The Effect of Interleukin-4 and Dexamethasone on RNA-Seq-Based Transcriptomic Profiling of Human Podocytes: A Potential Role in Minimal Change Nephrotic Syndrome

 ,
,  , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture of Human Podocytes
2.2. Podocyte Treatment with IL-4 and Visualization and Quantification of Intracytoplasmic Actin Filaments
2.3. Podocyte Treatment with IL-4 with or without Dexamethasone
2.4. Whole Transcriptome Sequencing
2.5. Data Analysis and Statistical Modeling
2.5.1. Principal Component Analysis
2.5.2. Identification of DEGs
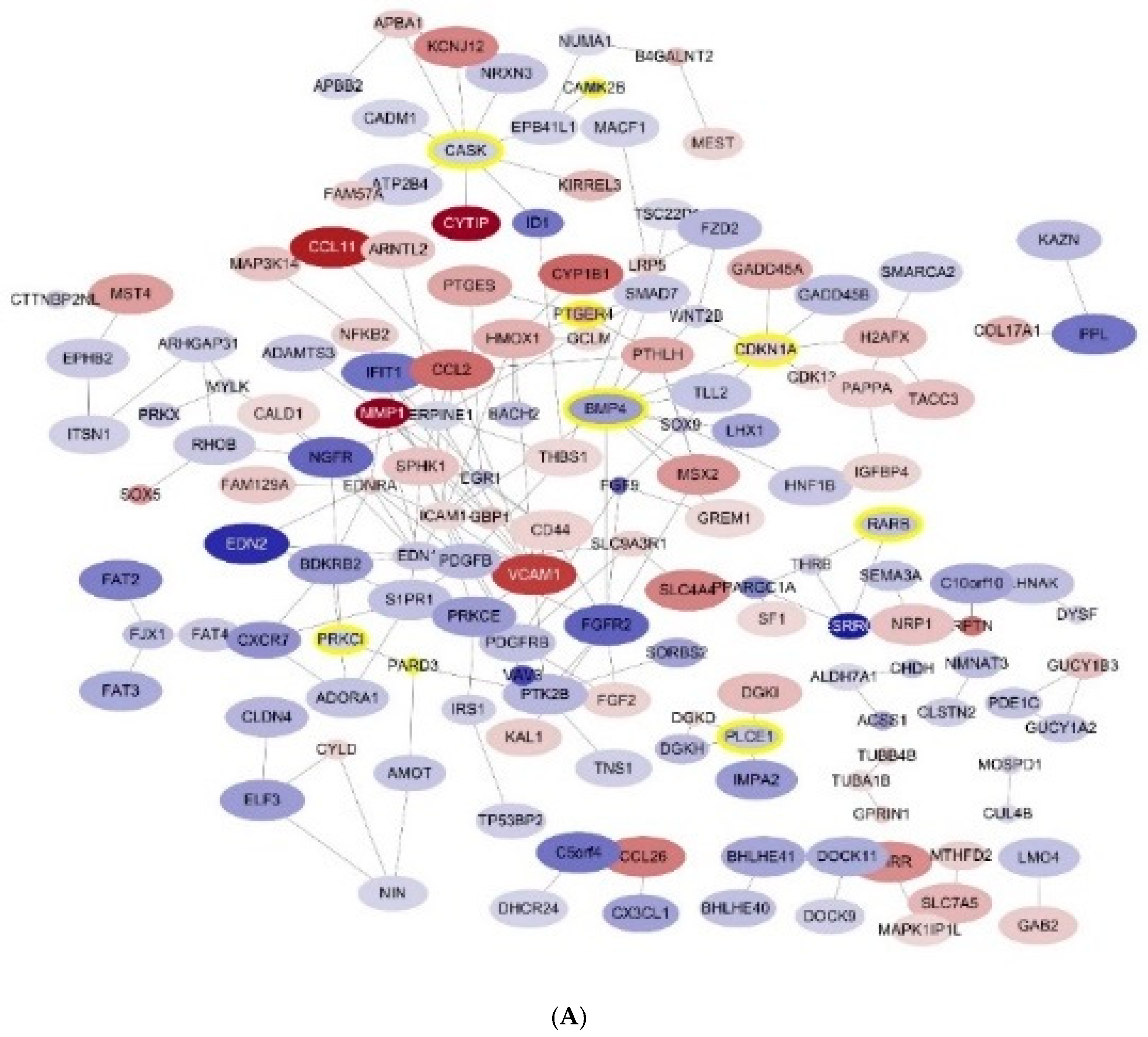
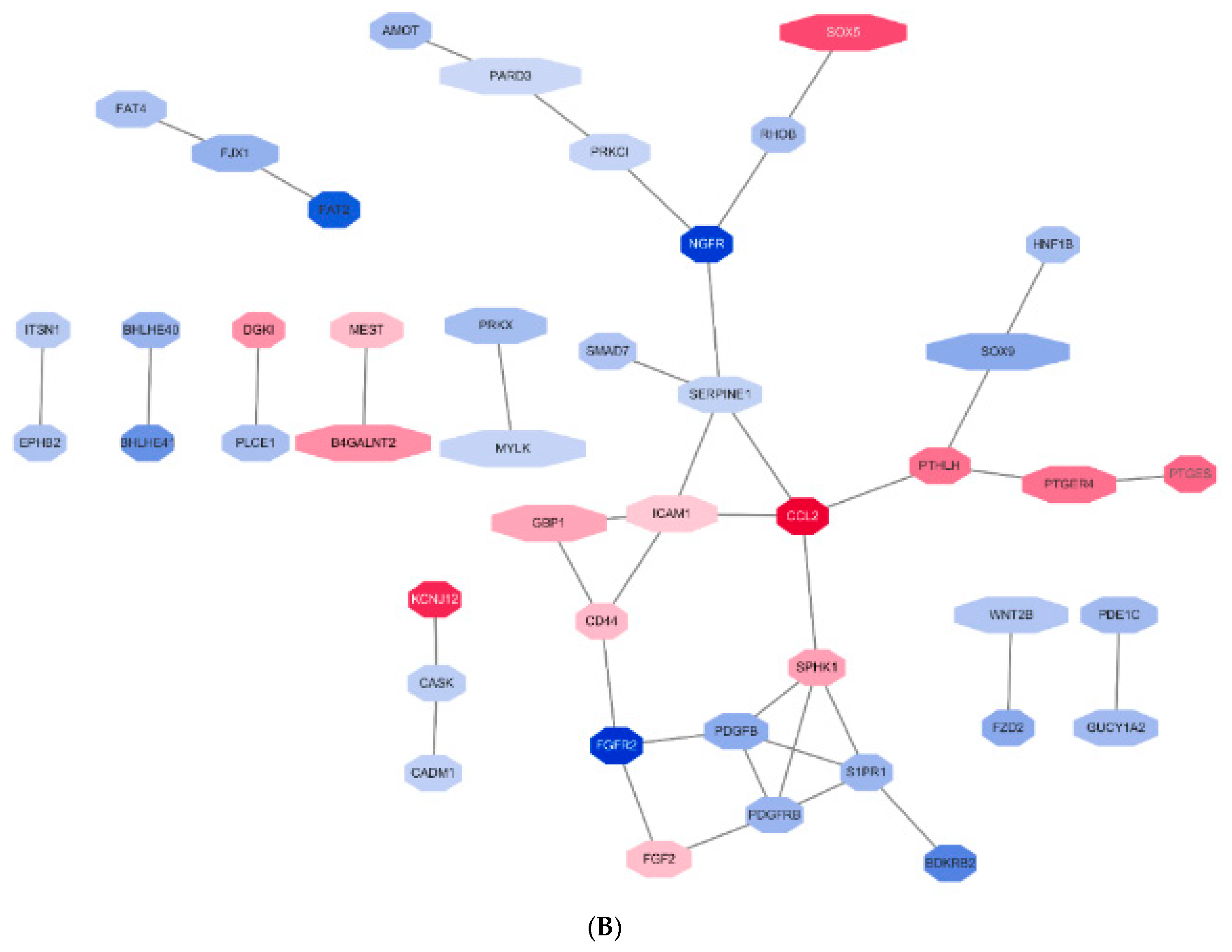
2.5.3. Construction of Protein-Protein Interaction Networks
2.5.4. Gene Ontology Enrichment Analysis
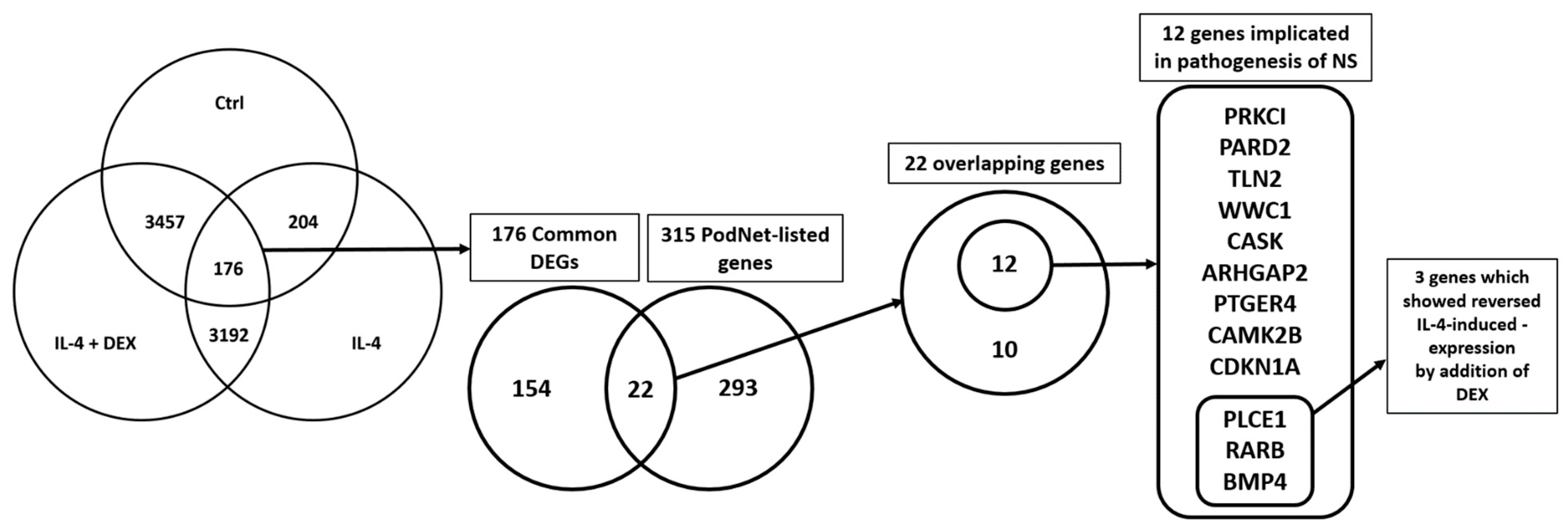
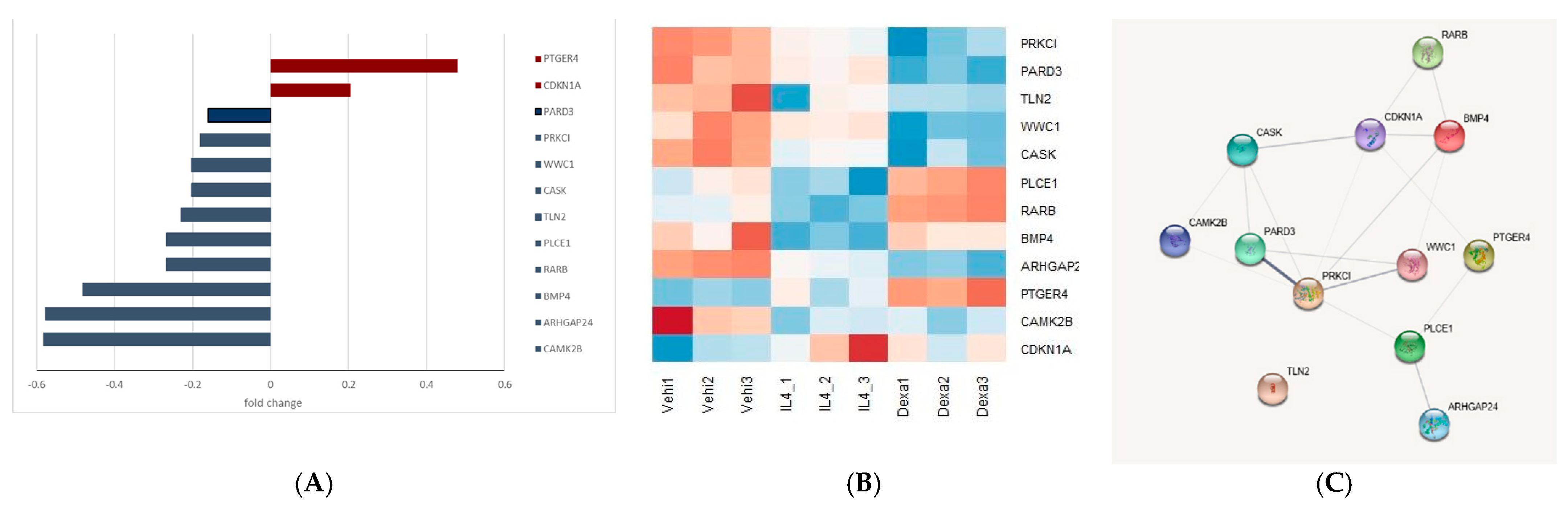
2.5.5. Selection of the Genes of Potential Relevance
3. Results
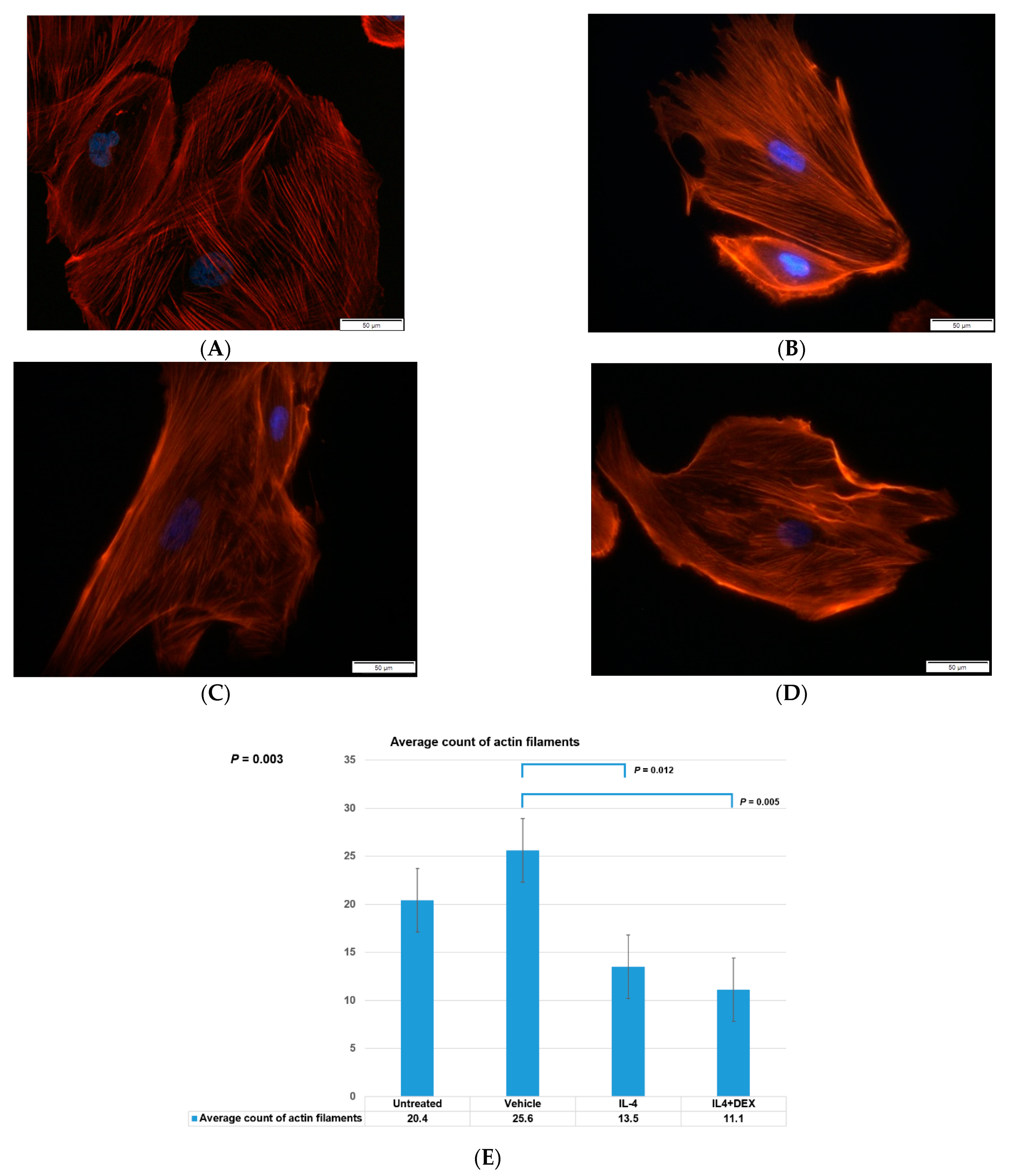
3.1. Visualization of Intracytoplasmic Actin Filaments in Human Podocytes
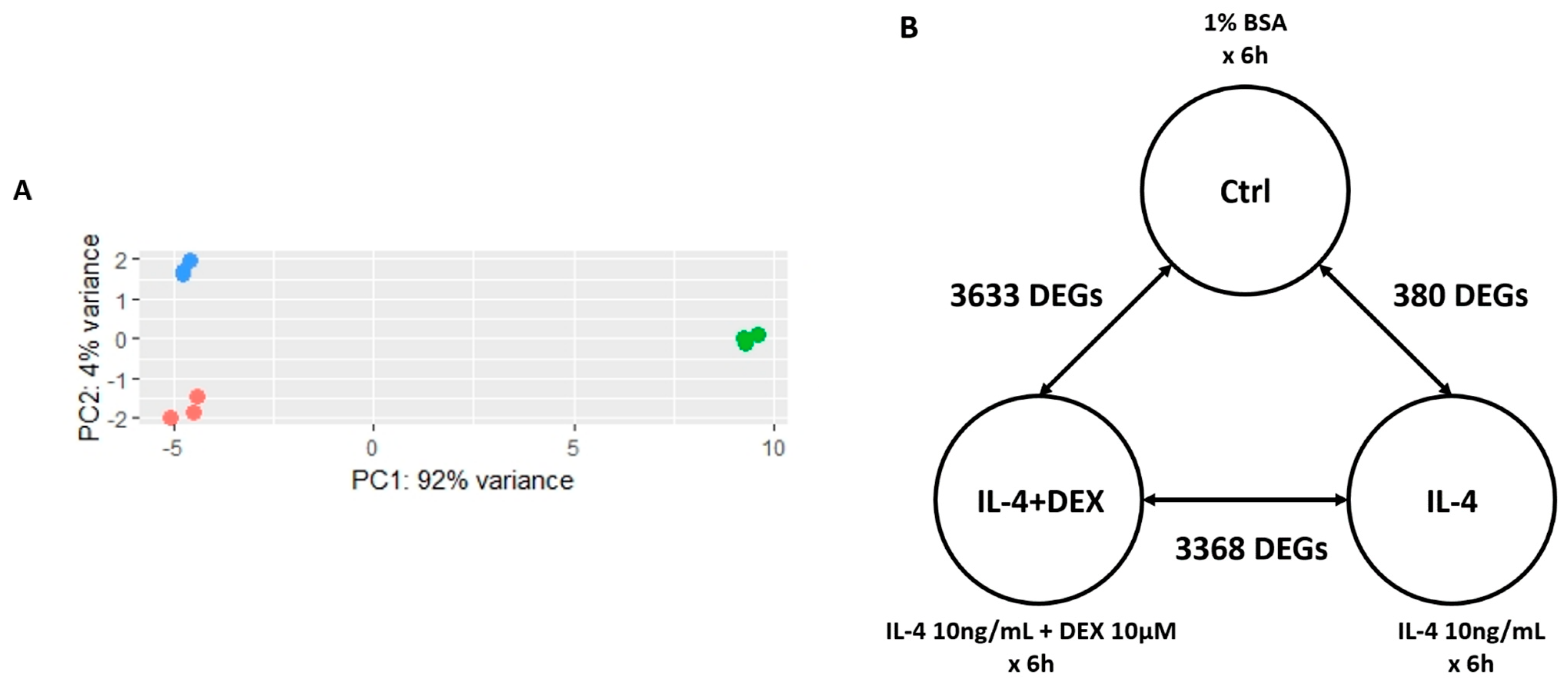
3.2. PCA and Analysis of DEGs
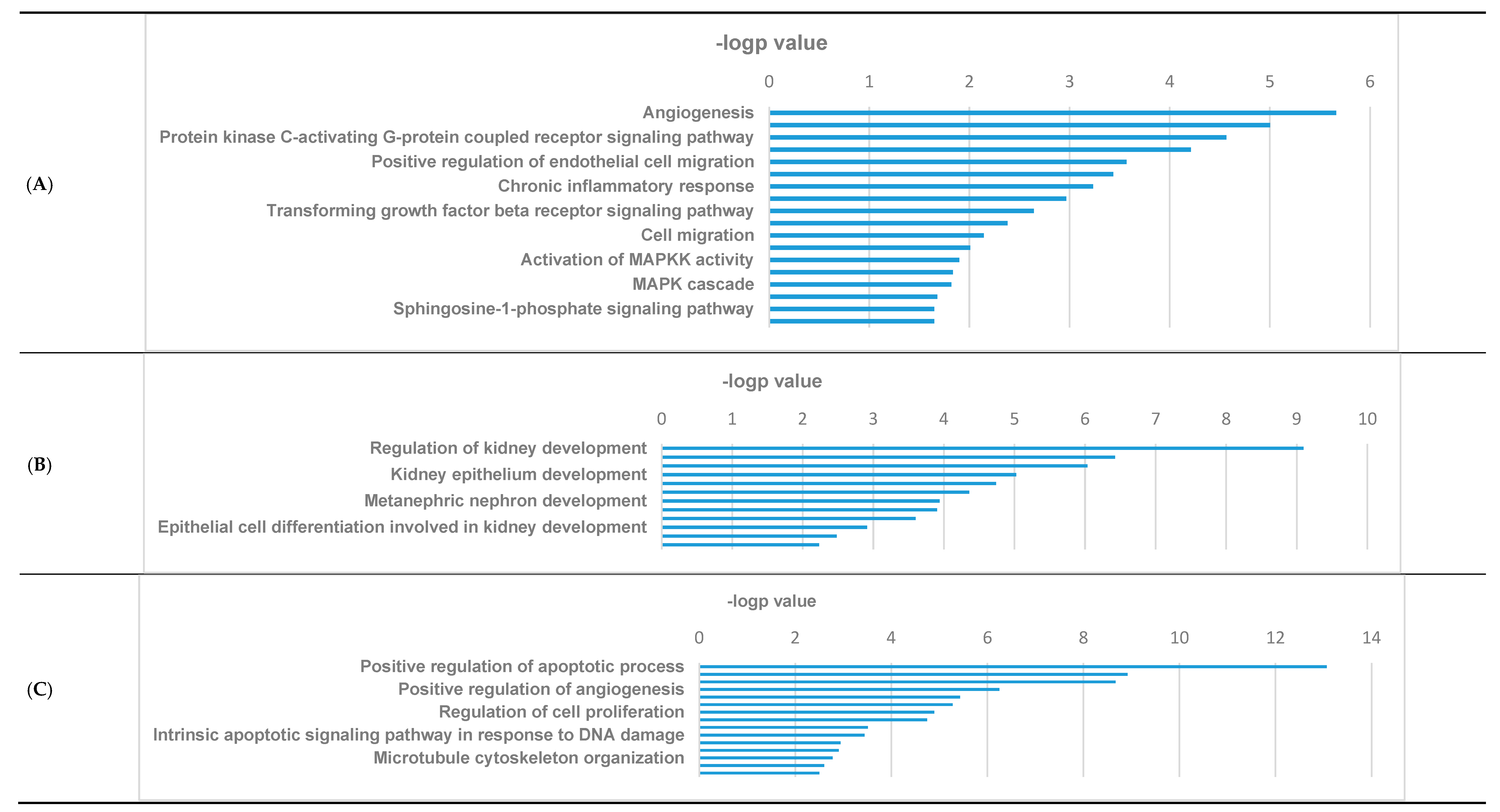
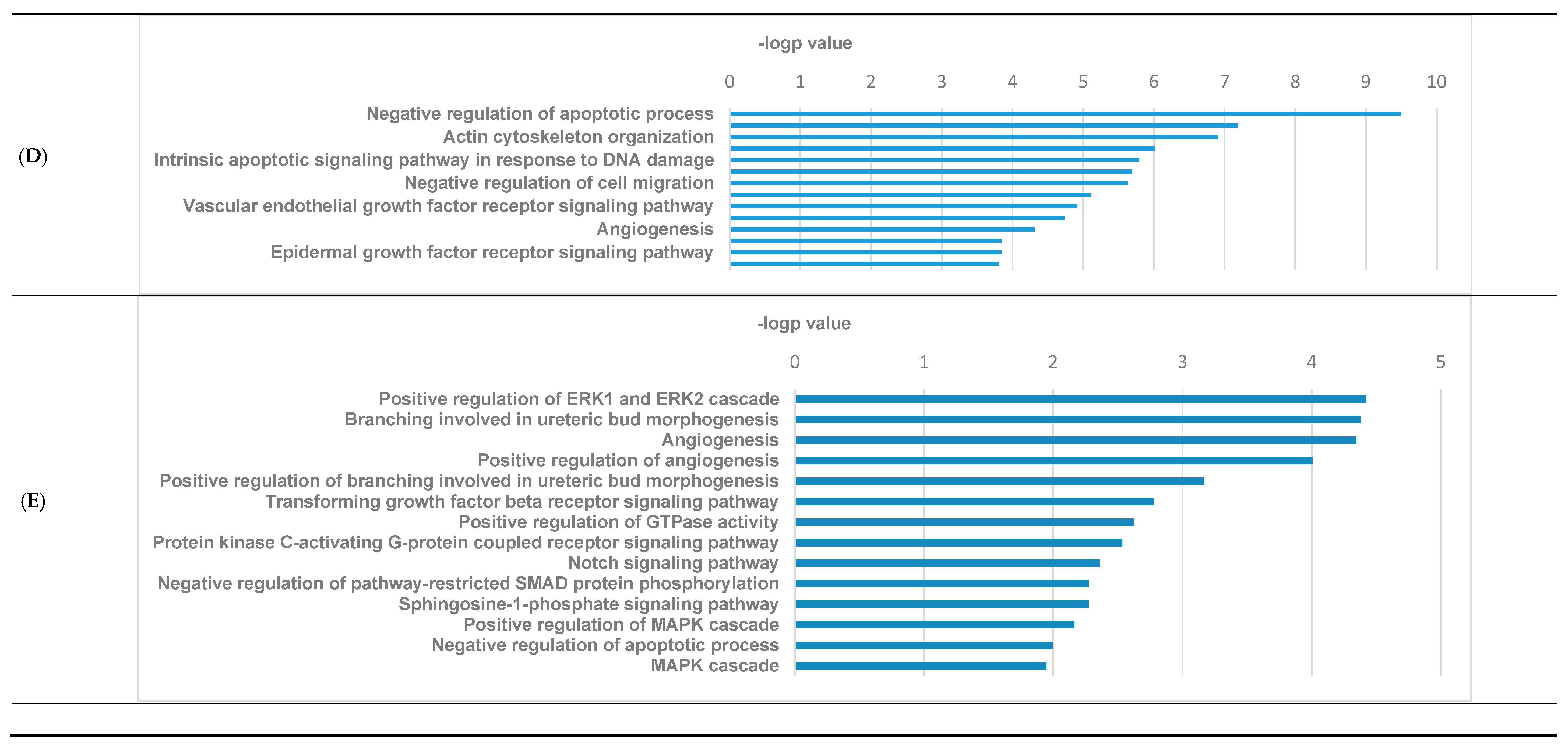
3.3. GO Pathways Related to the DEGs
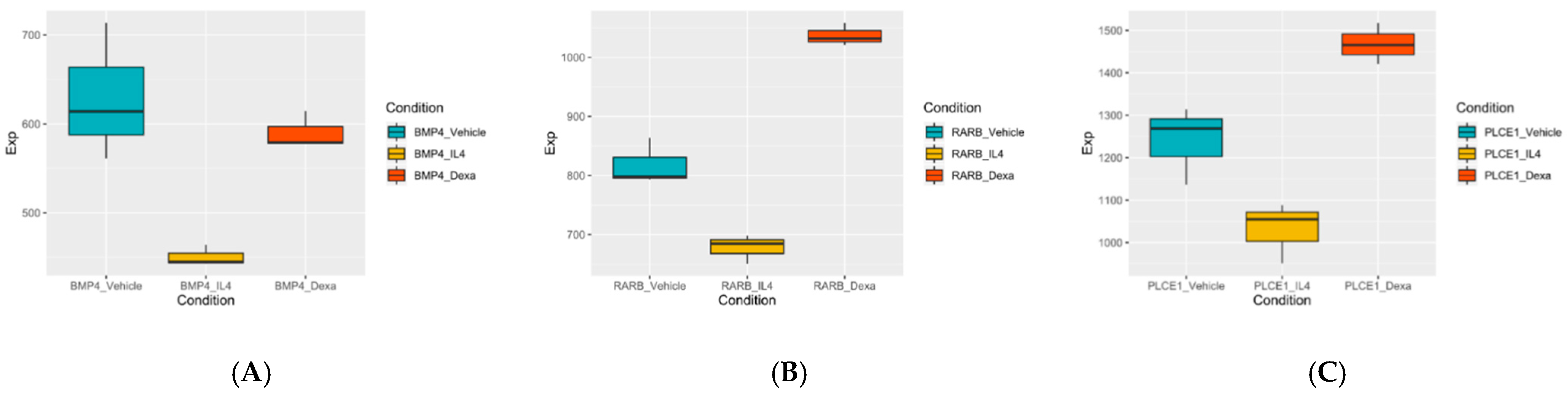
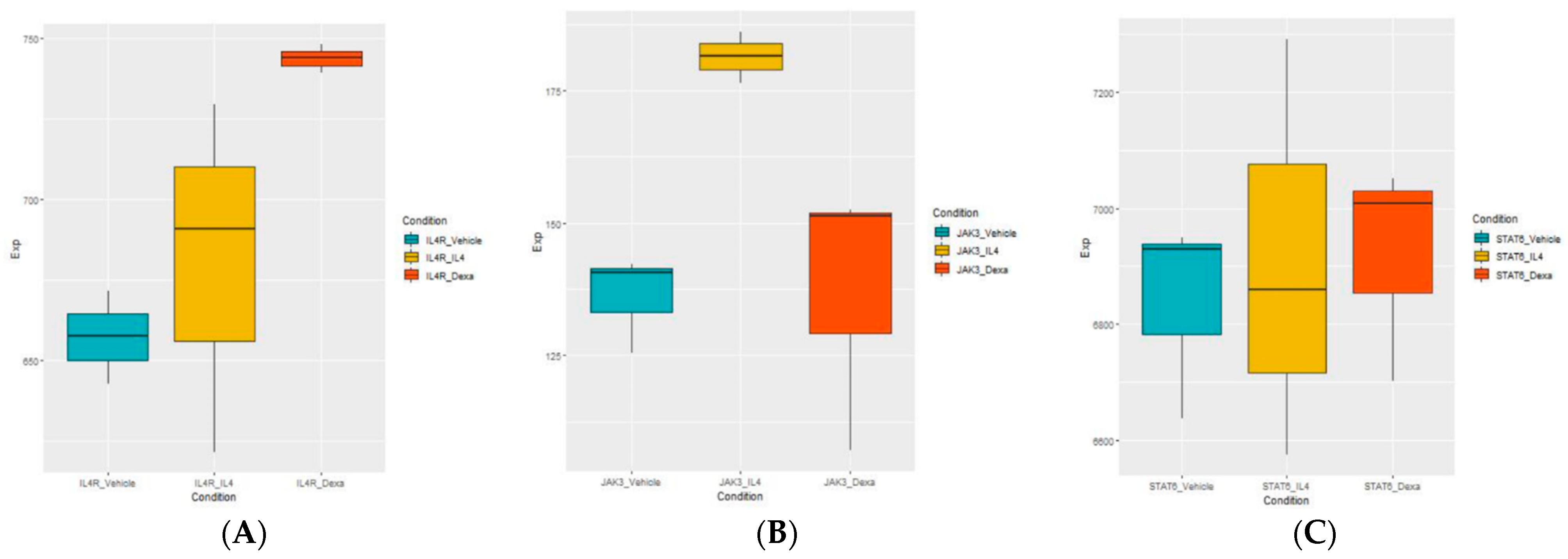
3.4. Selection of the Genes of Potential Relevance to IL-4-Induced Pathogenesis of NS
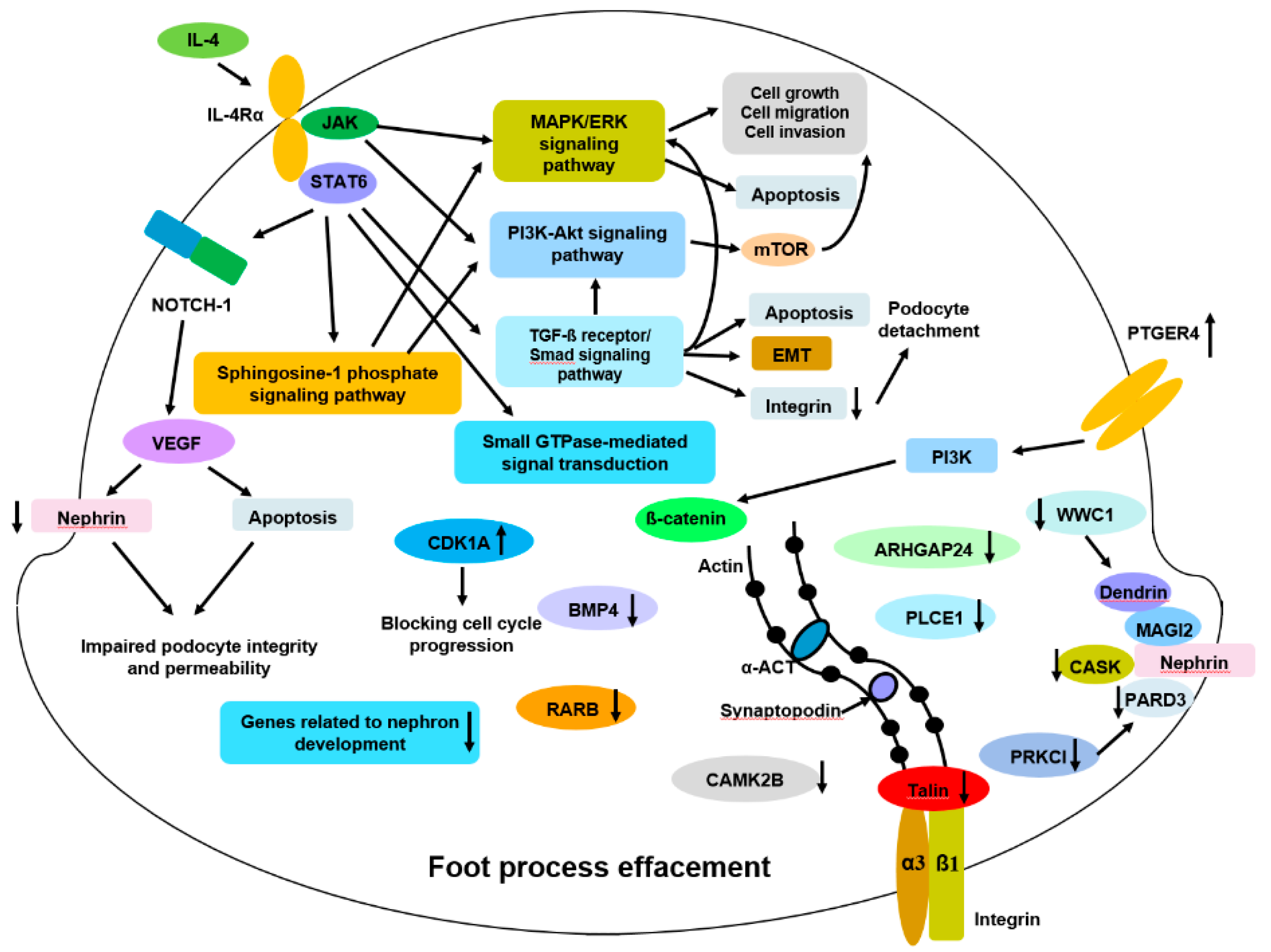
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, S.H.; Park, S.J.; Han, K.H.; Kronbichler, A.; Saleem, M.A.; Oh, J.; Lim, B.J.; Shin, J.I. Pathogenesis of minimal change nephrotic syndrome: An immunological concept. Korean J. Pediatr. 2016, 59, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Eddy, A.A.; Symons, J.M. Nephrotic syndrome in childhood. Lancet 2003, 362, 629–639. [Google Scholar] [CrossRef]
- Greenbaum, L.A.; Benndorf, R.; Smoyer, W.E. Childhood nephrotic syndrome--current and future therapies. Nat. Rev. Nephrol. 2012, 8, 445–458. [Google Scholar] [CrossRef]
- Kang, H.G.; Seo, H.; Lim, J.H.; Kim, J.I.; Han, K.H.; Park, H.W.; Koo, J.W.; Kim, K.H.; Kim, J.H.; Cheong, H.I.; et al. Markers of disease and steroid responsiveness in paediatric idiopathic nephrotic syndrome: Whole-transcriptome sequencing of peripheral blood mononuclear cells. J. Int. Med. Res. 2017, 45, 948–963. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, Q.; Wang, L.; Li, C.; Yang, H.; Wang, X.; Tao, H. The role of Th17/IL-17 in the pathogenesis of primary nephrotic syndrome in children. Kidney Blood Press. Res. 2013, 37, 332–345. [Google Scholar] [CrossRef]
- Liu, L.L.; Qin, Y.; Cai, J.F.; Wang, H.Y.; Tao, J.L.; Li, H.; Chen, L.M.; Li, M.X.; Li, X.M.; Li, X.W. Th17/Treg imbalance in adult patients with minimal change nephrotic syndrome. Clin. Immunol. 2011, 139, 314–320. [Google Scholar] [CrossRef]
- Shimada, M.; Araya, C.; Rivard, C.; Ishimoto, T.; Johnson, R.J.; Garin, E.H. Minimal change disease: A "two-hit" podocyte immune disorder? Pediatr. Nephrol. 2011, 26, 645–649. [Google Scholar] [CrossRef]
- Kanai, T.; Shiraishi, H.; Yamagata, T.; Ito, T.; Odaka, J.; Saito, T.; Aoyagi, J.; Momoi, M.Y. Th2 cells predominate in idiopathic steroid-sensitive nephrotic syndrome. Clin. Exp. Nephrol. 2010, 14, 578–583. [Google Scholar] [CrossRef]
- Zhai, S.; Zhao, L.; Zhang, Y.; Ma, Q. Interleukin-7 stimulation inhibits nephrin activation and induces podocyte injury. Biochem. Biophys. Res. Commun. 2018, 507, 100–105. [Google Scholar] [CrossRef]
- Ha, T.S.; Nam, J.A.; Seong, S.B.; Saleem, M.A.; Park, S.J.; Shin, J.I. Montelukast improves the changes of cytoskeletal and adaptor proteins of human podocytes by interleukin-13. Inflamm Res. 2017, 66, 793–802. [Google Scholar] [CrossRef]
- Zea, A.H.; Stewart, T.; Ascani, J.; Tate, D.J.; Finkel-Jimenez, B.; Wilk, A.; Reiss, K.; Smoyer, W.E.; Aviles, D.H. Activation of the IL-2 Receptor in Podocytes: A Potential Mechanism for Podocyte Injury in Idiopathic Nephrotic Syndrome? PLoS ONE 2016, 11, e0157907. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Saleem, M.A.; Nam, J.A.; Ha, T.S.; Shin, J.I. Effects of interleukin-13 and montelukast on the expression of zonula occludens-1 in human podocytes. Yonsei Med. J. 2015, 56, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Kanai, T.; Shiraishi, H.; Yamagata, T.; Ito, T.; Odaka, J.; Saito, T.; Aoyagi, J.; Momoi, M.Y. Elevated serum interleukin-7 level in idiopathic steroid-sensitive nephrotic syndrome. Pediatr. Int. 2011, 53, 906–909. [Google Scholar] [CrossRef] [PubMed]
- Youssef, D.M.; Elbehidy, R.M.; El-Shal, A.S.; Sherief, L.M. T helper 1 and T helper 2 cytokines in atopic children with steroid-sensitive nephrotic syndrome. Iran. J. Kidney Dis. 2015, 9, 298–305. [Google Scholar]
- Shalaby, S.A.; Al-Edressi, H.M.; El-Tarhouny, S.A.; Fath El-Bab, M.; Zolaly, M.A. Type 1/type 2 cytokine serum levels and role of interleukin-18 in children with steroid-sensitive nephrotic syndrome. Arab J. Nephrol. Transplant. 2013, 6, 83–88. [Google Scholar]
- Cheung, W.; Wei, C.L.; Seah, C.C.; Jordan, S.C.; Yap, H.K. Atopy, serum IgE, and interleukin-13 in steroid-responsive nephrotic syndrome. Pediatr. Nephrol. 2004, 19, 627–632. [Google Scholar] [CrossRef]
- Kim, A.H.; Chung, J.J.; Akilesh, S.; Koziell, A.; Jain, S.; Hodgin, J.B.; Miller, M.J.; Stappenbeck, T.S.; Miner, J.H.; Shaw, A.S. B cell-derived IL-4 acts on podocytes to induce proteinuria and foot process effacement. JCI Insight 2017, 2, e81836. [Google Scholar] [CrossRef]
- Wei, C.; Cheung, W.; Arty, N.; Yap, H.K. Interleukin-4 receptor alpha subunit (IL-4RA) gene polymorphisms and Singapore childhood minimal change disease. Ann. Acad. Med. 2003, 32 (Suppl. 5), S71–S72. [Google Scholar]
- Van Den Berg, J.G.; Aten, J.; Annink, C.; Ravesloot, J.H.; Weber, E.; Weening, J.J. Interleukin-4 and -13 promote basolateral secretion of H(+) and cathepsin L by glomerular epithelial cells. Am. J. Physiol. Ren. Physiol. 2002, 282, F26–F33. [Google Scholar] [CrossRef]
- Van Den Berg, J.G.; Aten, J.; Chand, M.A.; Claessen, N.; Dijkink, L.; Wijdenes, J.; Lakkis, F.G.; Weening, J.J. Interleukin-4 and interleukin-13 act on glomerular visceral epithelial cells. J. Am. Soc. Nephrol. 2000, 11, 413–422. [Google Scholar]
- Kang, J.; Bai, K.M.; Wang, B.L.; Yao, Z.; Pang, X.W.; Chen, W.F. Increased production of interleukin 4 in children with simple idiopathic nephrotic syndrome. Chin. Med. J. 1994, 107, 347–350. [Google Scholar] [PubMed]
- Cho, B.S.; Lee, C.E.; Pyun, K.H. Studies on the role of interleukin-4 and Fc epsilon RII in the pathogenesis of minimal change nephrotic syndrome. J. Korean Med. Sci. 1992, 7, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Yap, H.K.; Cheung, W.; Murugasu, B.; Sim, S.K.; Seah, C.C.; Jordan, S.C. Th1 and Th2 cytokine mRNA profiles in childhood nephrotic syndrome: Evidence for increased IL-13 mRNA expression in relapse. J. Am. Soc. Nephrol. 1999, 10, 529–537. [Google Scholar] [PubMed]
- Lai, K.W.; Wei, C.L.; Tan, L.K.; Tan, P.H.; Chiang, G.S.; Lee, C.G.; Jordan, S.C.; Yap, H.K. Overexpression of interleukin-13 induces minimal-change-like nephropathy in rats. J. Am. Soc. Nephrol. 2007, 18, 1476–1485. [Google Scholar] [CrossRef] [PubMed]
- Prasad, N.; Jaiswal, A.K.; Agarwal, V.; Yadav, B.; Sharma, R.K.; Rai, M.; Singh, H.; Chaturvedi, S.; Singh, A. Differential alteration in peripheral T-regulatory and T-effector cells with change in P-glycoprotein expression in Childhood Nephrotic Syndrome: A longitudinal study. Cytokine 2015, 72, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Youn, Y.S.; Lim, H.H.; Lee, J.H. The clinical characteristics of steroid responsive nephrotic syndrome of children according to the serum immunoglobulin E levels and cytokines. Yonsei Med. J. 2012, 53, 715–722. [Google Scholar] [CrossRef]
- Saleem, M.A.; O’Hare, M.J.; Reiser, J.; Coward, R.J.; Inward, C.D.; Farren, T.; Xing, C.Y.; Ni, L.; Mathieson, P.W.; Mundel, P. A conditionally immortalized human podocyte cell line demonstrating nephrin and podocin expression. J. Am. Soc. Nephrol. 2002, 13, 630–638. [Google Scholar]
- Xing, C.Y.; Saleem, M.A.; Coward, R.J.; Ni, L.; Witherden, I.R.; Mathieson, P.W. Direct effects of dexamethasone on human podocytes. Kidney Int. 2006, 70, 1038–1045. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Ghosh, S.; Chan, C.K. Analysis of RNA-Seq Data Using TopHat and Cufflinks. Methods Mol. Biol. 2016, 1374, 339–361. [Google Scholar] [CrossRef]
- Warsow, G.; Endlich, N.; Schordan, E.; Schordan, S.; Chilukoti, R.K.; Homuth, G.; Moeller, M.J.; Fuellen, G.; Endlich, K. PodNet, a protein-protein interaction network of the podocyte. Kidney Int. 2013, 84, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ding, J.; Fan, Q.; Zhang, H. The activation of extracellular signal-regulated kinase is responsible for podocyte injury. Mol. Biol. Rep. 2010, 37, 2477–2484. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.Q.; Shi, W.; Liu, S.X.; Zhang, B.; Xu, L.X.; Liang, X.L.; Liang, Y.Z. Podocyte injury is suppressed by 1,25-dihydroxyvitamin D via modulation of transforming growth factor-beta 1/bone morphogenetic protein-7 signalling in puromycin aminonucleoside nephropathy rats. Clin. Exp. Pharm. Physiol. 2009, 36, 682–689. [Google Scholar] [CrossRef]
- Niranjan, T.; Murea, M.; Susztak, K. The pathogenic role of Notch activation in podocytes. Nephron Exp. Nephrol. 2009, 111, e73–e79. [Google Scholar] [CrossRef]
- Sparks, M.A.; Coffman, T.M. The EP4 receptor for prostaglandin E2 in glomerular disease: A good receptor turned bad? J. Am. Soc. Nephrol. 2010, 21, 1597–1599. [Google Scholar] [CrossRef] [PubMed]
- Faour, W.H.; Gomi, K.; Kennedy, C.R. PGE(2) induces COX-2 expression in podocytes via the EP(4) receptor through a PKA-independent mechanism. Cell. Signal. 2008, 20, 2156–2164. [Google Scholar] [CrossRef]
- Stitt-Cavanagh, E.M.; Faour, W.H.; Takami, K.; Carter, A.; Vanderhyden, B.; Guan, Y.; Schneider, A.; Breyer, M.D.; Kennedy, C.R. A maladaptive role for EP4 receptors in podocytes. J. Am. Soc. Nephrol. 2010, 21, 1678–1690. [Google Scholar] [CrossRef]
- Thieme, K.; Majumder, S.; Brijmohan, A.S.; Batchu, S.N.; Bowskill, B.B.; Alghamdi, T.A.; Advani, S.L.; Kabir, M.G.; Liu, Y.; Advani, A. EP4 inhibition attenuates the development of diabetic and non-diabetic experimental kidney disease. Sci. Rep. 2017, 7, 3442. [Google Scholar] [CrossRef]
- Aringer, I.; Artinger, K.; Kirsch, A.H.; Schabhuttl, C.; Jandl, K.; Barnthaler, T.; Mooslechner, A.A.; Herzog, S.A.; Uhlig, M.; Kirsch, A.; et al. Blockade of prostaglandin E2 receptor 4 ameliorates nephrotoxic serum nephritis. Am. J. Physiol. Ren. Physiol. 2018, 315, F1869–F1880. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Pippin, J.W.; Hagmann, H.; Krofft, R.D.; Chang, A.M.; Zhang, J.; Terada, Y.; Brinkkoetter, P.; Shankland, S.J. Both cyclin I and p35 are required for maximal survival benefit of cyclin-dependent kinase 5 in kidney podocytes. Am. J. Physiol. Ren. Physiol. 2012, 302, F1161–F1171. [Google Scholar] [CrossRef][Green Version]
- Griffin, S.V.; Olivier, J.P.; Pippin, J.W.; Roberts, J.M.; Shankland, S.J. Cyclin I protects podocytes from apoptosis. J. Biol. Chem. 2006, 281, 28048–28057. [Google Scholar] [CrossRef] [PubMed]
- Koehler, S.; Tellkamp, F.; Niessen, C.M.; Bloch, W.; Kerjaschki, D.; Schermer, B.; Benzing, T.; Brinkkoetter, P.T. Par3A is dispensable for the function of the glomerular filtration barrier of the kidney. Am. J. Physiol. Ren. Physiol. 2016, 311, F112–F119. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Gu, Y.; Coates, G.; Groome, L.J.; Saleem, M.A.; Mathieson, P.W.; Wang, Y. Altered nephrin and podoplanin distribution is associated with disturbed polarity protein PARD-3 and PARD-6 expressions in podocytes from preeclampsia. Reprod. Sci. 2011, 18, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Chung, C.; Kim, J.M.; Lee, D.; Cho, S.Y.; Lee, T.H.; Cho, H.J.; Yeo, M.K. Clinical significance of atypical protein kinase C (PKCiota and PKCzeta) and its relationship with yes-associated protein in lung adenocarcinoma. BMC Cancer 2019, 19, 804. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Hartleben, B.; Huber, T.B. Podocyte polarity signalling. Curr. Opin. Nephrol. Hypertens. 2009, 18, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Huber, T.B.; Hartleben, B.; Winkelmann, K.; Schneider, L.; Becker, J.U.; Leitges, M.; Walz, G.; Haller, H.; Schiffer, M. Loss of podocyte aPKClambda/iota causes polarity defects and nephrotic syndrome. J. Am. Soc. Nephrol. 2009, 20, 798–806. [Google Scholar] [CrossRef]
- Song, L.; Tang, S.; Han, X.; Jiang, Z.; Dong, L.; Liu, C.; Liang, X.; Dong, J.; Qiu, C.; Wang, Y.; et al. KIBRA controls exosome secretion via inhibiting the proteasomal degradation of Rab27a. Nat. Commun. 2019, 10, 1639. [Google Scholar] [CrossRef]
- Lin, T.; Zhang, L.; Liu, S.; Chen, Y.; Zhang, H.; Zhao, X.; Li, R.; Zhang, Q.; Liao, R.; Huang, Z.; et al. WWC1 promotes podocyte survival via stabilizing slit diaphragm protein dendrin. Mol. Med. Rep. 2017, 16, 8685–8690. [Google Scholar] [CrossRef][Green Version]
- Duning, K.; Schurek, E.M.; Schluter, M.; Bayer, M.; Reinhardt, H.C.; Schwab, A.; Schaefer, L.; Benzing, T.; Schermer, B.; Saleem, M.A.; et al. KIBRA modulates directional migration of podocytes. J. Am. Soc. Nephrol. 2008, 19, 1891–1903. [Google Scholar] [CrossRef]
- Meliambro, K.; Wong, J.S.; Ray, J.; Calizo, R.C.; Towne, S.; Cole, B.; El Salem, F.; Gordon, R.E.; Kaufman, L.; He, J.C.; et al. The Hippo pathway regulator KIBRA promotes podocyte injury by inhibiting YAP signaling and disrupting actin cytoskeletal dynamics. J. Biol. Chem. 2017, 292, 21137–21148. [Google Scholar] [CrossRef]
- Kwok, E.; Rodriguez, D.J.; Kremerskothen, J.; Nyarko, A. Intrinsic disorder and amino acid specificity modulate binding of the WW2 domain in kidney and brain protein (KIBRA) to synaptopodin. J. Biol. Chem. 2019, 294, 17383–17394. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, S.; Lehtonen, E.; Kudlicka, K.; Holthofer, H.; Farquhar, M.G. Nephrin forms a complex with adherens junction proteins and CASK in podocytes and in Madin-Darby canine kidney cells expressing nephrin. Am. J. Pathol. 2004, 165, 923–936. [Google Scholar] [CrossRef]
- Beaudreuil, S.; Zhang, X.; Herr, F.; Harper, F.; Candelier, J.J.; Fan, Y.; Yeter, H.; Dudreuilh, C.; Lecru, L.; Vazquez, A.; et al. Circulating CASK is associated with recurrent focal segmental glomerulosclerosis after transplantation. PLoS ONE 2019, 14, e0219353. [Google Scholar] [CrossRef] [PubMed]
- Gough, R.E.; Goult, B.T. The tale of two talins-two isoforms to fine-tune integrin signalling. FEBS Lett. 2018, 592, 2108–2125. [Google Scholar] [CrossRef]
- Mathew, S.; Palamuttam, R.J.; Mernaugh, G.; Ramalingam, H.; Lu, Z.; Zhang, M.Z.; Ishibe, S.; Critchley, D.R.; Fassler, R.; Pozzi, A.; et al. Talin regulates integrin beta1-dependent and -independent cell functions in ureteric bud development. Development 2017, 144, 4148–4158. [Google Scholar] [CrossRef]
- Akilesh, S.; Suleiman, H.; Yu, H.; Stander, M.C.; Lavin, P.; Gbadegesin, R.; Antignac, C.; Pollak, M.; Kopp, J.B.; Winn, M.P.; et al. Arhgap24 inactivates Rac1 in mouse podocytes, and a mutant form is associated with familial focal segmental glomerulosclerosis. J. Clin. Investig. 2011, 121, 4127–4137. [Google Scholar] [CrossRef]
- Maeda, K.; Otomo, K.; Yoshida, N.; Abu-Asab, M.S.; Ichinose, K.; Nishino, T.; Kono, M.; Ferretti, A.; Bhargava, R.; Maruyama, S.; et al. CaMK4 compromises podocyte function in autoimmune and nonautoimmune kidney disease. J. Clin. Investig. 2018, 128, 3445–3459. [Google Scholar] [CrossRef]
- Jiang, L.; Xu, L.; Song, Y.; Li, J.; Mao, J.; Zhao, A.Z.; He, W.; Yang, J.; Dai, C. Calmodulin-dependent protein kinase II/cAMP response element-binding protein/Wnt/beta-catenin signaling cascade regulates angiotensin II-induced podocyte injury and albuminuria. J. Biol. Chem. 2013, 288, 23368–23379. [Google Scholar] [CrossRef]
- Hinkes, B.; Wiggins, R.C.; Gbadegesin, R.; Vlangos, C.N.; Seelow, D.; Nurnberg, G.; Garg, P.; Verma, R.; Chaib, H.; Hoskins, B.E.; et al. Positional cloning uncovers mutations in PLCE1 responsible for a nephrotic syndrome variant that may be reversible. Nat. Genet. 2006, 38, 1397–1405. [Google Scholar] [CrossRef]
- Gbadegesin, R.; Hinkes, B.G.; Hoskins, B.E.; Vlangos, C.N.; Heeringa, S.F.; Liu, J.; Loirat, C.; Ozaltin, F.; Hashmi, S.; Ulmer, F.; et al. Mutations in PLCE1 are a major cause of isolated diffuse mesangial sclerosis (IDMS). Nephrol. Dial. Transplant. 2008, 23, 1291–1297. [Google Scholar] [CrossRef]
- Khurana, S.; Chakraborty, S.; Lam, M.; Liu, Y.; Su, Y.T.; Zhao, X.; Saleem, M.A.; Mathieson, P.W.; Bruggeman, L.A.; Kao, H.Y. Familial focal segmental glomerulosclerosis (FSGS)-linked alpha-actinin 4 (ACTN4) protein mutants lose ability to activate transcription by nuclear hormone receptors. J. Biol. Chem. 2012, 287, 12027–12035. [Google Scholar] [CrossRef] [PubMed]
- Trasino, S.E.; Tang, X.H.; Shevchuk, M.M.; Choi, M.E.; Gudas, L.J. Amelioration of Diabetic Nephropathy Using a Retinoic Acid Receptor beta2 Agonist. J. Pharm. Exp. Ther. 2018, 367, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Miyazaki, Y.; Matsusaka, T.; Utsunomiya, Y.; Kawamura, T.; Hosoya, T.; Ichikawa, I. Bmp in podocytes is essential for normal glomerular capillary formation. J. Am. Soc. Nephrol. 2008, 19, 685–694. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Ontology (GO) Pathway | Genes |
|---|---|
| Positive Regulation of ERK1 and ERK2 cascade | FGFR2, FGF2, PDGFB, ICAM1, GCNT2, CCL2, CD44, SEMA7A, PDGFRB, CX3CL1 |
| Positive Regulation of MAPK cascade | FGFR2, PDGFB, C1QTNF1, WWC1, NGFR |
| Negative Regulation of apoptotic process | PDGFRB, PAK2, CD44, SPHK1, PRKCI, MALT1, NGFR, RARB, GREM1, APBB2, SOX9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.M.; Ko, Y.; Lee, C.H.; Jeon, N.; Lee, K.H.; Oh, J.; Kronbichler, A.; Saleem, M.A.; Lim, B.J.; Shin, J.I. The Effect of Interleukin-4 and Dexamethasone on RNA-Seq-Based Transcriptomic Profiling of Human Podocytes: A Potential Role in Minimal Change Nephrotic Syndrome. J. Clin. Med. 2021, 10, 496. https://doi.org/10.3390/jcm10030496
Lee JM, Ko Y, Lee CH, Jeon N, Lee KH, Oh J, Kronbichler A, Saleem MA, Lim BJ, Shin JI. The Effect of Interleukin-4 and Dexamethasone on RNA-Seq-Based Transcriptomic Profiling of Human Podocytes: A Potential Role in Minimal Change Nephrotic Syndrome. Journal of Clinical Medicine. 2021; 10(3):496. https://doi.org/10.3390/jcm10030496
Chicago/Turabian StyleLee, Jiwon M., Younhee Ko, Chul Ho Lee, Nara Jeon, Keum Hwa Lee, Jun Oh, Andreas Kronbichler, Moin A. Saleem, Beom Jin Lim, and Jae Il Shin. 2021. "The Effect of Interleukin-4 and Dexamethasone on RNA-Seq-Based Transcriptomic Profiling of Human Podocytes: A Potential Role in Minimal Change Nephrotic Syndrome" Journal of Clinical Medicine 10, no. 3: 496. https://doi.org/10.3390/jcm10030496
APA StyleLee, J. M., Ko, Y., Lee, C. H., Jeon, N., Lee, K. H., Oh, J., Kronbichler, A., Saleem, M. A., Lim, B. J., & Shin, J. I. (2021). The Effect of Interleukin-4 and Dexamethasone on RNA-Seq-Based Transcriptomic Profiling of Human Podocytes: A Potential Role in Minimal Change Nephrotic Syndrome. Journal of Clinical Medicine, 10(3), 496. https://doi.org/10.3390/jcm10030496