Heavy Water (D2O) Containing Preservation Solution Reduces Hepatic Cold Preservation and Reperfusion Injury in an Isolated Perfused Rat Liver (IPRL) Model

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of Preservation Solutions
2.3. Animals
2.4. Procedures
2.5. Experimental Protocol
2.6. Isolated Perfused Rat Liver (IPRL)
2.7. Sample Collection
2.8. IPRL Calculations
2.9. Histology and Apoptotic Index
2.10. ATP and Energy Charge Assay
2.11. Glutathione Assay
2.12. Western Blot Analysis
2.13. Statistical Analysis
3. Results
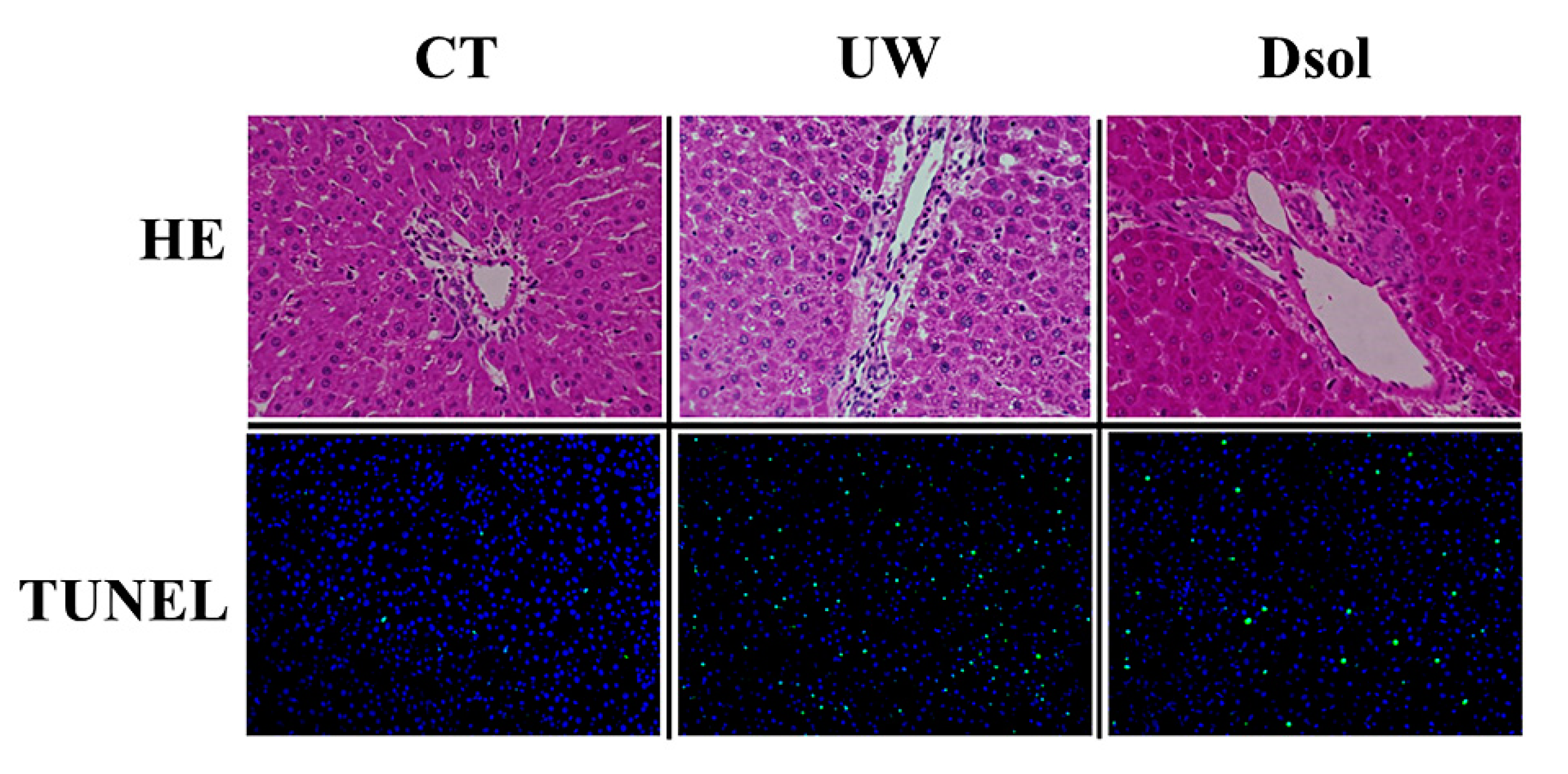
3.1. Liver Histopathology and TUNEL Staining
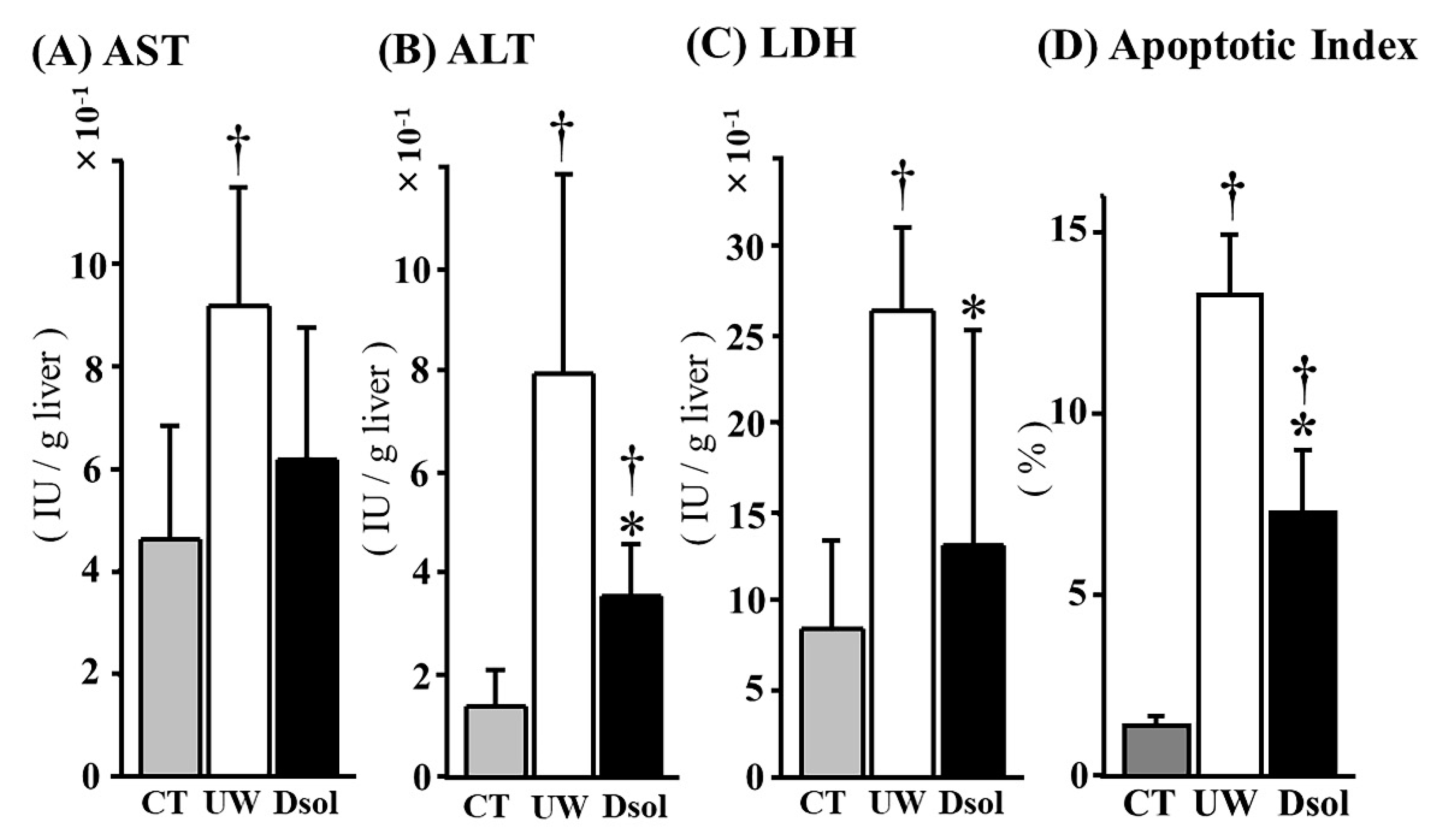
3.2. Liver Function Tests and Apoptotic Cell Ratio
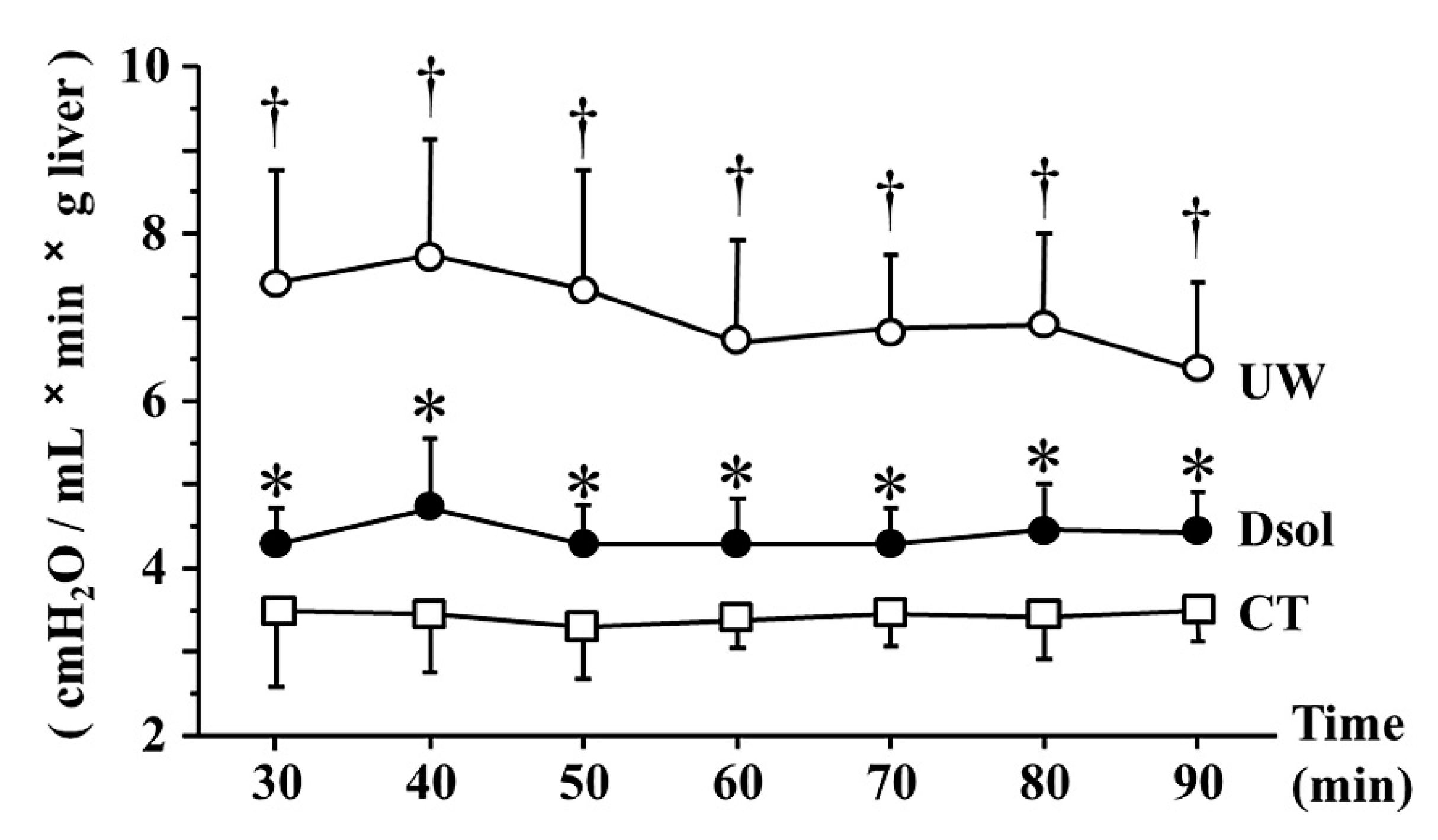
3.3. Portal Venous Resistance (PVR)
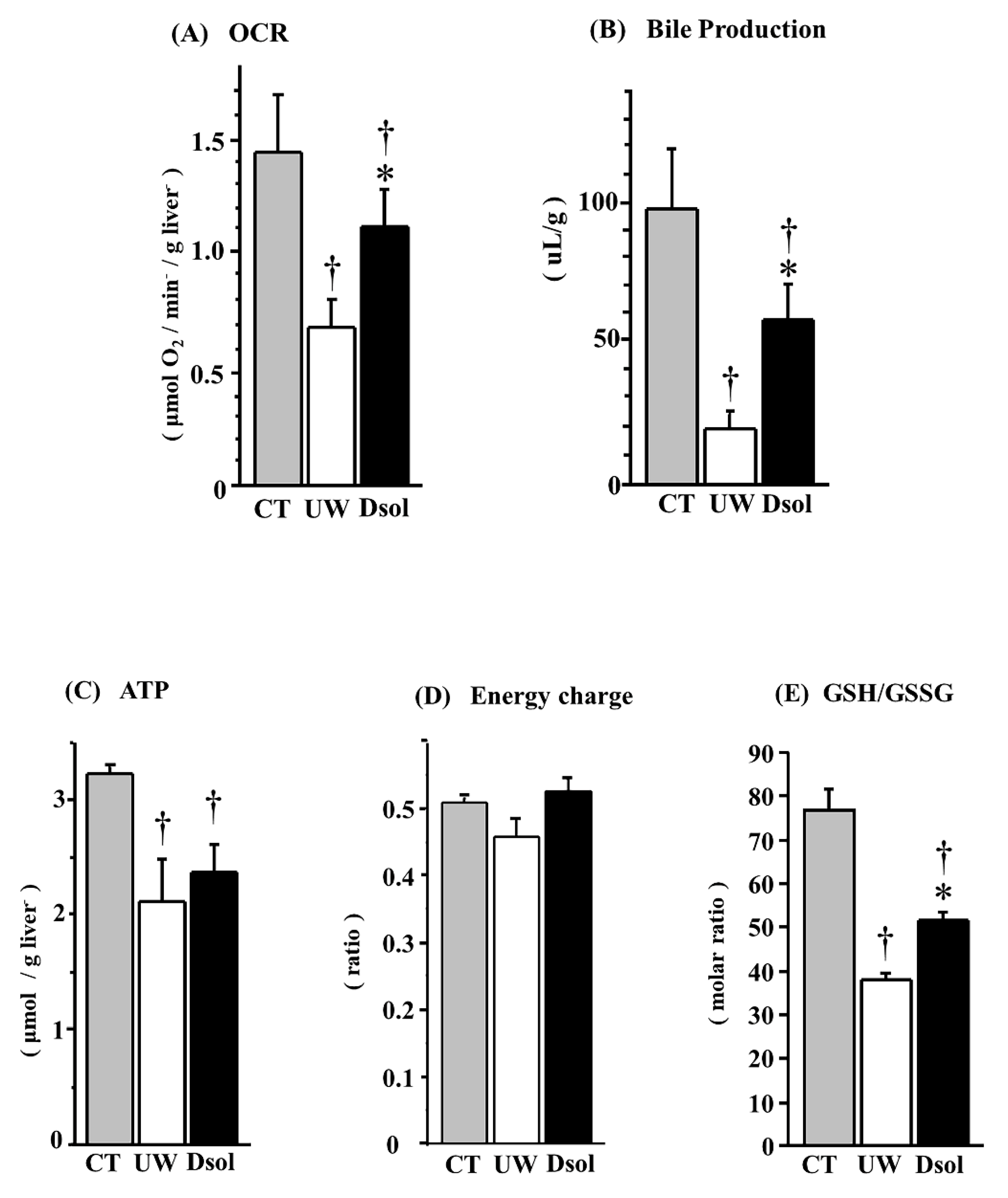
3.4. Oxygen Consumption Rate (OCR)
3.5. Bile Production
3.6. Energy Status
3.7. Redox Status of Glutathione
3.8. Cytoskeletal Derangement
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hoyer, D.P.; Paul, A.; Gallinat, A.; Molmenti, E.P.; Reinhardt, R.; Minor, T.; Saner, F.H.; Canbay, A.; Treckmann, J.W.; Sotiropoulos, G.C.; et al. Donor information based prediction of early allograft dysfunction and outcome in liver transplantation. Liver Int. 2015, 35, 156–163. [Google Scholar]
- Agopian, V.G.; Petrowsky, H.; Kaldas, F.M.; Zarrinpar, A.; Farmer, D.G.; Yersiz, H.; Holt, C.; Harlander-Locke, M.; Hong, J.C.; Rana, A.R.; et al. The evolution of liver transplantation during 3 decades: Analysis of 5347 consecutive liver transplants at a single center. Ann. Surg. 2013, 258, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Minor, T.; Isselhard, W. Synthesis of high energy phosphates during cold ischemic rat liver preservation with gaseous oxygen insufflation. Transplantation 1996, 61, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Wyllie, S.; Seu, P.; Gao, F.Q.; Goss, J.A. Deregulation of iron homeostasis and cold-preservation injury to rat liver stored in University of Wisconsin solution. Liver Transplant. 2003, 9, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Belous, A.; Knox, C.; Nicoud, I.B.; Pierce, J.; Anderson, C.; Pinson, C.W.; Chari, R.S. Altered ATP-dependent mitochondrial Ca2+ uptake in cold ischemia is attenuated by ruthenium red. J. Surg. Res. 2003, 111, 284–289. [Google Scholar] [CrossRef]
- Tian, Y.H.; Fukuda, C.; Schilling, M.K. Interstitial accumulation of Na+ and K+ during flush-out and cold storage of rat livers: Implications for graft survival. Hepatology 1998, 28, 1327–1331. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Southard, J.H. Membrane stabilizing effects of calcium and taxol during the cold storage of isolated rat hepatocytes. Transplantation 1999, 68, 938–943. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Antoine, D.J.; Weemhoff, J.L.; Jenkins, R.E.; Farhood, A.; Park, B.K.; Jaeschke, H. Biomarkers distinguish apoptotic and necrotic cell death during hepatic ischemia/reperfusion injury in mice. Liver Transplant. 2014, 20, 1372–1382. [Google Scholar] [CrossRef] [PubMed]
- Shimada, S.; Wakayama, K.; Fukai, M.; Shimamura, T.; Ishikawa, T.; Fukumori, D.; Shibata, M.; Yamashita, K.; Kimura, T.; Todo, S.; et al. Hydrogen gas ameliorates hepatic reperfusion injury after prolonged cold preservation in isolated perfused rat liver. Artif. Organs 2016, 40, 1128–1136. [Google Scholar] [CrossRef]
- Wakayama, K.; Fukai, M.; Yamashita, K.; Kimura, T.; Hirokata, G.; Shibasaki, S.; Fukumori, D.; Haga, S.; Sugawara, M.; Suzuki, T.; et al. Successful transplantation of rat hearts subjected to extended cold preservation with a novel preservation solution. Transpl. Int. 2012, 25, 696–706. [Google Scholar] [CrossRef]
- Marsland, D.; Tilney, L.G.; Hirshfield, M. Stabilizing effects of D2O on the microtubular components and needle-like form of heliozoan axopods: A pressure-temperature analysis. J. Cell. Physiol. 1971, 77, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Omori, H.; Kuroda, M.; Naora, H.; Takeda, H.; Nio, Y.; Otani, H.; Tamura, K. Deuterium oxide (heavy water) accelerates actin assembly in vitro and changes microfilament distribution in cultured cells. Eur. J. Cell Biol. 1997, 74, 273–280. [Google Scholar] [PubMed]
- Wenzel, M.; Fischer, J.H. Organ preservation with heavy water—Effect of D2O concentration and temperature on organ swelling. J. Clin. Chem. Clin. Biochem. 1983, 21, 83–89. [Google Scholar] [PubMed]
- Wals, P.A.; Katz, J. The effect of D2O on glycolysis by rat hepatocytes. Int. J. Biochem. 1993, 25, 1561–1564. [Google Scholar] [CrossRef]
- Urbauer, J.L.; Dorgan, L.J.; Schuster, S.M. Effects of deuterium on the kinetics of beef heart mitochondrial ATPase. Arch. Biochem. Biophys. 1984, 231, 498–502. [Google Scholar] [CrossRef]
- Fukai, M.; Hayashi, T.; Yokota, R.; Shimamura, T.; Suzuki, T.; Taniguchi, M.; Matsushita, M.; Furukawa, H.; Todo, S. Lipid peroxidation during ischemia depends on ischemia time in warm ischemia and reperfusion of rat liver. Free Radic. Biol. Med. 2005, 38, 1372–1381. [Google Scholar] [CrossRef]
- Gnaiger, E. Oxygen solubility in experimental media. Mitochondrial Physiol. Netw. 2001, 6, 1–6. [Google Scholar]
- Shimada, S.; Fukai, M.; Wakayama, K.; Ishikawa, T.; Kobayashi, N.; Kimura, T.; Yamashita, K.; Kamiyama, T.; Shimamura, T.; Taketomi, A.; et al. Hydrogen sulfide augments survival signals in warm ischemia and reperfusion of the mouse liver. Surg. Today 2015, 45, 892–903. [Google Scholar] [CrossRef]
- Fukuda, C.; Kollmar, O.; Schafer, T.; Tian, Y.H.; Schilling, M.K. Anionic polysaccharides. A class of substances with hepatoprotective and antiadhesive properties in rat liver preservation. Transpl. Int. 2002, 15, 17–23. [Google Scholar]
- Wenzel, M.; Hölscher, B.; Günther, T.; Merker, H.J. Organ preservation by heavy water (D2O). Morphological and biochemical studies on heart and liver (author’s transl). J. Clin. Chem. Clin. Biochem. 1979, 17, 123–128. [Google Scholar]
- Fischer, J.H.; Knupfer, P.; Beyer, M. Flush solution 2, a new concept for one-to-three-day hypothermic renal storage preservation. Functional recovery after preservation in Euro-Collins, Collins’ C2, hypertonic citrate, and F.2 solution. Transplantation 1985, 39, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Hesse, U.J.; Gores, P.F.; Florack, G.; Sutherland, D.E. The use of D2O (heavy water)-based solution for hypothermic preservation of the pancreas. Transplant. Proc. 1987, 19, 4167. [Google Scholar] [PubMed]
- Inserte, J.; Garcia-Dorado, D.; Hernando, V.; Barba, I.; Soler-Soler, J. Ischemic preconditioning prevents calpain-mediated impairment of Na+/K+-ATPase activity during early reperfusion. Cardiovasc. Res. 2006, 70, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Halestrap, A.P. Calcium, mitochondria and reperfusion injury: A pore way to die. Biochem. Soc. Trans. 2006, 34, 232–237. [Google Scholar] [CrossRef]
- Kohli, V.; Gao, W.; Camargo, C.A.; Clavien, P.A. Calpain is a mediator of preservation-reperfusion injury in rat liver transplantation. Proc. Natl. Acad. Sci. USA 1997, 94, 9354–9359. [Google Scholar] [CrossRef]
- Wang, M.; Sakon, M.; Miyoshi, H.; Umeshita, K.; Kishimoto, S.; Taniguchi, K.; Gotoh, M.; Imajoh-Ohmi, S.; Monden, M. Prostacyclin analog-suppressed ischemia-reperfusion injury of the rat liver: Evaluation by calpain mu activation. J. Surg. Res. 1997, 73, 101–106. [Google Scholar] [CrossRef]
- Tsuji, T.; Ohga, Y.; Yoshikawa, Y.; Sakata, S.; Abe, T.; Tabayashi, N.; Kobayashi, S.; Kohzuki, H.; Yoshida, K.I.; Suga, H.; et al. Rat cardiac contractile dysfunction induced by Ca2+ overload: Possible link to the proteolysis of alpha-fodrin. Am. J. Physiol. Heart Circ. Physiol. 2001, 281, H1286–H1294. [Google Scholar] [CrossRef]
- Shinohara, H.; Tanaka, A.; Fujimoto, T.; Hatano, E.; Satoh, S.; Fujimoto, K.; Noda, T.; Ide, C.; Yamaoka, Y. Disorganization of microtubular network in postischemic liver dysfunction: Its functional and morphological changes. Biochim. Biophys. Acta 1996, 1317, 27–35. [Google Scholar] [CrossRef]
- Zaouali, M.A.; Bejaoui, M.; Calvo, M.; Folch-Puy, E.; Pantazi, E.; Pasut, G.; Rimola, A.; Ben Abdennebi, H.; Adam, R.; Roselló-Catafau, J. Polyethylene glycol rinse solution: An effective way to prevent ischemia-reperfusion injury. World J. Gastroenterol. 2014, 20, 16203–16214. [Google Scholar] [CrossRef]
- Ono, S. Mechanism of depolymerization and severing of actin filaments and its significance in cytoskeletal dynamics. Int. Rev. Cytol. 2007, 258, 1–82. [Google Scholar]
- Niwa, R.; Nagata-Ohashi, K.; Takeichi, M.; Mizuno, K.; Uemura, T. Control of actin reorganization by Slingshot, a family of phosphatases that dephosphorylate ADF/cofilin. Cell 2002, 108, 233–246. [Google Scholar] [CrossRef]
- Yang, N.; Higuchi, O.; Ohashi, K.; Nagata, K.; Wada, A.; Kangawa, K.; Nishida, E.; Mizuno, K. Cofilin phosphorylation by LIM-kinase 1 and its role in Rac-mediated actin reorganization. Nature 1998, 393, 809–812. [Google Scholar] [CrossRef] [PubMed]
- Bellini, M.I.; Yiu, J.; Nozdrin, M.; Papalois, V. The effect of preservation temperature on liver, kidney, and pancreas tissue ATP in animal and preclinical human models. J. Clin. Med. 2019, 8, 1421. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Dose |
|---|---|
| Additives (mM) | |
| NaOH | 125 |
| MgSO4 | 5 |
| KH2PO4 | 25 |
| Lactobionate | 100 |
| Sucrose | 20 |
| Mannitol | 10 |
| Adenosine | 5 |
| Allopurinol | 1 |
| Glutathione | 3 |
| Solvent (%) | |
| H2O | 70 |
| D2O | 30 |
| Freezing point (℃) | 0.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimada, S.; Fukai, M.; Shibata, K.; Sakamoto, S.; Wakayama, K.; Ishikawa, T.; Kawamura, N.; Fujiyoshi, M.; Shimamura, T.; Taketomi, A. Heavy Water (D2O) Containing Preservation Solution Reduces Hepatic Cold Preservation and Reperfusion Injury in an Isolated Perfused Rat Liver (IPRL) Model. J. Clin. Med. 2019, 8, 1818. https://doi.org/10.3390/jcm8111818
Shimada S, Fukai M, Shibata K, Sakamoto S, Wakayama K, Ishikawa T, Kawamura N, Fujiyoshi M, Shimamura T, Taketomi A. Heavy Water (D2O) Containing Preservation Solution Reduces Hepatic Cold Preservation and Reperfusion Injury in an Isolated Perfused Rat Liver (IPRL) Model. Journal of Clinical Medicine. 2019; 8(11):1818. https://doi.org/10.3390/jcm8111818
Chicago/Turabian StyleShimada, Shingo, Moto Fukai, Kengo Shibata, Sodai Sakamoto, Kenji Wakayama, Takahisa Ishikawa, Norio Kawamura, Masato Fujiyoshi, Tsuyoshi Shimamura, and Akinobu Taketomi. 2019. "Heavy Water (D2O) Containing Preservation Solution Reduces Hepatic Cold Preservation and Reperfusion Injury in an Isolated Perfused Rat Liver (IPRL) Model" Journal of Clinical Medicine 8, no. 11: 1818. https://doi.org/10.3390/jcm8111818
APA StyleShimada, S., Fukai, M., Shibata, K., Sakamoto, S., Wakayama, K., Ishikawa, T., Kawamura, N., Fujiyoshi, M., Shimamura, T., & Taketomi, A. (2019). Heavy Water (D2O) Containing Preservation Solution Reduces Hepatic Cold Preservation and Reperfusion Injury in an Isolated Perfused Rat Liver (IPRL) Model. Journal of Clinical Medicine, 8(11), 1818. https://doi.org/10.3390/jcm8111818