
Early Succession Patterns of Benthic Assemblages on Artificial Reefs in the Oligotrophic Eastern Mediterranean Basin

 , ,
, ,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Image Processing
2.3. Statistical Analysis
3. Results
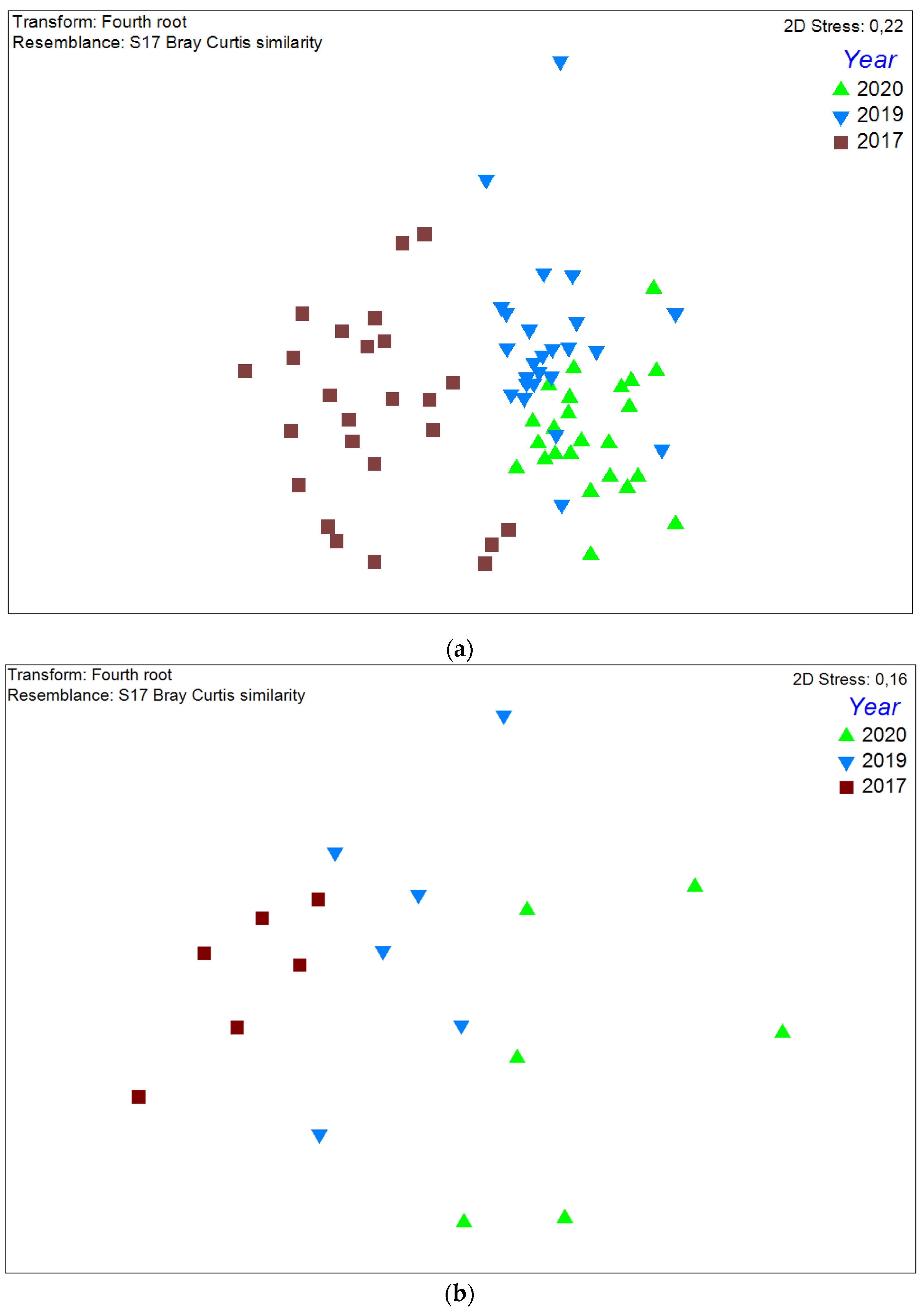
3.1. Community Structure
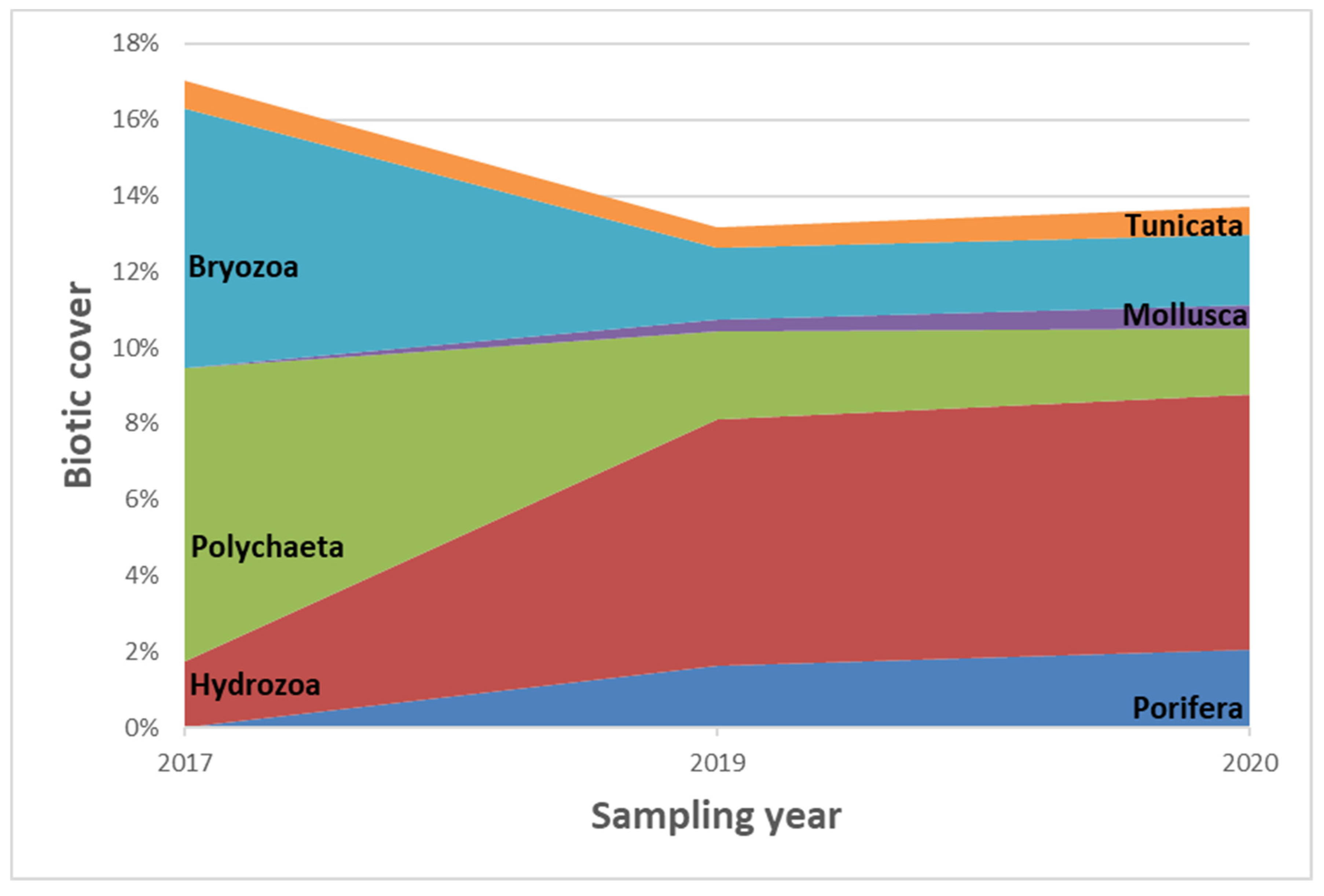
3.1.1. Sessile Community
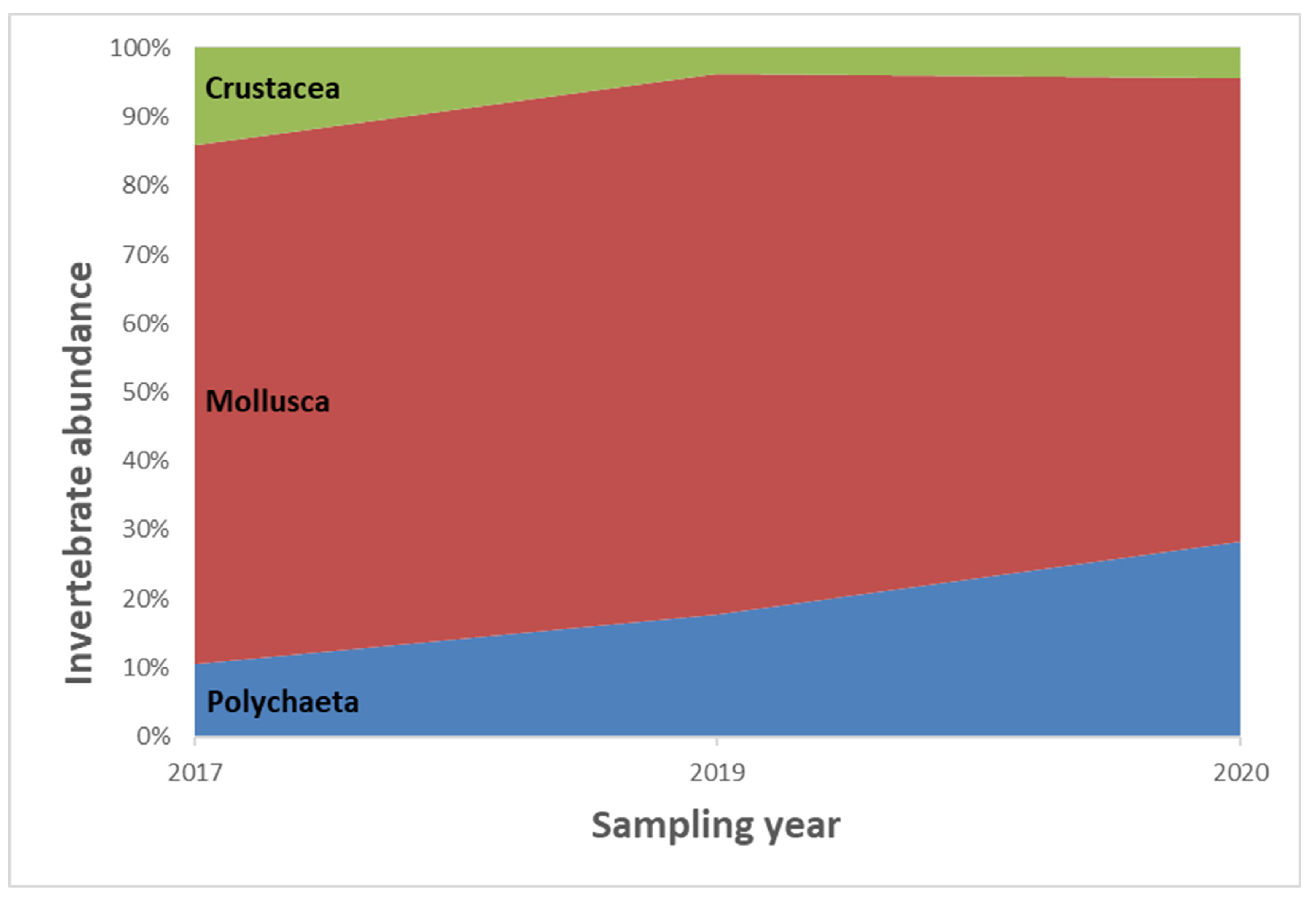
3.1.2. Motile Community
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Manoudis, G.; Antoniadou, C.; Dounas, K.; Chintiroglou, C. Successional stages of experimental artificial reefs deployed in vistonikos gulf (N. Aegean Sea, Greece): Preliminary results. Belg. J. Zool. 2005, 135, 209–215. [Google Scholar]
- Airoldi, L.; Beck, M.W. Loss, status and trends for coastal marine habitats of Europe. Oceanogr. Mar. Biol. 2007, 45, 345–405. [Google Scholar] [CrossRef]
- Fabi, G.; Spagnolo, A.; Bellan-Santini, D.; Charbonnel, E.; Çiçek, B.A.; García, J.J.G.; Jensen, A.C.; Kallianiotis, A. Overview on artificial reefs in Europe. Braz. J. Oceanogr. 2011, 59, 155–166. [Google Scholar] [CrossRef]
- Sinis, A.I.; Chintiroglou, C.C.; Stergiou, K.I. Preliminary results from the establishment of experimental artificial reefs in the N. Aegean Sea (Chalkidiki, Greece). Belg. J. Zool. 2000, 130, 143–147. [Google Scholar]
- Klaoudatos, D.S.; Anastasopoulou, A.; Papaconstantinou, C.; Conides, A.I. The Greek experience of artificial reef construction and management. J. Environ. Prot. Ecol. 2012, 13, 1647–1655. [Google Scholar]
- Seaman, W. Unifying trends and opportunities in global artificial reef research, including evaluation. ICES J. Mar. Sci. 2002, 59, S14–S16. [Google Scholar] [CrossRef]
- Achilleos, K.; Patsalidou, M.; Jimenez, C.; Kamidis, N.; Georgiou, A.; Petrou, A.; Kallianiotis, A. Epibenthic communities on artificial reefs in Greece, Mediterranean Sea. Water 2018, 10, 347. [Google Scholar] [CrossRef]
- Psarra, S.; Tselepides, A.; Ignatiades, L. Primary productivity in the Oligotrophic Cretan Sea (NE Mediterranean): Seasonal and interannual variability. Prog. Oceanogr. 2000, 46, 187–204. [Google Scholar] [CrossRef]
- Tselepides, A.; Papadopoulou, K.N.; Podaras, D.; Plaiti, W.; Koutsoubas, D. Macrobenthic Community Structure over the continental margin of Crete (South Aegean Sea, NE Mediterranean). Prog. Oceanogr. 2000, 46, 401–428. [Google Scholar] [CrossRef]
- Georgopoulos, D.; Theocharis, A.; Zodiatis, G. Intermediate water formation in the Cretan Sea (South Aegean Sea). Oceanol. Acta 1989, 12, 353–359. [Google Scholar]
- Androulakis, D.N.; Dounas, C.G.; Margaris, D.P. The seafloor observatory of the underwater biotechnological park of Crete. In Proceedings of the 6th International Conference on “Experiments/Process/System Modelling/Simulation/Optimization”, Athens, Greece, 8–11 July 2015. [Google Scholar]
- Dounas, C.; Androulakis, D.; Dailianis, A.; Koulouri, P. Recreational diving Oasis with artificial habitats. Rapp. Comm. Int. Mer Médit 2016, 41, 476. [Google Scholar]
- Jensen, A.; Collins, K.; Lockwood, P. Current issues relating to artificial reefs in European seas. In Artificial Reefs in European Seas; Jensen, A.C., Collins, K.J., Lockwood, A.P.M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 489–499. ISBN 978-0-7923-6144-2. [Google Scholar]
- Jimenez, C.; Hadjioannou, L.; Petrou, A.; Andreou, V.; Georgiou, A. Fouling communities of two accidental artificial reefs (modern shipwrecks) in Cyprus (Levantine sea). Water 2016, 9, 11. [Google Scholar] [CrossRef]
- Jimenez, C.; Andreou, V.; Evriviadou, M.; Munkes, B.; Hadjioannou, L.; Petrou, A.; Abu Alhaija, R. Epibenthic communities associated with unintentional artificial reefs (modern shipwrecks) under contrasting regimes of nutrients in the Levantine sea (Cyprus and Lebanon). PLoS ONE 2017, 12, e0182486. [Google Scholar] [CrossRef] [PubMed]
- Gerovasileiou, V.; Voultsiadou, E. Sponge diversity gradients in Marine caves of the Eastern Mediterranean. J. Mar. Biol. Assoc. UK 2016, 96, 407–416. [Google Scholar] [CrossRef]
- Dimarchopoulou, D.; Gerovasileiou, V.; Voultsiadou, E. Spatial variability of sessile benthos in a semi-submerged Marine cave of a remote Aegean Island (Eastern Mediterranean sea). Reg. Stud. Mar. Sci. 2018, 17, 102–111. [Google Scholar] [CrossRef]
- Sedano, F.; Florido, M.; Rallis, I.; Espinosa, F.; Gerovasileiou, V. Comparing sessile benthos on shallow artificial versus natural hard substrates in the Eastern Mediterranean sea. Mediterr. Mar. Sci. 2019, 20, 688. [Google Scholar] [CrossRef]
- Çinar, M.E.; Féral, J.; Arvanitidis, C.; David, R.; Taşkin, E.; Sini, M.; Dailianis, T.; Doğan, A.; Gerovasileiou, V.; Evcen, A.; et al. Coralligenous assemblages along their geographical distribution: Testing of concepts and implications for management. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 1578–1594. [Google Scholar] [CrossRef]
- Chatzigeorgiou, G.; Faulwetter, S.; López, E.; Sardá, R.; Arvanitidis, C. Can coastal biodiversity measured in four Mediterranean sites be representative of the region? A test for the robustness of the NaGISA protocol by using the hard substrate syllid (Annelida, Polychaeta) taxo-communities as a Surrogate. Hydrobiologia 2012, 691, 147–156. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Pronzato, R.; Cattaneo-Vietti, R.; Benedetti Cecchi, L.; Morri, C.; Pansini, M.; Chemello, R.; Milazzo, M.; Fraschetti, S.; Terlizzi, A.; et al. Hard bottoms. In Mediterranean Marine Benthos: A Manual of Methods for Its Sampling and Study; Gambi, M.C., Dappiano, M., Eds.; Società Italiana di Biologia Marina: Genova, Italy, 2004; Volume 11, (Suppl. S1), pp. 185–215. [Google Scholar]
- Androulakis, D.; Dounas, C.; Banks, A.; Magoulas, A.; Margaris, D. An assessment of computational fluid dynamics as a tool to aid the design of the HCMR-artificial-reefsTM diving Oasis in the underwater biotechnological park of Crete. Sustainability 2020, 12, 4847. [Google Scholar] [CrossRef]
- Trygonis, V.; Sini, M. PhotoQuad: A dedicated seabed image processing software, and a comparative error analysis of four photoquadrat methods. J. Exp. Mar. Biol. Ecol. 2012, 424–425, 99–108. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Dimitriadis, C.; Arvanitidis, C.; Voultsiadou, E. Taxonomic and functional surrogates of sessile benthic diversity in Mediterranean Marine caves. PLoS ONE 2017, 12, e0183707. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E Ltd.: Plymouth, UK, 2006. [Google Scholar]
- Clarke, K.; Warwick, R. A further biodiversity index applicable to species lists: Variation in taxonomic distinctness. Mar. Ecol. Prog. Ser. 2001, 216, 265–278. [Google Scholar] [CrossRef]
- Ardizzone, G.; Somaschini, A.; Belluscio, A. Prediction of benthic and fish colonization on the fregene and other Mediterranean artificial reefs. In Artificial Reefs in European Seas; Jensen, A.C., Collins, K.J., Lockwood, A.P.M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 113–128. ISBN 978-0-7923-6144-2. [Google Scholar]
- Riggio, S.; Badalamenti, F.; D’Anna, G. Artificial reefs in Sicily: An overview. In Artificial Reefs in European Seas; Jensen, A.C., Collins, K.J., Lockwood, A.P.M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 65–74. ISBN 978-0-7923-6144-2. [Google Scholar]
- Kocak, F.; Ergen, Z.; Çinar, M.E. Fouling organisms and their developments in a polluted and an unpolluted marina in the Aegean sea (Turkey). Ophelia 1999, 50, 1–20. [Google Scholar] [CrossRef]
- Badalamenti, F.; D’Anna, G.; Riggio, S. Artificial reefs in the gulf of castellammare (North-West Sicily): A case study. In Artificial Reefs in European Seas; Jensen, A.C., Collins, K.J., Lockwood, A.P.M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 75–96. ISBN 978-0-7923-6144-2. [Google Scholar]
- D’Anna, G.; Badalamenti, F.; Riggio, S. Artificial reefs in North West Sicily: Comparisons and conclusions. In Artificial Reefs in European Seas; Jensen, A.C., Collins, K.J., Lockwood, A.P.M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 97–112. ISBN 978-0-7923-6144-2. [Google Scholar]
- Rindi, F.; Cinelli, F. Phenology and Small-scale distribution of some rhodomelacean red algae on a Western Mediterranean Rocky Shore. Eur. J. Phycol. 2000, 35, 115–125. [Google Scholar] [CrossRef]
- Airoldi, L. Effects of disturbance, life histories, and overgrowth on coexistence of algal crusts and turfs. Ecology 2000, 81, 798–814. [Google Scholar] [CrossRef]
- Antoniadou, C.; Voultsiadou, E.; Chintiroglou, C. Seasonal patterns of colonization and early succession on sublittoral rocky cliffs. J. Exp. Mar. Biol. Ecol. 2011, 403, 21–30. [Google Scholar] [CrossRef]
- Falace, A. Evaluation of the influence of inclination of substrate panels on seasonal changes in a macrophytobenthic community. ICES J. Mar. Sci. 2002, 59, S116–S121. [Google Scholar] [CrossRef]
- Burt, J.; Bartholomew, A.; Sale, P.F. Benthic development on large-scale engineered reefs: A comparison of communities among breakwaters of different age and natural reefs. Ecol. Eng. 2011, 37, 191–198. [Google Scholar] [CrossRef]
- Tsiamis, K.; Salomidi, M.; Gerakaris, V.; Mogg, A.O.M.; Porter, E.S.; Sayer, M.D.J.; Küpper, F.C. Macroalgal vegetation on a North European artificial reef (Loch Linnhe, Scotland): Biodiversity, community types and role of abiotic factors. J. Appl. Phycol. 2020, 32, 1353–1363. [Google Scholar] [CrossRef]
- Bailey-Brock, J.H. Fouling community development on an artificial reef in Hawaiian waters. Bull. Mar. Sci. 1989, 42, 580–591. [Google Scholar]
- Palmer-Zwahlen, M.L.; Aseltine, D.A. Successional development of the turf community on a Quarry rock artificial reef. Bull. Mar. Sci. 1994, 55, 902–923. [Google Scholar]
- Casoli, E.; Ricci, S.; Antonelli, F.; Sacco Perasso, C.; Ardizzone, G.; Gravina, M.F. Colonization dynamic on experimental limestone substrata: The role of encrusting epilithics favouring boring polychaetes. Hydrobiologia 2019, 842, 101–112. [Google Scholar] [CrossRef]
- Ponti, M.; Fava, F.; Perlini, R.A.; Giovanardi, O.; Abbiati, M. Benthic assemblages on artificial reefs in the Northwestern Adriatic sea: Does structure type and age matter? Mar. Environ. Res. 2015, 104, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Antoniadou, C.; Voultsiadou, E.; Chintiroglou, C. Benthic colonization and succession on temperate sublittoral rocky cliffs. J. Exp. Mar. Biol. Ecol. 2010, 382, 145–153. [Google Scholar] [CrossRef]
- Choi, D.R. Ecological succession of reef cavity-dwellers (Coelobites) in Coral Rubble. Bull. Mar. Sci. 1984, 31, 72–79. [Google Scholar]
- Gili, J.M.; Ballesteros, E. Structure of Cnidarian populations in Mediterranean sublittoral benthic communities as a result of adaptation to different environmental conditions. Oecologia Aquat. 1991, 10, 243–254. [Google Scholar]
- Harms, J.; Anger, K. Seasonal, annual, and spatial variation in the development of hard bottom communities. Helgoländer Meeresunters. 1983, 36, 137–150. [Google Scholar] [CrossRef][Green Version]
- Fariñas-Franco, J.M.; Roberts, D. Early faunal successional patterns in artificial reefs used for restoration of impacted biogenic habitats. Hydrobiologia 2014, 727, 75–94. [Google Scholar] [CrossRef]
- Toledo, M.-I.; Torres, P.; Díaz, C.; Zamora, V.; López, J.; Olivares, G. Ecological succession of benthic organisms on niche-type artificial reefs. Ecol. Process. 2020, 9, 38. [Google Scholar] [CrossRef]
- Dean, T.A.; Hurd, L.E. Development in an estuarine fouling community: The influence of early colonists on later arrivals. Oecologia 1980, 46, 295–301. [Google Scholar] [CrossRef]
- Herbert, R.J.H.; Collins, K.; Mallinson, J.; Hall, A.E.; Pegg, J.; Ross, K.; Clarke, L.; Clements, T. Epibenthic and mobile species colonisation of a geotextile artificial surf reef on the south coast of England. PLoS ONE 2017, 12, e0184100. [Google Scholar] [CrossRef] [PubMed]
- Bastida, R.; Grosso, M.; Elkin, D. The role of benthic communities and environmental agents in the formation of underwater archeological sites. In Underwater and maritime archaeology in Latin America and the Caribbean; Leshikar-Denton, M.E., Luna Erreguerena, P., Eds.; One world archaeology series; Left Coast Press: Walnut Creek, CA, USA, 2008; pp. 173–185. ISBN 978-1-59874-262-6. [Google Scholar]
- Connell, J.H.; Slatyer, R.O. Mechanisms of succession in natural communities and their role in community stability and organization. Am. Nat. 1977, 111, 1119–1144. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Albouy, C.; Ben Rais Lasram, F.; Cheung, W.W.L.; Christensen, V.; Karpouzi, V.S.; Guilhaumon, F.; Mouillot, D.; Paleczny, M.; et al. The Mediterranean sea under Siege: Spatial overlap between marine biodiversity, cumulative threats and marine reserves. Glob. Ecol. Biogeogr. 2012, 21, 465–480. [Google Scholar] [CrossRef]
- Cahill, A.E.; Pearman, J.K.; Borja, A.; Carugati, L.; Carvalho, S.; Danovaro, R.; Dashfield, S.; David, R.; Féral, J.-P.; Olenin, S.; et al. A Comparative analysis of metabarcoding and morphology-based identification of benthic communities across different regional. Seas Ecol. Evol. 2018, 8, 8908–8920. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Rittschof, D. Love at first taste: Induction of larval settlement by marine microbes. Int. J. Mol. Sci. 2020, 21, 731. [Google Scholar] [CrossRef]
- Olabarria, C. Role of colonization in spatio-temporal patchiness of microgastropods in coralline turf habitat. J. Exp. Mar. Biol. Ecol. 2002, 274, 121–140. [Google Scholar] [CrossRef][Green Version]
- Underwood, A.J.; Chapman, M.G. Early development of subtidal macrofaunal assemblages: Relationships to period and timing of colonization. J. Exp. Mar. Biol. Ecol. 2006, 330, 221–233. [Google Scholar] [CrossRef]
- Antoniadou, C.; Koutsoubas, D.; Chintiroglou, C.C. Mollusca Fauna from Infralittoral Hard Substrate Assem- Blages in the North Aegean Sea. Belg. J. Zool. 2005, 135, 119–126. [Google Scholar]
- Poursanidis, D.; Koutsoubas, D.; Arvanitidis, C.; Chatzigeorgiou, G. ReefMedMol: Mollusca from the Infralittoral Rocky Shores—The biocoenosis of photophilic algae—In the Mediterranean Sea. Biodivers. Data J. 2016, 4, e7516. [Google Scholar] [CrossRef]
- Poursanidis, D.; Chatzigeorgiou, G.; Dimitriadis, C.; Koutsoubas, D.; Arvanitidis, C. Testing the robustness of a coastal biodiversity data protocol in the Mediterranean: Insights from the molluskan assemblages from the sublittoral macroalgae communities. Hydrobiologia 2019, 826, 159–172. [Google Scholar] [CrossRef]
- Russo, G.F.; Fraschetti, S.; Terlizzi, A. Population ecology and production of Bittium latreillii (Gastropoda, Cerithidae) in a Posidonia Oceanica seagrass bed. Ital. J. Zool. 2002, 69, 215–222. [Google Scholar] [CrossRef]
- Arvanitidis, C.; Bellan, G.; Drakopoulos, P.; Valavanis, V.; Dounas, C.; Koukouras, A.; Eleftheriou, A. Seascape biodiversity patterns along the mediterranean and the Black sea: Lessons from the biogeography of benthic polychaetes. Mar. Ecol. Prog. Ser. 2002, 244, 139–152. [Google Scholar] [CrossRef]
- Giangrande, A.; Gravina, M.F. Brackish-water polychaetes, good descriptors of environmental changes in space and time. Transit. Waters Bull. 2015, 9, 42–55. [Google Scholar] [CrossRef]
- Çinar, M.E.; Katagan, T.; Öztürk, B.; Bakir, K.; Dagli, E.; Açik, S.; Dogan, A.; Bıtlıs, B. Spatio-temporal distributions of zoobenthos in soft substratum of izmir bay (Aegean sea, Eastern Mediterranean), with special emphasis on alien species and ecological quality status. J. Mar. Biol. Assoc. UK 2012, 92, 1457–1477. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Cabezas, M.P.; Baeza-Rojano, E.; García-Gómez, J.C. Spatial patterns and seasonal fluctuations of intertidal macroalgal assemblages from Tarifa Island, Southern Spain: Relationship with associated crustacea. J. Mar. Biol. Assoc. UK 2011, 91, 107–116. [Google Scholar] [CrossRef]
- Keklikoglou, K.; Chatzigeorgiou, G.; Faulwetter, S.; Kalogeropoulou, V.; Plaiti, W.; Maidanou, M.; Dounas, C.; Lampadariou, N.; Arvanitidis, C. ‘Simple’ can be good, too: Testing three hard bottom sampling methods on macrobenthic and meiobenthic assemblages. J. Mar. Biol. Assoc. UK 2019, 99, 777–784. [Google Scholar] [CrossRef]
- Azov, Y. Eastern Mediterranean—A Marine desert? Mar. Pollut. Bull. 1991, 23, 225–232. [Google Scholar] [CrossRef]
- Voultsiadou, E. Demosponge distribution in the Eastern Mediterranean: A NW–SE Gradient. Helgol. Mar. Res. 2005, 59, 237–251. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Azzola, A.; Parravicini, V.; Peirano, A.; Morri, C.; Montefalcone, M. Abrupt change in a subtidal rocky reef community coincided with a rapid acceleration of sea water warming. Diversity 2019, 11, 215. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sessile Assemblages | Motile Assemblages | ||
|---|---|---|---|
| Sampling Periods | Average Dissimilarity | Sampling Periods | Average Dissimilarity |
| 2017–2019 | 52.37 | 2017–2019 | 61.52 |
| 2017–2020 | 54.71 | 2017–2020 | 74.45 |
| 2019–2020 | 39.2 | 2019–2020 | 67.83 |
| Major Taxonomic Group | 2017 | 2019 | 2020 | |
|---|---|---|---|---|
| Average Similarity | 56.12 | 63.99 | 64.47 | |
| Sessile Assemblages | Taxonomic Group/Species | |||
| Macroalgae | Turf-forming algae | 34.43 | 49.25 | 39.23 |
| Encrusting macroalgae | 7.31 | 12.92 | 16.46 | |
| Mucilaginous algae | 19.82 | - | - | |
| Acetabularia sp. | 5.56 | - | - | |
| Dictyota implexa | 3.87 | - | - | |
| Cnidaria | Hydrozoa | 4.97 | 16.92 | 17.36 |
| Polychaeta | Other Serpulidae | 17.77 | 16.90 | 12.71 |
| Protula spp. | - | - | 5.66 | |
| Average Similarity | 53.87 | 40.58 | 33.77 | |
| Motile assemblages | Family | |||
| Polychaeta | Capitellidae | - | 6.42 | - |
| Chrysopetalidae | 6.22 | 13.74 | 7.78 | |
| Dorvilleidae | - | 4.13 | - | |
| Eunicidae | 3.28 | - | - | |
| Nereididae | 3.39 | - | 2.57 | |
| Paraonidae | 4.06 | - | - | |
| Syllidae | 4.13 | 8.65 | - | |
| Mollusca | Cerithiidae | 24.92 | 36.59 | 73.86 |
| Rissoidae | 7.78 | 9.19 | - | |
| Trochidae | - | - | - | |
| Crustacea | Aoridae | 9.41 | 7.89 | - |
| Bodotriidae | 4.20 | - | - | |
| Caprellidae | 7.78 | - | - | |
| Dexaminidae | 10.50 | - | - | |
| Lysianassidae | 6.09 | - | - | |
| Pasiphaeidae | - | 4.24 | 7.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rallis, I.; Chatzigeorgiou, G.; Florido, M.; Sedano, F.; Procopiou, A.; Chertz-Bynichaki, M.; Vernadou, E.; Plaiti, W.; Koulouri, P.; Dounas, C.; et al. Early Succession Patterns of Benthic Assemblages on Artificial Reefs in the Oligotrophic Eastern Mediterranean Basin. J. Mar. Sci. Eng. 2022, 10, 620. https://doi.org/10.3390/jmse10050620
Rallis I, Chatzigeorgiou G, Florido M, Sedano F, Procopiou A, Chertz-Bynichaki M, Vernadou E, Plaiti W, Koulouri P, Dounas C, et al. Early Succession Patterns of Benthic Assemblages on Artificial Reefs in the Oligotrophic Eastern Mediterranean Basin. Journal of Marine Science and Engineering. 2022; 10(5):620. https://doi.org/10.3390/jmse10050620
Chicago/Turabian StyleRallis, Ioannis, Giorgos Chatzigeorgiou, Marta Florido, Francisco Sedano, Avgi Procopiou, Melina Chertz-Bynichaki, Emmanouela Vernadou, Wanda Plaiti, Panayota Koulouri, Costas Dounas, and et al. 2022. "Early Succession Patterns of Benthic Assemblages on Artificial Reefs in the Oligotrophic Eastern Mediterranean Basin" Journal of Marine Science and Engineering 10, no. 5: 620. https://doi.org/10.3390/jmse10050620
APA StyleRallis, I., Chatzigeorgiou, G., Florido, M., Sedano, F., Procopiou, A., Chertz-Bynichaki, M., Vernadou, E., Plaiti, W., Koulouri, P., Dounas, C., Gerovasileiou, V., & Dailianis, T. (2022). Early Succession Patterns of Benthic Assemblages on Artificial Reefs in the Oligotrophic Eastern Mediterranean Basin. Journal of Marine Science and Engineering, 10(5), 620. https://doi.org/10.3390/jmse10050620