NK Cell Reconstitution in Paediatric Leukemic Patients after T-Cell-Depleted HLA-Haploidentical Haematopoietic Stem Cell Transplantation Followed by the Reinfusion of iCasp9-Modified Donor T Cells

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Cell Source
2.2. Multicolour Immunofluorescence and Flow Cytometry Analysis
2.3. Degranulation Assay and IFN-γ Production
2.4. Statistical Analysis
3. Results and Discussion
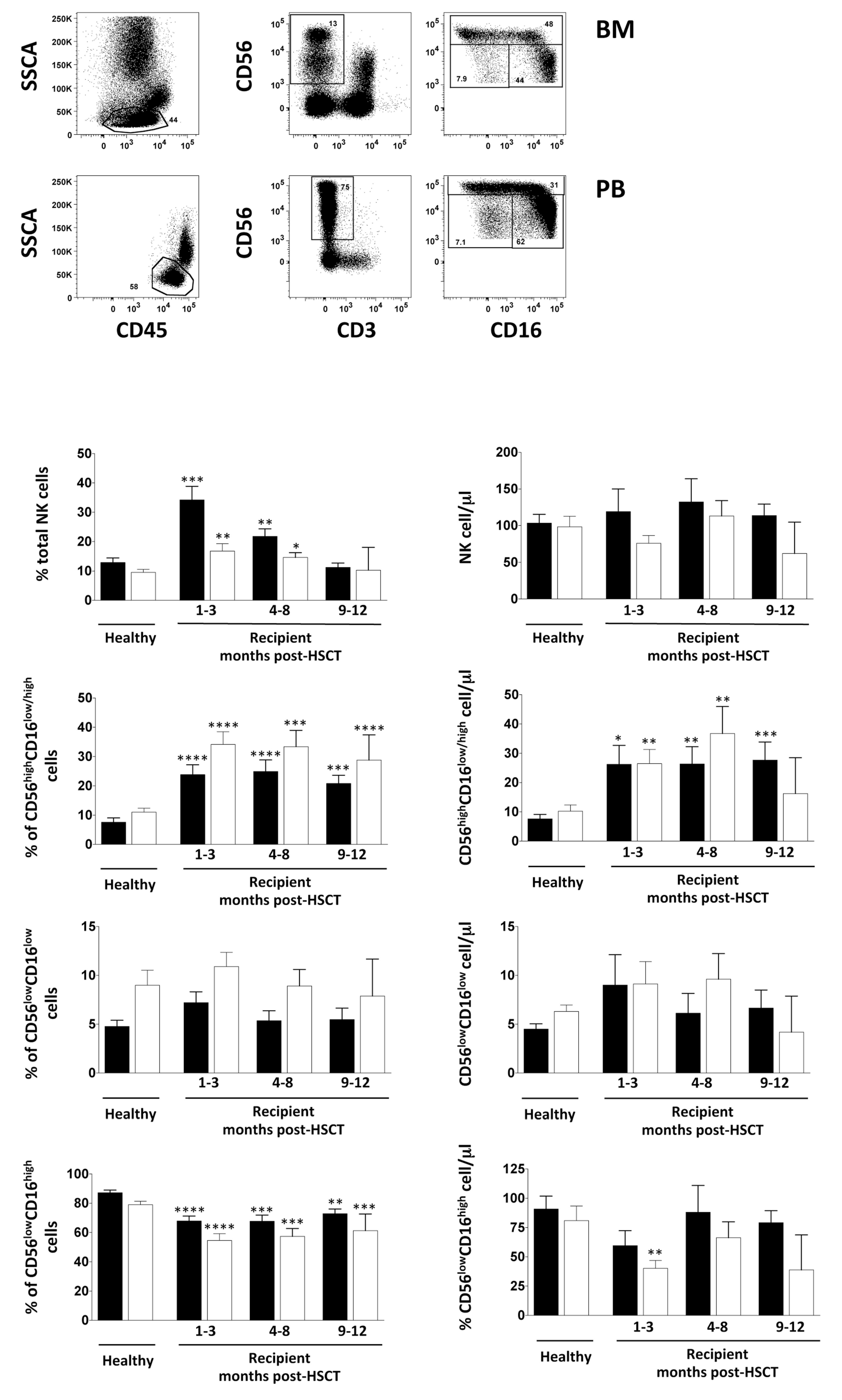
3.1. Impact of iCasp9-Modified Donor T Cells on NK Cell Recovery after HSCT
3.2. NK Cell Distribution in BM and PB of iCasp9-HSCT Patients
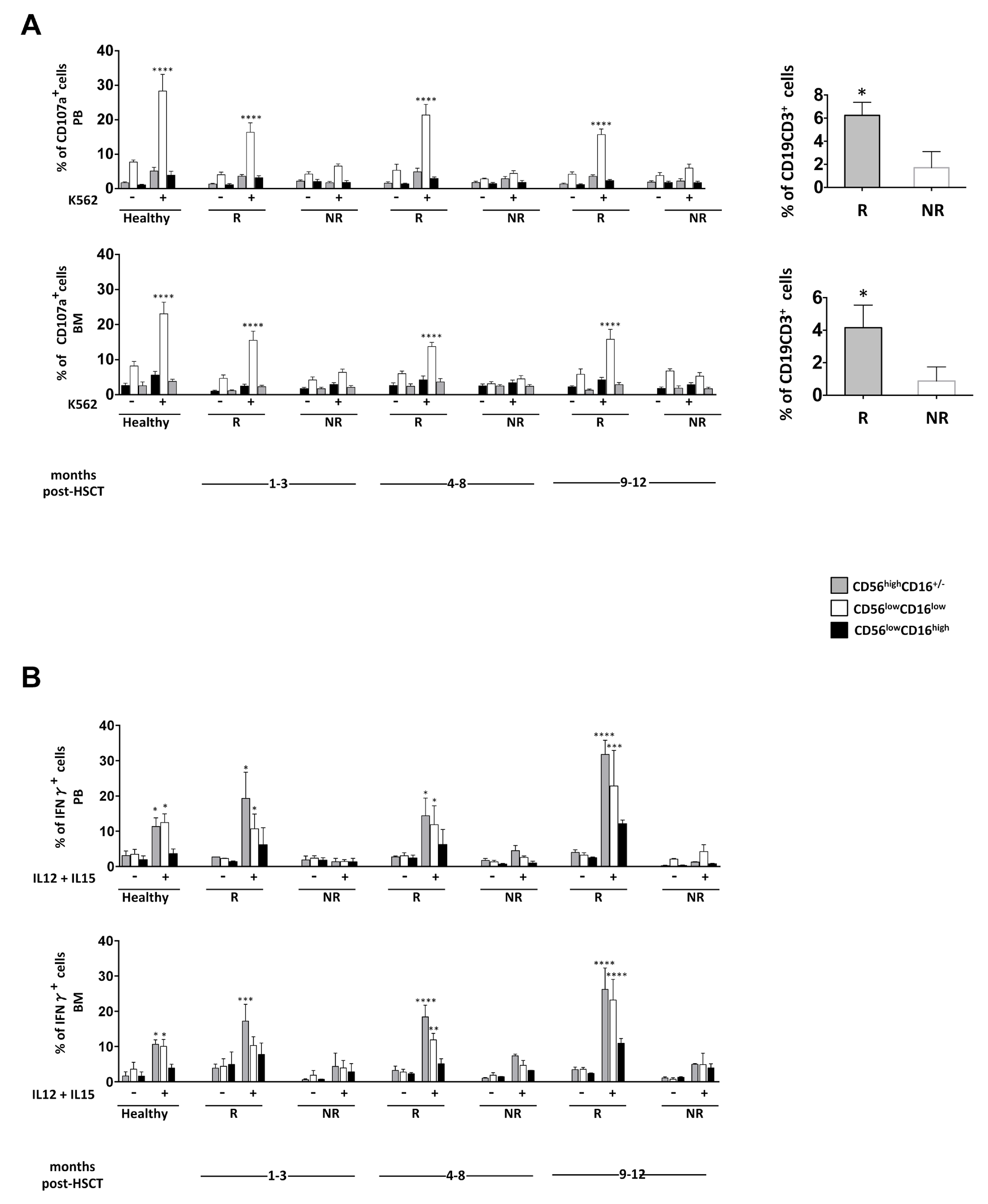
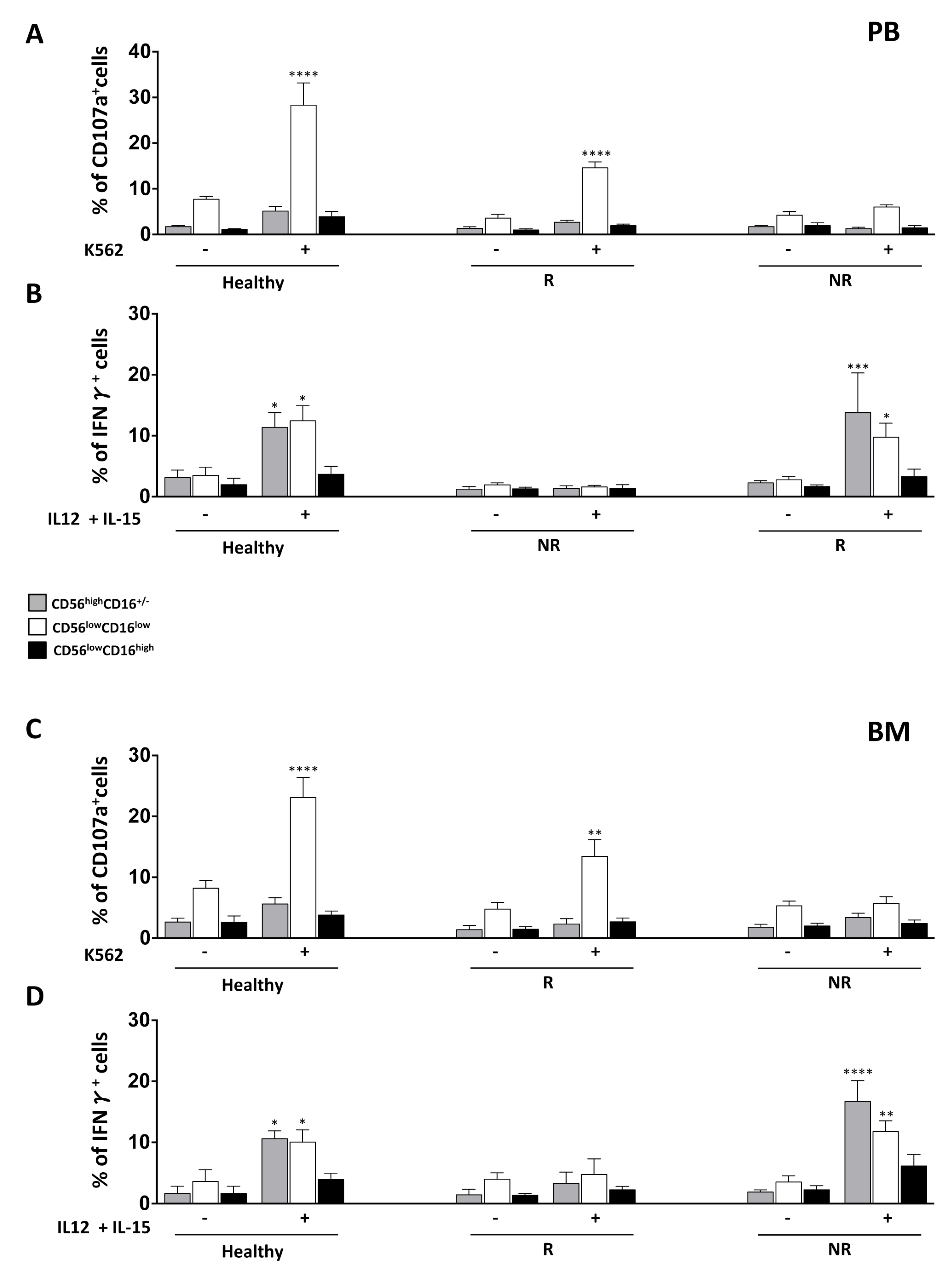
3.3. NK Cell Effector Functions in iCasp9-HSCT Patients
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Spitzer, T.R.; Dey, B.R.; Chen, Y.; Bin Attar, E.; Ballen, K.K. The expanding frontier of hematopoietic cell transplantation. Cytom. Part B Clin. Cytom. 2012, 82, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Ballen, K.K.; Koreth, J.; Chen, Y.; Bin Dey, B.R.; Spitzer, T.R. Selection of optimal alternative graft source: Mismatched unrelated donor, umbilical cord blood, or haploidentical transplant. Blood 2012, 119, 1972–1980. [Google Scholar] [CrossRef] [PubMed]
- Tabilio, A.; Terenzi, A.; Falzetti, F.; Barbabietola, G.; Latini, P.; Felicini, R.; Falcinelli, F.; Carotti, A.; Ballanti, S.; Santucci, A.; et al. Treatment of high-risk acute leukemia with T-cell-depleted stem cells from related donors with one fully mismatched HLA haplotype. N. Engl. J. Med. 1998, 339, 1186–1193. [Google Scholar] [CrossRef]
- Marek, A.; Stern, M.; Chalandon, Y.; Ansari, M.; Ozsahin, H.; Güngör, T.; Gerber, B.; Kühne, T.; Passweg, J.R.; Gratwohl, A.; et al. The impact of T-cell depletion techniques on the outcome after haploidentical hematopoietic SCT. Bone Marrow Transplant. 2014, 49, 55–61. [Google Scholar] [CrossRef]
- O’Reilly, R.J.; Koehne, G.; Hasan, A.N.; Doubrovina, E.; Prockop, S. T-cell depleted allogeneic hematopoietic cell transplants as a platform for adoptive therapy with leukemia selective or virus-specific T-cells. Bone Marrow Transplant. 2015, 50, S43–S50. [Google Scholar] [CrossRef]
- Leen, A.M.; Christin, A.; Myers, G.D.; Liu, H.; Cruz, C.R.; Hanley, P.J.; Kennedy-Nasser, A.A.; Leung, K.S.; Gee, A.P.; Krance, R.A.; et al. Cytotoxic T lymphocyte therapy with donor T cells prevents and treats adenovirus and Epstein-Barr virus infections after haploidentical and matched unrelated stem cell transplantation. Blood 2009, 114, 4283–4292. [Google Scholar] [CrossRef]
- Amrolia, P.J.; Muccioli-Casadei, G.; Huls, H.; Adams, S.; Durett, A.; Gee, A.; Yvon, E.; Weiss, H.; Cobbold, M.; Gaspar, H.B.; et al. Adoptive immunotherapy with allodepleted donor T-cells improves immune reconstitution after haploidentical stem cell transplantation. Blood 2006, 108, 1797–1808. [Google Scholar] [CrossRef]
- Locatelli, F.; Merli, P.; Rutella, S. At the Bedside: Innate immunity as an immunotherapy tool for hematological malignancies. J. Leukoc. Biol. 2013, 94, 1141–1157. [Google Scholar] [CrossRef]
- Tey, S.-K. Adoptive T-cell therapy: Adverse events and safety switches. Clin. Transl. Immunol. 2014, 3, e17. [Google Scholar] [CrossRef]
- Di Stasi, A.; Tey, S.-K.; Dotti, G.; Fujita, Y.; Kennedy-Nasser, A.; Martinez, C.; Straathof, K.; Liu, E.; Durett, A.G.; Grilley, B.; et al. Inducible apoptosis as a safety switch for adoptive cell therapy. N. Engl. J. Med. 2011, 365, 1673–1683. [Google Scholar] [CrossRef]
- Jones, B.S.; Lamb, L.S.; Goldman, F.; Di Stasi, A. Improving the safety of cell therapy products by suicide gene transfer. Front. Pharmacol. 2014, 5, 254. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Di Stasi, A.; Tey, S.K.; Krance, R.A.; Martinez, C.; Leung, K.S.; Durett, A.G.; Wu, M.-F.; Liu, H.; Leen, A.M.; et al. Long-term outcome after haploidentical stem cell transplant and infusion of T cells expressing the inducible caspase 9 safety transgene. Blood 2014, 123, 3895–3905. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, L.; Capanni, M.; Urbani, E.; Shlomchik, W.D.; Posati, S.; Rogaia, D.; Martelli, M.F.; Velardi, A.; Perruccio, K.; Tosti, A.; et al. Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science 2002, 295, 2097–2100. [Google Scholar] [CrossRef] [PubMed]
- Cichocki, F.; Verneris, M.R.; Cooley, S.; Bachanova, V.; Brunstein, C.G.; Blazar, B.R.; Wagner, J.; Schlums, H.; Bryceson, Y.T.; Weisdorf, D.J.; et al. The Past, Present, and Future of NK Cells in Hematopoietic Cell Transplantation and Adoptive Transfer. Curr. Top. Microbiol. Immunol. 2015. [Google Scholar] [CrossRef]
- Foley, B.; Cooley, S.; Verneris, M.R.; Pitt, M.; Curtsinger, J.; Luo, X.; Lopez-Vergès, S.; Lanier, L.L.; Weisdorf, D.; Miller, J.S. Cytomegalovirus reactivation after allogeneic transplantation promotes a lasting increase in educated NKG2C+ natural killer cells with potent function. Blood 2012, 119, 2665–2674. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.; Guethlein, L.A.; Nemat-Gorgani, N.; Norman, P.J.; Cooley, S.; Miller, J.S.; Parham, P. Regulation of Adaptive NK Cells and CD8 T Cells by HLA-C Correlates with Allogeneic Hematopoietic Cell Transplantation and with Cytomegalovirus Reactivation. J. Immunol. 2015, 195, 4524–4536. [Google Scholar] [CrossRef]
- Muccio, L.; Bertaina, A.; Falco, M.; Pende, D.; Meazza, R.; Lopez-Botet, M.; Moretta, L.; Locatelli, F.; Moretta, A.; Chiesa, M.D. Analysis of memory-like natural killer cells in human cytomegalovirus-infected children undergoing +T and B cell-depleted hematopoietic stem cell transplantation for hematological malignancies. Haematologica 2016, 101, 371–381. [Google Scholar] [CrossRef]
- Cichocki, F.; Schlums, H.; Theorell, J.; Tesi, B.; Miller, J.S.; Ljunggren, H.-G.; Bryceson, Y.T. Diversification and Functional Specialization of Human NK Cell Subsets. Curr. Top. Microbiol. Immunol. 2015, 358, 63–93. [Google Scholar] [CrossRef]
- Stabile, H.; Fionda, C.; Gismondi, A.; Santoni, A. Role of distinct natural killer cell subsets in anticancer response. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Stabile, H.; Nisti, P.; Morrone, S.; Pagliara, D.; Bertaina, A.; Locatelli, F.; Santoni, A.; Gismondi, A. Multifunctional human CD56lowCD16lownatural killer cells are the prominent subset in bone marrow of both healthy pediatric donors and leukemic patients. Haematologica 2015, 100, 489–498. [Google Scholar] [CrossRef]
- Vulpis, E.; Stabile, H.; Soriani, A.; Fionda, C.; Petrucci, M.T.; Mariggio’, E.; Ricciardi, M.R.; Cippitelli, M.; Gismondi, A.; Santoni, A.; et al. Key role of the CD56 low CD16 low natural killer cell subset in the recognition and killing of multiple myeloma cells. Cancers 2018, 10, 473. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.A.; Hill, G.R.; Tey, S.-K. Functional Reconstitution of Natural Killer Cells in Allogeneic Hematopoietic Stem Cell Transplantation. Front. Immunol. 2016, 7, 144. [Google Scholar] [CrossRef] [PubMed]
- Helena, S.; Paolo, N.; Giovanna, P.; Cinzia, F.; Daria, P.; Letizia, P.B.; Pietro, M.; Franco, L.; Santoni, A.; Gismondi, A. Reconstitution of multifunctional CD56lowCD16low natural killer cell subset in children with acute leukemia given α/β T cell-depleted HLA-haploidentical haematopoietic stem cell transplantation. Oncoimmunology 2017, 6. [Google Scholar] [CrossRef]
- Björklund, A.T.; Schaffer, M.; Fauriat, C.; Ringdén, O.; Remberger, M.; Hammarstedt, C.; Barrett, A.J.; Ljungman, P.; Ljunggren, H.-G.; Malmberg, K.-J. NK cells expressing inhibitory KIR for non-self-ligands remain tolerant in HLA-matched sibling stem cell transplantation. Blood 2010, 115, 2686–2694. [Google Scholar] [CrossRef]
- Dulphy, N.; Haas, P.; Busson, M.; Belhadj, S.; De Latour, R.P.; Robin, M.; Carmagnat, M.; Loiseau, P.; Tamouza, R.; Scieux, C.; et al. An unusual CD56(bright) CD16(low) NK cell subset dominates the early posttransplant period following HLA-matched hematopoietic stem cell transplantation. J. Immunol. 2008, 181, 2227–2237. [Google Scholar] [CrossRef]
- Cooley, S.; McCullar, V.; Wangen, R.; Bergemann, T.L.; Spellman, S.; Weisdorf, D.J.; Miller, J.S. KIR reconstitution is altered by T cells in the graft and correlates with clinical outcomes after unrelated donor transplantation. Blood 2005, 106, 4370–4376. [Google Scholar] [CrossRef]
- Martínez, C.; Urbano-Ispizua, A.; Rozman, C.; Marín, P.; Rovira, M.; Sierra, J.; Montfort, N.; Carreras, E.; Montserrat, E. Immune reconstitution following allogeneic peripheral blood progenitor cell transplantation: Comparison of recipients of positive CD34+ selected grafts with recipients of unmanipulated grafts. Exp. Hematol. 1999, 27, 561–568. [Google Scholar] [CrossRef]
- Vago, L.; Forno, B.; Sormani, M.P.; Crocchiolo, R.; Zino, E.; Di Terlizzi, S.; Stanghellini, M.T.L.; Mazzi, B.; Perna, S.K.; Bondanza, A.; et al. Temporal, quantitative, and functional characteristics of single-KIR-positive alloreactive natural killer cell recovery account for impaired graft-versus-leukemia activity after haploidentical hematopoietic stem cell transplantation. Blood 2008, 112, 3488–3499. [Google Scholar] [CrossRef]
- Björkström, N.K.; Riese, P.; Heuts, F.; Fauriat, C.; Ivarsson, M.A.; Björklund, A.T.; Flodström-Tullberg, M.; Michaëlsson, J.; Rottenberg, M.E.; Guzman, C.A.; et al. Expression patterns of NKG2A, KIR, and CD57 define a process of CD56 dim NK-cell differentiation uncoupled from NK-cell education. Blood 2010, 116, 3853–3864. [Google Scholar] [CrossRef]
- Foley, B.; Cooley, S.; Verneris, M.R.; Curtsinger, J.; Luo, X.; Waller, E.K.; Weisdorf, D.J.; Miller, J.S. NK cell education after allogeneic transplantation: Dissociation between recovery of cytokine-producing and cytotoxic functions. Blood 2011, 118, 2784–2792. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stabile, H.; Nisti, P.; Fionda, C.; Pagliara, D.; Gaspari, S.; Locatelli, F.; Santoni, A.; Gismondi, A. NK Cell Reconstitution in Paediatric Leukemic Patients after T-Cell-Depleted HLA-Haploidentical Haematopoietic Stem Cell Transplantation Followed by the Reinfusion of iCasp9-Modified Donor T Cells. J. Clin. Med. 2019, 8, 1904. https://doi.org/10.3390/jcm8111904
Stabile H, Nisti P, Fionda C, Pagliara D, Gaspari S, Locatelli F, Santoni A, Gismondi A. NK Cell Reconstitution in Paediatric Leukemic Patients after T-Cell-Depleted HLA-Haploidentical Haematopoietic Stem Cell Transplantation Followed by the Reinfusion of iCasp9-Modified Donor T Cells. Journal of Clinical Medicine. 2019; 8(11):1904. https://doi.org/10.3390/jcm8111904
Chicago/Turabian StyleStabile, Helena, Paolo Nisti, Cinzia Fionda, Daria Pagliara, Stefania Gaspari, Franco Locatelli, Angela Santoni, and Angela Gismondi. 2019. "NK Cell Reconstitution in Paediatric Leukemic Patients after T-Cell-Depleted HLA-Haploidentical Haematopoietic Stem Cell Transplantation Followed by the Reinfusion of iCasp9-Modified Donor T Cells" Journal of Clinical Medicine 8, no. 11: 1904. https://doi.org/10.3390/jcm8111904
APA StyleStabile, H., Nisti, P., Fionda, C., Pagliara, D., Gaspari, S., Locatelli, F., Santoni, A., & Gismondi, A. (2019). NK Cell Reconstitution in Paediatric Leukemic Patients after T-Cell-Depleted HLA-Haploidentical Haematopoietic Stem Cell Transplantation Followed by the Reinfusion of iCasp9-Modified Donor T Cells. Journal of Clinical Medicine, 8(11), 1904. https://doi.org/10.3390/jcm8111904