-Petersen, N.; Christensen, P.B. Temporal and spatial variation in nitrogen fixation activity in the eelgrass Zostera marina rhizosphere. Mar. Ecol. Prog. Ser. 1998, 168, 245–258. [Google Scholar] [CrossRef]
- Capone, D.G.; Taylor, B.F. N2 fixation in the rhizosphere of Thalassia testudinum. Can. J. Microbiol. 1980, 26, 998–1005. [Google Scholar] [CrossRef]
- Capone, D.G. Nitrogen fixation (acetylene reduction) by rhizosphere sediments of the eelgrass Zostera marina. Mar. Ecol. Prog. Ser. 1982, 10, 67–75. [Google Scholar] [CrossRef]
- Howarth, R.W.; Butler, T.; Lunde, K.; Swaney, D.; Chu, C.R. Turbulence and planktonic nitrogen fixation: A mesocosm experiment. Limnol. Oceanogr. 1993, 38, 1696–1711. [Google Scholar] [CrossRef]
- Hemminga, M.A. The roots/rhizome system of seagrasses: An asset and a burden. J. Sea Res. 1998, 39, 183–196. [Google Scholar] [CrossRef]
- Capone, D.G.; Oremland, R.S.; Taylor, B.F. Significance of N2 fixation to the production of Thalassia testudinum communities. In Proceedings of the CICAR-II: Symposium on Progress in Marine Research in the Caribbean and Adjacent Regions, Caracas, Venezuela, 12–16 July 1976; pp. 71–85. [Google Scholar]
- Capone, D.G. Benthic Nitrogen Fixation. In Nitrogen in the Marine Environment; Carpenter, E.J., Capone, D.G., Eds.; Academic Press: New York, NY, USA, 1983; pp. 105–138. [Google Scholar]
- Nedwell, D.; Aziz, S. Heterotrophic nitrogen fixation in intertidal salt marsh sediment. Estuar. Coast. Mar. Sci. 1980, 10, 699–702. [Google Scholar] [CrossRef]
- Oremland, R.S.; Capone, D.G. Use of “specific” inhibitors in biogeochemistry and microbial ecology. Adv. Microb. Ecol. 1988, 10, 285–383. [Google Scholar]
- Cook, P.L.M.; Evrard, V.; Woodland, R.J. Factor controlling nitrogen fixation in temperate seagrass beds. Mar. Ecol. Prog. Ser. 2015, 525, 41–51. [Google Scholar] [CrossRef]
- Bertics, V.J.; Löscher, C.R.; Salonen, I.; Dale, A.W.; Gier, J.; Schmitz, R.A.; Treude, T. Occurrence of benthic microbial nitrogen fixation coupled to sulfate reduction in the seasonally hypoxic Eckernförde Bay, Baltic Sea. Biogeosciences 2013, 10, 1243–1258. [Google Scholar] [CrossRef] [Green Version]
- Fulweiler, R.W.; Nixon, S.W.; Buckley, B.A.; Granger, S.L. Reversal of the net dinitrogen gas flux in coastal marine sediments. Nature 2007, 448, 180–182. [Google Scholar] [CrossRef]
- Sim, M.S.; Ono, S.; Bosak, T. Effects of iron and nitrogen limitation on sulfur isotope fractionation during microbial sulfate reduction. Appl. Environ. Microbiol. 2012, 78, 8368–8376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaabjerg, V.; Mouritsen, K.N.; Finster, K. Diel cycles of sulphate reduction rates in sediments of a Zostera marina bed (Denmark). Aquat. Microb. Ecol. 1998, 15, 97–102. [Google Scholar] [CrossRef]
- Blackburn, T.H.; Nedwell, D.B.; Wiebe, W.J. Active mineral cycling in a Jamaican seagrass sediment. Mar. Ecol. Prog. Ser. 1994, 110, 233–239. [Google Scholar] [CrossRef]
- Lamers, L.P.M.; Govers, L.L.; Janssen, I.C.J.M.; Geurts, J.J.M.; Van Der Welle, M.E.W.; Van Katwijk, M.M.; Van Der Heide, T.; Roelofs, J.G.M.; Smolders, A.J.P. Sulfide as soil phytotoxin—A review. Front. Plant Sci. 2013, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.M. The future of seagrass meadows. Environ. Conserv. 2002, 29, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Borum, J.; Pedersen, O.; Greve, T.M.; Frankovich, T.A.; Zieman, J.C.; Fourqurean, J.W.; Madden, C.J. The potential role of plant oxygen and sulphide dynamics in die-off events of the tropical seagrass, Thalassia testudinum. J. Ecol. 2005, 93, 148–158. [Google Scholar] [CrossRef]
- Olinger, L.W.; Rogers, R.; Fore, P.; Todd, R.; Mullins, B.; Blisterfield, F.; Wise, L. Environmental and Recory Studies of Escambia Bay and the Pensacola Bay System; Florida US Environmental Protection Agency, Region IV: Gulf Breeze, FL, USA, 1975; pp. 1–345.
- Lehnen, N.; Marchant, H.K.; Schewdt, A.; Milucka, J.; Lott, C.; Weber, M.; Dekaezemacker, J.; Seah, B.K.B.; Hach, P.F.; Mohr, W.; et al. High rates of microbial dinitrogen fixation and sulfate reduction associated with the Mediterranean seagrass Posidonia oceanica. Syst. Appl. Microbiol. 2016, 39, 476–483. [Google Scholar] [CrossRef] [PubMed]
- USEPA. The Ecological Condition of the Pensacola Bay System, Northwest Florida (1994–2001); U.S. Environmental Protection Agency, Office of Research and Development, National Health and Ecological Effects Research Laboratory, Gulf Ecology Division: Gulf Breeze, FL, USA, 2004.
- NWFWMD. Pensacola Bay System Surface Water Improvement and Management Plan; Northwest Florida Water Management District: Havana, FL, USA, 2017. [Google Scholar]
- Devereaux, R.; Yates, D.F.; Aukamp, J.; Quarles, R.L.; Jordan, S.J.; Stanley, R.S.; Eldridge, P.M. Interactions of Thalassia testudinum and sediment biogeochemistry in Santa Rosa Sound, NW Florida. Mar. Biol. Res. 2011, 7, 317–331. [Google Scholar] [CrossRef]
- Macauley, J.M.; Clark, J.R.; Price, W.A. Seasonal changes in the standing crop and chlorophyll content of Thalassia testudinum Banks ex König and its epiphytes in the northern Gulf of Mexico. Aquat. Bot. 1988, 31, 277–287. [Google Scholar] [CrossRef]
- Lores, E.M.; Pasko, E.; Patrick, J.M.; Quarles, R.L.; Campbell, J.; Macauley, J. Mapping and monitoring of submerged aquatic egetation in Escambia-Pensacola Bay System, Florida. Gulf Mex. Sci. 2000, 18, 1–14. [Google Scholar]
- Scott, T.M. Geology of the Florida Platform. In Gulf of Mexico Origin, Waters, and Biota; Buster, N.A., Holmes, C.W., Eds.; Texas A&M University Press: College Station, TX, USA, 2011; Volume 3, pp. 17–31. [Google Scholar]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. Determination of Phosphate. In A Manual of Chemical and Biological Methods for Seawater Analysis; Pergamon Press: New York, NY, USA, 1984; pp. 22–24. [Google Scholar]
- Holmes, R.M.; Aminot, A.; Kérouel, R.; Hooker, B.A.; Peterson, B.J. A simple and precise method for measuring ammonium in marine and freshwater ecosystems. Can. J. Fish. Aquat. Sci. 1999, 56, 1801–1808. [Google Scholar] [CrossRef]
- Braman, R.S.; Hendrix, S.A. Nanogram nitrite and nitrate determination in environmental and biological materials by vanadium(III) reduction with chemiluminescence detection. Anal. Chem. 1989, 61, 2715–2718. [Google Scholar] [CrossRef]
- Fonselius, S.H. Determination of Hydrogen Sulphide. In Methods of Seawater Analysis; Grasshoff, K., Ehrhardt, M., Kremling, K., Eds.; Wiley-VCH Verlag GmbH: Weinheim, Germany, 1983; pp. 73–80. [Google Scholar]
- Welshmeyer, N.A. Fluorometric analysis of chlorophyll a in the presence of chlorphyll b and pheopigments. Limnol. Oceanogr. 1994, 39, 1985–1992. [Google Scholar] [CrossRef]
- Capone, D.G. Determination of Nitrogenase Activity in Aquatic Samples Using the Acetylene Reduction Procedure. In Handbook of Methods in Aquatic Microbial Ecology; Kemp, P.F., Cole, J.J., Sherr, B.F., Sherr, E.B., Eds.; CRC Press: Boca Raton, FL, USA, 1993; pp. 621–632. [Google Scholar]
- Capone, D.G.; Montoya, J.P. Nitrogen fixation and denitrification. Method. Microbiol. 2001, 30, 501–515. [Google Scholar]
- Atkinson, M.J.; Smith, S.V. C:N:P ratios of benthic marine plants. Limnol. Oceanogr. 1983, 28, 568–574. [Google Scholar] [CrossRef]
- Carlson, P.R.; Yarbro, L.A.; Barber, T.R. Relationship of sediment sulfide to mortality of Thalassia testudinum in Florida Bay. Bull. Mar. Sci. 1994, 54, 733–746. [Google Scholar]
- Price, W.A. A Comparison of Nitrogen Fixation (Acetylene Reduction) Rates in Seagrass Communities Dominated by Thalassia Testudinum and Halodule Wrightii; University of West Flordia: Pensacola, FL, USA, 1988. [Google Scholar]
- McRoy, C.P.; Goering, J.J.; Chaney, B. Nitrogen fixation associated with seagrasses. Limnol. Oceanogr. 1973, 18, 998–1002. [Google Scholar] [CrossRef]
- Capone, D.G.; Penhale, P.A.; Oremland, R.S.; Taylor, B.F. Relationship between productivity and N2 (C2H2) fixation in a Thalassia testudinum community. Limnol. Oceanogr. 1979, 24, 117–125. [Google Scholar] [CrossRef]
- Patriquin, D.; Knowles, R. Nitrogen fixation in the rhizosphere of marine angiosperms. Mar. Biol. 1972, 16, 49–58. [Google Scholar] [CrossRef]
- Smith, G.W.; Hayasaka, S.S. Nitrogenase actvity associated with Halodule wrightii roots. Appl. Environ. Microbiol. 1982, 43, 1244–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartog, C.D. The Sea-Grasses of the World; North-Holland Publishing Co.: Amsterdam, The Netherlands, 1970. [Google Scholar]
- Moshiri, G.A.; Aumen, N.G.; Swann, W.G. Water Quality Studies in Santa Rosa Sound; US Environmental Protection Agency: Gulf Breeze, FL, USA, 1980; pp. 1–71.
- Lee, K.S.; Park, S.R.; Park, Y.K. Effects of irradiance, temperature, and nutrients on growth dynamics of seagrass: A review. J. Exp. Mar. Biol. Ecol. 2007, 350, 144–175. [Google Scholar] [CrossRef]
- Moore, D.R. Distribution of the sea grass, Thalassia, in the United States. Bull. Mar. Sci. 1963, 13, 329–342. [Google Scholar]
- Lee, K.S.; Dunton, K.H. Effects of nitrogen enrichment on biomass allocation, growth, and leaf morphology of the seagrass Thalassia testudinum. Mar. Ecol. Prog. Ser. 2000, 196, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Fourqurean, J.W.; Cai, Y. Arsenic and phosphorus in seagrass leaves from the Gulf of Mexico. Aquat. Bot. 2001, 71, 247–258. [Google Scholar] [CrossRef]
- Johnson, M.W.; Heck, K.L., Jr.; Fourqurean, J.W. Nutrient content of seagrasses and epiphytes in the northern Gulf of Mexico: Evidence of phosphorus and nitrogen limitation. Aquat. Bot. 2006, 85, 103–111. [Google Scholar] [CrossRef]
- Lee, K.S.; Dunton, K.H. Diurnal changes in pore water sulfide concentrations in the seagrass Thalassia testudinum beds: The effects of seagrasses on sulfide dynamic. J. Exp. Mar. Biol. Ecol. 2000, 255, 201–214. [Google Scholar] [CrossRef]
- Borum, J.; Sand-Jensen, K.; Binzer, T.; Pedersen, O. Oxygen movement in seagrasses. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 255–270. [Google Scholar]
- Sand-Jensen, K.; Prahl, C.; Stokholm, H. Oxygen release from roots of submerged aquatic macrophytes. Oikos 1982, 38, 349–354. [Google Scholar] [CrossRef]
- Jovanovic, Z.; Pedersen, M.Ø.; Larsen, M.; Kristensen, E.; Glud, R.N. Rhizosphere O2 dynamics in young Zostera marina and Ruppia maritima. Mar. Ecol. Prog. Ser. 2015, 518, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Caffrey, J.M.; Kemp, W.M. Seasonal and spatial patterns of oxygen production, respiration and root-rhizome release in Potamogeton perfoliatus L. and Zostera marina L. Aquat. Bot. 1991, 40, 109–128. [Google Scholar] [CrossRef]
- Koch, M.S.; Erskine, J.M. Sulfide as a phytotoxin to the tropical seagrass Thalassia testudinum interactions with light, salinity and temperature. J. Exp. Mar. Biol. Ecol. 2001, 266, 81–95. [Google Scholar] [CrossRef]
- Koch, M.S.; Schopmeyer, S.; Kyhn-Hansen, C.; Madden, C.J. Synergistic effects of high temperature and sulfide on tropical seagrass. J. Exp. Mar. Biol. Ecol. 2007, 341, 91–101. [Google Scholar] [CrossRef]
- Johnson, C.R.; Koch, M.S.; Pederson, O.; Madden, C.J. Hypersalinity as a trigger of seagrass (Thalassia testudinum) die-off events in Florida Bay: Evidence based on shoot meristem O2 and H2S dynamics. J. Exp. Mar. Biol. Ecol. 2018, 504, 47–52. [Google Scholar] [CrossRef]
- Knapp, A.N. The sensitivity of marine N2 fixation to dissolved inorganic nitrogen. Front. Microbiol. 2012, 3, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caffrey, J.M.; Albrecht, B.; Capps, R.E. Seagrass Abundance and Productivity in Pensacola Bay and Santa Rosa Sound; Florida Fish and Wildlife Comission FIsh and Wildlife Research Insitute: St. Petersburg, FL, USA, 2017. [Google Scholar]
- Duarte, C.M.; Chiscano, C.L. Seagrass biomass and production: A reassessment. Aquat. Bot. 1999, 65, 159–174. [Google Scholar] [CrossRef]
- Cole, L.W.; McGlathery, K.J. Nitrogen fixation in restored eelgrass meadows. Mar. Ecol. Prog. Ser. 2012, 448, 235–246. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate Type | Unvegetated | H. wrightii | T. testudinum | |
|---|---|---|---|---|
| Sediment Water (%) | Range | 17.2–22.2 | 20.9–32.2 | 21.2–44.9 |
| Average ± SE | 20.2 ± 0.2 | 26.2 ± 0.3 | 28.9 ± 0.7 | |
| Sediment Organic Matter (%) | Range | 0.09–0.89 | 0.30–2.75 | 0.34–2.07 |
| Average ± SE | 0.29 ± 0.02 | 1.14 ± 0.07 | 1.27 ± 0.06 | |
| Dissolved Inorganic Phosphate (DIP) (µM) | Range | 0.1–1.7 | 0.6–3.0 | 0.6–3.3 |
| Average ± SE | 0.7 ± 0.1 | 1.6 ± 0.2 | 1.6 ± 0.2 | |
| Ammonium (µM) | Range | 8.0–46.9 | 2.9–48.0 | 2.2–33.7 |
| Average ± SE | 21.6 ± 3.4 | 10.8 ± 3.1 | 9.6 ± 2.7 | |
| Porewater Nitrogen:Phosphorus Ratio (N:P) | Range | 16.8–107 | 1.6–18.2 | 0.7–19.0 |
| Average ± SE | 39.6 ± 6.1 | 7.0 ± 1.3 | 6.0 ± 1.3 | |
| N:P Seagrass Leaves | N/A | 16 | 26 | |
| Substrate Type | Location | Season or Month | Nitrogen Fixation Rate (mg N m−2 d−1) | Reference |
|---|---|---|---|---|
| T. testudinum | NW Florida, USA | Early Growth (Mar.-Apr.) | 2.40 ± 0.95 | This Study |
| Peak Growth (May-Jul.) | 1.40 ± 0.89 | This Study | ||
| Late Growth (Aug.-Sept.) | 2.48 ± 0.66 | This Study | ||
| Senescence (Sept.-Nov.) | 3.33 ± 0.57 | This Study | ||
| Dormancy (Dec.-Feb.) | 2.04 ± 0.74 | This Study | ||
| Spring | 2.54 | [53] | ||
| Summer | 4.21 | [53] | ||
| Fall | 10.45 | [53] | ||
| Winter | 4.95 | [53] | ||
| Florida, USA | January/March | 0.03 | [54] | |
| Biscayne Bay, Florida, USA | August/October | 5–24 | [19] | |
| Bimini Harbor, Bahamas | July | 5.1–5.3 | [55] | |
| August | 6–9 | [55] | ||
| Barbados | September | 27–140 | [56] | |
| H. wrightii | NW Florida, USA | Early Growth (Mar.-Apr.) | 1.42 ± 0.57 | This Study |
| Peak Growth (May-Jul.) | 1.74 ± 0.94 | This Study | ||
| Late Growth (Aug.-Sept.) | 3.06 ± 0.69 | This Study | ||
| Senescence (Sept.-Nov.) | 4.66 ± 0.97 | This Study | ||
| Dormancy (Dec.-Feb.) | 3.53 ± 0.68 | This Study | ||
| Spring | 5.50 | [53] | ||
| Summer | 3.15 | [53] | ||
| Fall | 7.39 | [53] | ||
| Winter | 8.25 | [53] | ||
| North Carolina, USA | June | 0.021 | [57] | |
| August | 0.023 | [57] | ||
| November | 0.001 | [57] | ||
| H. beaudettei | Jamaica | December | 28 | [32] |
| Unvegetated | NW Florida, USA | Early Growth (Mar.-Apr.) | 0.53 ± 0.31 | This Study |
| Peak Growth (May-Jul.) | 0.82 ± 0.39 | This Study | ||
| Late Growth (Aug.-Sept.) | 1.27 ± 0.46 | This Study | ||
| Senescence (Sept.-Nov.) | 1.68 ± 1.06 | This Study | ||
| Dormancy (Dec.-Feb.) | 0.13 ± 0.07 | This Study | ||
| Spring | 2.61 | [53] | ||
| Summer | 12.05 | [53] | ||
| Fall | 3.89 | [53] | ||
| Winter | 64.71 | [53] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Presley, R.; Caffrey, J.M. Nitrogen Fixation in Subtropical Seagrass Sediments: Seasonal Patterns in Activity in Santa Rosa Sound, Florida, USA. J. Mar. Sci. Eng. 2021, 9, 766. https://doi.org/10.3390/jmse9070766
Presley R, Caffrey JM. Nitrogen Fixation in Subtropical Seagrass Sediments: Seasonal Patterns in Activity in Santa Rosa Sound, Florida, USA. Journal of Marine Science and Engineering. 2021; 9(7):766. https://doi.org/10.3390/jmse9070766
Chicago/Turabian StylePresley, Rachel, and Jane M. Caffrey. 2021. "Nitrogen Fixation in Subtropical Seagrass Sediments: Seasonal Patterns in Activity in Santa Rosa Sound, Florida, USA" Journal of Marine Science and Engineering 9, no. 7: 766. https://doi.org/10.3390/jmse9070766
APA StylePresley, R., & Caffrey, J. M. (2021). Nitrogen Fixation in Subtropical Seagrass Sediments: Seasonal Patterns in Activity in Santa Rosa Sound, Florida, USA. Journal of Marine Science and Engineering, 9(7), 766. https://doi.org/10.3390/jmse9070766