
Comparative Study of Marine Cave Communities in a Protected Area of the South-Eastern Aegean Sea, Greece

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
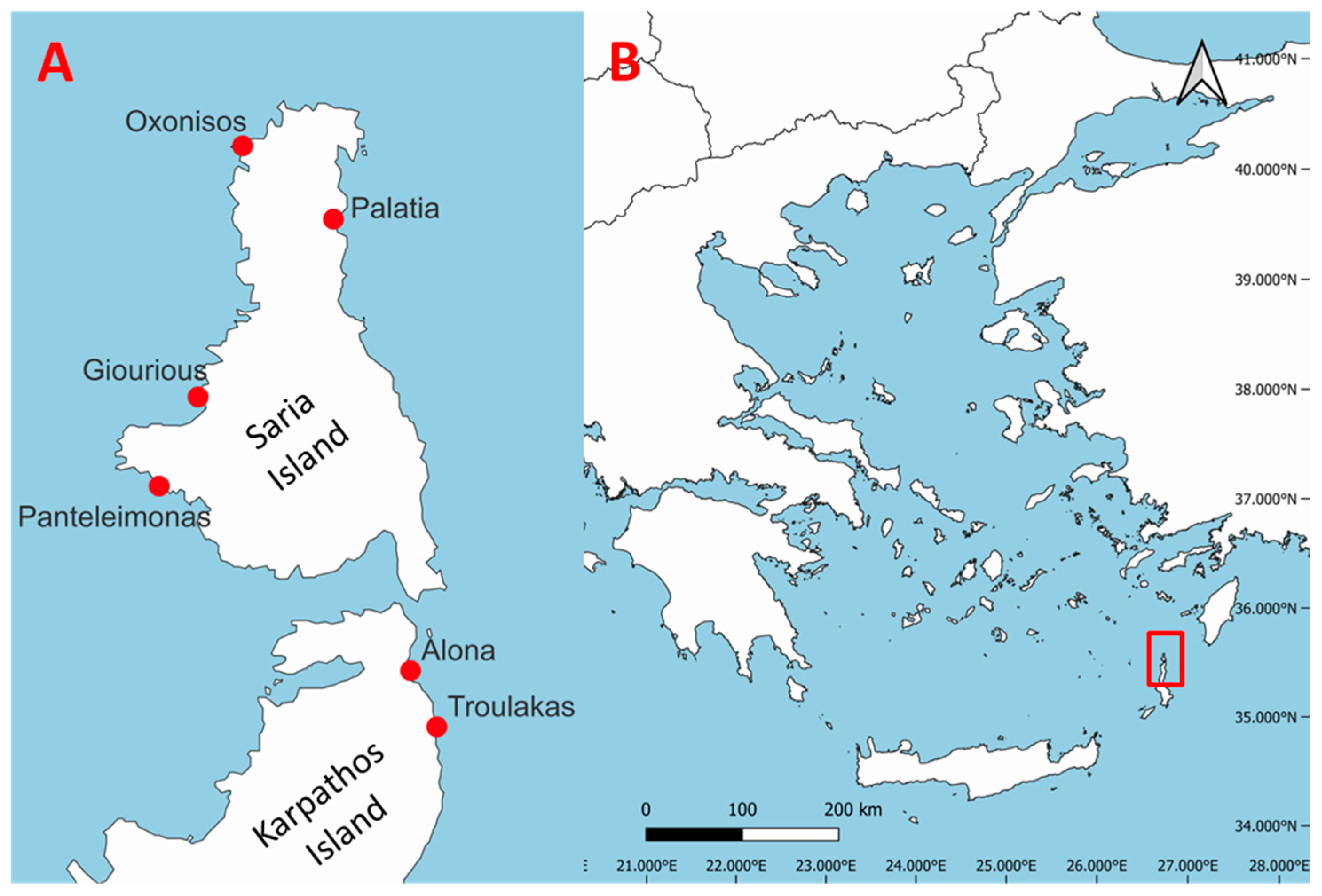
2.1. Study Area
2.2. Sampling
2.3. Photoquadrats Processing
2.4. Ecological Quality Assessment
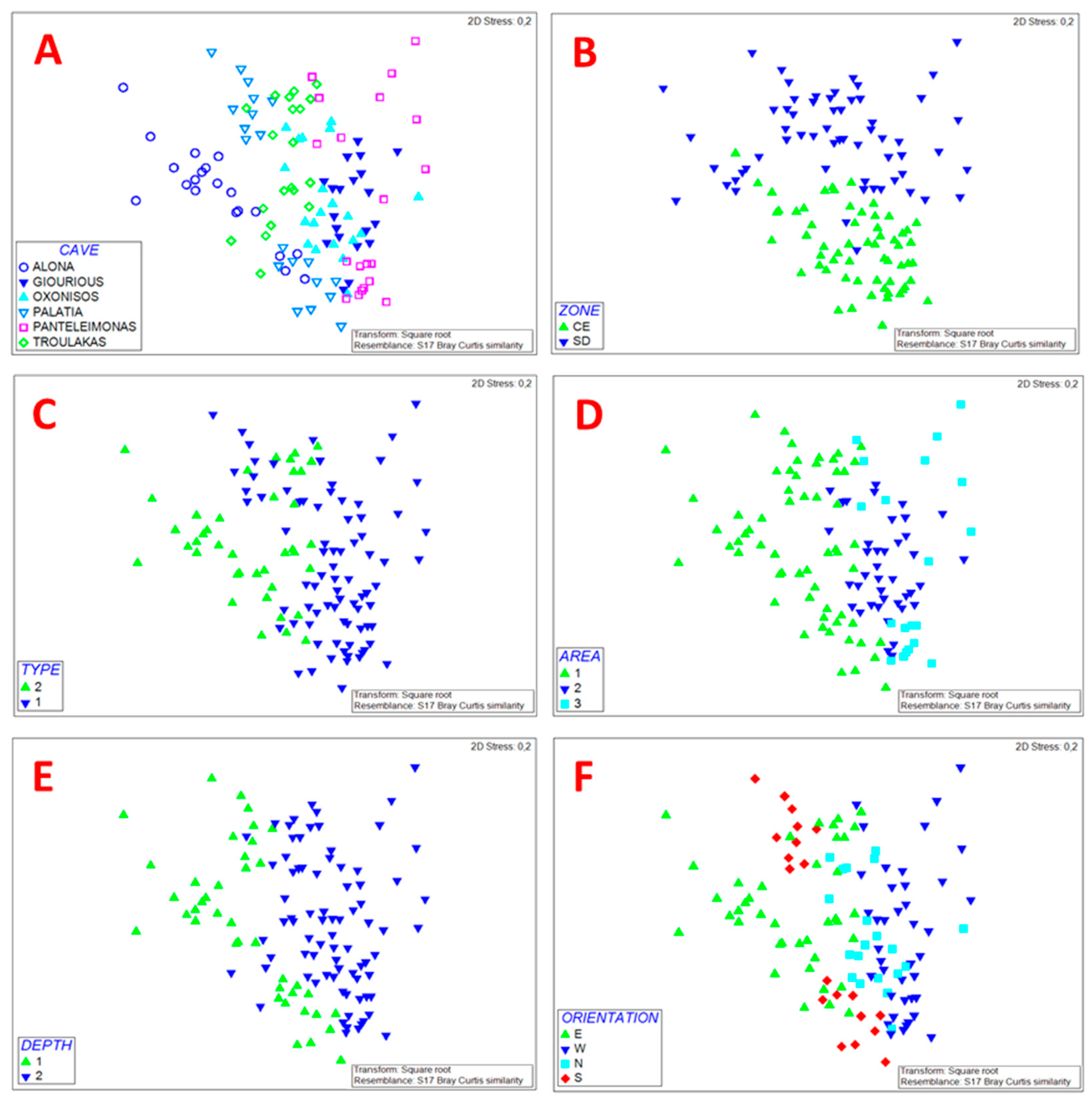
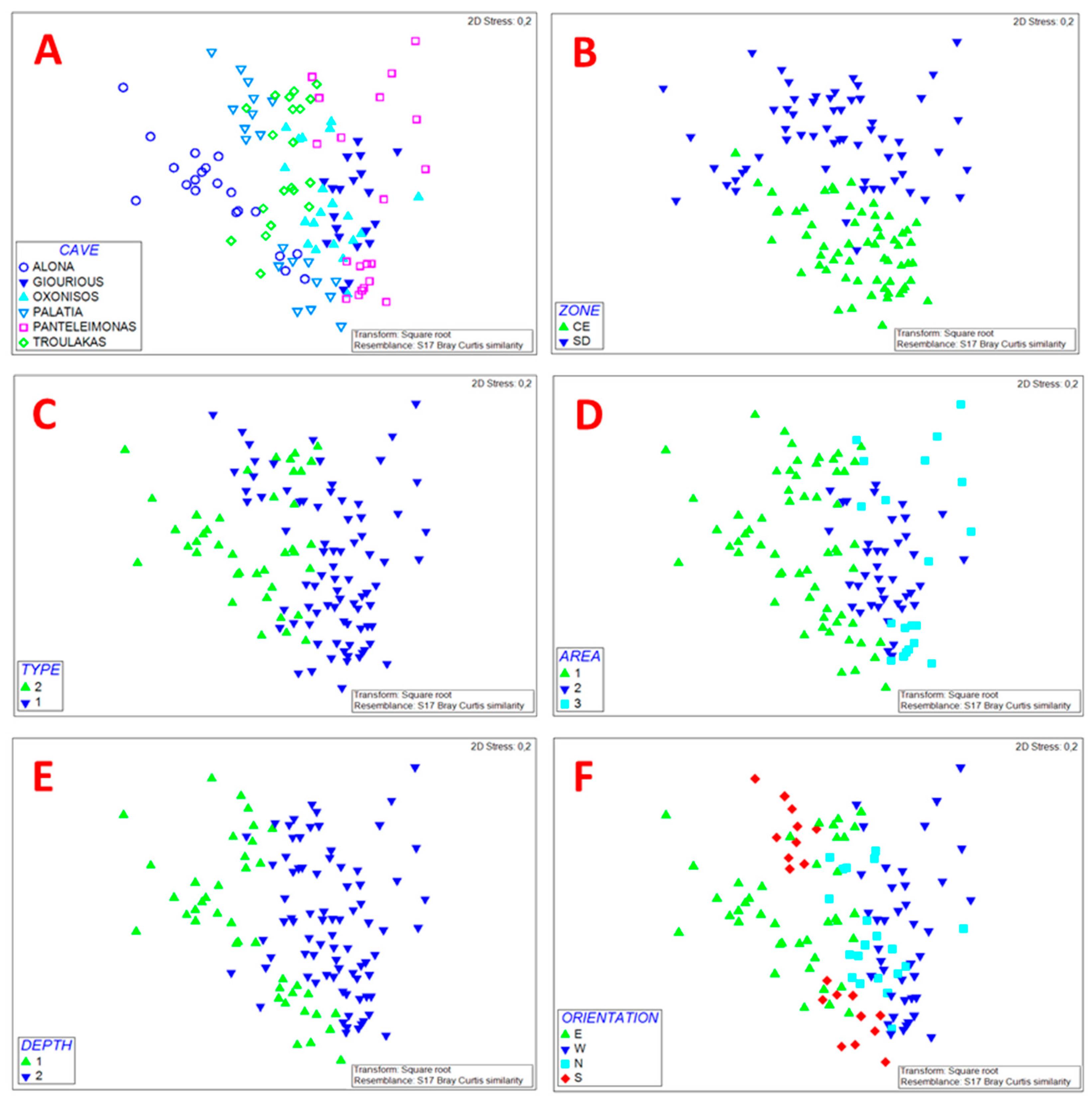
2.5. Structure Assessment and Statistical Analysis
3. Results
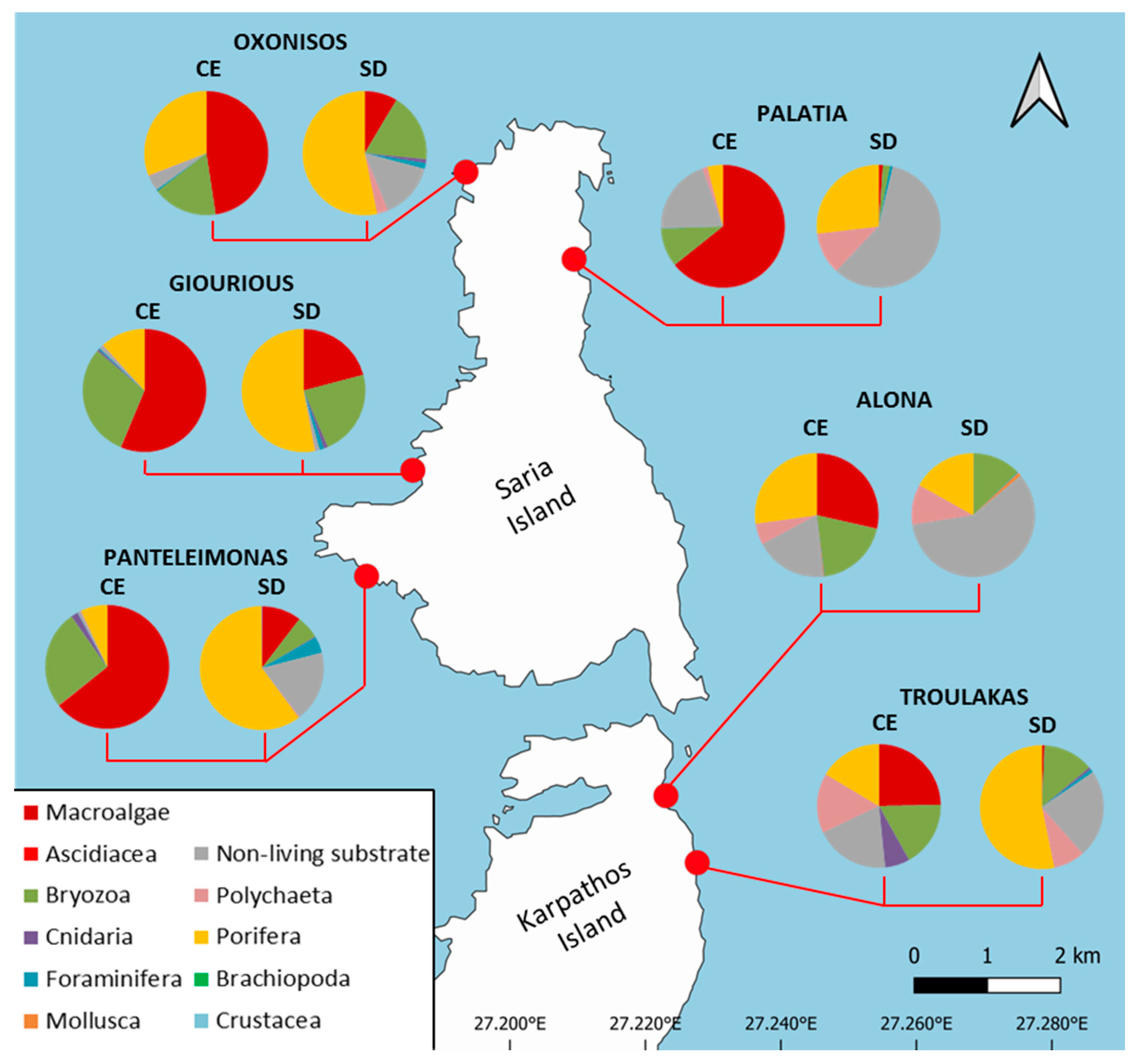
3.1. Sessile Community Structure
3.2. Threats and Pressures on the Sessile Community
3.3. Motile Fauna
3.4. Ecological Quality Assessment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ouerghi, A.; Gerovasileiou, V.; Bianchi, C.N. Mediterranean marine caves: A synthesis of current knowledge and the Mediterranean Action Plan for the conservation of “dark habitats”. In Marine Caves of the Eastern Mediterranean Sea: Biodiversity, Threats and Conservation; Publication no. 53; Öztürk, B., Ed.; Turkish Marine Research Foundation (TUDAV): Istanbul, Turkey, 2019; pp. 1–13. [Google Scholar]
- Gerovasileiou, V.; Bianchi, C. Mediterranean marine caves: A synthesis of current knowledge. Oceanogr. Mar. Biol. Annu. Rev. 2021, 59, 1–88. [Google Scholar] [CrossRef]
- Riedl, R. Biologie der Meereshöhlen; Paul Parey: Hamburg, Germany, 1966; pp. 1–636. [Google Scholar]
- Bussotti, S.; Terlizzi, A.; Fraschetti, S.; Belmonte, G.; Boero, F. Spatial and temporal variability of sessile benthos in shallow Mediterranean marine caves. Mar. Ecol. Progr. Ser. 2006, 325, 109–119. [Google Scholar] [CrossRef]
- Russ, K.; Rützler, K. Zur Kenntnis der Schwammfauna unterseeischer Höhlen. Pubbl. Stn. Zool. Napoli 1959, 30, 756–787. [Google Scholar]
- Sarà, M. Zonazione dei Porifera in biotope litorali. Pubbl. Stn. Zool. Napoli 1962, 32, 44–57. [Google Scholar]
- Pouliquen, L. Les spongiaires des grottes sous-marines de la région de Marseille—Ecologie et systematique. Téthys 1972, 3, 717–758. [Google Scholar]
- Balduzzi, A.; Bianchi, C.N.; Boero, F.; Cattaneo-Vietti, R.; Pansini, M.; Sarà, M. The suspension-feeder communities of a Mediterranean Sea cave. Sci. Mar. 1989, 53, 387–395. [Google Scholar]
- Martí, R.; Uriz, M.J.; Ballesteros, E.; Turon, X. Benthic assemblages in two Mediterranean caves: Species diversity and coverage as a function of abiotic parameters and geographic distance. J. Mar. Biol. Ass. UK 2004, 84, 557–572. [Google Scholar] [CrossRef] [Green Version]
- Gerovasileiou, V.; Dimitriadis, C.; Arvanitidis, C.; Voultsiadou, E. Taxonomic and functional surrogates of sessile benthic diversity in Mediterranean marine caves. PLoS ONE 2017, 12, e0183707. [Google Scholar] [CrossRef] [Green Version]
- Harmelin, J.G.; Vacelet, J.; Vasseur, P. Les grottes sous-marines obscures: Un milieu extrême et un remarquable biotope refuge. Téthys 1985, 11, 214–229. [Google Scholar]
- Gerovasileiou, V.; Aguilar, R.; Marín, P. Guidelines for Inventorying and Monitoring of Dark Habitats in the Mediterranean Sea; SPA/RAC-UNEP/MAP, OCEANA: Tunis, Tunisia, 2017; pp. 1–40. [Google Scholar]
- Giakoumi, S.; Sini, M.; Gerovasileiou, V.; Mazor, T.; Beher, J.; Possingham, H.P.; Abdulla, A.; Çinar, M.E.; Dendrinos, P.; Gucu, A.C.; et al. Ecoregion-based conservation planning in the Mediterranean: Dealing with large-scale heterogeneity. PLoS ONE 2013, 8, e76449. [Google Scholar] [CrossRef] [Green Version]
- Sini, M.; Katsanevakis, S.; Koukourouvli, N.; Gerovasileiou, V.; Dailianis, T.; Buhl-Mortensen, L.; Damalas, D.; Dendrinos, P.; Dimas, X.; Frantzis, A.; et al. Assembling ecological pieces to reconstruct the conservation puzzle of the Aegean Sea. Front. Mar. Sci. 2017, 4, 347. [Google Scholar] [CrossRef]
- Abdulla, A.; Gomei, M.; Maison, E.; Piante, C. Status of Marine Protected Areas in the Mediterranean Sea; International Union for Conservation of Nature (IUCN): Malaga, Spain; World Wide Fund for Nature (WWF): Paris, France, 2008; pp. 1–152. [Google Scholar]
- Petricioli, D.; Buzzacott, P.; Radolović, M.; Bakran-Petricioli, T.; Gerovasileiou, V. Visitation and conservation of marine caves. In Proceedings of the International Symposium “Inside and outside the Mountain”, Custonaci, Sicily, Italy, 23–26 April 2015; pp. 29–30. [Google Scholar]
- Chevaldonné, P.; Lejeusne, C. Regional warming-induced species shift in north-west Mediterranean marine caves. Ecol. Lett. 2003, 6, 371–379. [Google Scholar] [CrossRef]
- Parravicini, V.; Guidetti, P.; Morri, C.; Montefalcone, M.; Donato, M.; Bianchi, C.N. Consequences of sea water temperature anomalies on a Mediterranean submarine cave ecosystem. Estuar. Coast. Shelf Sci. 2010, 86, 276–282. [Google Scholar] [CrossRef]
- Costa, G.; Betti, F.; Nepote, E.; Cattaneo-Vietti, R.; Pansini, M.; Bavestrello, G.; Bertolino, M. Sponge community variations within two semi-submerged caves of the Ligurian Sea (Mediterranean Sea) over a half-century time span. Eur. Zool. J. 2018, 85, 381–391. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Dailianis, T.; Sini, M.; del Mar Otero, M.; Numa, C.; Katsanevakis, S.; Voultsiadou, E. Assessing the regional conservation status of sponges (Porifera): The case of the Aegean ecoregion. Mediterr. Mar. Sci. 2018, 19, 526–537. [Google Scholar] [CrossRef] [Green Version]
- Sempere-Valverde, J.; Sabino Lorenzo, Á.; Espinosa, F.; Gerovasileiou, V.; Sánchez-Tocino, L.; Navarro-Barranco, C. Taxonomic and morphological descriptors reveal high benthic temporal variability in a Mediterranean marine submerged cave over a decade. Hydrobiologia 2019, 839, 177–194. [Google Scholar] [CrossRef]
- Rastorgueff, P.A.; Bellan-Santini, D.; Bianchi, C.N.; Bussotti, S.; Chevaldonné, P.; Guidetti, P.; Harmelin, J.G.; Montefalcone, M.; Morri, C.; Perez, T.; et al. An ecosystem-based approach to evaluate the ecological quality of Mediterranean undersea caves. Ecol. Indic. 2015, 54, 137–152. [Google Scholar] [CrossRef]
- Nepote, E.; Bianchi, C.N.; Morri, C.; Ferrari, M.; Montefalcone, M. Impact of a harbour construction on the benthic community of two shallow marine caves. Mar. Pollut. Bull. 2017, 114, 35–45. [Google Scholar] [CrossRef]
- Montefalcone, M.; De Falco, G.; Nepote, E.; Canessa, M.; Bertolino, M.; Bavestrello, G.; Morri, C.; Bianchi, C.N. Thirty year ecosystem trajectories in a submerged marine cave under changing pressure regime. Mar. Environ. Res. 2018, 137, 98–110. [Google Scholar] [CrossRef]
- Di Franco, A.; Milazzo, M.; Baiata, P.; Tomasello, A.; Chemello, R. Scuba diver behaviour and its effects on the biota of a Mediterranean marine protected area. Environ. Conserv. 2009, 36, 32–40. [Google Scholar] [CrossRef]
- Guarnieri, G.; Terlizzi, A.; Bevilacqua, S.; Fraschetti, S. Increasing heterogeneity of sensitive assemblages as a consequence of human impact in submarine caves. Mar. Biol. 2012, 159, 1155–1164. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Chintiroglou, C.; Vafidis, D.; Koutsoubas, D.; Sini, M.; Dailianis, T.; Issaris, Y.; Akritopoulou, E.; Dimarchopoulou, D.; Voutsiadou, E. Census of biodiversity in marine caves of the eastern Mediterranean Sea. Mediterr. Mar. Sci. 2015, 16, 245–265. [Google Scholar] [CrossRef] [Green Version]
- Gerovasileiou, V.; Voultsiadou, E.; Issaris, Y.; Zenetos, A. Alien biodiversity in Mediterranean marine caves. Mar. Ecol. 2016, 37, 239–256. [Google Scholar] [CrossRef]
- Ragkousis, M.; Abdelali, N.; Azzurro, E.; Badreddine, A.; Bariche, M.; Bitar, G.; Crocetta, F.; Denitto, F.; Digenis, M.; El Zrelli, R.; et al. New Alien Mediterranean Biodiversity Records (October 2020). Mediterr. Mar. Sci. 2020, 21, 631–652. [Google Scholar]
- Digenis, M.; Ragkousis, M.; Vasileiadou, K.; Gerovasileiou, V.; Katsanevakis, S. New records of the Indo-Pacific shrimp Urocaridella pulchella Yokes & Galil, 2006 from the Eastern Mediterranean Sea. Bioinvasions Rec. 2021, 10, 295–303. [Google Scholar] [CrossRef]
- Dailianis, T.; Gerovasileiou, V.; Chatzigeorgiou, G.; Vernadou, E.; Glampedakis, I.; Arvanitidis, C.; Protopappas, K.; Piligotsi, G.; Kargiolaki, H. An extended baseline survey of cave habitats within a marine protected area in the Aegean Sea. In Proceedings of the 6th European Conference on Scientific Diving, Freiberg, Germany, 21–22 April 2021. [Google Scholar]
- Gerovasileiou, V.; Voultsiadou, Ε. Sponge diversity gradients in marine caves of the eastern Mediterranean. J. Mar. Biol. Assoc. UK 2016, 96, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Dimarchopoulou, D.; Gerovasileiou, V.; Voultsiadou, E. Spatial variability of sessile benthos in a semi-submerged marine cave of a remote Aegean Island (eastern Mediterranean Sea). Reg. Stud. Mar. Sci. 2018, 17, 102–111. [Google Scholar] [CrossRef]
- Pérès, J.M. Mediterranean Benthos. Oceanogr. Mar. Biol. Annu. Rev. 1967, 5, 449–533. [Google Scholar]
- Trygonis, V.; Sini, M. photoQuad: A dedicated seabed image processing software, and a comparative error analysis of four photoquadrat methods. J. Exp. Mar. Biol. Ecol. 2012, 424–425, 99–108. [Google Scholar] [CrossRef]
- Bacchiocchi, F.; Airoldi, L. Distribution and dynamics of epibiota on hard structures for coastal protection. Estuar. Coast. Shelf Sci. 2003, 56, 1157–1166. [Google Scholar] [CrossRef]
- Marzinelli, E.M.; Underwood, A.J.; Coleman, R.A. Modified habitats influence kelp epibiota via direct and indirect effects. PLoS ONE 2011, 6, e21936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostalé-Valriberas, E.; Sempere-Valverde, J.; Coppa, S.; García-Gómez, J.C.; Espinosa, F. Creation of microhabitats (tidepools) in ripraps with climax communities as a way to mitigate negative effects of artificial substrate on marine biodiversity. Ecol. Eng. 2018, 120, 522–531. [Google Scholar] [CrossRef]
- Sedano, F.; Pavón-Paneque, A.; Navarro-Barranco, C.; Guerra-García, J.M.; Digenis, M.; Sempere-Valverde, J.; Espinosa, F. Coastal armouring affects intertidal biodiversity across the Alboran Sea (Western Mediterranean Sea). Mar. Environ. Res. 2021, 171, 105475. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006; pp. 1–192. [Google Scholar]
- Clarke, K.; Warwick, R. A Further Biodiversity Index Applicable to Species Lists: Variation in Taxonomic Distinctness. Mar. Ecol. Prog. Ser. 2001, 216, 265–278. [Google Scholar] [CrossRef]
- Mead, A. Review of the development of multidimensional scaling methods. J. R. Stat. Soc. Ser. D 1992, 41, 27–39. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Galani, A.; Dailianis, T.; Sini, M.; Katsanevakis, S.; Gerovasileiou, V. Characterization of benthic communities in marine caves of the island-dominated Aegean Sea (Eastern Mediterranean). In Proceedings of the 2nd Mediterranean Symposium on the Conservation of Dark Habitats, Antalya, Turkey, 16 January 2019; pp. 67–68. [Google Scholar]
- Morri, C.; Bianchi, C.N. Zonazione biologica. In Grotte Marine: Cinquant’anni di Ricerca in Italia; Cicogna, F., Bianchi, C.N., Ferrari, G., Forti, P., Eds.; Ministero dell’Ambiente e della Tutela del Territorio: Roma, Italy, 2003; pp. 257–265. [Google Scholar]
- Radolović, M.; Bakran-Petricioli, T.; Petricioli, D.; Surić, M.; Perica, D. Biological response to geochemical and hydrological processes in a shallow submarine cave. Mediterr. Mar. Sci. 2015, 16, 305–324. [Google Scholar] [CrossRef]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Cave features, seasonality and subterranean distribution of non-obligate cave dwellers. PeerJ 2017, 5, e3169. [Google Scholar] [CrossRef]
- Rosso, A.; Gerovasileiou, V.; Sanfilippo, R.; Guido, A. Bryozoan assemblages from two submarine caves in the Aegean Sea (Eastern Mediterranean). Mar. Biodiv. 2018, 49, 707–726. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Pronzato, R.; Cattaneo-Vietti, R.; Benedetti-Cecchi, L.; Morri, C.; Pansini, M.; Chemello, R.; Milazzo, M.; Fraschetti, S.; Terlizzi, A.; et al. Mediterranean marine benthos: A manual of methods for its sampling and study. Hard bottoms. Biol. Mar. Mediterr. 2004, 11, 185–215. [Google Scholar]
- Zapata-Ramírez, P.A.; Scaradozzi, D.; Sorbi, L.; Palma, M.; Pantaleo, U.; Ponti, M.; Cerrano, C. Innovative study methods for the Mediterranean coralligenous habitats. Adv. Oceanogr. Limnol. 2013, 4, 102–119. [Google Scholar] [CrossRef]
- Lage, A.; Araujo, H.P.M.; Gerovasileiou, V.; Muricy, G. A new cave-dwelling species of Plakina (Porifera: Homoscleromorpha) from Crete, Greece (South Aegean Sea). Zootaxa 2018, 4466, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Lage, A.; Gerovasileiou, V.; Voultsiadou, E.; Muricy, G. Taxonomy of Plakina (Porifera: Homoscleromorpha) from Aegean submarine caves, with descriptions of three new species and new characters for the genus. Mar. Biodivers. 2019, 49, 727–747. [Google Scholar] [CrossRef]
- Bitner, M.A.; Gerovasileiou, V. Taxonomic composition and assemblage structure of brachiopods from two submarine caves in the Aegean Sea, Eastern Mediterranean. Eur. Zool. J. 2021, 88, 316–327. [Google Scholar] [CrossRef]
- Taviani, M.; Angeletti, L.; Campiani, E.; Ceregato, A.; Foglini, F.; Maselli, V.; Morsilli, M.; Parise, M.; Trincardi, F. Drowned karst landscape offshore the Apulian margin (Southern Adriatic Sea, Italy). J. Cave Karst Stud. 2012, 74, 197–212. [Google Scholar] [CrossRef]
- Novosel, M.; Bakran-Petricioli, T.; Požar-Domac, A.; Kružić, P.; Radić, I. The benthos of the northern part of the Velebit Channel (Adriatic Sea, Croatia). Nat. Croat. 2002, 11, 387–409. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. Available online: www.fishbase.org (accessed on 11 April 2022).
- Ragkousis, M.; Digenis, M.; Kovačić, M.; Katsanevakis, S.; Gerovasileiou, V. Rarely Reported Cryptobenthic Fish in Marine Caves of the Eastern Mediterranean Sea. J. Mar. Sci. Eng. 2021, 9, 557. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Gerovasileiou, V.; Morri, C.; Froglia, C. Distribution and Ecology of Decapod Crustaceans in Mediterranean Marine Caves: A Review. Diversity 2022, 14, 176. [Google Scholar] [CrossRef]
- MOm. Status Report of the Mediterranean Monk Seal Populations in Kimolos-Polyaigos and Karpathos Saria Areas; Final Report to EC LIFE-NATURE 2000 Project (LIFE00NAT/GR/7248); Hellenic Society for the Study and Protection of the Monk Seal (MOm): Athens, Greece, 2005. [Google Scholar]
- Sanfilippo, R.; Rosso, A.; Guido, A.; Mastandrea, A.; Russo, F.; Riding, R.; Taddei Ruggiero, E. Metazoan/microbial biostalactites from present-day submarine caves in the Mediterranean Sea. Mar. Ecol. 2015, 36, 1277–1293. [Google Scholar] [CrossRef]
- Guido, A.; Jimenez, C.; Achilleos, K.; Rosso, A.; Sanfilippo, R.; Hadjioannou, L.; Petrou, A.; Russo, F.; Mastandrea, A. Cryptic serpulid-microbialite bioconstructions in the Kakoskali submarine cave (Cyprus, Eastern Mediterranean). Facies 2017, 63, 21. [Google Scholar] [CrossRef]
- Rosso, A.; Sanfilippo, R.; Guido, A.; Gerovasileiou, V.; Taddei Ruggiero, E.; Belmonte, G. Colonisers of the dark: Biostalactite-associated metazoans from “lu Lampiùne” submarine cave (Apulia, Mediterranean Sea). Mar. Ecol. 2021, 42, e12634. [Google Scholar] [CrossRef]
- Kazanidis, G.; Guido, A.; Rosso, A.; Sanfilippo, R.; Roberts, J.M.; Gerovasileiou, V. One on Top of the Other: Exploring the Habitat Cascades Phenomenon in Iconic Biogenic Marine Habitats. Diversity 2022, 14, 290. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Zenetos, A.; Belchior, C.; Cardoso, A.C. Invading European Seas: Assessing pathways of introduction of marine aliens. Ocean Coast. Manag. 2013, 76, 64–74. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Zenetos, A.; Corsini-Foka, M.; Tsiamis, K. Biological Invasions in the Aegean Sea: Temporal Trends, Pathways, and Impacts; The Handbook of Environmental Chemistry; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar] [CrossRef]
- García-Gómez, J.C.; González, A.R.; Maestre, M.J.; Espinosa, F. Detect coastal disturbances and climate change effects in coralligenous community through sentinel stations. PLoS ONE 2020, 15, e0231641. [Google Scholar] [CrossRef] [PubMed]
- Di Camillo, C.G.; Cerrano, C. Mass Mortality Events in the NW Adriatic Sea: Phase Shift from Slow- to Fast-Growing Organisms. PLoS ONE 2015, 10, e0126689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idan, T.; Goren, L.; Shefer, S.; Ilan, M. Sponges in a changing climate: Survival of Agelas oroides in a warming Mediterranean Sea. Front. Mar. Sci. 2020, 7, 1064. [Google Scholar] [CrossRef]
- Garrabou, J.; Gómez-Gras, D.; Ledoux, J.B.; Linares, C.; Bensoussan, N.; López-Sendino, P.; Bazairi, H.; Espinosa, F.; Ramdani, M.; Grimes, S.; et al. Collaborative Database to Track Mass Mortality Events in the Mediterranean Sea. Front. Mar. Sci. 2019, 6, 707. [Google Scholar] [CrossRef] [Green Version]
- Mačić, V.; Ðorđević, N.; Petović, S.; Malovrazić, N.; Bajković, M. Typology of marine litter in “Papuča” (Slipper) cave (Montenegro, South Adriatic Sea). Stud. Mar. 2018, 31, 38–43. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Voultsiadou, E. Marine Caves of the Mediterranean Sea: A Sponge Biodiversity Reservoir within a Biodiversity Hotspot. PLoS ONE 2012, 7, e39873. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Tsirintanis, K.; Sini, M.; Gerovasileiou, V.; Koukourouvli, N. Aliens in the Aegean—A sea under siege (ALAS). Res. Ideas Outcomes 2020, 6, e53057. [Google Scholar] [CrossRef]
- Mammola, S.; Meierhofer, M.B.; Borges, P.A.V.; Colado, R.; Culver, D.C.; Deharveng, L.; Delić, T.; Di Lorenzo, T.; Dražina, T.; Ferreira, R.L.; et al. Toward evidence-based conservation of subterranean ecosystems. Biol. Rev. 2022. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; Manenti, R. Cave communities: From the surface border to the deep darkness. Diversity 2020, 12, 167. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
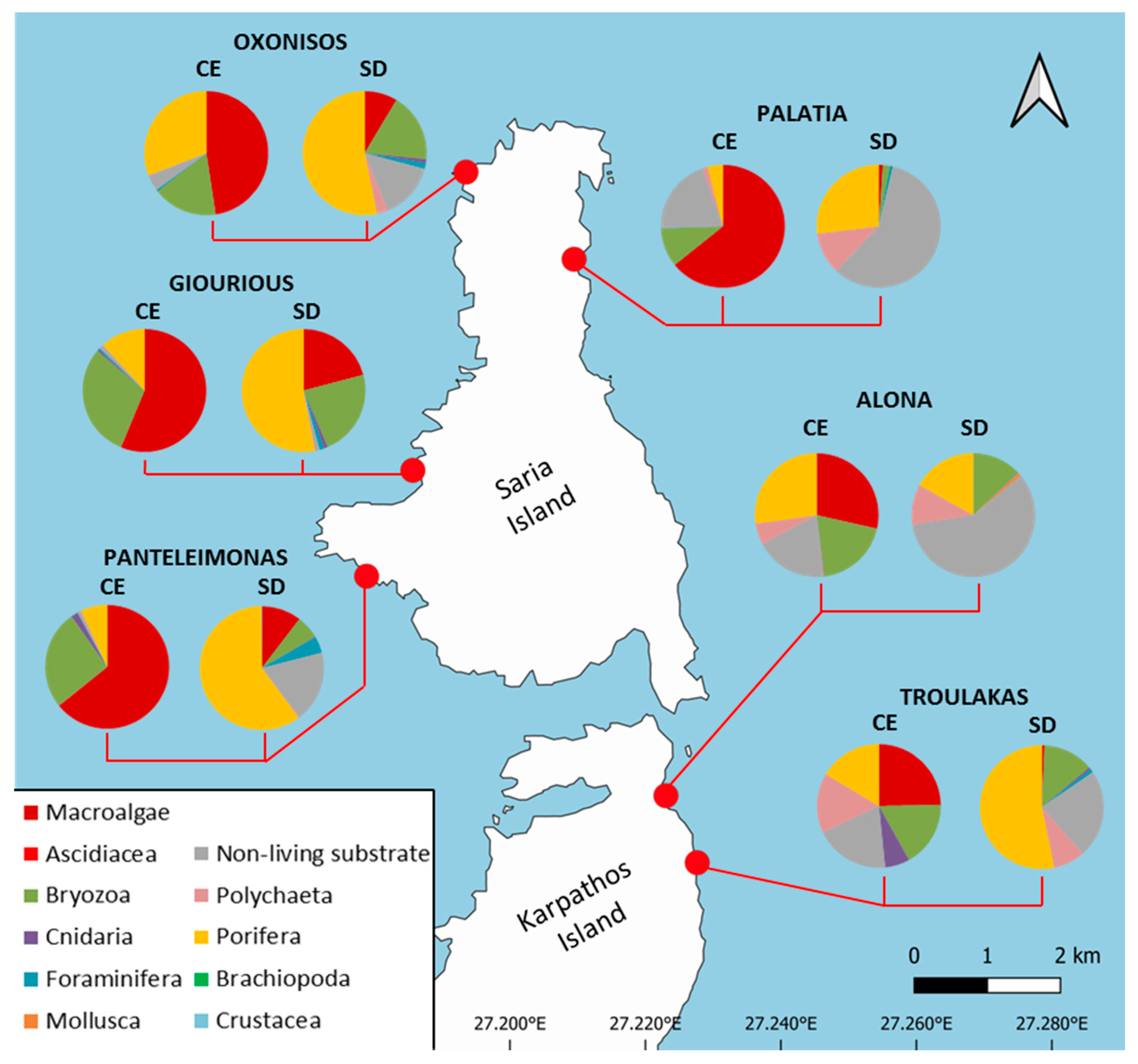
| Cave Name | Latitude (DD) | Longitude (DD) | Type | Depth (m) | Length (m) | Entrance Width (m) | Entrance Orientation |
|---|---|---|---|---|---|---|---|
| Alona | 35.81969 | 27.23049 | Sub. | 1–4 | 24 | 11 | E |
| Troulakas | 35.81072 | 27.23314 | Sub. | 8–11 | 16 | 13 | E |
| Giourious | 35.86615 | 27.20269 | Semi-sub. | 0–14 | 17 | 10 | W |
| Oxonisos | 35.90087 | 27.21935 | Semi-sub. | 0–16 | 17 | 10 | N |
| Palatia | 35.88730 | 27.23276 | Semi-sub. | 0–9 | 17 | 4 | S |
| Panteleimonas | 35.85458 | 27.19273 | Semi-sub. | 0–15 | 37 | 30 | W |
| Taxa/categories | ALO | GIOU | OXO | PAL | PANT | TROU | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CE | SD | CE | SD | CE | SD | CE | SD | CE | SD | CE | SD | |
| Macroalgae | 28.5 | - | 56.2 | 20.9 | 47.6 | 8.6 | 64.2 | 1.1 | 64.1 | 10.4 | 24.7 | 0.7 |
| Foraminifera | 0.1 | 0.2 | 0.4 | 1.2 | 4.0 | 1.5 | 1.0 | 0.7 | 0.1 | 4.3 | - | 0.9 |
| Porifera | 27.2 | 16.9 | 11.7 | 52.9 | 30.8 | 53.2 | 4.0 | 26.9 | 6.9 | 59.9 | 16.4 | 53.2 |
| Cnidaria | 0.1 | - | 0.5 | 1.0 | - | 1.0 | - | - | 1.8 | 0.2 | 6.5 | 0.6 |
| Polychaeta | 5.6 | 10.6 | 0.2 | 0.2 | 3.0 | 3.0 | 1.4 | 11.1 | 0.3 | 0.4 | 15.7 | 8.3 |
| Mollusca | 0.3 | 1.0 | - | - | - | - | - | - | - | - | - | - |
| Crustacea | - | - | - | - | - | - | - | - | - | 0.2 | - | - |
| Bryozoa | 19.3 | 12.8 | 29.9 | 22.7 | 16.9 | 18.0 | 10.2 | 1.9 | 25.9 | 6.1 | 17.2 | 13.2 |
| Ascidiacea | 0.1 | - | 0.1 | - | - | - | - | - | 0.2 | - | - | - |
| NLS | 18.8 | 58.5 | 1.0 | 1.1 | 4.0 | 14.7 | 20.1 | 58.3 | 0.7 | 18.5 | 19.5 | 23.1 |
| Taxa | ALO | GIOU | OXO | PAL | PANT | TROU | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CE | SD | CE | SD | CE | SD | CE | SD | CE | SD | CE | SD | |
| Macroalgae | 6 | - | 5 | 3 | 4 | 5 | 5 | 1 | 6 | 2 | 7 | 1 |
| Foraminifera | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | - | 1 |
| Porifera | 10 | 6 | 15 | 16 | 14 | 22 | 15 | 22 | 14 | 9 | 17 | 23 |
| Cnidaria | 2 | 1 | 1 | 4 | 1 | 4 | - | 1 | 3 | 2 | 2 | 4 |
| Polychaeta | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Mollusca | 2 | 1 | 1 | 1 | - | - | - | - | 1 | - | - | - |
| Crustacea | - | - | - | 1 | - | - | - | - | - | 1 | - | - |
| Bryozoa | 7 | 3 | 4 | 5 | 4 | 6 | 2 | 4 | 9 | 5 | 5 | 7 |
| Brachiopoda | - | 3 | - | 2 | - | - | 2 | 2 | 1 | - | - | 2 |
| Ascidiacea | 3 | 1 | 1 | 3 | 2 | 2 | 2 | 2 | 4 | - | 1 | 5 |
| SUM | 32 | 17 | 29 | 37 | 27 | 41 | 28 | 34 | 40 | 21 | 33 | 44 |
| Cave | CavEBQI | Ecological Quality | CIEBQI | Confidence Index |
|---|---|---|---|---|
| Alona | 5.2 | Moderate | 10 | High |
| Giourious | 3.7 | Poor | 10 | High |
| Oxonisos | 3.8 | Poor | 10 | High |
| Palatia | 2.7 | Poor | 10 | High |
| Panteleimonas | 3.5 | Poor | 10 | High |
| Troulakas | 4.5 | Moderate | 10 | High |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Digenis, M.; Arvanitidis, C.; Dailianis, T.; Gerovasileiou, V. Comparative Study of Marine Cave Communities in a Protected Area of the South-Eastern Aegean Sea, Greece. J. Mar. Sci. Eng. 2022, 10, 660. https://doi.org/10.3390/jmse10050660
Digenis M, Arvanitidis C, Dailianis T, Gerovasileiou V. Comparative Study of Marine Cave Communities in a Protected Area of the South-Eastern Aegean Sea, Greece. Journal of Marine Science and Engineering. 2022; 10(5):660. https://doi.org/10.3390/jmse10050660
Chicago/Turabian StyleDigenis, Markos, Christos Arvanitidis, Thanos Dailianis, and Vasilis Gerovasileiou. 2022. "Comparative Study of Marine Cave Communities in a Protected Area of the South-Eastern Aegean Sea, Greece" Journal of Marine Science and Engineering 10, no. 5: 660. https://doi.org/10.3390/jmse10050660
APA StyleDigenis, M., Arvanitidis, C., Dailianis, T., & Gerovasileiou, V. (2022). Comparative Study of Marine Cave Communities in a Protected Area of the South-Eastern Aegean Sea, Greece. Journal of Marine Science and Engineering, 10(5), 660. https://doi.org/10.3390/jmse10050660