Cardiac and Vascular Sympathetic Baroreflex Control during Orthostatic Pre-Syncope

 ,
,  ,
,  ,
, 
 and
and 
Abstract
1. Introduction
2. Material and Methods
2.1. Experimental Protocol
2.2. Data Analysis
2.3. Power Spectral Analysis of RR Interval and SAP Variability
2.4. MSNA Assessment and Calibrated MSNA (cMSNA) Series
2.5. Relationship between MSNA and DAP Oscillations in the Frequency Domain
2.6. Cardiac Baroreflex Modulation (cBRS)
2.7. Sympathetic Neural Discharge Baroreflex Modulation (sBRS)
2.8. Statistics
3. Results
4. Discussion
Limitations of the Study and Future Developments
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brignole, M.; Moya, A.; De Lange, F.J.; Deharo, J.-C.; Elliott, P.M.; Fanciulli, A.; Fedorowski, A.; Furlan, R.; Kenny, R.A.; Martín, A.; et al. 2018 ESC Guidelines for the diagnosis and management of syncope. Eur. Heart J. 2018, 39, 1883–1948. [Google Scholar] [CrossRef] [PubMed]
- Costantino, G.; Perego, F.; Dipaola, F.; Borella, M.; Galli, A.; Cantoni, G.; Dell’Orto, S.; Dassi, S.; Filardo, N.; Duca, P.G.; et al. Short- and long-term prognosis of syncope, risk factors, and role of hospital admission: results from the STePS (Short-Term Prognosis of Syncope) study. J. Am. Coll. Cardiol. 2008, 51, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Mosqueda-Garcia, R.; Furlan, R.; Md, J.T.; Fernandez-Violante, R. The Elusive Pathophysiology of Neurally Mediated Syncope. Circulation 2000, 102, 2898–2906. [Google Scholar] [CrossRef] [PubMed]
- Furlan, R.; Alboni, P.; Mosqueda-Garcia, R. Pathophysiology of Vasovagal Syncope: Conclusive Remarks. In Vasovagal Syncope; Alboni, P., Furlan, R., Eds.; Springer: Cham, Switzerland, 2015; Volume 11, pp. 95–102. [Google Scholar]
- Mosqueda-Garcia, R.; Furlan, R.; Fernandez-Violante, R.; Desai, T.; Snell, M.; Járai, Z.; Ananthram, V.; Robertson, R.M.; Robertson, D. Sympathetic and baroreceptor reflex function in neurally mediated syncope evoked by tilt. J. Clin. Investig. 1997, 99, 2736–2744. [Google Scholar] [CrossRef] [PubMed]
- Diedrich, A.; Paranjape, S.Y.; Robertson, D. Plasma and Blood Volume in Space. Am. J. Med. Sci. 2007, 334, 80–86. [Google Scholar] [CrossRef]
- Robertson, D.; Diedrich, A.; Chapleau, M.W. Editorial on arterial baroreflex issue. Auton. Neurosci. 2012, 172, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Furlan, R.; Porta, A.; Costa, F.; Tank, J.; Baker, L.; Schiavi, R.; Robertson, D.; Malliani, A.; Mosqueda-Garcia, R. Oscillatory Patterns in Sympathetic Neural Discharge and Cardiovascular Variables During Orthostatic Stimulus. Circulation 2000, 101, 886–892. [Google Scholar] [CrossRef]
- Diedrich, A.; Porta, A.; Barbic, F.; Brychta, R.J.; Bonizzi, P.; Diedrich, L.; Cerutti, S.; Robertson, D.; Furlan, R. Lateralization of expression of neural sympathetic activity to the vessels and effects of carotid baroreceptor stimulation. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1758–H1765. [Google Scholar] [CrossRef][Green Version]
- Kamiya, A.; Hayano, J.; Kawada, T.; Michikami, D.; Yamamoto, K.; Ariumi, H.; Shimizu, S.; Uemura, K.; Miyamoto, T.; Aiba, T.; et al. Low-frequency oscillation of sympathetic nerve activity decreases during development of tilt-induced syncope preceding sympathetic withdrawal and bradycardia. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H1758–H1769. [Google Scholar] [CrossRef]
- Schwartz, C.E.; Lambert, E.; Medow, M.S.; Stewart, J.M. Disruption of phase synchronization between blood pressure and muscle sympathetic nerve activity in postural vasovagal syncope. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H1238–H1245. [Google Scholar] [CrossRef]
- Mosqueda-Garcia, R.; Robertson, R.M.; Hollister, A.S.; Biaggioni, I.; Netterville, J.L.; Robertson, D. The Diagnosis and Treatment of Baroreflex Failure. N. Engl. J. Med. 1993, 329, 1449–1455. [Google Scholar]
- Furlan, R.; Magatelli, R.; Palazzolo, L.; Rimoldi, A.; Colombo, S.; Porta, A. Orthostatic intolerance: different abnormalities in the neural sympathetic response to a gravitational stimulus. Auton. Neurosci. 2001, 90, 83–88. [Google Scholar] [CrossRef]
- Jordan, J.; Shannon, J.R.; Black, B.K.; Costa, F.; Ertl, A.C.; Furlan, R.; Biaggioni, I.; Robertson, D. Malignant Vagotonia Due to Selective Baroreflex Failure. Hypertension 1997, 30, 1072–1077. [Google Scholar] [CrossRef] [PubMed]
- Heusser, K.; Tank, J.; Luft, F.C.; Jordan, J. Baroreflex Failure. Hypertension 2005, 45, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Morillo, C.A.; Eckberg, D.L.; Ellenbogen, K.A.; Beightol, L.A.; Hoag, J.B.; Tahvanainen, K.U.; Kuusela, T.A.; Diedrich, A.M. Vagal and Sympathetic Mechanisms in Patients With Orthostatic Vasovagal Syncope. Circulation 1997, 96, 2509–2513. [Google Scholar] [CrossRef] [PubMed]
- Ichinose, M.; Saito, M.; Fujii, N.; Kondo, N.; Nishiyasu, T. Modulation of the control of muscle sympathetic nerve activity during severe orthostatic stress. J. Physiol. 2006, 576, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Barbic, F.; Heusser, K.; Marchi, A.; Zamunér, A.R.; Gauger, P.; Tank, J.; Jordan, J.; Diedrich, A.; Robertson, D.; DiPaola, F.; et al. Cardiovascular parameters and neural sympathetic discharge variability before orthostatic syncope: Role of sympathetic baroreflex control to the vessels. Physiol. Meas. 2015, 36, 633–641. [Google Scholar] [CrossRef]
- Mosqueda-Garcia, R. Pathophysiology of Vasovagal Syncope: Role of Baroreceptor Mechanisms. In Vasovagal Syncope; Alboni, P., Furlan, R., Eds.; Springer: Cham, Switzerland, 2015; Volume 11, pp. 67–74. [Google Scholar]
- Cooke, W.H.; Rickards, C.A.; Ryan, K.L.; Kuusela, T.A.; Convertino, V.A. Muscle sympathetic nerve activity during intense lower body negative pressure to presyncope in humans. J. Physiol. 2009, 587, 4987–4999. [Google Scholar] [CrossRef]
- Furlan, R.; Montano, N.; Porta, A. Cardiovascular Rhythms in Vasovagal Syncope. In Vasovagal Syncope; Alboni, P., Furlan, R., Eds.; Springer: Cham, Switzerland, 2015; Volume 11, pp. 83–93. [Google Scholar]
- Stauss, H.M.; Anderson, E.A.; Haynes, W.G.; Kregel, K.C. Frequency response characteristics of sympathetically mediated vasomotor waves in humans. Am. J. Physiol. 1998, 274, H1277–H1283. [Google Scholar] [CrossRef]
- Diedrich, A.; Crossman, A.A.; Beightol, L.A.; Tahvanainen, K.U.; Kuusela, T.A.; Ertl, A.C.; Eckberg, D.L. Baroreflex physiology studied in healthy subjects with very infrequent muscle sympathetic bursts. J. Appl. Physiol. (1985) 2013, 114, 203–210. [Google Scholar] [CrossRef][Green Version]
- Buehlmeier, J.; Mulder, E.; Noppe, A.; Frings-Meuthen, P.; Angerer, O.; Rudwill, F.; Biolo, G.; Smith, S.M.; Blanc, S.; Heer, M. A combination of whey protein and potassium bicarbonate supplements during head-down-tilt bed rest: Presentation of a multidisciplinary randomized controlled trial (MEP study). Acta Astronaut. 2014, 95, 82–91. [Google Scholar] [CrossRef]
- Mosqueda-Garcia, R. Microneurography in neurological research. Am. Acad. Neurol. 1996, 2, 4–5. [Google Scholar]
- Pagani, M.; Lombardi, F.; Guzzetti, S.; Rimoldi, O.; Furlan, R.; Pizzinelli, P.; Sandrone, G.; Malfatto, G.; Dell’Orto, S.; Piccaluga, E. Power spectral analysis of heart rate and arterial pressure variabilities as a marker of sympatho-vagal interaction in man and conscious dog. Circ. Res. 1986, 59, 178–193. [Google Scholar] [CrossRef] [PubMed]
- Malik, M. Heart rate variability: standards of measurement, physiological interpretation and clinical use. Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology. Circulation 1996, 93, 1043–1065. [Google Scholar] [CrossRef]
- Parati, G.; Saul, J.P.; Di Rienzo, M.; Mancia, G. Spectral Analysis of Blood Pressure and Heart Rate Variability in Evaluating Cardiovascular Regulation. A critical appraisal. Hypertension 1995, 25, 1276–1286. [Google Scholar] [CrossRef]
- Pomeranz, B.; Macaulay, R.J.; Caudill, M.A.; Kutz, I.; Adam, D.; Gordon, D.; Kilborn, K.M.; Barger, A.C.; Shannon, D.C.; Cohen, R.J.; et al. Assessment of autonomic function in humans by heart rate spectral analysis. Am. J. Physiol. 1985, 248, H151–H153. [Google Scholar] [CrossRef]
- Barbic, F.; Perego, F.; Canesi, M.; Gianni, M.; Biagiotti, S.; Costantino, G.; Pezzoli, G.; Porta, A.; Malliani, A.; Furlan, R. Early Abnormalities of Vascular and Cardiac Autonomic Control in Parkinson’s Disease Without Orthostatic Hypotension. Hypertension 2007, 49, 120–126. [Google Scholar] [CrossRef]
- Hamner, J.W.; Taylor, J.A. Automated quantification of sympathetic beat-by-beat activity, independent of signal quality. J. Appl. Physiol. (1985) 2001, 91, 1199–1206. [Google Scholar] [CrossRef]
- Wallin, B.G.; Burke, D.; Gandevia, S. Coupling between variations in strength and baroreflex latency of sympathetic discharges in human muscle nerves. J. Physiol. 1994, 474, 331–338. [Google Scholar] [CrossRef]
- Marchi, A.; Bari, V.; De Maria, B.; Esler, M.D.; Lambert, E.A.; Baumert, M.; Porta, A. Calibrated variability of muscle sympathetic nerve activity during graded head-up tilt in humans and its link with noradrenaline data and cardiovascular rhythms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R1134–R1143. [Google Scholar] [CrossRef]
- Pagani, M.; Montano, N.; Porta, A.; Malliani, A.; Abboud, F.M.; Birkett, C.; Somers, V.K. Relationship Between Spectral Components of Cardiovascular Variabilities and Direct Measures of Muscle Sympathetic Nerve Activity in Humans. Circulation 1997, 95, 1441–1448. [Google Scholar] [CrossRef] [PubMed]
- Porta, A.; Bari, V.; Bassani, T.; Marchi, A.; Pistuddi, V.; Ranucci, M. Model-based causal closed-loop approach to the estimate of baroreflex sensitivity during propofol anesthesia in patients undergoing coronary artery bypass graft. J. Appl. Physiol. (1985) 2013, 115, 1032–1042. [Google Scholar] [CrossRef] [PubMed]
- Magagnin, V.; Bassani, T.; Bari, V.; Turiel, M.; Maestri, R.; Pinna, G.D.; Porta, A. Non-stationarities significantly distort short-term spectral, symbolic and entropy heart rate variability indices. Physiol. Meas. 2011, 32, 1775–1786. [Google Scholar] [CrossRef] [PubMed]
- Bertinieri, G.; Di Rienzo, M.; Cavallazzi, A.; Ferrari, A.U.; Pedotti, A.; Mancia, G. Evaluation of baroreceptor reflex by blood pressure monitoring in unanesthetized cats. Am. J. Physiol. 1988, 254, H377–H383. [Google Scholar] [PubMed]
- Parati, G.; Di Rienzo, M.; Bertinieri, G.; Pomidossi, G.; Casadei, R.; Groppelli, A.; Pedotti, A.; Zanchetti, A.; Mancia, G. Evaluation of the baroreceptor-heart rate reflex by 24-hour intra-arterial blood pressure monitoring in humans. Hypertension 1988, 12, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Hart, E.C.; Joyner, M.J.; Wallin, B.G.; Karlsson, T.; Curry, T.B.; Charkoudian, N. Baroreflex control of muscle sympathetic nerve activity: A nonpharmacological measure of baroreflex sensitivity. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H816–H822. [Google Scholar] [CrossRef] [PubMed]
- Marchi, A.; Bari, V.; De Maria, B.; Esler, M.; Lambert, E.; Baumert, M.; Porta, A. Simultaneous Characterization of Sympathetic and Cardiac Arms of the Baroreflex through Sequence Techniques during Incremental Head-Up Tilt. Front. Physiol. 2016, 7, 438. [Google Scholar] [CrossRef] [PubMed]
- Ei-Bedawi, K.M.; Hainsworth, R. Combined head-up tilt and lower body suction: A test of orthostatic tolerance. Clin. Auton. Res. 1994, 4, 41–47. [Google Scholar] [CrossRef]
- Sutton, R.; Brignole, M. Twenty-eight years of research permit reinterpretation of tilt-testing: hypotensive susceptibility rather than diagnosis. Eur. Hear. J. 2014, 35, 2211–2212. [Google Scholar] [CrossRef] [PubMed]
- Piccirillo, G.; Naso, C.; Moisè, A.; Lionetti, M.; Nocco, M.; Di Carlo, S.; De Laurentis, T.; Magrí, D.; Cacciafesta, M.; Marigliano, V. Heart rate and blood pressure variability in subjects with vasovagal syncope. Clin. Sci. 2004, 107, 55–61. [Google Scholar] [CrossRef]
- Ogoh, S.; Volianitis, S.; Raven, P.B.; Secher, N.H. Carotid baroreflex function ceases during vasovagal syncope. Clin. Auton. Res. 2004, 14, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Jardine, D.L.; Melton, I.C.; Crozier, I.G.; English, S.; Bennett, S.I.; Frampton, C.M.; Ikram, H. Decrease in cardiac output and muscle sympathetic activity during vasovagal syncope. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, H1804–H1809. [Google Scholar] [CrossRef] [PubMed]
- Jardine, D.L. Vasovagal syncope: new physiologic insights. Cardiol. Clin. 2013, 31, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Vaddadi, G.; Esler, M.D.; Dawood, T.; Lambert, E. Persistence of muscle sympathetic nerve activity during vasovagal syncope. Eur. Hear. J. 2010, 31, 2027–2033. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Verheyden, B.; Wieling, W.; Levine, B.D. Cardiac output and sympathetic vasoconstrictor responses during upright tilt to presyncope in healthy humans. J. Physiol. 2012, 590, 1839–1848. [Google Scholar] [CrossRef] [PubMed]
- Iwase, S.; Mano, T.; Kamiya, A.; Niimi, Y.; Fu, Q.; Suzumura, A. Syncopal attack alters the burst properties of muscle sympathetic nerve activity in humans. Auton. Neurosci. 2002, 95, 141–145. [Google Scholar] [CrossRef]
- Eckberg, D.L.; Nerhed, C.; Wallin, B.G. Respiratory modulation of muscle sympathetic and vagal cardiac outflow in man. J. Physiol. 1985, 365, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Malliani, A.; Schwartz, P.; Zanchetti, A. A sympathetic reflex elicited by experimental coronary occlusion. Am. J. Physiol. 1969, 217, 703–709. [Google Scholar]



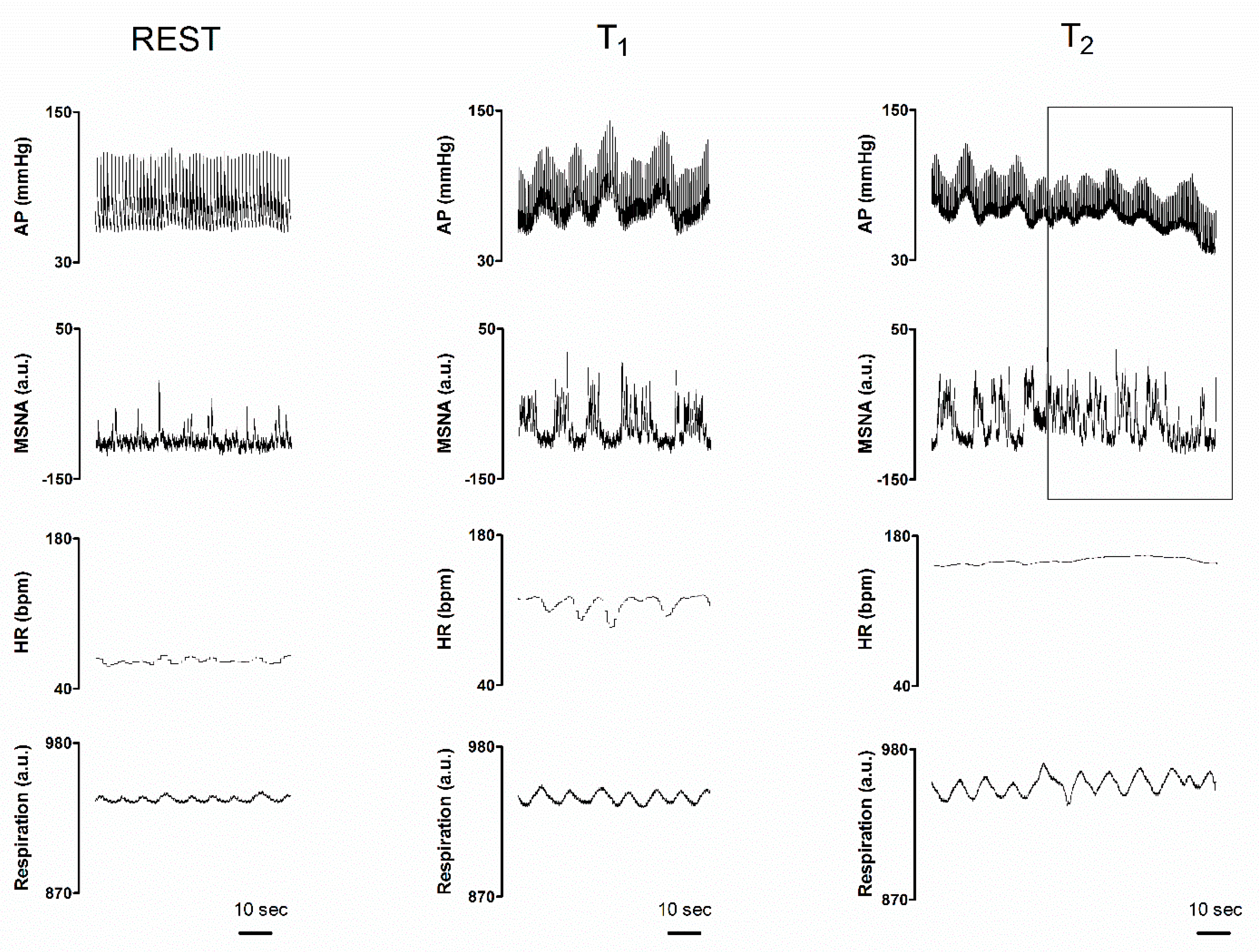{kind=link}
{kind=link}
| Parameters | REST | T1 | T2 |
|---|---|---|---|
| RR (msec) | 847 ± 99 | 609 ± 84 * | 444 ± 87 § |
| SAP (mmHg) | 131 ± 12 | 133 ± 10 | 114 ± 14 #§ |
| DAP (mmHg) | 76 ± 7 | 88 ± 8 * | 71 ± 9 # |
| Resp (breaths/min) | 17 ± 6 | 16 ± 3 | 18 ± 4 |
| MSNA (bursts/min) | 17 ± 6 | 30 ± 5 * | 34 ± 9 § |
| MSNA (bursts/100 beats) | 25 ± 8 | 31 ± 6 | 25 ± 5 |
| Parameters | REST | T1 | T2 |
|---|---|---|---|
| α LF (msec/mmHg) | 14.0 ± 6.5 | 5.7 ± 3.0 * | 1.8 ± 1.9 § |
| cBRSseq | 17.5 ± 5.4 | 6.8 ± 3.7 * | 2.8 ± 1.5 § |
| sBRS (% burst/mmHg) | −6.0 ± 3.3 | −4.4 ± 1.2 | −0.4 ± 0.9 #§ |
| Parameters | REST | T1 | T2 |
|---|---|---|---|
| RRvar (msec2) | 2228 ± 710 | 1711 ± 1388 | 142 ± 223 #§ |
| LFRR (msec2) (n.u.) | 797 ± 665 62.5 ± 25.3 | 1316 ± 1189 88.5 ± 11.3 * | 100 ± 209 #§ 87.7 ± 8.9 § |
| HFRR (msec2) (n.u.) | 328 ± 200 35.9 ± 25.4 | 107 ± 144 9.3 ± 9.7 * | 7 ± 14 #§ 5.8 ± 2.0 § |
| LF/HFRR | 3.7 ± 3.7 | 28.3 ± 35.9 * | 17.7 ± 9.2 § |
| LFSAP (mmHg2) | 3.3 ± 2.4 | 32.6 ± 16.9 * | 10.0 ± 8.5 # |
| LFcMSNA (bursts2·sec−2/Hz) | 0.034 ± 0.014 | 0.058 ± 0.014 | 0.029 ± 0.019 # |
| K2cMSNA-DAP (LF) | 0.75 ± 0.19 | 0.90 ± 0.10 | 0.69 ± 0.15 # |
| HFcMSNA (bursts2·sec−2/Hz) | 0.0025 ± 0.0007 | 0.0028 ± 0.0008 | 0.0044 ± 0.0018 § |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furlan, R.; Heusser, K.; Minonzio, M.; Shiffer, D.; Cairo, B.; Tank, J.; Jordan, J.; Diedrich, A.; Gauger, P.; Zamuner, A.R.; et al. Cardiac and Vascular Sympathetic Baroreflex Control during Orthostatic Pre-Syncope. J. Clin. Med. 2019, 8, 1434. https://doi.org/10.3390/jcm8091434
Furlan R, Heusser K, Minonzio M, Shiffer D, Cairo B, Tank J, Jordan J, Diedrich A, Gauger P, Zamuner AR, et al. Cardiac and Vascular Sympathetic Baroreflex Control during Orthostatic Pre-Syncope. Journal of Clinical Medicine. 2019; 8(9):1434. https://doi.org/10.3390/jcm8091434
Chicago/Turabian StyleFurlan, Raffaello, Karsten Heusser, Maura Minonzio, Dana Shiffer, Beatrice Cairo, Jens Tank, Jens Jordan, André Diedrich, Peter Gauger, Antonio Roberto Zamuner, and et al. 2019. "Cardiac and Vascular Sympathetic Baroreflex Control during Orthostatic Pre-Syncope" Journal of Clinical Medicine 8, no. 9: 1434. https://doi.org/10.3390/jcm8091434
APA StyleFurlan, R., Heusser, K., Minonzio, M., Shiffer, D., Cairo, B., Tank, J., Jordan, J., Diedrich, A., Gauger, P., Zamuner, A. R., Dipaola, F., Porta, A., & Barbic, F. (2019). Cardiac and Vascular Sympathetic Baroreflex Control during Orthostatic Pre-Syncope. Journal of Clinical Medicine, 8(9), 1434. https://doi.org/10.3390/jcm8091434