Assessment of Halamphora coffeaeformis Growth and Biochemical Composition for Aquaculture Purposes

 ,
,
Abstract
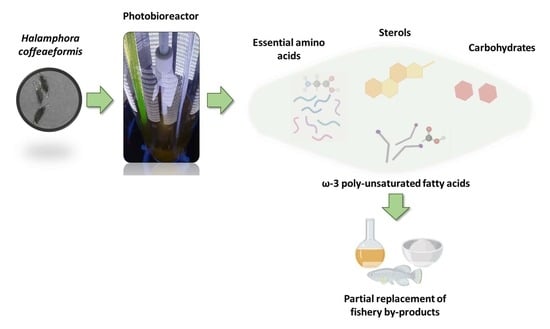
1. Introduction
2. Material and Methods
2.1. Experimental Design
2.2. Growth rate and Biomass
2.3. Lipid Determinations
2.3.1. Quantification of Triacylglycerides (TAG) and Sterols
2.3.2. Fatty Acid Characterization
2.3.3. Sterol Characterization
2.4. Protein Determinations
2.4.1. Total Protein Quantification
2.4.2. Amino Acid Characterization
2.5. Determination of Total Carbohydrates
2.6. Statistical Analysis
3. Results
3.1. Growth and Biomass Production of H. coffeaeformis
3.2. Triglyceride Production and Fatty Acid and Sterol Composition
3.3. Protein Production and Amino Acid Composition
3.4. Carbohydrate Production
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Borowitzka, M.A. High-value products from microalgae-their development and commercialization. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Senroy, S.; Pal, R. Microalgae in aquaculture: A review with special references to nutritional value and fish dietetics. Proc. Zool. Soc. Lond. 2015, 68, 1–8. [Google Scholar] [CrossRef]
- Sirakov, I.; Velichkova, K.; Stoyanova, S.; Staykov, Y. The importance of microalgae for aquaculture industry. Review. Int. J. Fish. Aquat. Stud. 2015, 2, 81–84. [Google Scholar]
- Muller-Feuga, A. The role of microalgae in aquaculture: Situation and trends. J. Appl. Phycol. 2000, 12, 527–534. [Google Scholar] [CrossRef]
- Brown, M.R.; Jeffrey, S.W.; Volkman, J.K.; Dunstan, G.A. Nutritional properties of microalgae for mariculture. Aquaculture. 1997, 151, 315–331. [Google Scholar] [CrossRef]
- D’Souza, F.M.L.; Lorenagan, N.R. Effects of monospecific and mixed-algae diets, on survival, development and fatty acid composition of penaeid prawn (Penaeus spp.) larvae. Mar. Biol. 1999, 133, 621–633. [Google Scholar] [CrossRef]
- Martín, L.A.; Popovich, C.A.; Martinez, A.M.; Damiani, M.C.; Leonardi, P.I. Oil assessment of Halamphora coffeaeformis diatom growing in a hybrid two-stage system for biodiesel production. Renew. Energy 2016, 92, 127–135. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Kilham, P.; Jackson, T.A. Kinetics of silicon-limited growth in the marina diatom Thalassiosira pseudonana. J. Phycol. 1973, 9, 233–237. [Google Scholar]
- Barnech Bielsa, G.; Popovich, C.; Rodríguez, M.C.; Martínez, A.; Martín, L.A.; Matulewicz, M.C.; Leonardi, P.I. Simultaneous production assessment of triacylglycerols for biodiesel and exopolysaccharides as valuable co-products in Navicula cincta. Algal Res. 2016, 15, 120–128. [Google Scholar] [CrossRef]
- Martín, L.A.; Popovich, C.A.; Martinez, A.M.; Scodelaro Bilbao, P.G.; Damiani, M.C.; Leonardi, P.I. Hybrid two-stage culture of Halamphora coffeaeformis for biodiesel production: Growth phases, nutritional stages and biorefinery approach. Renew. Energy 2018, 118, 984–992. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Scodelaro Bilbao, P.G.; Damiani, M.C.; Salvador, G.A.; Leonardi, P.I. Haematococcus pluvialis as a source of fatty acids and phytosterols: Potential nutritional and biological implications. J. Appl. Phycol. 2016, 28, 3283–3294. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Brown, M.R. The amino-acid and sugar composition of 16 species of microalgae used in mariculture. J. Exp. Mar. Biol. Ecol. 1991, 145, 79–99. [Google Scholar] [CrossRef]
- Simpson, R.J.; Neuberger, M.R.; Liu, T.Y. Complete amino acid analysis of proteins from a single hydrolysate. J. Biol. Chem. 1976, 251, 1936–1940. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Li, P.; Mai, K.; Trushenski, J.; Wu, G. New developments in fish amino acid nutrition: Towards functional and environmentally oriented aquafeeds. Amino Acids 2009, 37, 43–53. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef]
- Shah, M.R.; Lutzu, G.A.; Alam, A.; Sarker, P.; Chowdhury, K.; Parsaeimehr, A.; Liang, Y.; Daroch, M. Microalgae in aquafeeds for a sustainable aquaculture industry. J. Appl. Phycol. 2018, 30, 197–213. [Google Scholar] [CrossRef]
- Scholz, B.; Liebezeit, G. Growth responses of 25 benthic marine Wadden Sea diatoms isolated from the Solthörn tidal flat (southern North Sea) in relation to varying culture conditions. Diatom Res. 2012, 27, 65–73. [Google Scholar] [CrossRef]
- Yi, Z.; Xu, M.; Di, X.; Brynjolfsson, S.; Fu, W. Exploring Valuable Lipids in Diatoms. Front. Mar. Sci. 2017, 4, 17. [Google Scholar] [CrossRef]
- Yang, Y.J.; Du, L.; Hosokawa, M.; Miyashita, K.; Kokubun, Y.; Arai, H.; Taroda, H. Fatty acid and lipid class composition of the microalga Phaeodactylum tricornutum. J. Oleo Sci. 2017, 66, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Zulu, N.N.; Zienkiewicz, K.; Vollheyde, K.; Feussner, I. Current trends to comprehend lipid metabolism in diatoms. Progr. Lipid Res. 2018, 70, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Almeyda, M.D.; Scodelaro Bilbao, P.G.; Popovich, C.A.; Constenla, D.; Leonardi, P.I. Enhancement of polyunsaturated fatty acid production under low-temperature stress in Cylindrotheca closterium. J. Appl. Phycol. 2020. [Google Scholar] [CrossRef]
- Sayanova, O.; Mimouni, V.; Ulmann, L.; Morant-Manceau, A.; Pasquet, V.; Schoefs, B.; Napier, J.A. Modulation of lipid biosynthesis by stress in diatoms. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160407. [Google Scholar] [CrossRef]
- Benemann, J.R. Microalgae aquaculture feeds. J. Appl. Phycol. 1992, 4, 233–245. [Google Scholar] [CrossRef]
- Ryckebosch, E.; Bruneel, C.; Termote-Verhalle, R.; Goiris, K.; Muylaert, K.; Fouberta, I. Nutritional evaluation of microalgae oils rich in omega-3 long chain polyunsaturated fatty acids as an alternative for fish oil. Food Chem. 2014, 160, 393–400. [Google Scholar] [CrossRef] [PubMed]
- De Silva, S.; Francis, D.S.; Tacon, A.G.J. Fish oil in aquaculture: In retrospect. In Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; Turchini, G.M., Ng, W.K., Tocher, D.R., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 1–20. [Google Scholar]
- Watanabe, T.; Tamiya, T.; Oka, A.; Hirata, M.; Kitajima, C.; Fujita, S. Improvement of dietary value of live foods for fish larvae by feeding them on ω-3 highly unsaturated fatty acids and fat-soluble vitamins. NIPPON SUISAN GAKKAISHI 1983, 49, 471–479. [Google Scholar] [CrossRef][Green Version]
- Volkman, J.K. Sterols in microalgae. In The Physiology of Microalgae. Developments in Applied Phycology; Borowitzka, M., Beardall, J., Raven, J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 6, pp. 485–505. [Google Scholar] [CrossRef]
- Gladu, P.K.; Patterson, G.W.; Wikfors, G.H.; Chitwood, D.J.; Lusby, W.R. Sterols of some diatoms. Phytochemistry 1991, 30, 2301–2303. [Google Scholar] [CrossRef]
- Rampen, S.W.; Abbas, B.A.; Schouten, S.; Sinninghe Damsté, J.S. A comprehensive study of sterols in marine diatoms (Bacillariophyta): Implications for their use as tracers for diatom productivity. Limnol. Oceanogr. 2010, 55, 91–105. [Google Scholar] [CrossRef]
- Barrett, S.M.; Volkman, J.K.; Dunstan, G.A.; LeRoi, J.-M. Sterols of 14 species of marine diatoms (Bacillariophyta). J. Phycol. 1995, 31, 360–369. [Google Scholar] [CrossRef]
- Norambuena, F.; Lewis, M.; Hamid, N.K.A.; Hermon, K.; Donald, J.A.; Turchini, G.M. Fish oil replacement in current aquaculture feed: Is cholesterol a hidden treasure for fish nutrition? PLoS ONE 2013, 8, e81705. [Google Scholar] [CrossRef] [PubMed]
- Cowey, C.B. Amino acid requirements of fish: A critical appraisal of present values. Aquaculture 1994, 124, 1–11. [Google Scholar] [CrossRef]
- Brunty, J.L.; Bucklin, R.A.; Davis, J.; Baird, C.D.; Nordstedt, R.A. The influence of feed protein intake on tilapia ammonia production. Aquacult. Eng. 1997, 16, 161–166. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018. [Google Scholar]
- Wilson, R.P. Utilization of dietary carbohydrate by fish. Aquaculture 1994, 124, 67–80. [Google Scholar] [CrossRef]
- Buhler, D.R.; Halver, J.E. Nutrition of salmonoid fishes. Carbohydrate requirements of chinook salmon. J. Nutr. 1961, 74, 307–318. [Google Scholar] [CrossRef]
- Tibbetts, S.; Milley, J.; Lall, S. Chemical composition and nutritional properties of freshwater and marine microalgal biomass cultured in photobioreactors. J. Appl. Phycol. 2014, 27, 1109–1119. [Google Scholar] [CrossRef]
- Krogdahl, A.; Hemre, G.-I.; Mommsen, T.P. Carbohydrates in fish nutrition: Digestion and absorption in postlarval stages. Aquac. Nutr. 2005, 11, 103–122. [Google Scholar] [CrossRef]
- Courtois de Viçose, G.; Porta, A.; Viera, M.P.; Fernández-Palacios, H.; Izquierdo, M.S. Effects of density on growth rates of four benthic diatoms and variations in biochemical composition associated with growth phase. J. Appl. Phycol. 2012, 24, 1427–1437. [Google Scholar] [CrossRef]
- Brown, M.R.; Dunstan, G.A.; Norwood, S.J.; Miller, K.A. Effects of harvest stage and light on the biochemical composition of the diatom Thalassiosira pseudonana. J. Phycol. 1996, 32, 64–73. [Google Scholar] [CrossRef]
- Chu, W.-L.; Phang, S.-M.; Goh, S.-H. Environmental effects on growth and biochemical composition of Nitzschia inconspicua Grunow. J. Appl. Phycol. 1996, 8, 389–396. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids (%) | |||
|---|---|---|---|
| C 14:0 (myristic) | 14.11 | ± | 0.390 |
| C 16:0 (palmitic) | 25.82 | ± | 0.291 |
| C 16:1 (palmitoleic) | 38.05 | ± | 0.513 |
| C 18:0 (stearic) | 0.55 | ± | 0.027 |
| C 18:1ω9c (oleic) | 4.03 | ± | 0.402 |
| C 18:2ω6c (linoleic) | 3.52 | ± | 0.074 |
| C 18:3ω6 (ɤ linolenic) | 0.95 | ± | 0.053 |
| C 20:2 (eicosadienoic) | 2.56 | ± | 0.191 |
| C 20:4ω6 (arachidonic) | 1.02 | ± | 0.094 |
| C 20:5ω3 (eicosapentaenoic) | 9.40 | ± | 1.062 |
| Amino Acids | % of Total Amino Acids | ||
| Essential* | |||
| Arginine | 7.04 | ± | 1.48 |
| Histidine | 2.30 | ± | 0.36 |
| Isoleucine | 4.15 | ± | 0.41 |
| Leucine | 8.21 | ± | 0.67 |
| Lysine | 6.01 | ± | 0.63 |
| Methionine | 4.47 | ± | 0.09 |
| Phenylalanine | 6.50 | ± | 0.49 |
| Threonine | 5.65 | ± | 0.58 |
| Tryptophan | nd | ||
| Valine | 6.19 | ± | 0.18 |
| Conditionally essential* | |||
| Cysteine | 0.21 | ± | 0.02 |
| Glutamine | nd | ||
| Hydroxyproline | nd | ||
| Proline | 3.07 | ± | 0.09 |
| Taurine | nd | ||
| Non-essential AA | |||
| Alanine | 6.63 | ± | 0.67 |
| Asparagine | nd | ||
| Aspartate | 11.14 | ± | 1.17 |
| Glutamate | 12.72 | ± | 0.58 |
| Glycine | 5.71 | ± | 0.36 |
| Serine | 6.18 | ± | 0.54 |
| Tyrosine | 4.05 | ± | 0.45 |
| H. coffeaeformis | Fish Oils * | |
|---|---|---|
| Essential Fatty Acids | % | % |
| EPA | 9.40 ± 1.06 | 7.175–15.375 |
| ARA | 1.02 ± 0.09 | 0.375 |
| DHA | n.d. | 4.875–10.075 |
| Sterols | ||
| Cholesterol | 12.4 ± 0.12 | 4.4 |
| Phytosterols | 87.5 ± 0.20 | n.d. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scodelaro Bilbao, P.G.; Martín, L.A.; Popovich, C.A.; Almeyda, M.D.; Chamorro, V.; Leonardi, P.I. Assessment of Halamphora coffeaeformis Growth and Biochemical Composition for Aquaculture Purposes. J. Mar. Sci. Eng. 2020, 8, 282. https://doi.org/10.3390/jmse8040282
Scodelaro Bilbao PG, Martín LA, Popovich CA, Almeyda MD, Chamorro V, Leonardi PI. Assessment of Halamphora coffeaeformis Growth and Biochemical Composition for Aquaculture Purposes. Journal of Marine Science and Engineering. 2020; 8(4):282. https://doi.org/10.3390/jmse8040282
Chicago/Turabian StyleScodelaro Bilbao, Paola G., Lucas A. Martín, Cecilia A. Popovich, María Delfina Almeyda, Verónica Chamorro, and Patricia I. Leonardi. 2020. "Assessment of Halamphora coffeaeformis Growth and Biochemical Composition for Aquaculture Purposes" Journal of Marine Science and Engineering 8, no. 4: 282. https://doi.org/10.3390/jmse8040282
APA StyleScodelaro Bilbao, P. G., Martín, L. A., Popovich, C. A., Almeyda, M. D., Chamorro, V., & Leonardi, P. I. (2020). Assessment of Halamphora coffeaeformis Growth and Biochemical Composition for Aquaculture Purposes. Journal of Marine Science and Engineering, 8(4), 282. https://doi.org/10.3390/jmse8040282