Effects of Light and Autotoxicity on the Reproduction of Bidens pilosa L.: From Laboratory to the Field



Abstract
1. Introduction
2. Materials and Methods
2.1. Seeds and Plant Materials
2.2. Laboratory Experiment: The Effects of Light and Residue Extract on Cobbler Pegs in the Controlled Conditions
2.2.1. Light Treatments
2.2.2. Residue Extract Preparation
2.2.3. Germination Condition
2.2.4. Radicle Length Measurement
2.3. Pot Experiment: The Effects of Light and Residue on Cobbler Pegs in the Seminatural Conditions
2.4. Field Experiment: The Effects of FR “Pretreatment” and Residues on Cobbler Pegs in the Natural Conditions
2.5. Statistical Analysis
3. Results
3.1. The Effects of Light and Residue Extract on Bidens pilosa L. (Cobbers Pegs) in the Controlled Conditions
3.1.1. Light Requirement of Germination
3.1.2. Responses to FR Pretreatment
3.1.3. Responses to FR Irradiation
3.1.4. Response of Radicle Growth to Light and Residue Extract
3.2. The Effects of Light and Residue on Cobbler’s Pegs in the Seminatural Conditions
3.2.1. Light Requirement of Germination
3.2.2. Germination Responses to FR Pretreatments
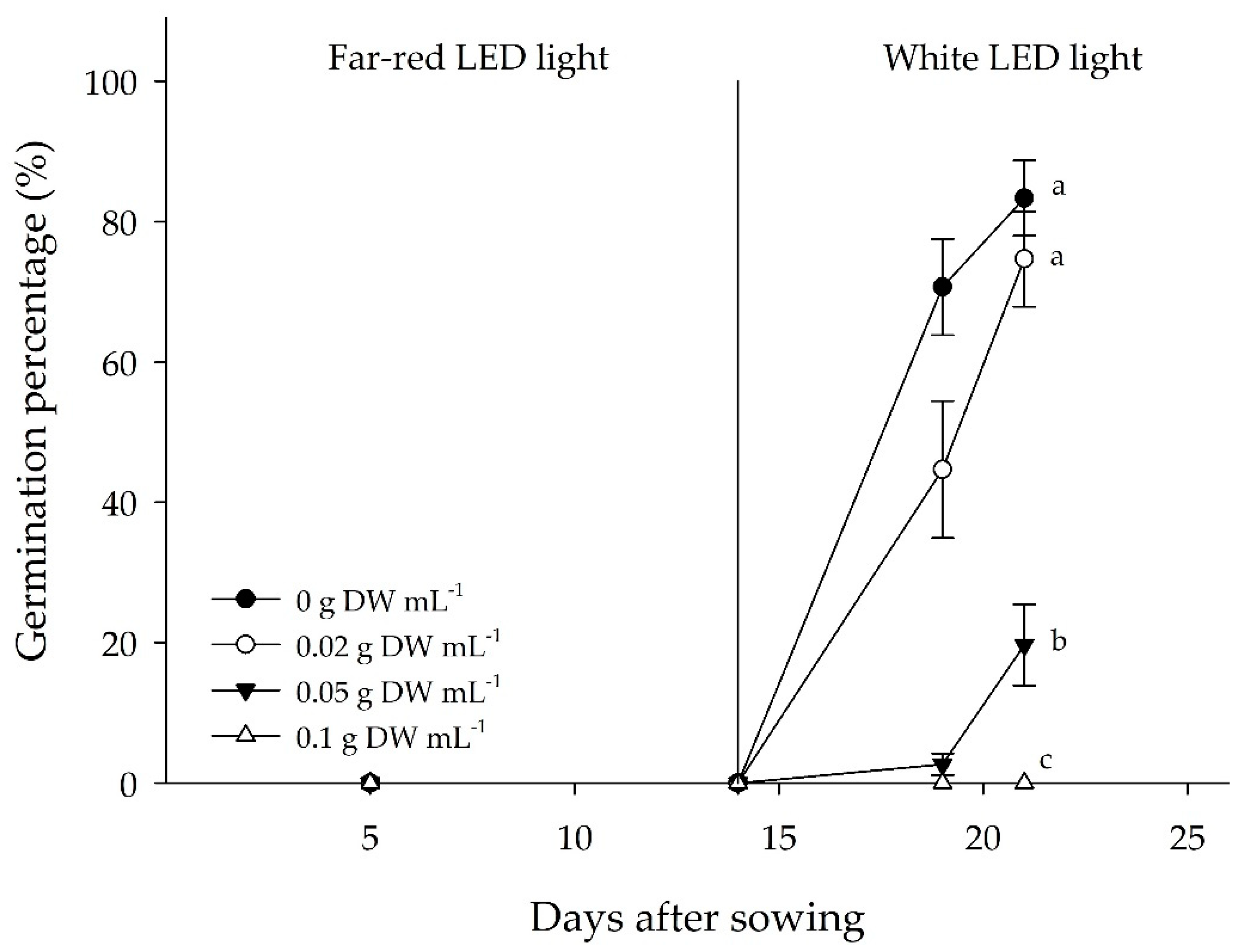
3.2.3. Germination Responses to FR Irradiation
3.3. The Effects of Light and Residue on Cobbler’s Pegs in the Natural Conditions
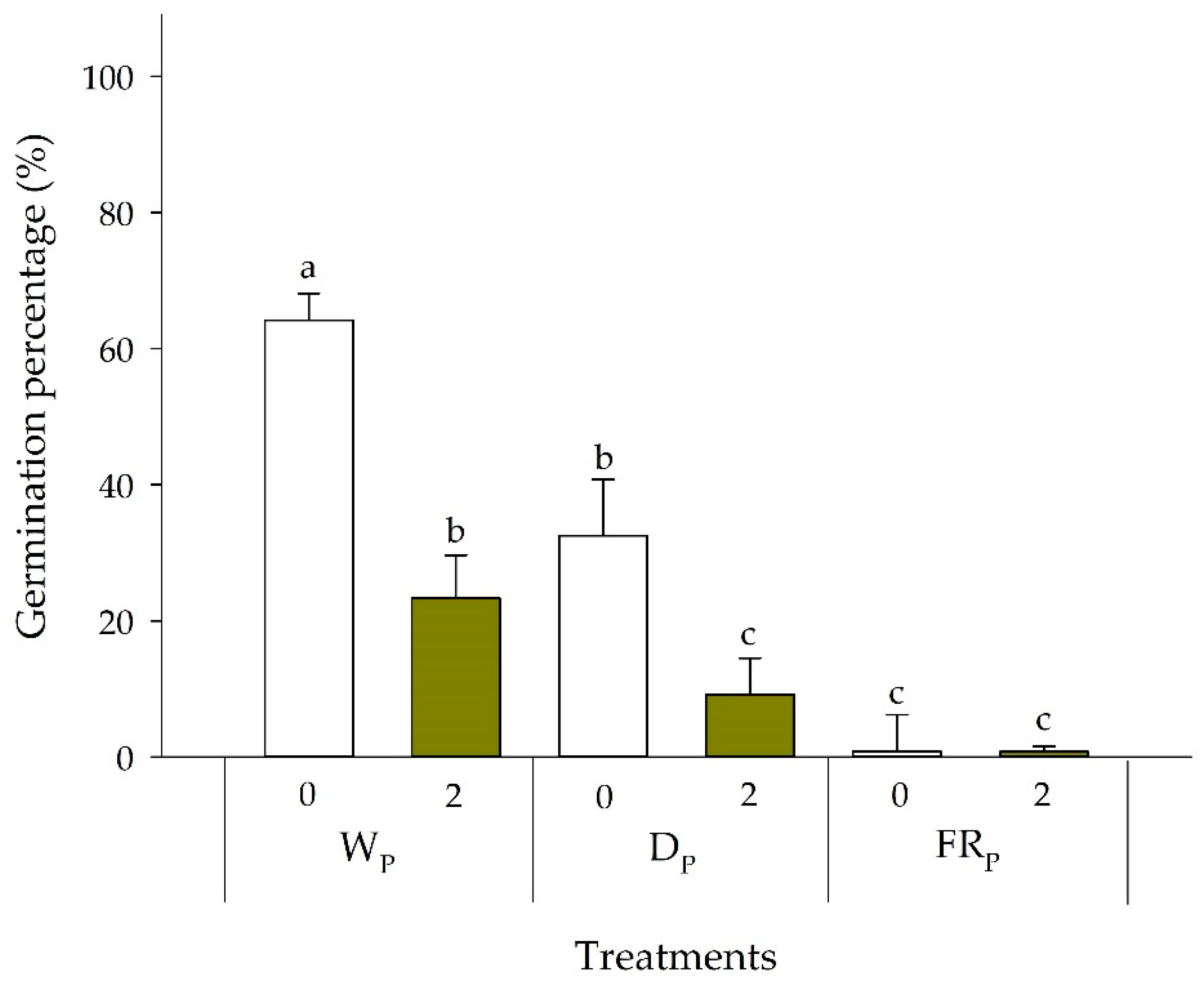
3.3.1. Germination Responses to FR Pretreatments
3.3.2. Germination Responses to Residue Application
4. Discussion
4.1. Effects of Light on Seed Germination
4.1.1. Light Requirements of Cobbler’s Pegs Germination
4.1.2. The Responses of Germination to Far-Red Light (FR) Pretreatments
4.1.3. The Responses of Germination to FR Irradiation
4.2. Other Factors That May Affect the Seed Germination
4.3. Autotoxic/Phytotoxic Effects of Residues on the Seed Germination and Radicle Growth
4.3.1. Responses of Germination
4.3.2. Responses of Radicle Growth
4.4. Implications for Agricultural Practices
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Silva, F.L.; Fischer, D.C.H.; Tavares, J.F.; Silva, M.S.; de Athayde-Filho, P.F.; Barbosa-Filho, J.M. Compilation of secondary metabolites from Bidens pilosa L. Molecules 2011, 16, 1070–1102. [Google Scholar] [CrossRef] [PubMed]
- Bartolome, A.P.; Villaseñor, I.M.; Yang, W.C. Bidens pilosa L. (Asteraceae): Botanical properties, traditional uses, phytochemistry, and pharmacology. Evid. Based Compl. Alt. 2013, 2013, 51. [Google Scholar] [CrossRef] [PubMed]
- Khanh, T.D.; Cong, L.C.; Xuan, T.D.; Uezato, Y.; Deba, F.; Toyama, T.; Tawata, S. Allelopathic plants: 20. hairy beggarticks (Bidens pilosa L.). Allelopathy J. 2009, 24, 243–254. [Google Scholar]
- Lok, A.; Tan, K.-x.; Tan, H. The ecology and distribution in Singapore of Bidens pilosa L.(Asteraceae). Cosmos 2010, 6, 39–44. [Google Scholar] [CrossRef]
- Luo, Y.T.; Wang, Z.M.; Cui, X.L.; Zhao, L.K.; Wang, J.H.; Luo, Y.L. The reproductive traits and invasiveness of Bidens pilosa var. radiata. Chin. J. Ecol. 2019, 38, 655–662. [Google Scholar]
- Shen, S.; Xu, G.; Li, D.; Clements, D.R.; Jin, G.; Yin, X.; Gao, R.; Zhang, F. Occurrence and damage of invasive alien plants in Dehong Prefecture, western of Yunnan Province. Acta Ecol. Sin. 2017, 37, 195–200. [Google Scholar] [CrossRef]
- Deba, F.; Xuan, T.D.; Yasuda, M.; Tawata, S. Herbicidal and fungicidal activities and identification of potential phytotoxins from Bidens pilosa L. var. radiata Scherff. Weed Biol. Manag. 2007, 7, 77–83. [Google Scholar] [CrossRef]
- Shimamoto, Y.; Nomura, N.; Takaso, T.; Setoguchi, H. Overcompensation of seed production caused by clipping of Bidens pilosa var. radiata (Compositae): Implications for weed control on Iriomote-Jima Island, Japan. Weed Biol. Manag. 2011, 11, 118–126. [Google Scholar]
- Chiang, M.Y.; Hsu, L.M.; Yuan, C.Y.; Chen, F.Y.; Chiang, Y.C. Harmful and ecology of invasive plants of Taiwan. In Proceedings of the Harmful Effect and Field Management of Mikania micrantha, Hualien, Taiwan, 1 October 2003; HDARES: Hualien, Taiwan, 2003; pp. 97–109. [Google Scholar]
- Hsu, H.M.; Kao, W.Y. Contrasting effects of aqueous tissue extracts from an invasive plant, Bidens pilosa L. var. radiata, on the performance of its sympatric plant species. Taiwania 2009, 54, 255–260. [Google Scholar]
- Huang, Y.L.; Kao, W.Y. Comparisons of growth, biomass allocation, and morphology of an invasive and two non-invasive varieties of Bidens pilosa in Taiwan. Taiwania 2016, 61, 288–294. [Google Scholar]
- Poonpaiboonpipat, T.; Poolkum, S. Utilization of Bidens pilosa var. radiata (Sch. Bip.) Sherff integrated with water irrigation for paddy weed control and rice yield production. Weed Biol. Manag. 2019, 19, 31–38. [Google Scholar] [CrossRef]
- Zungsontiporn, S. Some characteristics of Bidens pilosa L. var. radiata Scheff., a new invasive species in Thailand. In Proceedings of the 21st Asian Pacific Weed Science Society (APWSS) Conference, Colombo, Sri Lanka, 2–6 October 2007; Asian Pacific Weed Science Society: Colombo, Sri Lanka, 2007; pp. 558–564. [Google Scholar]
- Arthur, G.D.; Naidoo, K.K.; Coopoosamy, R.M. Bidens pilosa L.: Agricultural and pharmaceutical importance. J. Med. Plants Res. 2012, 6, 3282–3287. [Google Scholar] [CrossRef]
- Wang, Y.C. Vanishing Diet: Searching for the Appearance of Diet of Amis Tribe’s Elders—The Case of Wild Vegetable. Master’s Thesis, Tunghai University, Taitung, Taiwan, December 2017. [Google Scholar]
- Li, F.H. Analysis of Antioxidant Capacity of Eight Common Wild Vegetables in Taiwan Flatland. Master’s Thesis, Chung Yuan Christian University, Taoyuan, Taiwan, January 2019. [Google Scholar]
- Chen, S.L.; Lin, Y.R.; Wang, R.M.; Chou, P.H.; Chen, M.H.; Huang, M.F.; Huang, C.T.; Lin, R.T.; Lin, W.J. A study of the antioxidant and anti-inflammatory potential of supercritical carbon dioxide extract of Bidens pilosa. Radiol. Tech. 2011, 42, 9–17. [Google Scholar]
- Hsueh, M.T.; Fan, C.H.; Chang, W.L. Allelopathic effects of Bidens pilosa L. var. radiata Sch. Bip. on the tuber sprouting and seedling growth of Cyperus rotundus L. Plants 2020, 9, 742. [Google Scholar]
- Hsiao, C.L. Safe production techniques of Bidens pilosa for high content of functional compounds. Stock-Farming Tendays 2017, 1908, 43–44. [Google Scholar]
- Hsu, H.M.; Kao, W.Y. Vegetative and reproductive growth of an invasive weed Bidens pilosa L. var. radiata and its noninvasive congener Bidens bipinnata in Taiwan. Taiwania 2014, 59, 119–126. [Google Scholar]
- Batlla, D.; Benech-Arnold, R.L. Weed seed germination and the light environment: Implications for weed management. Weed Biol. Manag. 2014, 14, 77–87. [Google Scholar] [CrossRef]
- Scopel, A.L.; Ballaré, C.L.; Sánchez, R.A. Induction of extreme light sensitivity in buried weed seeds and its role in the perception of soil cultivations. Plant Cell Environ. 1991, 14, 501–508. [Google Scholar] [CrossRef]
- Botto, J.F.; Scopel, A.L.; Ballaré, C.L.; Sánchez, R.A. The effect of light during and after soil cultivation with different tillage implements on weed seedling emergence. Weed Sci. 1998, 46, 351–357. [Google Scholar] [CrossRef]
- Buhler, D.D. Effects of tillage and light environment on emergence of 13 annual weeds. Weed Technol. 1997, 11, 496–501. [Google Scholar] [CrossRef]
- Jensen, P.K. Effect of light environment during soil disturbance on germination and emergence pattern of weeds. Ann. Appl. Biol. 1995, 127, 561–571. [Google Scholar] [CrossRef]
- Hsu, L.M.; Lin, H.S. Comparison of morphology and seed germination of three Bidens Species. Weed Sci. Bull. 2005, 26, 33–42. [Google Scholar]
- Hsueh, M.T.; Chang, W.L. The effect of light and pH on the seed germination of Bidens pilosa var. radiata. Weed Sci. Bull. 2017, 38, 89–98. [Google Scholar]
- Gallagher, R.S.; Cardina, J. Phytochrome-mediated Amaranthus germination II: Development of very low fluence sensitivity. Weed Sci. 1998, 46, 53–58. [Google Scholar] [CrossRef]
- Rashid, M.H.; Asaeda, T.; Uddin, M.N. Litter-mediated allelopathic effects of kudzu (Pueraria montana) on Bidens pilosa and Lolium perenne and its persistence in soil. Weed Biol. Manag. 2010, 10, 48–56. [Google Scholar] [CrossRef]
- El-Gawad, A.M.A.; Mashaly, I.A.; Ziada, M.E.A.; Deweeb, M.R. Phytotoxicity of three Plantago species on germination and seedling growth of hairy beggarticks (Bidens pilosa L.). Egypt J. Basic Appl. Sci. 2015, 2, 303–309. [Google Scholar] [CrossRef]
- Wat, C.; Biswas, R.K.; Graham, E.A.; Bohm, L.; Towers, G.H.N.; Waygood, E.R. Ultraviolet-mediated cytotoxic activity of phenylheptatriyne from Bidens pilosa L. J. Nat. Prod. 1979, 42, 103–111. [Google Scholar] [CrossRef]
- Campbell, G.; Lambert, J.D.H.; Arnason, T.; Towers, G.H.N. Allelopathic properties of α-terthienyl and phenylheptatriyne, naturally occurring compounds from species of Asteraceae. J. Chem. Ecol. 1982, 8, 961–972. [Google Scholar] [CrossRef]
- Shen, M.L. Experimental Designs; Jeou Chou Book Co, Ltd.: Taipei, Taiwan, 2010. [Google Scholar]
- Fenner, M. The inhibition of germination of Bidens pilosa seeds by leaf canopy shade in some natural vegetation types. New Phytol. 1980, 84, 95–101. [Google Scholar] [CrossRef]
- Fenner, M. The induction of a light requirement in Bidens pilosa seeds by leaf canopy shade. New Phytol. 1980, 84, 103–106. [Google Scholar] [CrossRef]
- Reddy, K.N.; Singh, M. Germination and emergence of hairy beggarticks (Bidens pilosa). Weed Sci. 1992, 40, 195–199. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Ali, H.H.; Florentine, S. Seed germination ecology of Bidens pilosa and its implications for weed management. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, C.; Brown, N.A.C. Germination of the dimorphic fruits of Bidens pilosa L. New Phytol. 1982, 90, 151–164. [Google Scholar] [CrossRef]
- Van Rooden, J.; Akkermans, L.M.A.; Van Der Veen, R. A study on photoblastism in seeds of some tropical weeds. Acta Bot. Neerl. 1970, 19, 257–264. [Google Scholar] [CrossRef]
- Holmes, M.; Smith, H. The function of phytochrome in the natural environment—II. The influence of vegetation canopies on the spectral energy distribution of natural daylight. Photochem. Photobiol. 1977, 25, 539–545. [Google Scholar] [CrossRef]
- Fenner, M. Germination tests on thirty-two East African weed species. Weed Res. 1980, 20, 135–138. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and, Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014; pp. 5–35. [Google Scholar]
- Baskin, C.C.; Baskin, J.M.; Chester, E.W. Seed dormancy and germination in Rhexia mariana var. interior (Melastomataceae) and eco-evolutionary implications. Can. J. Botany. 1999, 77, 488–493. [Google Scholar]
- Shamsi, S.R.A.; Whitehead, F.H. Comparative eco-physiology of Epilobium hirsutum L. and Lythrum salicaria L.: I. General biology, distribution and germination. J. Ecol. 1974, 62, 279–290. [Google Scholar] [CrossRef]
- Hsiao, A.I. The effect of sodium hypochlorite, gibberellic acid and light on seed dormancy and germination of stinkweed and wild mustard. Can. J. Plant Sci. 1980, 60, 643–649. [Google Scholar] [CrossRef]
- Jiang, G.B.; Huang, D.Y.; Huang, L.M.; Lin, G.W. Self-allelopathy of aquatic extracts from different parts of Ageratum conyzoides and Bidens pilosa. J. Hunan Agric. Univ. 2011, 37, 624–626. [Google Scholar] [CrossRef]
- Wilson, R.E.; Rice, E.L. Allelopathy as Expressed by Helianthus annuus and Its Role in Old-Field Succession. Bull. Torrey Bot. Club 1968, 95, 432–448. [Google Scholar] [CrossRef]
- Picman, J.; Picman, A.K. Autotoxicity in Parthenium hysterophorus and its possible role in control of germination. Biochem. Syst. Ecol. 1984, 12, 287–292. [Google Scholar] [CrossRef]
- Qi, S.S.; Dai, Z.C.; Miao, S.L.; Zhai, D.L.; Si, C.C.; Huang, P.; Wang, R.P.; Du, D.L. Light limitation and litter of an invasive clonal plant, Wedelia trilobata, inhibit its seedling recruitment. Ann. Bot. 2014, 114, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Hazarika, U.K. Allelopathic effects of Galinsoga parviflora Car. and Bidens pilosa L. on germination and seedling growth of soybean and groundnut. Allelopathy J. 1996, 3, 89–92. [Google Scholar]
- Zeng, R.; Luo, S. Allelopathic effects of root exudates of Cymbopogon citratus, Ageratum conyzoides and Bidens pilosa. J. South China Agric. Univ. 1996, 17, 119–120. [Google Scholar]
- Zeng, R.; Luo, S. Study on allelopathic potentials of Cymbopogon citratus, Ageratum conyzoides and Bidens pilosa. J. South China Agric. Univ. 1993, 14, 8–14. [Google Scholar]
- Hong, N.H.; Xuan, T.D.; Eiji, T.; Khanh, T.D. Paddy weed control by higher plants from Southeast Asia. Crop Prot. 2004, 23, 255–261. [Google Scholar] [CrossRef]
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant Soil 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Blum, U. Effects of microbial utilization of phenolic acids and their phenolic acid breakdown products on allelopathic interactions. J. Chem. Ecol. 1998, 24, 685–708. [Google Scholar] [CrossRef]
- Blum, U. Allelopathy: A soil system perspective. In Allelopathy: A Physiological Process with Ecological Implications; Reigosa, M.J., Pedrol, N., Gonzalez, L., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 299–340. [Google Scholar]
- Central Weather Bureau Observation Data Inquire System. Available online: https://e-service.cwb.gov.tw/HistoryDataQuery/index.jsp (accessed on 24 August 2020).
- Wang, J.; Yang, H.; Lin, Z.W.; Sun, H.D. Flavonoids from Bidens pilosa var. radiata. Phytochemistry 1997, 46, 1275–1278. [Google Scholar] [CrossRef]
- Litts, J.C.; Kelly, J.M.; Lagarias, J.C. Structure-function studies on phytochrome. Preliminary characterization of highly purified phytochrome from Avena sativa enriched in the 124-kilodalton species. J. Biol. Chem. 1983, 258, 11025–11031. [Google Scholar] [PubMed]
- Clough, R.C.; Vierstra, R.D. Phytochrome degradation. Plant Cell Environ. 1997, 20, 713–721. [Google Scholar] [CrossRef]
- Haugland, E.; Brandsaeter, L.O. Experiments on bioassay sensitivity in the study of allelopathy. J. Chem. Ecol. 1996, 22, 1845–1859. [Google Scholar] [CrossRef] [PubMed]
- Krumsri, R.; Suwunnamek, U.; Homhaul, W.; Laosinwattana, C.; Poonpaiboonpipattana, T. Allelopathic effects of Bidens pilosa var. radiata and its preliminary utilization to control weeds in rice. J. Agric. Technol. 2015, 11, 1875–1886. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Factor | Phytotoxic Treatment 1 | Light Treatment 2 | Type of Light 3 | Hour(s) of Far-Red Light (FR) Pretreatment | Photoperiod | Incubation Days |
|---|---|---|---|---|---|---|
| Incubation temperature 25 °C | ||||||
| Light requirement | Residue extract | WL | White LED | 0 | 12/12 | 5 |
| DL | Darkness | 0 | Continuous | 5 | ||
| FR pretreatment | Residue extract | FR-WL | White LED | 3 | 12/12 | 5 |
| FR-DL | Darkness | 3 | Continuous | 5 | ||
| FR irradiation | Residue extract | FRL | Far-red LED White LED | 0 | 12/12 12/12 | 14 4 7 |
| Conditions of The Growth Chamber | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Light Factor | Phytotoxic Treatment 1 | Light Treatment 2 | Type of Light 3 | Hour(s) of FR Pretreatment | Photoperiod | Incubation Days | ||||
| Incubation temperature 25 °C | ||||||||||
| Light requirement | Residue | WP | White LED | 0 | 12/12 | 7 | ||||
| DP | Darkness | 0 | Continuous | 7 | ||||||
| Different FR pretreatment time | N/A | FR0-WP | White LED | 0 | 12/12 | 7 | ||||
| FR0-DP | Darkness | 0 | Continuous | 7 | ||||||
| FR3-WP | White LED | 3 | 12/12 | 7 | ||||||
| FR3-Dp | Darkness | 3 | Continuous | 7 | ||||||
| FR24-WP | White LED | 24 | 12/12 | 7 | ||||||
| FR24-DP | Darkness | 24 | Continuous | 7 | ||||||
| FR irradiation | Residue | FRP | Far-red LED | 0 | 12/12 | 7 | ||||
| Conditions of The Soil | ||||||||||
| pH | EC (dS cm−1) | P2O5 (mg kg−1) | K2O (mg kg−1) | Organic Matter (%) | ||||||
| 6.35 | 0.07 | 116.01 | 95.49 | 3.57 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsueh, M.-T.; Fan, C.; Lo, H.-F.; Chang, W.-L. Effects of Light and Autotoxicity on the Reproduction of Bidens pilosa L.: From Laboratory to the Field. Agriculture 2020, 10, 555. https://doi.org/10.3390/agriculture10110555
Hsueh M-T, Fan C, Lo H-F, Chang W-L. Effects of Light and Autotoxicity on the Reproduction of Bidens pilosa L.: From Laboratory to the Field. Agriculture. 2020; 10(11):555. https://doi.org/10.3390/agriculture10110555
Chicago/Turabian StyleHsueh, Ming-Tung, Chihhao Fan, Hsiao-Feng Lo, and Wen-Lian Chang. 2020. "Effects of Light and Autotoxicity on the Reproduction of Bidens pilosa L.: From Laboratory to the Field" Agriculture 10, no. 11: 555. https://doi.org/10.3390/agriculture10110555
APA StyleHsueh, M.-T., Fan, C., Lo, H.-F., & Chang, W.-L. (2020). Effects of Light and Autotoxicity on the Reproduction of Bidens pilosa L.: From Laboratory to the Field. Agriculture, 10(11), 555. https://doi.org/10.3390/agriculture10110555